
Matrix Metalloproteinases in Helicobacter pylori–Associated Gastritis and Gastric Cancer


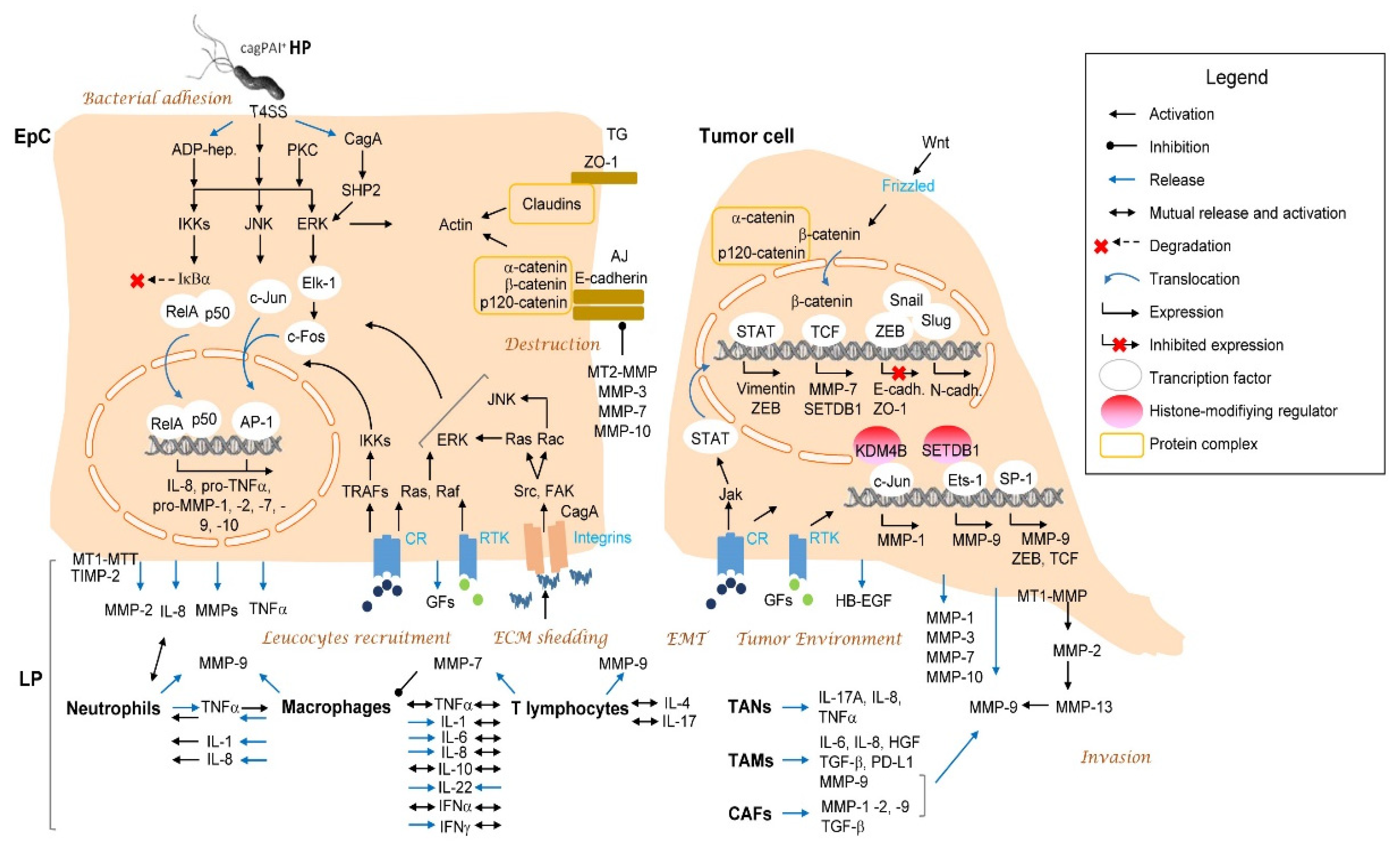{kind=link}
Abstract
1. Introduction
2. MMPs and Gastritis
2.1. MMP Levels and Localization
2.2. Bacterial Factors and Host Pathways Pivotal for MMP Activation
2.3. Cytokines and MMPs
2.4. MMPs in Ulcer
3. MMPs and Gastric Cancer
3.1. MMPs Expression in Tumor
3.2. MMPs in Tumor Microenvironment
3.3. Pathways of MMPs Activation in Gastric Cancer
3.4. MMPs and E-Cadherin
4. MMPs as Tools and Targets—Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pollard, T.D.; Earnshaw, W.C. Cell Biology; Elsevier Inc.: Philadelphia, PA, USA, 2008; pp. 468–472. [Google Scholar]
- Itoh, Y. Membrane-Type Matrix Metalloproteinases: Their Functions and Regulations. Matrix Biol. 2015, 44–46, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Jobin, P.G.; Butler, G.S.; Overall, C.M. New Intracellular Activities of Matrix Metalloproteinases Shine in the Moonlight. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864 Pt A, 2043–2055. [Google Scholar] [CrossRef] [PubMed]
- Chakraborti, S.; Mandal, M.; Das, S.; Mandal, A.; Chakraborti, T. Regulation of matrix metalloproteinases: An overview. Mol. Cell. Biochem. 2003, 253, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Dayer, C.; Stamenkovic, I. Recruitment of Matrix Metalloproteinase-9 (MMP-9) to the Fibroblast Cell Surface by Lysyl Hydroxylase 3 (LH3) Triggers Transforming Growth Factor-β (TGF-β) Activation and Fibroblast Differentiation. J. Biol. Chem. 2015, 290, 13763–13778. [Google Scholar] [CrossRef] [PubMed]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix Metalloproteinases and the Regulation of Tissue Remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Takino, T.; Watanabe, Y.; Matsui, M.; Miyamori, H.; Kudo, T.; Seiki, M.; Sato, H. Membrane-Type 1 Matrix Metalloproteinase Modulates Focal Adhesion Stability and Cell Migration. Exp. Cell Res. 2006, 312, 1381–1389. [Google Scholar] [CrossRef]
- Bassiouni, W.; Ali, M.A.M.; Schulz, R. Multifunctional Intracellular Matrix Metalloproteinases: Implications in Disease. FEBS J. 2021, 288, 7162–7182. [Google Scholar] [CrossRef]
- Vincenti, M.P.; Brinckerhoff, C.E. Signal Transduction and Cell-Type Specific Regulation of Matrix Metalloproteinase Gene Expression: Can MMPs Be Good for You? J. Cell. Physiol. 2007, 213, 355–364. [Google Scholar] [CrossRef]
- Madzharova, E.; Kastl, P.; Sabino, F.; Keller, U.A.D. Post-Translational Modification-Dependent Activity of Matrix Metalloproteinases. Int. J. Mol. Sci. 2019, 20, 3077. [Google Scholar] [CrossRef]
- McMahon, M.; Ye, S.; Pedrina, J.; Dlugolenski, D.; Stambas, J. Extracellular Matrix Enzymes and Immune Cell Biology. Front. Mol. Biosci. 2021, 8, 703868. [Google Scholar] [CrossRef]
- Correa, P. A Human Model of Gastric Carcinogenesis. Cancer Res. 1988, 48, 3554–3560. [Google Scholar] [PubMed]
- Businello, G.; Angerilli, V.; Parente, P.; Realdon, S.; Savarino, E.; Farinati, F.; Grillo, F.; Vanoli, A.; Galuppini, F.; Paccagnella, S.; et al. Molecular Landscapes of Gastric Pre-Neoplastic and Pre-Invasive Lesions. Int. J. Mol. Sci. 2021, 22, 9950. [Google Scholar] [CrossRef] [PubMed]
- Senchukova, M.A.; Tomchuk, O.; Shurygina, E.I. Helicobacter pylori in Gastric Cancer: Features of Infection and their Correlations with Long-Term Results of Treatment. World J. Gastroenterol. 2021, 27, 6290–6305. [Google Scholar] [CrossRef] [PubMed]
- Peek, R.M.; Blaser, M.J. Helicobacter pylori and Gastrointestinal Tract Adenocarcinomas. Nat. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Rad, R.; Dossumbekova, A.; Neu, B.; Lang, R.; Bauer, S.; Saur, D.; Gerhard, M.; Prinz, C. Cytokine Gene Polymorphisms Influence Mucosal Cytokine Expression, Gastric Inflammation, and Host Specific Colonisation during Helicobacter pylori Infection. Gut 2004, 53, 1082–1089. [Google Scholar] [CrossRef]
- Gobert, A.P.; Wilson, K.T. Human and Helicobacter pylori Interactions Determine the Outcome of Gastric Diseases. Mol. Pathog. Signal Transduct. Helicobacter Pylori 2017, 400, 27–52. [Google Scholar] [CrossRef]
- Sokolova, O.; Naumann, M. Crosstalk between DNA Damage and Inflammation in the Multiple Steps of Gastric Carcinogenesis. Mol. Mech. Inflamm. Induction Resolut. Escape Helicobacter Pylori 2019, 421, 107–137. [Google Scholar] [CrossRef]
- Sheu, B.-S.; Yang, H.-B.; Yeh, Y.-C.; Wu, J.-J. Helicobacter pylori colonization of the Human Gastric Epithelium: A Bug’s First Step Is a Novel Target for Us. J. Gastroenterol. Hepatol. 2010, 25, 26–32. [Google Scholar] [CrossRef]
- Posselt, G.; Backert, S.; Wessler, S. The Functional Interplay of Helicobacter pylori Factors with Gastric Epithelial Cells Induces a Multi-Step Process in Pathogenesis. Cell Commun. Signal. 2013, 11, 77. [Google Scholar] [CrossRef]
