
BAF-L Modulates Histone-to-Protamine Transition during Spermiogenesis

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
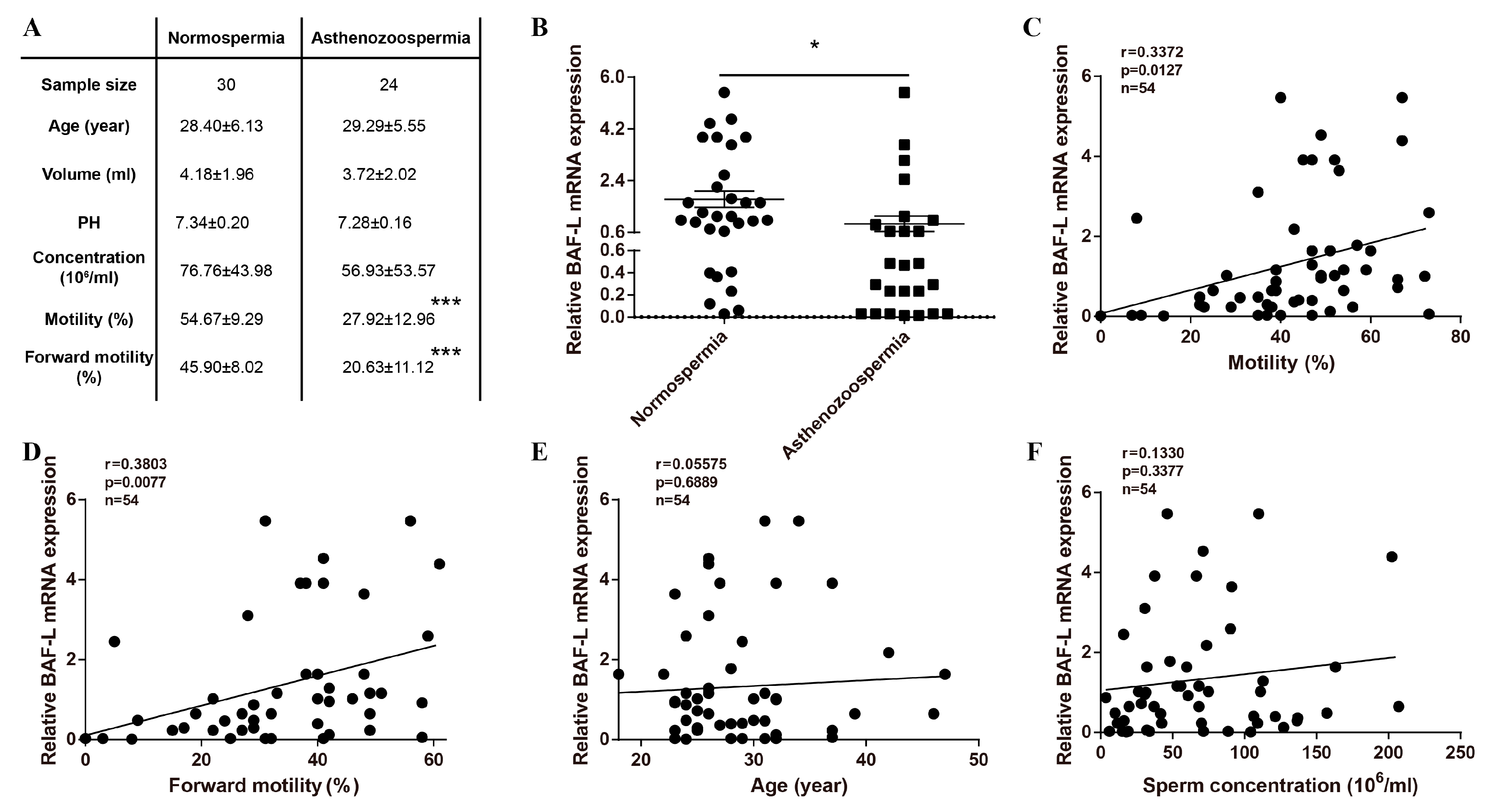
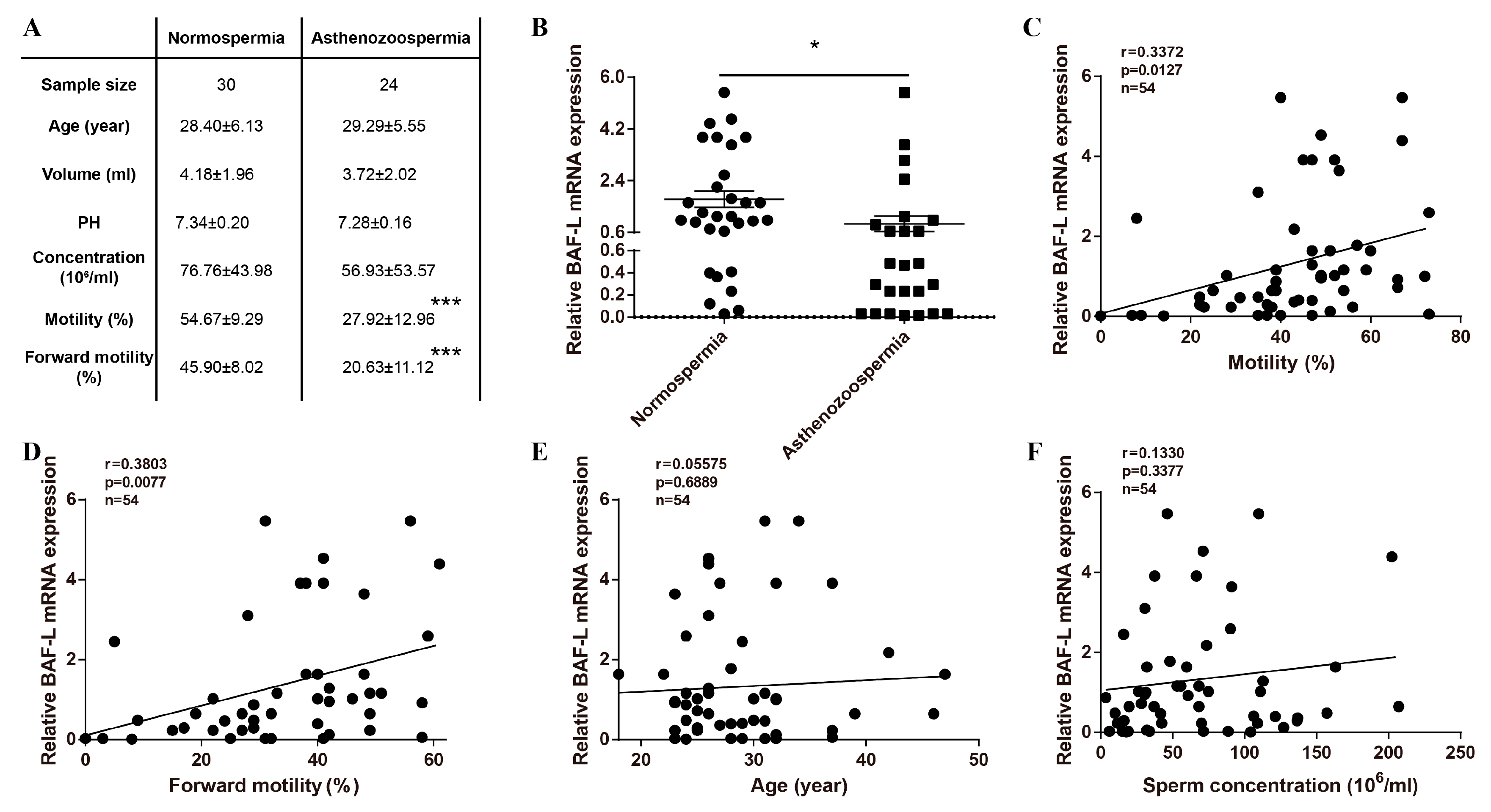
2.1. Reduced Expression of BAF-L Is Related to Human Asthenozoospermia
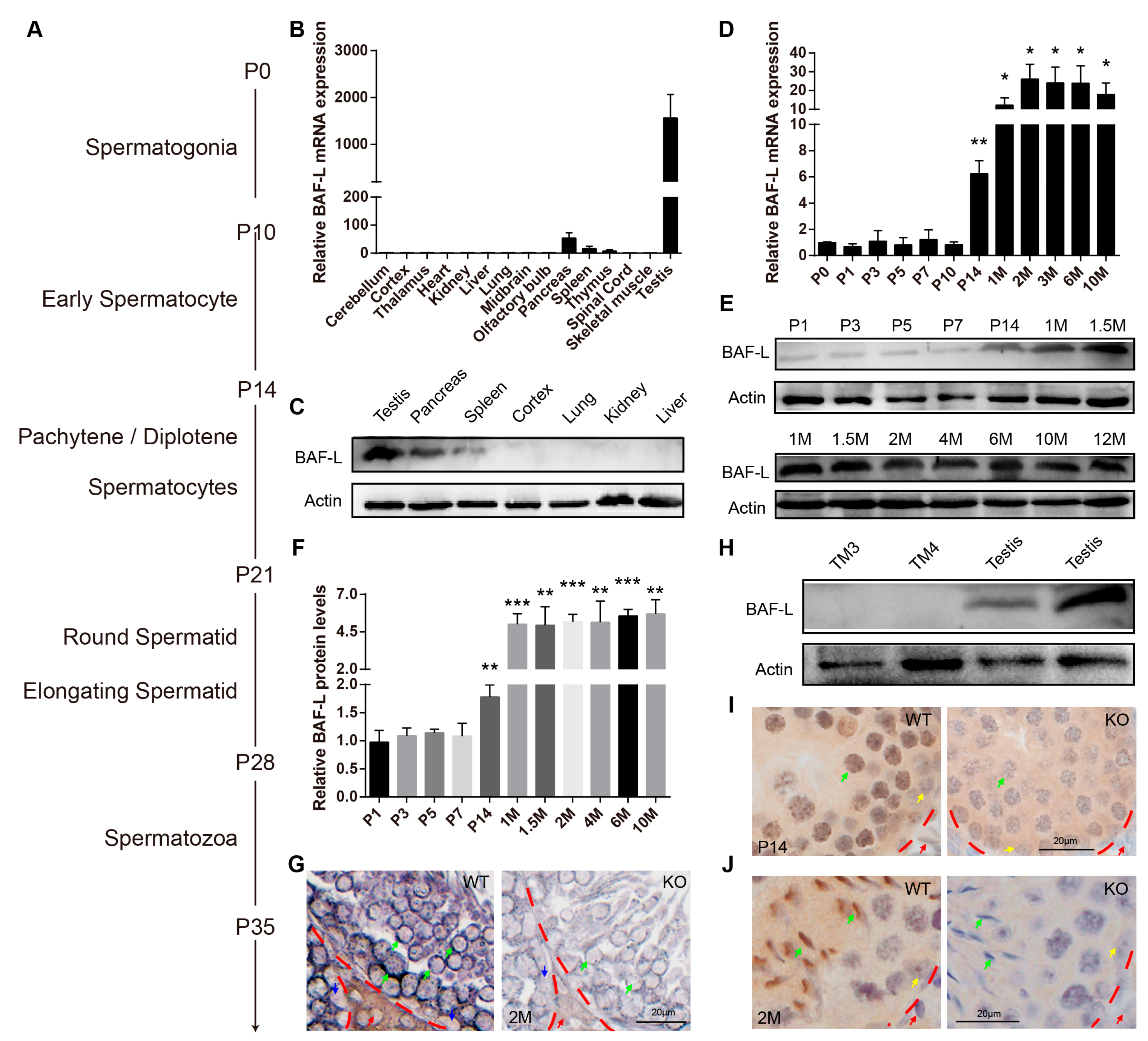
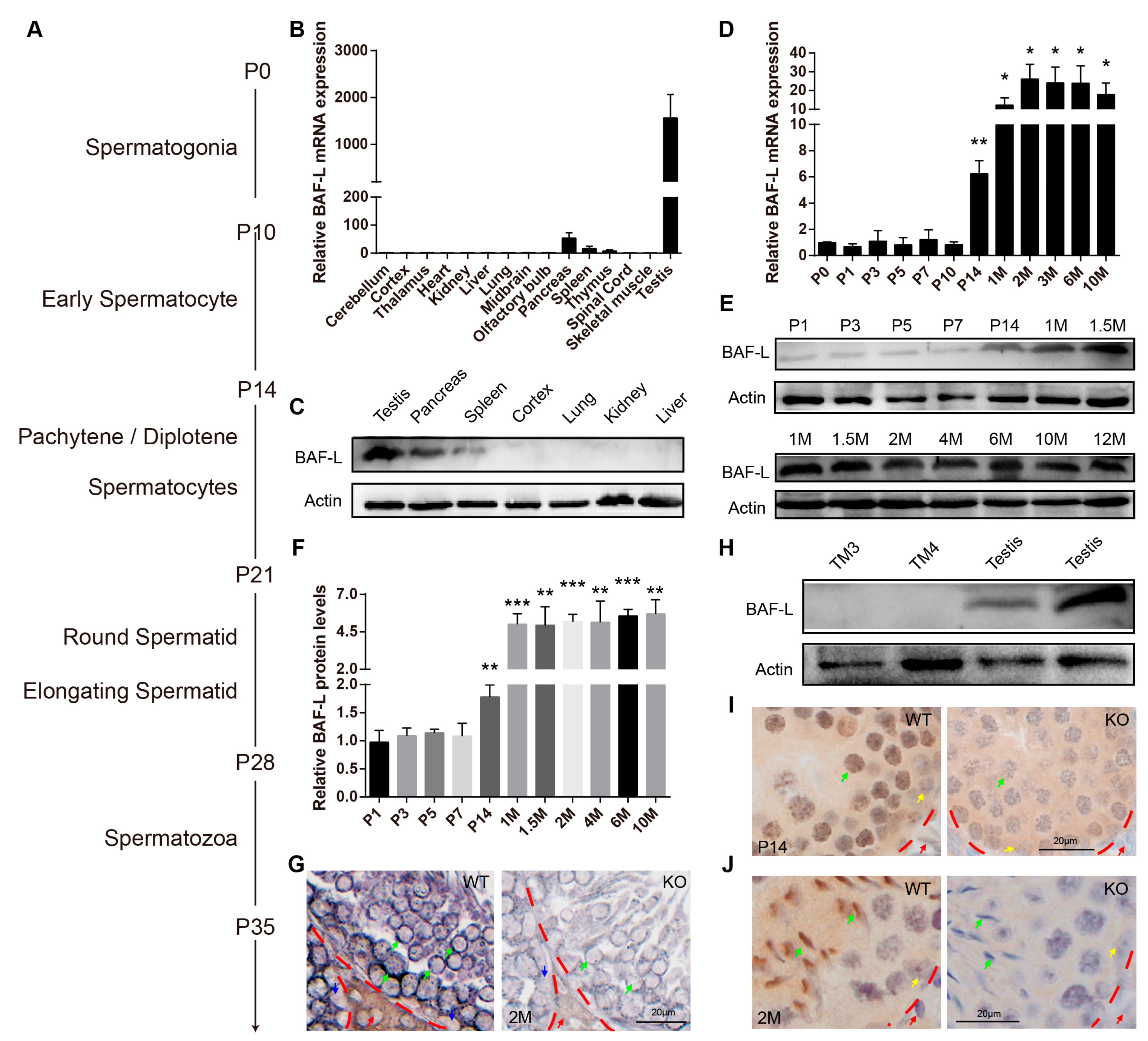
2.2. BAF-L Is a Male Germ Cell Marker That Corelates with Spermatozoal Maturation
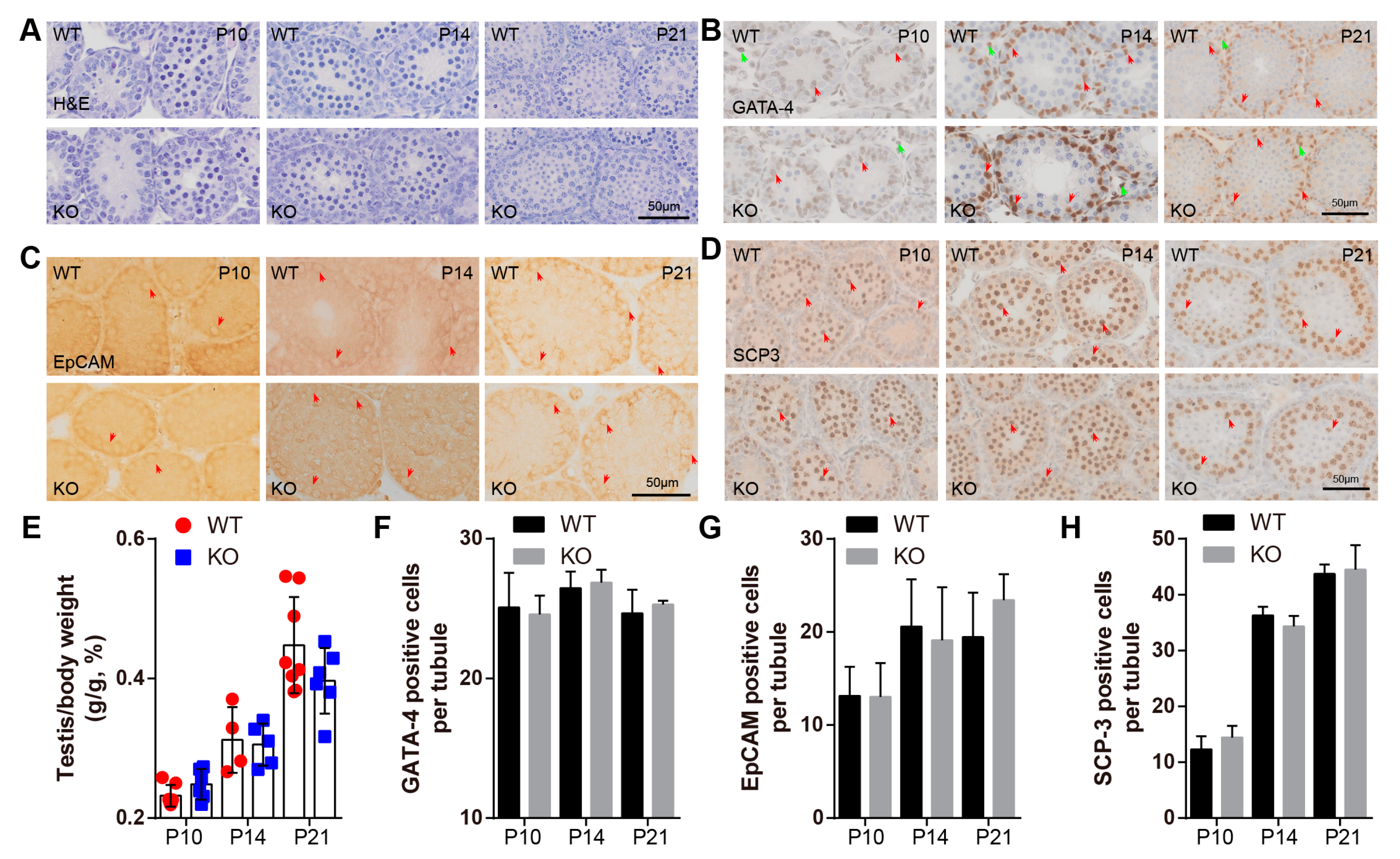
2.3. BAF-L Does Not Affect the Early Progress of Spermatogenesis
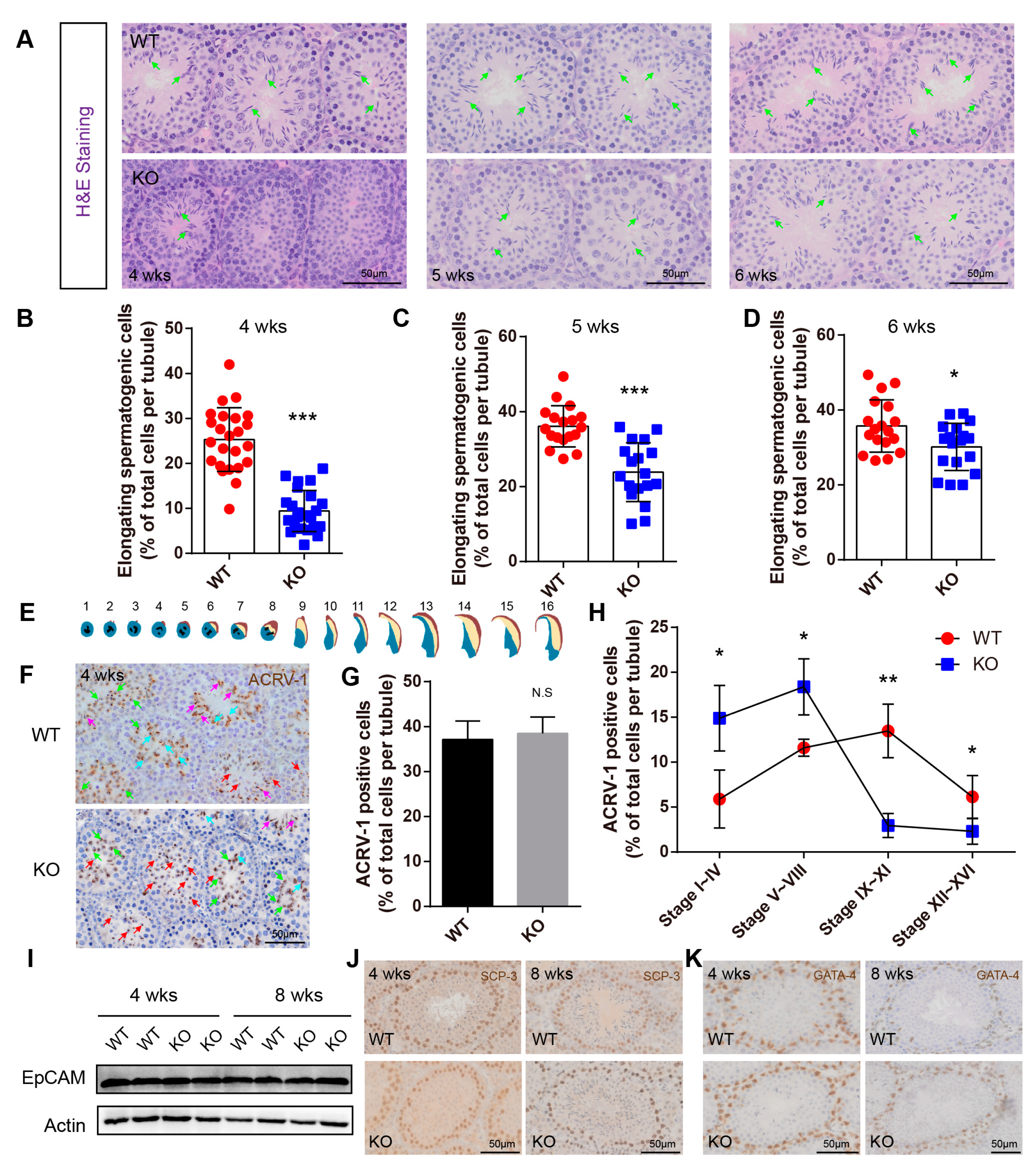
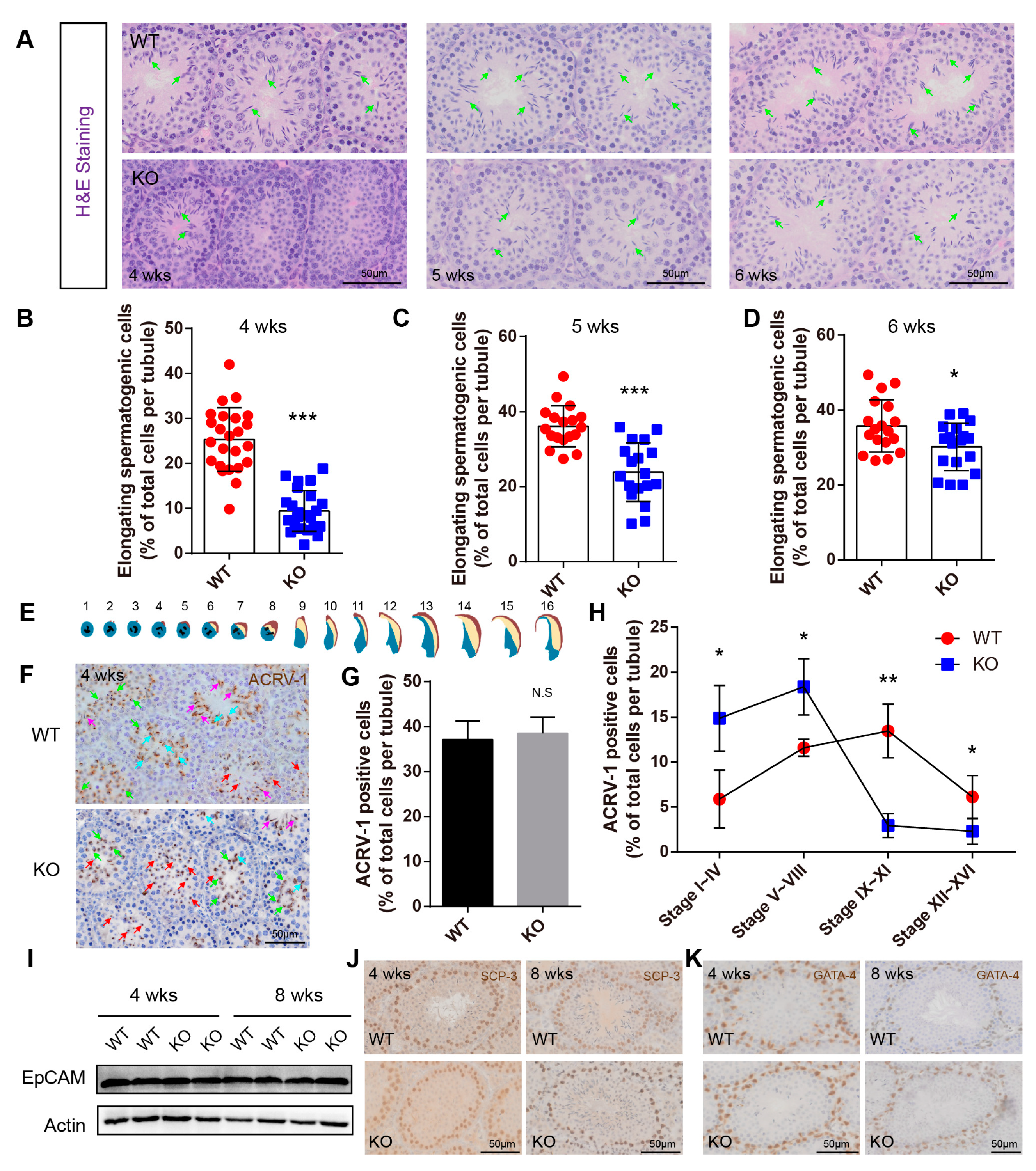
2.4. BAF-L Deficiency Affects the Progress of Spermiogenesis
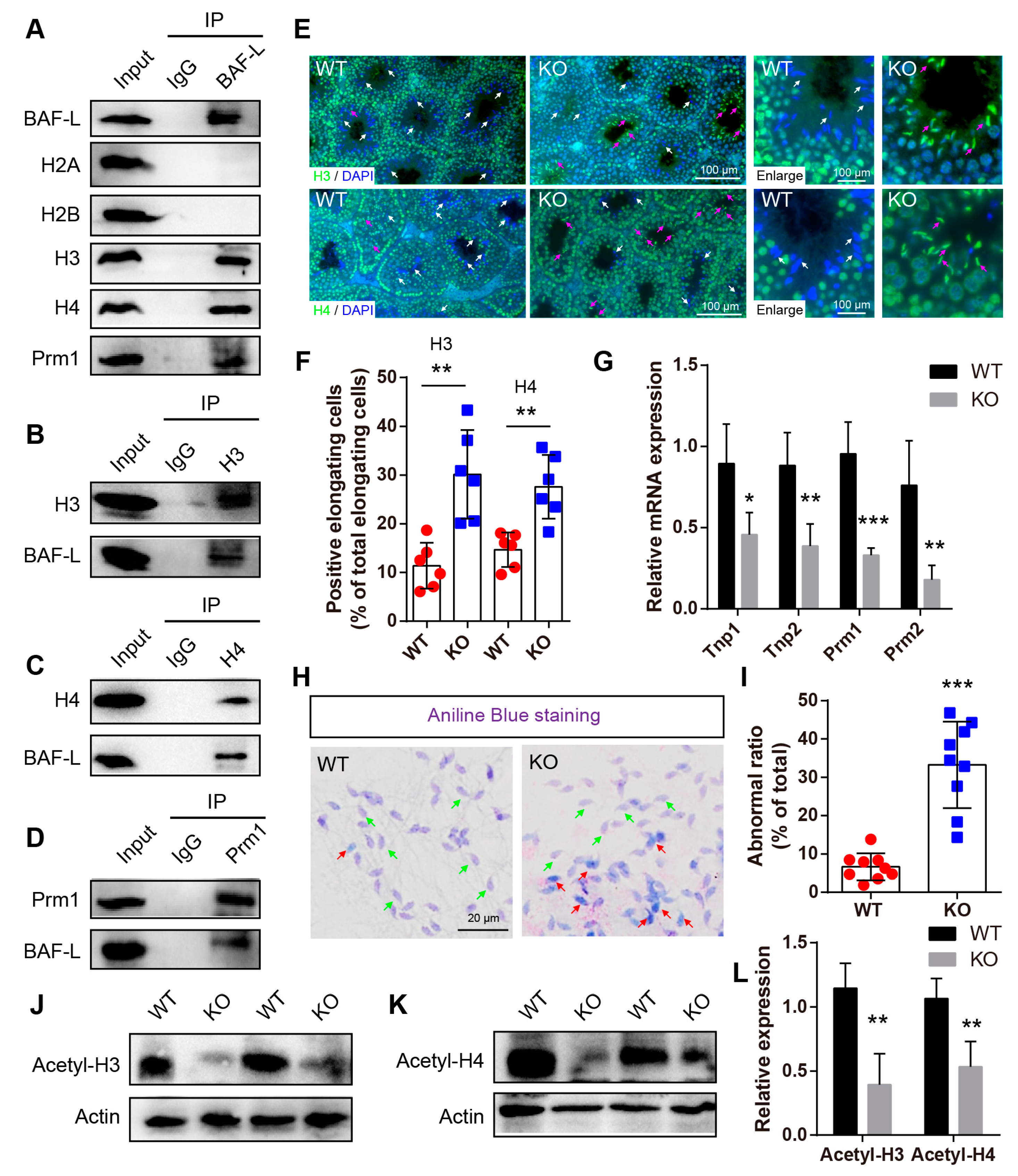
2.5. BAF-L Regulates Histone–Protamine Exchange in Spermiogenesis
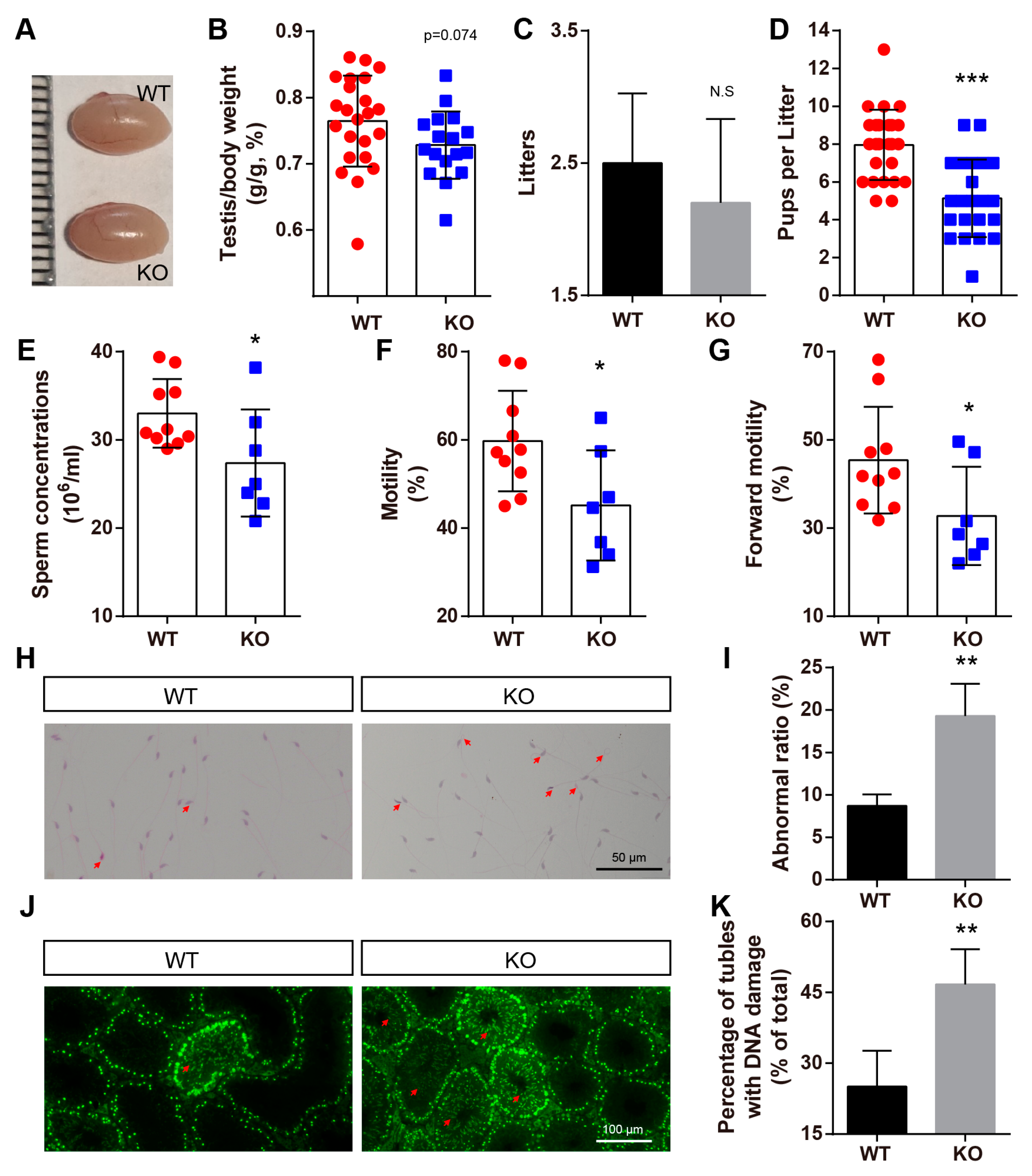
2.6. BAF-L KO Results in Male Subfertility
3. Discussion
4. Materials and Methods
4.1. Patients, Semen Collection, and Analysis
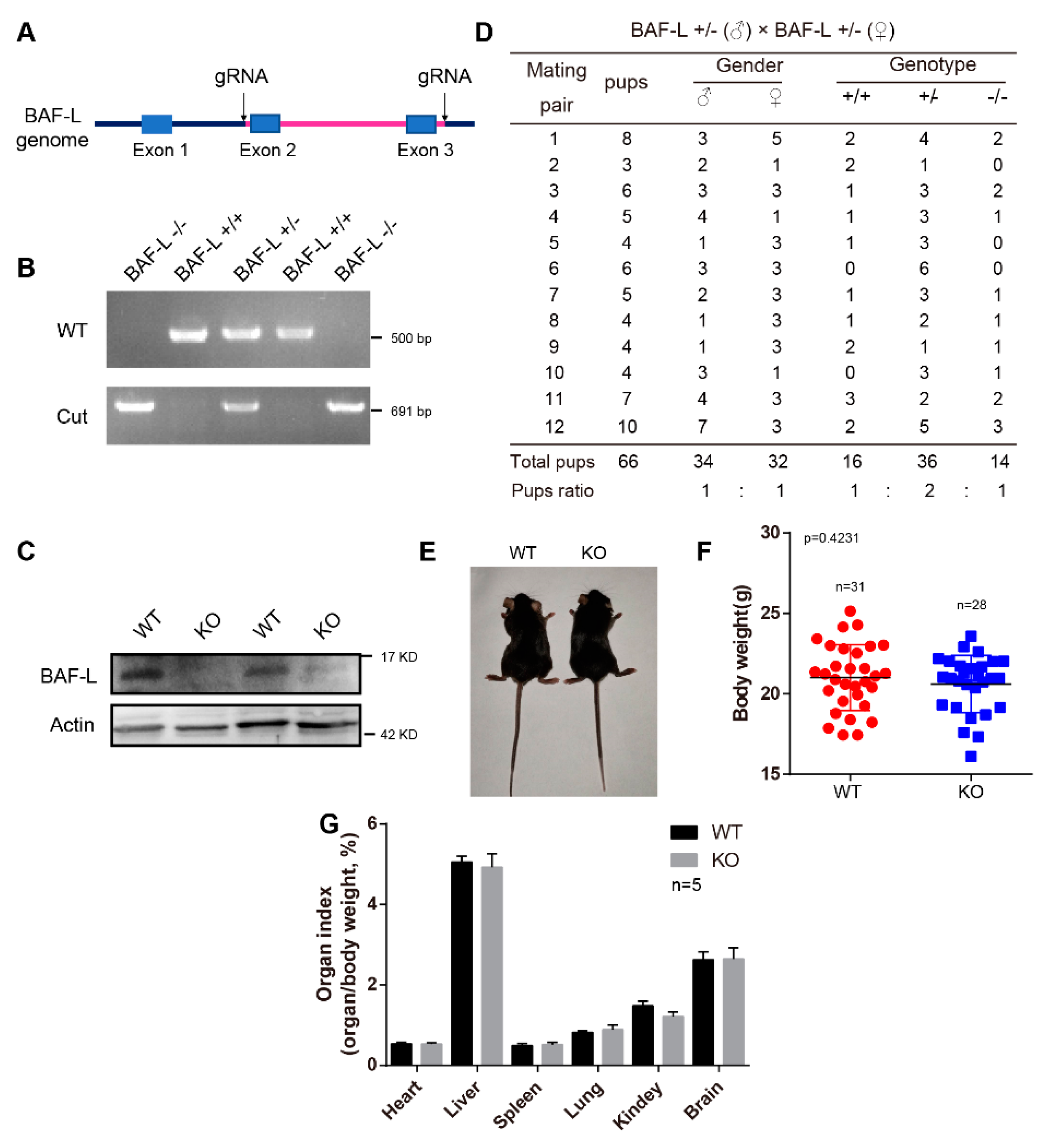
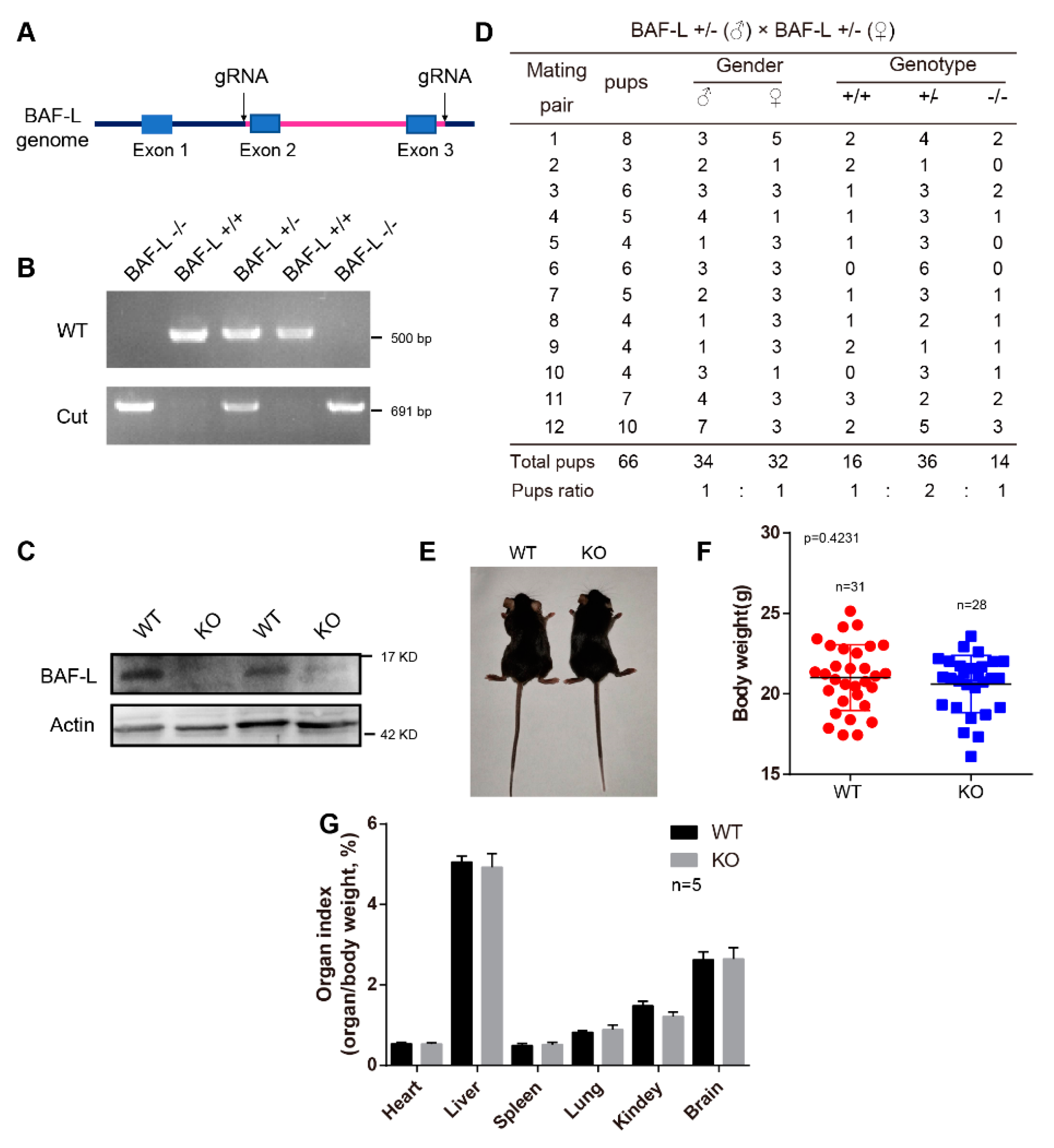
4.2. Animals
4.3. In Situ Hybridization
4.4. Quantitative Realtime PCR
4.5. Western Blotting
4.6. Immunochemical Staining
4.7. Immunofluorescence Staining
4.8. H&E Staining
4.9. Sperm Parameters Analysis
4.10. Cell Culture
4.11. Immunoprecipitation
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clermont, Y. The cycle of the seminiferous epithelium in man. Am. J. Anat. 1963, 112, 35–51. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.L.F.; Varghese, A.C.; Agarwal, A. The genetic causes of male factor infertility: A review. Fertil. Steril. 2010, 93, 1–12. [Google Scholar]
