
Analysis of mRNA and Protein Levels of CAP2, DLG1 and ADAM10 Genes in Post-Mortem Brain of Schizophrenia, Parkinson’s and Alzheimer’s Disease Patients

 , ,
, , 

Abstract
:1. Introduction
2. Results
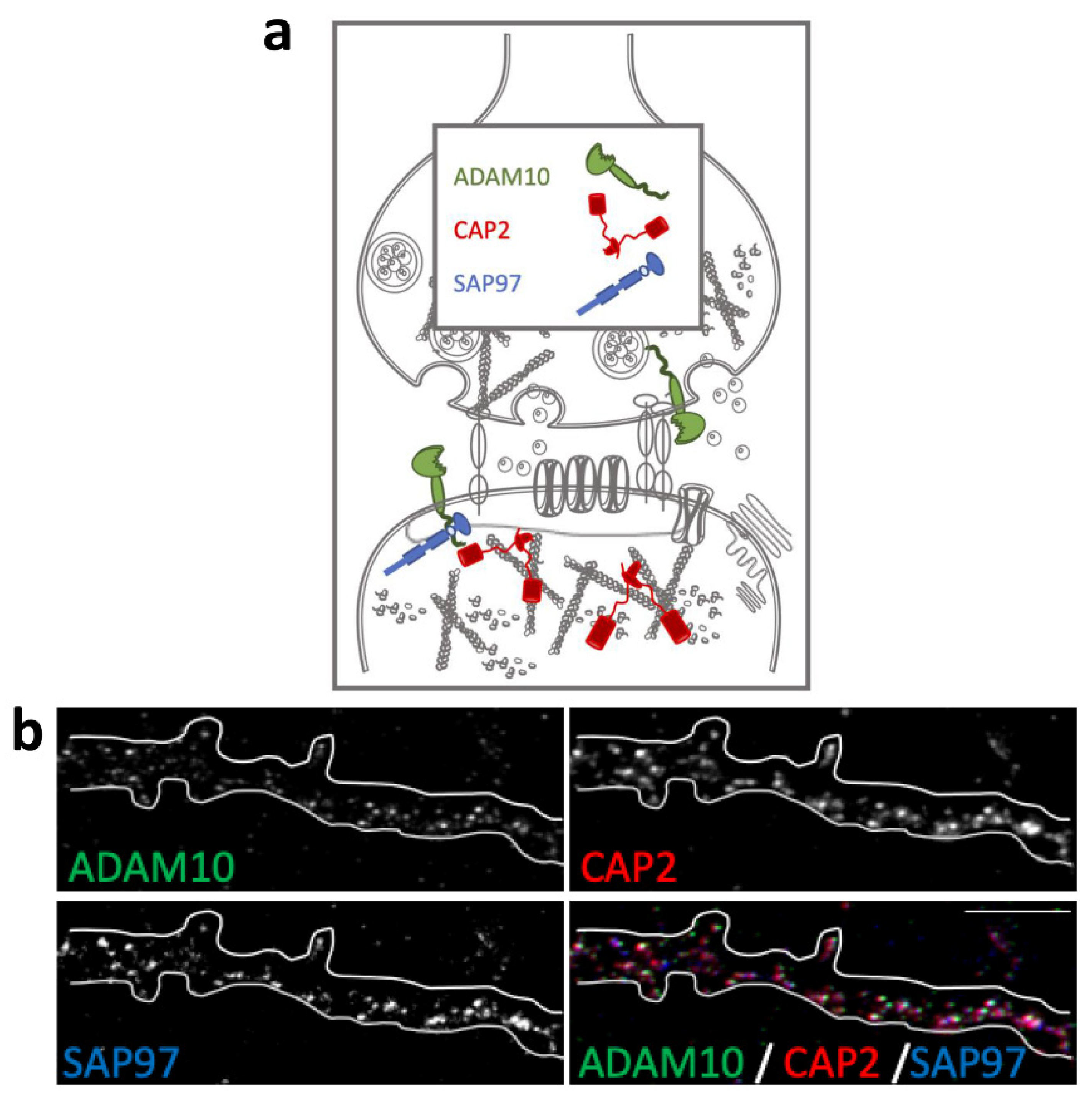