- Letley, D.P.; Rhead, J.L.; Twells, R.J.; Dove, B.; Atherton, J.C. Determinants of Non-Toxicity in the Gastric Pathogen Helicobacter pylori. J. Biol. Chem. 2003, 278, 26734–26741. [Google Scholar] [CrossRef]
- Censini, S.; Lange, C.; Xiang, Z.; Crabtree, J.E.; Ghiara, P.; Borodovsky, M.; Rappuoli, R.; Covacci, A. Cag, a Pathogenicity Island of Helicobacter pylori, Encodes Type I-Specific and Disease-Associated Virulence Factors. Proc. Natl. Acad. Sci. USA 1996, 93, 14648–14653. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, O.; Borgmann, M.; Rieke, C.; Schweitzer, K.; Rothkötter, H.-J.; Naumann, M. Helicobacter pylori Induces Type 4 Secretion System-Dependent, but CagA-Independent Activation of IκBs and NF-κB/RelA at Early Time Points. Int. J. Med. Microbiol. 2013, 303, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, O.; Vieth, M.; Naumann, M. Protein Kinase C Isozymes Regulate Matrix Metalloproteinase-1 Expression and Cell Invasion in Helicobacter pylori infection. Gut 2012, 62, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Odenbreit, S.; Püls, J.; Sedlmaier, B.; Gerland, E.; Fischer, W.; Haas, R. Translocation of Helicobacter pylori CagA into Gastric Epithelial Cells by Type IV Secretion. Science 2000, 287, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Kanemitsu, A.; Knight, C.T.; Hatakeyama, M. Molecular Anatomy and Pathogenic Actions of Helicobacter pylori CagA that underpin Gastric Carcinogenesis. Cell. Mol. Immunol. 2019, 17, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Casarotto, M.; Pratesi, C.; Bidoli, E.; Maiero, S.; Magris, R.; Steffan, A.; Basaglia, G.; Canzonieri, V.; De Re, V.; Cannizzaro, R.; et al. Differential Helicobacter pylori Plasticity in the Gastric Niche of Subjects at Increased Gastric Cancer Risk. Pathogens 2019, 8, 65. [Google Scholar] [CrossRef]
- Stein, S.C.; Faber, E.; Bats, S.H.; Murillo, T.; Speidel, Y.; Coombs, N.; Josenhans, C. Helicobacter pylori Modulates Host Cell Responses by CagT4SS-Dependent Translocation of an Intermediate Metabolite of LPS Inner Core Heptose Biosynthesis. PLoS Pathog. 2017, 13, e1006514. [Google Scholar] [CrossRef]
- Zimmermann, S.; Pfannkuch, L.; Al-Zeer, M.A.; Bartfeld, S.; Koch, M.; Liu, J.; Rechner, C.; Soerensen, M.; Sokolova, O.; Zamyatina, A.; et al. ALPK1- and TIFA-Dependent Innate Immune Response Triggered by the Helicobacter pylori Type IV Secretion System. Cell Rep. 2017, 20, 2384–2395. [Google Scholar] [CrossRef]
- Pfannkuch, L.; Hurwitz, R.; Trauisen, J.; Sigulla, J.; Poeschke, M.; Matzner, L.; Kosma, P.; Schmid, M.; Meyer, T.F. ADP Heptose, a Novel Pathogen-Associated Molecular Pattern Identified in Helicobacter pylori. FASEB J. 2019, 33, 9087–9099. [Google Scholar] [CrossRef]
- Toyoda, T.; Tsukamoto, T.; Takasu, S.; Shi, L.; Hirano, N.; Ban, H.; Kumagai, T.; Tatematsu, M. Anti-Inflammatory Effects of Caffeic Acid Phenethyl Ester (CAPE), a Nuclear Factor-κB Inhibitor, on Helicobacter pylori-Induced Gastritis in Mongolian Gerbils. Int. J. Cancer 2009, 125, 1786–1795. [Google Scholar] [CrossRef]
- Cook, K.W.; Letley, D.P.; Ingram, R.J.M.; Staples, E.; Skjoldmose, H.; Atherton, J.C.; Robinson, K. CCL20/CCR6-Mediated Migration of Regulatory T Cells to the Helicobacter pylori-Infected Human Gastric Mucosa. Gut 2014, 63, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-Y.; Liu, D.-J.; Liu, M.-X. The Protective Effect of NF-κB Signaling Pathway Inhibitor PDTC on Mice with Chronic Atrophic Gastritis. Scand. J. Gastroenterol. 2021, 56, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Posselt, G.; Crabtree, J.E.; Wessler, S. Proteolysis in Helicobacter pylori-Induced Gastric Cancer. Toxins 2017, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Argent, R.H.; Atherton, J.C. The Inflammatory and Immune Response to Helicobacter pylori Infection. Best Pract. Res. Clin. Gastroenterol. 2007, 21, 237–259. [Google Scholar] [CrossRef]
- Krakowiak, M.S.; Noto, J.M.; Piazuelo, M.B.; Hardbower, D.M.; Romero-Gallo, J.; Delgado, A.; Chaturvedi, R.; Correa, P.; Wilson, K.T.; Peek, R.M. Matrix Metalloproteinase 7 Restrains Helicobacter pylori-Induced Gastric Inflammation and Premalignant lesions in the Stomach by Altering Macrophage Polarization. Oncogene 2014, 34, 1865–1871. [Google Scholar] [CrossRef]
- Lv, Y.-P.; Cheng, P.; Zhang, J.-Y.; Mao, F.-Y.; Teng, Y.-S.; Liu, Y.-G.; Kong, H.; Wu, X.-L.; Hao, C.-J.; Han, B.; et al. Helicobacter pylori–Induced Matrix Metallopeptidase-10 Promotes Gastric Bacterial Colonization and Gastritis. Sci. Adv. 2019, 5, eaau6547. [Google Scholar] [CrossRef]
- Koyama, S. Significance of Cell-Surface Expression of Matrix Metalloproteinases and their Inhibitors on Gastric Epithelium and Infiltrating Mucosal Lymphocytes in Progression of Helicobacter pylori-Associated Gastritis. Scand. J. Gastroenterol. 2004, 39, 1046–1053. [Google Scholar] [CrossRef]