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis. Curr. Biol. CB 2017, 27, R988–R994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, L. Mechanisms of spermiogenesis and spermiation and how they are disturbed. Spermatogenesis 2014, 4, e979623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathke, C.; Baarends, W.M.; Awe, S.; Renkawitz-Pohl, R. Chromatin dynamics during spermiogenesis. Biochim. Biophys. Acta 2014, 1839, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Govin, J.; Caron, C.; Lestrat, C.; Rousseaux, S.; Khochbin, S. The role of histones in chromatin remodelling during mammalian spermiogenesis. Eur. J. Biochem. 2004, 271, 3459–3469. [Google Scholar] [CrossRef]
- Bao, J.; Bedford, M.T. Epigenetic regulation of the histone-to-protamine transition during spermiogenesis. Reproduction 2016, 151, R55. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Brachner, A.; Gotzmann, J.; Foisner, R.; Gruenbaum, Y. Barrier-to-autointegration factor—A BAFfling little protein. Trends Cell Biol. 2007, 17, 202–208. [Google Scholar] [CrossRef]
- Halfmann, C.T.; Sears, R.M.; Katiyar, A.; Busselman, B.W.; Aman, L.K.; Zhang, Q.; O’Bryan, C.S.; Angelini, T.E.; Lele, T.P.; Roux, K.J. Repair of nuclear ruptures requires barrier-to-autointegration factor. J. Cell Biol. 2019, 218, 2136–2149. [Google Scholar] [CrossRef] [Green Version]
- Jamin, A.; Wiebe, M.S. Barrier to Autointegration Factor (BANF1): Interwoven roles in nuclear structure, genome integrity, innate immunity, stress responses and progeria. Curr. Opin. Cell Biol. 2015, 34, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Bolderson, E.; Burgess, J.T.; Li, J.; Gandhi, N.S.; Boucher, D.; Croft, L.V.; Beard, S.; Plowman, J.J.; Suraweera, A.; Adams, M.N.; et al. Barrier-to-autointegration factor 1 (Banf1) regulates poly [ADP-ribose] polymerase 1 (PARP1) activity following oxidative DNA damage. Nat. Commun. 2019, 10, 5501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura-Totten, M.; Kowalski, A.K.; Craigie, R.; Wilson, K.L. Barrier-to-autointegration factor: Major roles in chromatin decondensation and nuclear assembly. J. Cell Biol. 2002, 158, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Tifft, K.E.; Segura-Totten, M.; Lee, K.K.; Wilson, K.L. Barrier-to-autointegration factor-like (BAF-L): A proposed regulator of BAF. Exp. Cell Res. 2006, 312, 478–487. [Google Scholar] [CrossRef]
- Elkhatib, R.A.; Paci, M.; Boissier, R.; Longepied, G.; Auguste, Y.; Achard, V.; Bourgeois, P.; Levy, N.; Branger, N.; Mitchell, M.J.; et al. LEM-domain proteins are lost during human spermiogenesis but BAF and BAF-L persist. Reproduction 2017, 154, 387–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maratou, K.; Forster, T.; Costa, Y.; Taggart, M.; Speed, R.M.; Ireland, J.; Teague, P.; Roy, D.; Cooke, H.J. Expression profiling of the developing testis in wild-type and Dazl knockout mice. Mol. Reprod. Dev. 2004, 67, 26–54. [Google Scholar] [CrossRef]
- Paci, M.; Elkhatib, R.; Longepied, G.; Hennebicq, S.; Bessonat, J.; Courbiere, B.; Bourgeois, P.; Levy, N.; Mitchell, M.J.; Metzler-Guillemain, C. Abnormal retention of nuclear lamina and disorganization of chromatin-related proteins in spermatozoa from DPY19L2-deleted globozoospermic patients. Reprod. Biomed. Online 2017, 35, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Griswold, M.D. Spermatogenesis: The commitment to meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ernst, C.; Eling, N.; Martinez-Jimenez, C.P.; Marioni, J.C.; Odom, D.T. Staged developmental mapping and X chromosome transcriptional dynamics during mouse spermatogenesis. Nat. Commun. 2019, 10, 1251. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, K.; Sugiyama, S.; Osouda, S.; Goto, H.; Inagaki, M.; Horigome, T.; Omata, S.; McConnell, M.; Fisher, P.A.; Nishida, Y. Barrier-to-autointegration factor plays crucial roles in cell cycle progression and nuclear organization in Drosophila. J. Cell Sci. 2003, 116 Pt 18, 3811–3823. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, R. Regulation of spermatogenesis. Physiol. Reprod. 1994, 1, 1363–1434. [Google Scholar]
- Oatley, J.M.; Brinster, R.L. Regulation of spermatogonial stem cell self-renewal in mammals. Annu. Rev. Cell Dev. Biol. 2008, 24, 263–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Bai, X.; Zhang, D.; Han, C.; Yuan, J.; Liu, W.; Cao, X.; Chen, Z.; Shangguan, F.; Zhu, Z.; et al. Mammalian elongation factor 4 regulates mitochondrial translation essential for spermatogenesis. Nat. Struct. Mol. Biol. 2016, 23, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, J.; Kim, S.Y.; Zhao, M.; Hearn, S.A.; Zhang, M.Q.; Meistrich, M.L.; Mills, A.A. Chd5 orchestrates chromatin remodelling during sperm development. Nat. Commun. 2014, 5, 3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, J.; Mesbah, M.; Huber, C.; Dadoune, J. Aniline blue staining as a marker of sperm chromatin defects associated with different semen characteristics discriminates between proven fertile and suspected infertile men. Int. J. Androl. 1990, 13, 452–462. [Google Scholar] [CrossRef]