- Mori, N.; Sato, H.; Hayashibara, T.; Senba, M.; Geleziunas, R.; Wada, A.; Hirayama, T.; Yamamoto, N. Helicobacter pylori Induces Matrix Metalloproteinase-9 through Activation of Nuclear Factor κB. Gastroenterology 2003, 124, 983–992. [Google Scholar] [CrossRef]
- Bergin, P.J.; Anders, E.; Sicheng, W.; Erik, J.; Jennie, A.; Hans, L.; Pierre, M.; Qiang, P.-H.; Marianne, Q.-J. Increased Production of Matrix Metalloproteinases in Helicobacter pylori-Associated Human Gastritis. Helicobacter 2004, 9, 201–210. [Google Scholar] [CrossRef]
- Li, S.-L.; Zhao, J.-R.; Ren, X.-Y.; Xie, J.-P.; Ma, Q.-Z.; Rong, Q.-H. Increased Expression of Matrix Metalloproteinase-9 Associated with Gastric Ulcer Recurrence. World J. Gastroenterol. 2013, 19, 4590–4595. [Google Scholar] [CrossRef]
- Kubben, F.J.G.M.; Sier, C.F.M.; Schram, M.; Witte, T.A.M.C.; Veenendaal, R.A.; Van Duijn, W.; Verheijen, J.H.; Hanemaaijer, R.; Lamers, C.B.H.W.; Verspaget, H.W. Eradication of Helicobacter pylori Infection Favourably Affects Altered Gastric Mucosal MMP-9 Levels. Helicobacter 2007, 12, 498–504. [Google Scholar] [CrossRef]
- Wroblewski, L.E.; Noble, P.-J.M.; Pagliocca, A.; Pritchard, D.M.; Hart, C.A.; Campbell, F.; Dodson, A.R.; Dockray, G.J.; Varro, A. Stimulation of MMP-7 (matrilysin) by Helicobacter pylori in Human Gastric Epithelial Cells: Role in Epithelial Cell Migration. J. Cell Sci. 2003, 116, 3017–3026. [Google Scholar] [CrossRef]
- Bebb, J.R.; Letley, D.P.; Thomas, R.J.; Aviles, F.; Collins, H.; Watson, S.A.; Hand, N.M.; Zaitoun, A.; Atherton, J.C. Helicobacter pylori Upregulates Matrilysin (MMP-7) in Epithelial Cells in Vivo and in Vitro in a Cag Dependent Manner. Gut 2003, 52, 1408–1413. [Google Scholar] [CrossRef]
- Chung, W.C.; Jung, S.H.; Lee, K.-M.; Paik, C.N.; Kawk, J.W.; Jung, J.H.; Lee, M.K.; Lee, Y.K. The Detection of Helicobacter pylori Cag Pathogenicity Islands (PAIs) and Expression of Matrix Metalloproteinase-7 (MMP-7) in Gastric Epithelial Dysplasia and Intramucosal Cancer. Gastric Cancer 2010, 13, 162–169. [Google Scholar] [CrossRef]
- Sadeghiani, M.; Bagheri, N.; Shahi, H.; Reiisi, S.; Rahimian, G.; Rashidi, R.; Mahsa, M.; Shafigh, M.; Salimi, E.; Rafieian-Kopaei, M.; et al. cag Pathogenicity Island-Dependent Upregulation of Matrix Metalloproteinase-7 in Infected Patients with Helicobacter pylori. J. Immunoass. Immunochem. 2017, 38, 595–607. [Google Scholar] [CrossRef]
- Varro, A.; Kenny, S.; Hemers, E.; McCaig, C.; Przemeck, S.; Wang, T.C.; Bodger, K.; Pritchard, D.M. Increased Gastric Expression of MMP-7 in Hypergastrinemia and Significance for Epithelial-Mesenchymal Signaling. Am. J. Physiol. Liver Physiol. 2007, 292, G1133–G1140. [Google Scholar] [CrossRef]
- Yin, Y.; Grabowska, A.M.; Clarke, P.A.; Whelband, E.; Robinson, K.; Argent, R.H.; Tobias, A.; Kumari, R.; Atherton, J.C.; Watson, S.A. Helicobacter pylori Potentiates Epithelial: Mesenchymal Transition in Gastric Cancer: Links to Soluble HB-EGF, Gastrin and Matrix Metalloproteinase-7. Gut 2010, 59, 1037–1045. [Google Scholar] [CrossRef]
- Ogden, S.R.; Noto, J.M.; Allen, S.S.; Patel, D.A.; Romero-Gallo, J.; Washington, M.K.; Fingleton, B.; Israel, D.A.; Lewis, N.; Wilson, K.; et al. Matrix Metalloproteinase-7 and Premalignant Host Responses in Helicobacter pylori–Infected Mice. Cancer Res. 2010, 70, 30–35. [Google Scholar] [CrossRef]
- Wilson, C.L.; Ouellette, A.J.; Satchell, D.P.; Ayabe, T.; López-Boado, Y.S.; Stratman, J.L.; Hultgren, S.J.; Matrisian, L.M.; Parks, W.C. Regulation of Intestinal α-Defensin Activation by the Metalloproteinase Matrilysin in Innate Host Defense. Science 1999, 286, 113–117. [Google Scholar] [CrossRef]
- Rautelin, H.; Tervahartiala, T.; Lauhio, A.; Sorsa, T.; Kolho, K.-L. Assessment of Systemic Matrix Metalloproteinase and their Regulator Response in Children with Helicobacter pylori gastritis. Scand. J. Clin. Lab. Investig. 2010, 70, 492–496. [Google Scholar] [CrossRef]
- Rautelin, H.I.; Oksanen, A.M.; Veijola, L.I.; Sipponen, P.I.; Tervahartiala, T.I.; Sorsa, T.A.; Lauhio, A. Enhanced Systemic Matrix Metalloproteinase Response in Helicobacter pylori Gastritis. Ann. Med. 2009, 41, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.-C.; Sheu, B.-S.; Cheng, H.-C.; Wang, Y.-L.; Yang, H.-B.; Wu, J.-J. Elevated Serum Matrix Metalloproteinase-3 and -7 in H. pylori-Related Gastric Cancer Can Be Biomarkers Correlating with a Poor Survival. Am. J. Dig. Dis. 2009, 55, 1649–1657. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-J. Inflammation-Related Factors Predicting Prognosis of Gastric Cancer. World J. Gastroenterol. 2014, 20, 4586–4596. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Lu, H.; Sun, Y.; Graham, D.Y.; Cheung, H.S.; Yamaoka, Y. Balance between Polyoma Enhancing Activator 3 and Activator Protein 1 Regulates Helicobacter pylori–Stimulated Matrix Metalloproteinase 1 Expression. Cancer Res. 2006, 66, 5111–5120. [Google Scholar] [CrossRef]