- Lu, L.Y.; Wu, J.; Ye, L.; Gavrilina, G.B.; Saunders, T.L.; Yu, X. RNF8-dependent histone modifications regulate nucleosome removal during spermatogenesis. Dev. Cell 2010, 18, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhao, R.; Guo, C.; Jiang, S.; Yang, J.; Xu, Y.; Liu, Y.; Fan, L.; Xiong, W.; Ma, J.; et al. Knockout of BRD7 results in impaired spermatogenesis and male infertility. Sci. Rep. 2016, 6, 21776. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.R.; Noda, T.; Kim, H.; Kim, G.; Park, S.; Na, Y.; Oura, S.; Shimada, K.; Bang, I.; Ahn, J.Y.; et al. PHF7 Modulates BRDT Stability and Histone-to-Protamine Exchange during Spermiogenesis. Cell Rep. 2020, 32, 107950. [Google Scholar] [CrossRef]
- World-Health-Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Nair, M.; Nagamori, I.; Sun, P.; Mishra, D.P.; Rheaume, C.; Li, B.; Sassone-Corsi, P.; Dai, X. Nuclear regulator Pygo2 controls spermiogenesis and histone H3 acetylation. Dev. Biol. 2008, 320, 446–455. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Scott, G.; Ray, M.K.; Mishina, Y.; Zhang, Y. Histone demethylase JHDM2A is critical for Tnp1 and Prm1 transcription and spermatogenesis. Nature 2007, 450, 119–123. [Google Scholar] [CrossRef]
- Niu, C.M.; Xia, M.M.; Zhong, Y.N.; Zheng, Y. Mus musculus Barrier-To-Autointegration Factor 2 (Banf2) is Not Essential for Spermatogenesis or Fertility. Cytogenet. Genome Res. 2021, 161, 167–177. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Rivolta, C.; Li, L.Y.; Peng, G.H.; Swain, P.K.; Sung, C.H.; Swaroop, A.; Berson, E.L.; Dryja, T.P.; et al. Barrier to autointegration factor interacts with the cone-rod homeobox and represses its transactivation function. J. Biol. Chem. 2002, 277, 43288–43300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, F.; Cheng, Y.; Breschi, A.; Vierstra, J.; Wu, W.; Ryba, T.; Sandstrom, R.; Ma, Z.; Davis, C.; Pope, B.D.; et al. A comparative encyclopedia of DNA elements in the mouse genome. Nature 2014, 515, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Gao, H.; Li, W.; Liu, C. Essential Role of Histone Replacement and Modifications in Male Fertility. Front. Genet. 2019, 10, 962. [Google Scholar] [CrossRef] [PubMed]
- Grunstein, M. Histone acetylation in chromatin structure and transcription. Nature 1997, 389, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Isono, K.I.; Ohbo, K.; Endo, T.A.; Ohara, O.; Maekawa, M.; Toyama, Y.; Ito, C.; Toshimori, K.; Helin, K.; et al. EPC1/TIP60-Mediated Histone Acetylation Facilitates Spermiogenesis in Mice. Mol. Cell. Biol. 2017, 37, e00082-17. [Google Scholar] [CrossRef] [Green Version]
- Curi, S.M.; Ariagno, J.I.; Chenlo, P.H.; Mendeluk, G.R.; Pugliese, M.N.; Sardi Segovia, L.M.; Repetto, H.E.; Blanco, A.M. Asthenozoospermia: Analysis of a large population. Arch. Androl. 2003, 49, 343–349. [Google Scholar] [CrossRef]
- Shahrokhi, S.Z.; Salehi, P.; Alyasin, A.; Taghiyar, S.; Deemeh, M.R. Asthenozoospermia: Cellular and molecular contributing factors and treatment strategies. Andrologia 2020, 52, e13463. [Google Scholar] [CrossRef]