- Pillinger, M.; Marjanovic, N.; Kim, S.-Y.; Lee, Y.-C.; Scher, J.U.; Roper, J.; Abeles, A.; Izmirly, P.I.; Axelrod, M.; Pillinger, M.; et al. Helicobacter pylori Stimulates Gastric Epithelial Cell MMP-1 Secretion via CagA-Dependent and -Independent ERK Activation. J. Biol. Chem. 2007, 282, 18722–18731. [Google Scholar] [CrossRef]
- Crawford, H.C.; Krishna, U.S.; Israel, D.A.; Matrisian, L.M.; Washington, M.; Peek, R.M. Helicobacter pylori Strain-Selective Induction of Matrix Metalloproteinase-7 in Vitro and within Gastric Mucosa. Gastroenterology 2003, 125, 1125–1136. [Google Scholar] [CrossRef]
- Ogden, S.R.; Wroblewski, L.E.; Weydig, C.; Romero-Gallo, J.; O’Brien, D.P.; Israel, D.A.; Krishna, U.S.; Fingleton, B.; Reynolds, A.B.; Wessler, S.; et al. p120 and Kaiso Regulate Helicobacter pylori-Induced Expression of Matrix Metalloproteinase-7. Mol. Biol. Cell 2008, 19, 4110–4121. [Google Scholar] [CrossRef]
- Oliveira, M.J.; Costa, A.; Costa, A.; Henriques, L.; Suriano, G.; Atherton, J.C.; Machado, J.C.; Carneiro, F.; Seruca, R.; Mareel, M.; et al. Helicobacter pylori Induces Gastric Epithelial Cell Invasion in a c-Met and Type IV Secretion System-dependent Manner. J. Biol. Chem. 2006, 281, 34888–34896. [Google Scholar] [CrossRef]
- Yang, Y.; Li, X.; Du, J.; Yin, Y.; Li, Y. Involvement of MicroRNAs-MMPs-E-Cadherin in the Migration and Invasion of Gastric Cancer Cells Infected with Helicobacter pylori. Exp. Cell Res. 2018, 367, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Sougleri, I.S.; Papadakos, K.; Zadik, M.P.; Mavri-Vavagianni, M.; Mentis, A.F.; Sgouras, D.N. Helicobacter pylori CagA Protein Induces Factors Involved in the Epithelial to Mesenchymal Transition (EMT) in Infected Gastric Epithelial Cells in an EPIYA-Phosphorylation-Dependent Manner. FEBS J. 2015, 283, 206–220. [Google Scholar] [CrossRef]
- Costa, A.; Ferreira, R.; Pinto-Ribeiro, I.; Sougleri, I.S.; Oliveira, M.J.; Carreto, L.; Santos, M.; Sgouras, D.N.; Carneiro, F.; Leite, M.; et al. Helicobacter pylori Activates Matrix Metalloproteinase 10 in Gastric Epithelial Cells via EGFR and ERK-Mediated Pathways. J. Infect. Dis. 2016, 213, 1767–1776. [Google Scholar] [CrossRef]
- Chang, Y.W.; Oh, H.-C.; Jang, J.Y.; Hwangbo, Y.; Lee, J.W.; Lee, H.J.; Joo, K.R.; Dong, S.H.; Kim, S.S.; Kim, H.J.; et al. IL-1β and IL-8, Matrix Metalloproteinase 3, and Pepsinogen Secretion before and after H. pylori eradication in Gastroduodenal Phenotypes. Scand. J. Gastroenterol. 2008, 43, 1184–1193. [Google Scholar] [CrossRef]
- Song, J.H.; Kim, S.G.; Jung, S.-A.; Lee, M.K.; Jung, H.C.; Song, I.S. The Interleukin-8–251 AA Genotype Is Associated with Angiogenesis in Gastric Carcinogenesis in Helicobacter pylori–Infected Koreans. Cytokine 2010, 51, 158–165. [Google Scholar] [CrossRef]
- Zhuang, Y.; Cheng, P.; Liu, X.-F.; Peng, L.-S.; Li, B.-S.; Wang, T.-T.; Chen, N.; Li, W.-H.; Shi, Y.; Chen, W.; et al. A Pro-Inflammatory Role for Th22 Cells in Helicobacter pylori–Associated Gastritis. Gut 2014, 64, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Piazuelo, M.B.; Riechelmann, R.P.; Wilson, K.T.; Algood, H.M.S. Resolution of Gastric Cancer-Promoting Inflammation: A Novel Strategy for Anti-cancer Therapy. Mol. Mech. Inflamm. Induction Resolut. Escape Helicobacter Pylori 2019, 421, 319–359. [Google Scholar] [CrossRef]
- Bodger, K.; Ahmed, S.; Pazmany, L.; Pritchard, D.M.; Micheal, A.; Khan, A.L.; Dimaline, R.; Dockray, G.J.; Varro, A. Altered Gastric Corpus Expression of Tissue Inhibitors of Metalloproteinases in Human and Murine Helicobacter Infection. J. Clin. Pathol. 2007, 61, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Hahm, K.B.; Kim, D.H.; Lee, K.M.; Lee, J.S.; Surh, Y.J.; Kim, Y.B.; Yoo, B.M.; Kim, J.H.; Joo, H.J.; Cho, Y.K.; et al. Effect of Long-Term Administration of Rebamipide on Helicobacter pylori Infection in Mice. Aliment. Pharmacol. Ther. 2003, 18, 24–38. [Google Scholar] [CrossRef]
- Cheng, H.-C.; Yang, H.-B.; Chang, W.-L.; Chen, W.-Y.; Yeh, Y.-C.; Sheu, B.-S. Expressions of MMPs and TIMP-1 in Gastric Ulcers May Differentiate H. pylori-Infected from NSAID-Related Ulcers. Sci. World J. 2012, 2012, 539316. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, N.; Sadeghiani, M.; Rahimian, G.; Mahsa, M.; Shafigh, M.; Rafieian-Kopaei, M.; Shirzad, H. Correlation between Expression of MMP-9 and MMP-3 in Helicobacter pylori Infected Patients with Different Gastroduodenal Diseases. Arab J. Gastroenterol. 2018, 19, 148–154. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424, Erratum in CA Cancer J. Clin. 2020, 70, 313. [Google Scholar] [CrossRef]