- Ravina, C.G.; Seda, M.; Pinto, F.M.; Orea, A.; Fernandez-Sanchez, M.; Pintado, C.O.; Candenas, M.L. A role for tachykinins in the regulation of human sperm motility. Hum. Reprod. 2007, 22, 1617–1625. [Google Scholar] [CrossRef] [Green Version]
- Preece, C.; Alghadban, S.; Bouchareb, A.; Moralli, D.; Biggs, D.; Davies, B. Replacement of surgical vasectomy through the use of wild-type sterile hybrids. Lab Anim. 2021, 50, 49–52. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5439. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Mahmood, T.; Yang, P.C. Western blot: Technique, theory and trouble shooting. N. Am. J. Med. Sci. 2014, 6, 160. [Google Scholar]
- Biggers, J.; Whitten, W.; Whittingham, D. The culture of mouse embryos in vitro. Fac. Res. 1970–1979 1971, 193. Available online: https://mouseion.jax.org/stfb1970_1979/193 (accessed on 29 January 2022).
- DeCaprio, J.; Kohl, T.O. Immunoprecipitation. Cold Spring Harb. Protoc. 2017, 2017, pdb.prot098640. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Source | Identifier/Application |
|---|---|---|
| GATA-4(D3A3M) Rabbit mAb | Cell Signaling Technology | Cat#36966, IHC: 1:800 |
| SCP3 Rabbit pAb | Abcam | ab15093, IHC: 1:500 |
| ACRV1 Rabbit pAb | Proteintech | Cat#14040-1-AP, IHC: 1:500 |
| pCAM Rabbit mAb | ABclonal | Cat#A19301 IHC: 1:100, WB: 1:500 |
| PRM1 Rabbit pAb | Proteintech | Cat#15697-1-AP WB: 1:1000 |
| BAF-L Rabbit pAb | Generated by immunizing rabbits with full-length GST fusion protein of mouse BAF-L | IHC: 1:200, IP: 1:100, WB: 1:1000 |
| Histone H4 Rabbit mAb | ABclonal | A19815, IF: 1:100, IP: 1:100, WB: 1:1000 |