- Laurén, P. The Two Histological Main Types of Gastric Carcinoma: Diffuse and So-Called Intestinal-Type Carcinoma. An Attempt at a Histo-Clinical Classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Hippo, Y.; Taniguchi, H.; Tsutsumi, S.; Machida, N.; Chong, J.-M.; Fukayama, M.; Kodama, T.; Aburatani, H. Global gene expression analysis of gastric cancer by oligonucleotide microarrays. Cancer Res. 2002, 62, 233–240. [Google Scholar] [PubMed]
- Murray, G.I.; Duncan, M.E.; Arbuckle, E.; Melvin, W.T.; Fothergill, J.E. Matrix Metalloproteinases and their Inhibitors in Gastric Cancer. Gut 1998, 43, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Yashiro, M.; Nishimura, S.; Maeda, K.; Sawada, T.; Ogawa, Y.; Sowa, M.; Chung, K.H. Matrix Metalloproteinase-1 Expression Is a Prognostic Factor for Patients with Advanced Gastric Cancer. Int. J. Mol. Med. 1999, 4, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Sier, C.; Kubben, F.; Ganesh, S.; Heerding, M.; Griffioen, G.; Hanemaaijer, R.; Van Krieken, J.; Lamers, C.; Verspaget, H.; Sier, C. Tissue Levels of Matrix Metalloproteinases MMP-2 and MMP-9 Are Related to the Overall Survival of Patients with Gastric Carcinoma. Br. J. Cancer 1996, 74, 413–417. [Google Scholar] [CrossRef]
- Wroblewski, L.E.; Pritchard, D.M.; Carter, S.; Varro, A. Gastrin-Stimulated Gastric Epithelial Cell Invasion: The Role and Mechanism of Increased Matrix Metalloproteinase 9 Expression. Biochem. J. 2002, 365, 873–879. [Google Scholar] [CrossRef]
- Gao, H.; Lan, X.; Li, S.; Xue, Y. Relationships of MMP-9, E-Cadherin, and VEGF Expression with Clinicopathological Features and Response to Chemosensitivity in Gastric Cancer. Tumor Biol. 2017, 39, 1010428317698368. [Google Scholar] [CrossRef]
- Sheibani, S.; Mahmoudian, R.A.; Abbaszadegan, M.R.; Chamani, J.; Memar, B.; Gholamin, M. Expression Analysis of Matrix Metalloproteinase-13 in Human Gastric Cancer in the Presence of Helicobacter pylori Infection. Cancer Biomark. 2017, 18, 349–356. [Google Scholar] [CrossRef]
- Adachi, Y.; Itoh, F.; Yamamoto, H.; Matsuno, K.; Arimura, Y.; Kusano, M.; Endoh, T.; Hinoda, Y.; Oohara, M.; Hosokawa, M.; et al. Matrix Metalloproteinase Matrilysin (MMP-7) Participates in the Progression of Human Gastric and Esophageal Cancers. Int. J. Oncol. 1998, 13, 1031–1036. [Google Scholar] [CrossRef]
- Senota, A.; Itoh, F.; Yamamoto, H.; Adachi, Y.; Hinoda, Y.; Imai, K. Relation of Matrilysin Messenger RNA Expression with Invasive Activity in Human Gastric Cancer. Clin. Exp. Metastasis 1997, 16, 313–321. [Google Scholar] [CrossRef]
- Yamashita, K.; Azumano, I.; Mai, M.; Okada, Y. Expression and Tissue Localization of Matrix Metalloproteinase 7 (Matrilysin) in Human Gastric Carcinomas. Implications for Vessel Invasion and Metastasis. Int. J. Cancer 1998, 79, 187–194. [Google Scholar] [CrossRef]
- Zheng, H.-C.; Sun, J.-M.; Li, X.-H.; Yang, X.-F.; Zhang, Y.-C.; Xin, Y. Role of PTEN and MMP-7 Expression in Growth, Invasion, Metastasis and Angiogenesis of Gastric Carcinoma. Pathol. Int. 2003, 53, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Ajisaka, H.; Fushida, S.; Yonemura, Y.; Miwa, K. Expression of Insulin-Like Growth Factor-2, C-MET, Matrix Metalloproteinase-7 and MUC-1 in Primary Lesions and Lymph Node Metastatic Lesions of Gastric Cancer. Hepatogastroenterology 2002, 48, 1788–1792. [Google Scholar]
- Soleyman-Jahi, S.; Nedjat, S.; Abdirad, A.; Hoorshad, N.; Heidari, R.; Zendehdel, K. Prognostic Significance of Matrix Metalloproteinase-7 in Gastric Cancer Survival: A Meta-Analysis. PLoS ONE 2015, 10, e0122316. [Google Scholar] [CrossRef]
- Nomura, H.; Sato, H.; Seiki, M.; Mai, M.; Okada, Y. Expression of Membrane-Type Matrix Metalloproteinase in Human Gastric Carcinomas. Cancer Res. 1995, 55, 3263–3266. [Google Scholar]
- He, L.; Chu, D.; Li, X.; Zheng, J.; Liu, S.; Li, J.; Zhao, Q.; Ji, G. Matrix Metalloproteinase-14 Is a Negative Prognostic Marker for Patients with Gastric Cancer. Am. J. Dig. Dis. 2013, 58, 1264–1270. [Google Scholar] [CrossRef]
- Ogawa, S.; Kubo, H.; Murayama, Y.; Kubota, T.; Yubakami, M.; Matsumoto, T.; Ohashi, T.; Okamoto, K.; Kuriki, Y.; Hanaoka, K.; et al. Matrix Metalloprotease–14 Is a Target Enzyme for Detecting Peritoneal Metastasis in Gastric Cancer. Photodiagn. Photodyn. Ther. 2021, 35, 102420. [Google Scholar] [CrossRef]
- Ueda, J.; Kajita, M.; Suenaga, N.; Fujii, K.; Seiki, M. Sequence-Specific Silencing of MT1-MMP Expression Suppresses Tumor Cell Migration and Invasion: Importance of MT1-MMP as a Therapeutic Target for Invasive Tumors. Oncogene 2003, 22, 8716–8722. [Google Scholar] [CrossRef][Green Version]
- Levin, M.; Udi, Y.; Solomonov, I.; Sagi, I. Next Generation Matrix Metalloproteinase Inhibitors—Novel Strategies Bring New Prospects. Biochim. Biophys. Acta 2017, 1864, 1927–1939. [Google Scholar] [CrossRef]
- Sakurai, Y.; Otani, Y.; Kameyama, K.; Igarashi, N.; Kubota, T.; Kumai, K.; Kitajima, M. The Role of Stromal Cells in the Expression of Interstitial Collagenase (Matrix Metalloproteinase-1) in the Invasion of Gastric Cancer. J. Surg. Oncol. 1997, 66, 168–172. [Google Scholar] [CrossRef]
- Yokoyama, T.; Otani, Y.; Kurihara, N.; Sakurai, Y.; Kameyama, K.; Suzuki, H.; Igarashi, N.; Kimata, M.; Wada, N.; Kubota, T.; et al. Matrix Metalloproteinase Expression in Cultured Human Gastric Wall Fibroblasts-Interactions with Helicobacter pylori Isolated from Patients with Ulcers. Aliment. Pharmacol. Ther. 2000, 14, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.L.; Eslami, G.; Goudarzi, H.; Hajishafieeha, Z.; Soleimani, M.; Mohammadzadeh, A.; Ardeshirylajimi, A. Role of Helicobacter pylori on Cancer of Human Adipose-Derived Mesenchymal Stem Cells and Metastasis of Tumor Cells—An In Vitro Study. Tumor Biol. 2015, 37, 3371–3378. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cong, X.; Gao, H.; Lan, X.; Li, Z.; Wang, W.; Song, S.; Wang, Y.; Li, C.; Zhang, H.; et al. Correction to: Tumor-Associated Neutrophils Induce EMT by IL-17a to Promote Migration and Invasion in Gastric Cancer Cells. J. Exp. Clin. Cancer Res. 2019, 38, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wu, X.; Wang, X.; Yu, Z.; Pan, T.; Li, Z.; Chang, X.; Jin, Z.; Li, J.; Zhu, Z.; et al. The Reciprocal Interaction between Tumor Cells and Activated Fibroblasts Mediated by TNF-α/IL-33/ST2L Signaling Promotes Gastric Cancer Metastasis. Oncogene 2019, 39, 1414–1428. [Google Scholar] [CrossRef]
- Ren, Q.; Zhu, P.; Zhang, H.; Ye, T.; Liu, D.; Gong, Z.; Xia, X. Identification and Validation of Stromal-Tumor Microenvironment-Based Subtypes Tightly Associated with PD-1/PD-L1 Immunotherapy and Outcomes in Patients with Gastric Cancer. Cancer Cell Int. 2020, 20, 92. [Google Scholar] [CrossRef]
- Wang, Y.; Zhai, J.; Zhang, T.; Han, S.; Zhang, Y.; Yao, X.; Shen, L. Tumor-Associated Neutrophils Can Predict Lymph Node Metastasis in Early Gastric Cancer. Front. Oncol. 2020, 10, 1790. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, M.; Huang, F.; Yang, T.; Cai, J.; Zhang, X.; Zhu, W.; Qian, H.; Xu, W. H. pylori Infection-Induced MSC Differentiation into CAFs Promotes Epithelial-Mesenchymal Transition in Gastric Epithelial Cells. Int. J. Mol. Med. 2013, 32, 1465–1473. [Google Scholar] [CrossRef][Green Version]
- Wu, X.; Tao, P.; Zhou, Q.; Li, J.; Yu, Z.; Wang, X.; Li, J.; Li, C.; Yan, M.; Zhu, Z.; et al. IL-6 Secreted by Cancer-Associated Fibroblasts Promotes Epithelial-Mesenchymal Transition and Metastasis of Gastric Cancer via JAK2/STAT3 Signaling Pathway. Oncotarget 2017, 8, 20741–20750. [Google Scholar] [CrossRef]
- Liu, L.; Ye, Y.; Zhu, X. MMP-9 Secreted by Tumor Associated Macrophages Promoted Gastric Cancer Metastasis through a PI3K/AKT/Snail Pathway. Biomed. Pharmacother. 2019, 117, 109096. [Google Scholar] [CrossRef]
- Wu, J.; Chai, H.; Shan, H.; Pan, C.; Xu, X.; Dong, W.; Yu, J.; Gu, Y. Histone Methyltransferase SETD1A Induces Epithelial-Mesenchymal Transition to Promote Invasion and Metastasis Through Epigenetic Reprogramming of Snail in Gastric Cancer. Front. Cell Dev. Biol. 2021, 9, 657888. [Google Scholar] [CrossRef]
- Etoh, T.; Inoue, H.; Tanaka, S.; Barnard, G.F.; Kitano, S.; Mori, M. Angiopoietin-2 Is Related to Tumor Angiogenesis in Gastric Carcinoma: Possible in Vivo Regulation via Induction of Proteases. Cancer Res. 2001, 61, 2145–2153. [Google Scholar]
- Wang, H.; Wu, X.; Chen, Y. Stromal-Immune Score-Based Gene Signature: A Prognosis Stratification Tool in Gastric Cancer. Front. Oncol. 2019, 9, 1212. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Gong, J.; Li, X.; Hu, Z.; Xu, Y.; Shi, H.; Li, D.; Liu, G.; Jie, Y.; Hu, B.; et al. Unsupervised Hierarchical Clustering Identifies Immune Gene Subtypes in Gastric Cancer. Front. Pharmacol. 2021, 12, 692454. [Google Scholar] [CrossRef]
- Fanelli, M.F.; Chinen, L.T.D.; Begnami, M.D.; Costa, W.L.; Fregnami, J.H.T.; Soares, F.A.; Montagnini, A.L. The Influence of Transforming Growth Factor-α, Cyclooxygenase-2, Matrix Metalloproteinase (MMP)-7, MMP-9 and CXCR4 Proteins Involved in Epithelial-Mesenchymal Transition on Overall Survival of Patients with Gastric Cancer. Histopathology 2012, 61, 153–161. [Google Scholar] [CrossRef]
- Kubben, F.J.G.M.; Sier, C.F.M.; Van Duijn, W.; Griffioen, G.; Hanemaaijer, R.; Van De Velde, C.J.H.; Van Krieken, J.H.J.M.; Lamers, C.B.H.W.; Verspaget, H.W. Matrix Metalloproteinase-2 Is a Consistent Prognostic Factor in Gastric Cancer. Br. J. Cancer 2006, 94, 1035–1040. [Google Scholar] [CrossRef]
- Chu, D.; Zhang, Z.; Li, Y.; Zheng, J.; Dong, G.; Wang, W.; Ji, G. Matrix Metalloproteinase-9 Is Associated with Disease-Free Survival and Overall Survival in Patients with Gastric Cancer. Int. J. Cancer 2010, 129, 887–895. [Google Scholar] [CrossRef]