| Histone H3 Rabbit mAb | ABclonal | A17562, IF: 1:100, IP: 1:100, WB: 1: 1000 |
| Histone H2A Rabbit mAb | ABclonal | A3692, IF: 1:100, WB: 1:1000 |
| Histone H2B Rabbit mAb | ABclonal | A19812, WB: 1:1000 |
| Ac-Histone H4 (E-5) Mouse mAb | Santa Cruz | sc-377520, WB: 1:500 |
| Ac-Histone H3 (AH3-120) Mouse mAb | Santa Cruz | sc-56616, WB: 1:500 |
| Phospho-Histone H2A.X(Ser139) (20E3) Rabbit mAb | Cell Signaling Technology | Cat#9718, IF: 1:400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Gong, H.; Mu, B.; Lan, X.; Yang, C.; Tan, J.; Liu, W.; Zou, Y.; Li, L.; Feng, B.; et al. BAF-L Modulates Histone-to-Protamine Transition during Spermiogenesis. Int. J. Mol. Sci. 2022, 23, 1985. https://doi.org/10.3390/ijms23041985
Huang C, Gong H, Mu B, Lan X, Yang C, Tan J, Liu W, Zou Y, Li L, Feng B, et al. BAF-L Modulates Histone-to-Protamine Transition during Spermiogenesis. International Journal of Molecular Sciences. 2022; 23(4):1985. https://doi.org/10.3390/ijms23041985
Chicago/Turabian StyleHuang, Chao, Huan Gong, Bin Mu, Xinting Lan, Chengcheng Yang, Jinlong Tan, Wentao Liu, Yuanfeng Zou, Lixia Li, Bin Feng, and et al. 2022. "BAF-L Modulates Histone-to-Protamine Transition during Spermiogenesis" International Journal of Molecular Sciences 23, no. 4: 1985. https://doi.org/10.3390/ijms23041985
APA StyleHuang, C., Gong, H., Mu, B., Lan, X., Yang, C., Tan, J., Liu, W., Zou, Y., Li, L., Feng, B., He, X., Luo, Q., & Chen, Z. (2022). BAF-L Modulates Histone-to-Protamine Transition during Spermiogenesis. International Journal of Molecular Sciences, 23(4), 1985. https://doi.org/10.3390/ijms23041985