- Okusha, Y.; Eguchi, T.; Tran, M.T.; Sogawa, C.; Yoshida, K.; Itagaki, M.; Taha, E.A.; Ono, K.; Aoyama, E.; Okamura, H.; et al. Extracellular Vesicles Enriched with Moonlighting Metalloproteinase Are Highly Transmissive, Pro-Tumorigenic, and Trans-Activates Cellular Communication Network Factor (CCN2/CTGF): CRISPR against Cancer. Cancers 2020, 12, 881. [Google Scholar] [CrossRef]
- Pillinger, M.H.; Marjanovic, N.; Kim, S.-Y.; Scher, J.U.; Izmirly, P.; Tolani, S.; Dinsell, V.; Lee, Y.C.; Blaser, M.J.; Abramson, S. Matrix Metalloproteinase Secretion by Gastric Epithelial Cells Is Regulated by E Prostaglandins and MAPKs. J. Biol. Chem. 2005, 280, 9973–9979. [Google Scholar] [CrossRef]
- Ancha, H.R.; Kurella, R.R.; Stewart, C.A.; Damera, G.; Ceresa, B.P.; Harty, R.F. Histamine Stimulation of MMP-1(Collagenase-1) Secretion and Gene Expression in Gastric Epithelial Cells: Role of EGFR Transactivation and the MAP Kinase Pathway. Int. J. Biochem. Cell Biol. 2007, 39, 2143–2152. [Google Scholar] [CrossRef]
- Kim, S.; Choi, M.G.; Lee, H.S.; Lee, S.K.; Kim, S.; Kim, W.W.; Hur, S.M.; Kim, J.-H.; Choe, J.-H.; Nam, S.J.; et al. Silibinin Suppresses TNF-α-Induced MMP-9 Expression in Gastric Cancer Cells through Inhibition of the MAPK Pathway. Molecules 2009, 14, 4300–4311. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Wang, C.-J.; Tseng, C.-C.; Chen, H.-P.; Wu, M.-S.; Lin, J.-T.; Inoue, H.; Chen, G.-H. Helicobacter pylori promote Gastric Cancer Cells Invasion through a NF-kB and COX-2-Mediated Pathway. World J. Gastroenterol. 2005, 11, 3197–3203. [Google Scholar] [CrossRef]
- Ye, T.; Yang, M.; Huang, D.; Wang, X.; Xue, B.; Tian, N.; Xu, X.; Bao, L.; Hu, H.; Lv, T.; et al. MicroRNA-7 as a Potential Therapeutic Target for Aberrant NF-κB-Driven Distant Metastasis of Gastric Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 55. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-C.; Zhang, K.-L.; Chen, X.-Y.; Jiang, H.-F.; Kong, Q.-Y.; Sun, Y.; Wu, M.-L.; Huang, L.; Li, H.; Liu, J. Expression of Seven Gastric Cancer-Associated Genes and its Relevance for Wnt, NF-κB and Stat3 Signaling. APMIS 2007, 115, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-C.; Cheng, H.-H.; Yeh, T.-S.; Li, Y.-C.; Chen, T.-J.; Sit, W.Y.; Chuu, C.-P.; Kung, H.-J.; Chien, S.; Wang, W.-C. KDM4B Is a Coactivator of C-Jun and Involved in Gastric Carcinogenesis. Cell Death Dis. 2019, 10, 68. [Google Scholar] [CrossRef]
- Shang, W.; Wang, Y.; Liang, X.; Li, T.; Shao, W.; Liu, F.; Cui, X.; Wang, Y.; Lv, L.; Chai, L.; et al. SETDB1 Promotes Gastric Carcinogenesis and Metastasis via Upregulation of CCND1 and MMP9 Expression. J. Pathol. 2020, 253, 148–159. [Google Scholar] [CrossRef]
- Matsumura, S.; Oue, N.; Kitadai, Y.; Chayama, K.; Yoshida, K.; Yamaguchi, Y.; Toge, T.; Imai, K.; Nakachi, K.; Yasui, W. A Single Nucleotide Polymorphism in the MMP-1 Promoter Is Correlated with Histological Differentiation of Gastric Cancer. J. Cancer Res. Clin. Oncol. 2004, 130, 259–265. [Google Scholar] [CrossRef]
- Hellmig, S.; Ott, S.; Rosenstiel, P.; Fölsch, U.R.; Hampe, J.; Schreiber, S. Genetic Variants in Matrix Metalloproteinase Genes Are Associated with Development of Gastric Ulcer in H. pylori Infection. Am. J. Gastroenterol. 2006, 101, 29–35. [Google Scholar] [CrossRef]
- Kubben, F.J.G.M.; Sier, C.F.M.; Meijer, M.J.W.; Berg, M.V.D.; van der Reijden, J.J.; Griffioen, G.; van de Velde, C.J.H.; Lamers, C.B.H.W.; Verspaget, H.W. Clinical Impact of MMP and TIMP Gene Polymorphisms in Gastric Cancer. Br. J. Cancer 2006, 95, 744–751. [Google Scholar] [CrossRef]
- Sugimoto, M.; Furuta, T.; Kodaira, C.; Nishino, M.; Yamade, M.; Ikuma, M.; Sugimura, H.; Hishida, A. Polymorphisms of Matrix Metalloproteinase-7 and Chymase Are Associated with Susceptibility to and Progression of Gastric Cancer in Japan. J. Gastroenterol. 2008, 43, 751–761. [Google Scholar] [CrossRef]
- Achyut, B.; Ghoshal, U.C.; Moorchung, N.; Mittal, B. Transforming Growth Factor-B1 and Matrix Metalloproteinase-7 Promoter Variants Induce Risk for Helicobacter pylori–Associated Gastric Precancerous Lesions. DNA Cell Biol. 2009, 28, 295–301. [Google Scholar] [CrossRef]
- Matsumura, S.; Oue, N.; Nakayama, H.; Kitadai, Y.; Yoshida, K.; Yamaguchi, Y.; Imai, K.; Nakachi, K.; Matsusaki, K.; Chayama, K.; et al. A Single Nucleotide Polymorphism in the MMP-9 Promoter Affects Tumor Progression and Invasive Phenotype of Gastric Cancer. J. Cancer Res. Clin. Oncol. 2004, 131, 19–25. [Google Scholar] [CrossRef]
- Verma, S.; Kesh, K.; Gupta, A.; Swarnakar, S. An Overview of Matrix Metalloproteinase 9 Polymorphism and Gastric Cancer Risk. Asian Pac. J. Cancer Prev. 2015, 16, 7393–7400. [Google Scholar] [CrossRef]
- Carneiro, P.; Fernandes, S.; Figueiredo, J.; Caldeira, J.; Carvalho, J.; Pinheiro, H.; Leite, M.; Melo, S.; Oliveira, P.; Correia, J.S.; et al. E-Cadherin Dysfunction in Gastric Cancer-Cellular Consequences, Clinical Applications and Open Questions. FEBS Lett. 2012, 586, 2981–2989. [Google Scholar] [CrossRef]
- Liu, X.; Chu, K.-M. E-Cadherin and Gastric Cancer: Cause, Consequence, and Applications. BioMed Res. Int. 2014, 2014, 637308. [Google Scholar] [CrossRef]
- Tutunchi, S.; Akhavan, S.; Bereimipour, A.; Ghaderian, S.M.H. Evaluation of Important Molecular Pathways and Candidate Diagnostic Biomarkers of Noninvasive to Invasive Stages in Gastric Cancer by In Silico Analysis. J. Oncol. 2021, 2021, 5571413. [Google Scholar] [CrossRef]
- Gómez-Escudero, J.; Moreno, V.; Martín-Alonso, M.; de Riquer, M.V.H.; Feinberg, T.; Colmenar, Á.; Calvo, E.; Camafeita, E.; Martínez, F.; Oudhoff, M.; et al. E-Cadherin Cleavage by MT2-MMP Regulates Apical Junctional Signaling and Epithelial Homeostasis in the Intestine. J. Cell Sci. 2017, 130, 4013–4027. [Google Scholar] [CrossRef]
- Yao, Z.; Yuan, T.; Wang, H.; Yao, S.; Zhao, Y.; Liu, Y.; Jin, S.; Chu, J.; Xu, Y.; Zhou, W.; et al. MMP-2 Together with MMP-9 Overexpression Correlated with Lymph Node Metastasis and Poor Prognosis in Early Gastric Carcinoma. Tumor Biol. 2017, 39, 1010428317700411. [Google Scholar] [CrossRef]
- Herrera-Pariente, C.; Montori, S.; Llach, J.; Bofill, A.; Albeniz, E.; Moreira, L. Biomarkers for Gastric Cancer Screening and Early Diagnosis. Biomedicines 2021, 9, 1448. [Google Scholar] [CrossRef]
- Shiota, S.; Yamaoka, Y. Biomarkers for Helicobacter pylori Infection and Gastroduodenal Diseases. Biomark. Med. 2014, 8, 1127–1137. [Google Scholar] [CrossRef]
- Kucera, R.; Smid, D.; Topolcan, O.; Karlikova, M.; Fiala, O.; Slouka, D.; Skalicky, T.; Treska, V.; Kulda, V.; Simanek, V.; et al. Searching for New Biomarkers and the Use of Multivariate Analysis in Gastric Cancer Diagnostics. Anticancer Res. 2016, 36, 1967–1971. [Google Scholar]
- Myochin, T.; Hanaoka, K.; Komatsu, T.; Terai, T.; Nagano, T. Design Strategy for a Near-Infrared Fluorescence Probe for Matrix Metalloproteinase Utilizing Highly Cell Permeable Boron Dipyrromethene. J. Am. Chem. Soc. 2012, 134, 13730–13737. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Wu, L.; Sun, Y.; Wang, Y.; Wang, J.; Ye, D. Ratiometric Imaging of MMP-2 Activity Facilitates Tumor Detection Using Activatable Near-Infrared Fluorescent Semiconducting Polymer Nanoparticles. Small 2021, 17, 2101924. [Google Scholar] [CrossRef] [PubMed]
- Eder, M.; Pavan, S.; Bauder-Wüst, U.; Van Rietschoten, K.; Baranski, A.-C.; Harrison, H.; Campbell, S.; Stace, C.L.; Walker, E.H.; Chen, L.; et al. Bicyclic Peptides as a New Modality for Imaging and Targeting of Proteins Overexpressed by Tumors. Cancer Res. 2019, 79, 841–852. [Google Scholar] [CrossRef]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix Metalloproteinases: Biologic Activity and Clinical Implications. J. Clin. Oncol. 2000, 18, 1135. [Google Scholar] [CrossRef]
- Fields, G.B. The Rebirth of Matrix Metalloproteinase Inhibitors: Moving Beyond the Dogma. Cells 2019, 8, 984. [Google Scholar] [CrossRef]
- Waller, V.; Pruschy, M. Combined Radiochemotherapy: Metalloproteinases Revisited. Front. Oncol. 2021, 11, 676583. [Google Scholar] [CrossRef]
- Love, E.A.; Sattikar, A.; Cook, H.; Gillen, K.; Large, J.M.; Patel, S.; Matthews, D.; Merritt, A. Developing an Antibody–Drug Conjugate Approach to Selective Inhibition of an Extracellular Protein. ChemBioChem 2018, 20, 754–758. [Google Scholar] [CrossRef]
- Devy, L.; Huang, L.; Naa, L.; Yanamandra, N.; Pieters, H.; Frans, N.; Chang, E.; Tao, Q.; Vanhove, M.; Lejeune, A.; et al. Selective Inhibition of Matrix Metalloproteinase-14 Blocks Tumor Growth, Invasion, and Angiogenesis. Cancer Res. 2009, 69, 1517–1526. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolova, O.; Naumann, M. Matrix Metalloproteinases in Helicobacter pylori–Associated Gastritis and Gastric Cancer. Int. J. Mol. Sci. 2022, 23, 1883. https://doi.org/10.3390/ijms23031883
Sokolova O, Naumann M. Matrix Metalloproteinases in Helicobacter pylori–Associated Gastritis and Gastric Cancer. International Journal of Molecular Sciences. 2022; 23(3):1883. https://doi.org/10.3390/ijms23031883
Chicago/Turabian StyleSokolova, Olga, and Michael Naumann. 2022. "Matrix Metalloproteinases in Helicobacter pylori–Associated Gastritis and Gastric Cancer" International Journal of Molecular Sciences 23, no. 3: 1883. https://doi.org/10.3390/ijms23031883
APA StyleSokolova, O., & Naumann, M. (2022). Matrix Metalloproteinases in Helicobacter pylori–Associated Gastritis and Gastric Cancer. International Journal of Molecular Sciences, 23(3), 1883. https://doi.org/10.3390/ijms23031883