2.1. Colocalization of CAP2, SAP97, and ADAM10 in Primary Hippocampal Neurons
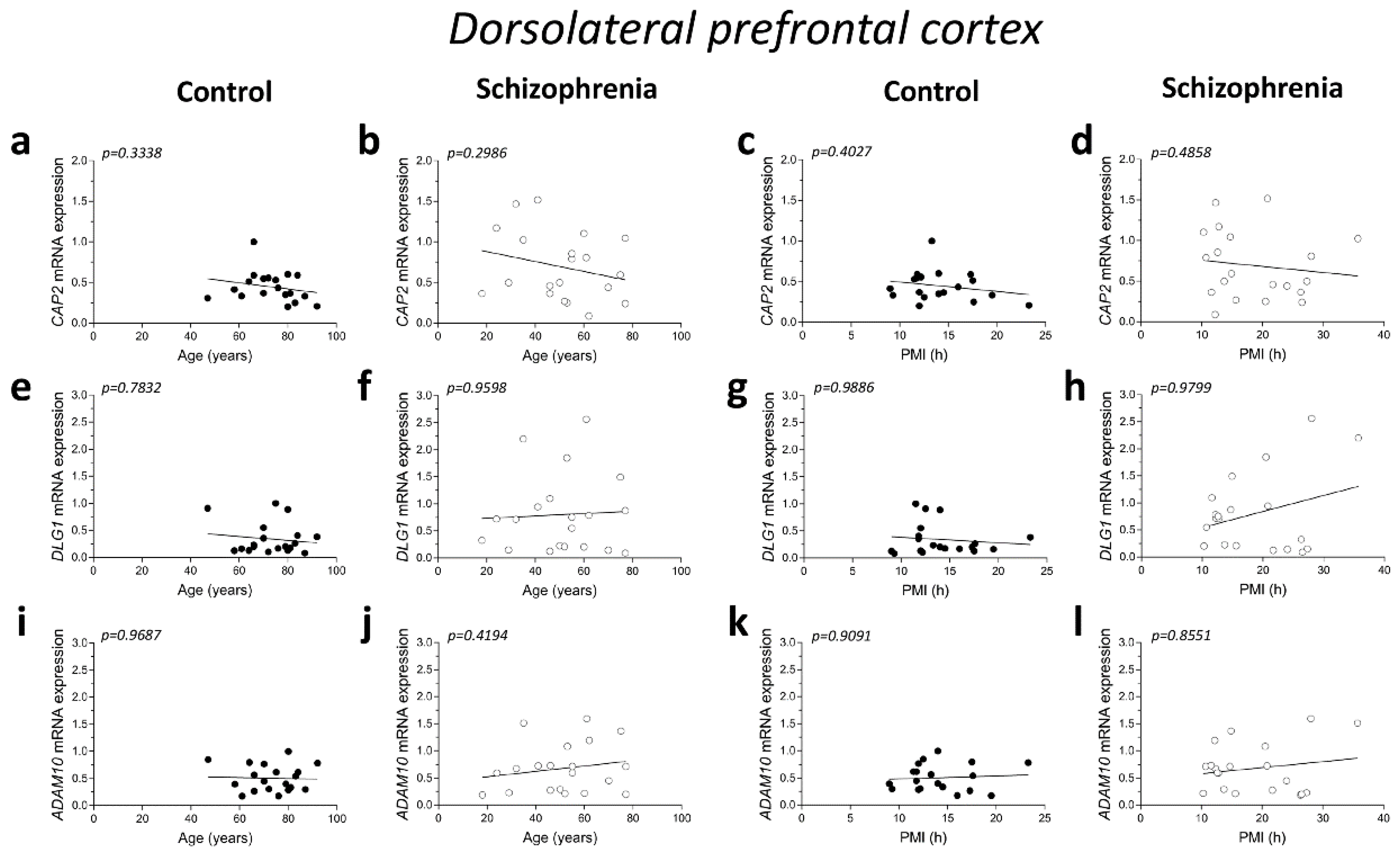
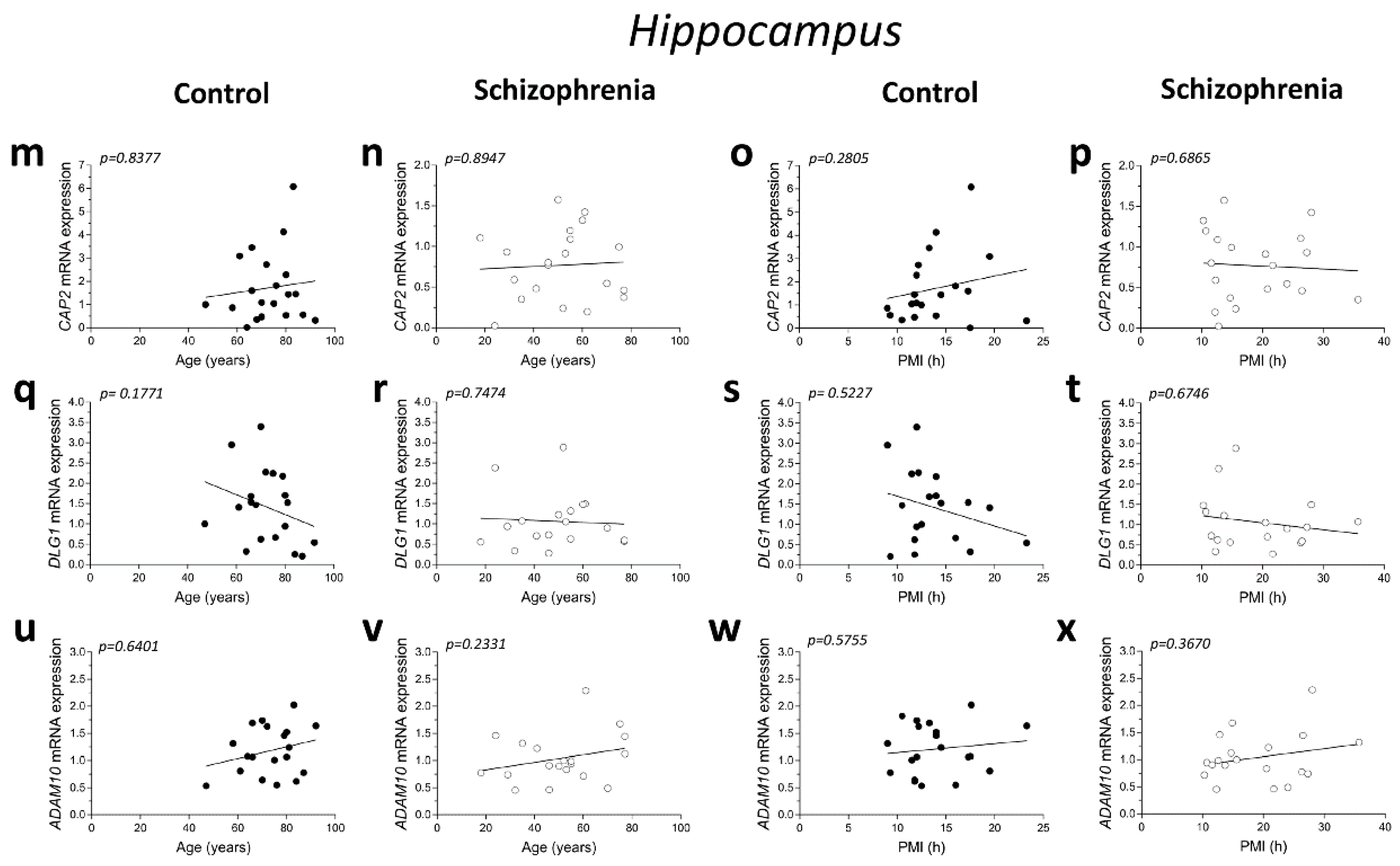
2.2. Correlation Analysis of CAP2, DLG1, and ADAM10 mRNA Expression with Age and PMI in the Post-Mortem Dorsolateral Prefrontal Cortex and Hippocampus of Schizophrenia Patients
2.3. Analysis of mRNA Expression of CAP2, DLG1, and ADAM10 in the Post-Mortem Dorsolateral Prefrontal Cortex and Hippocampus of SCZ Patients
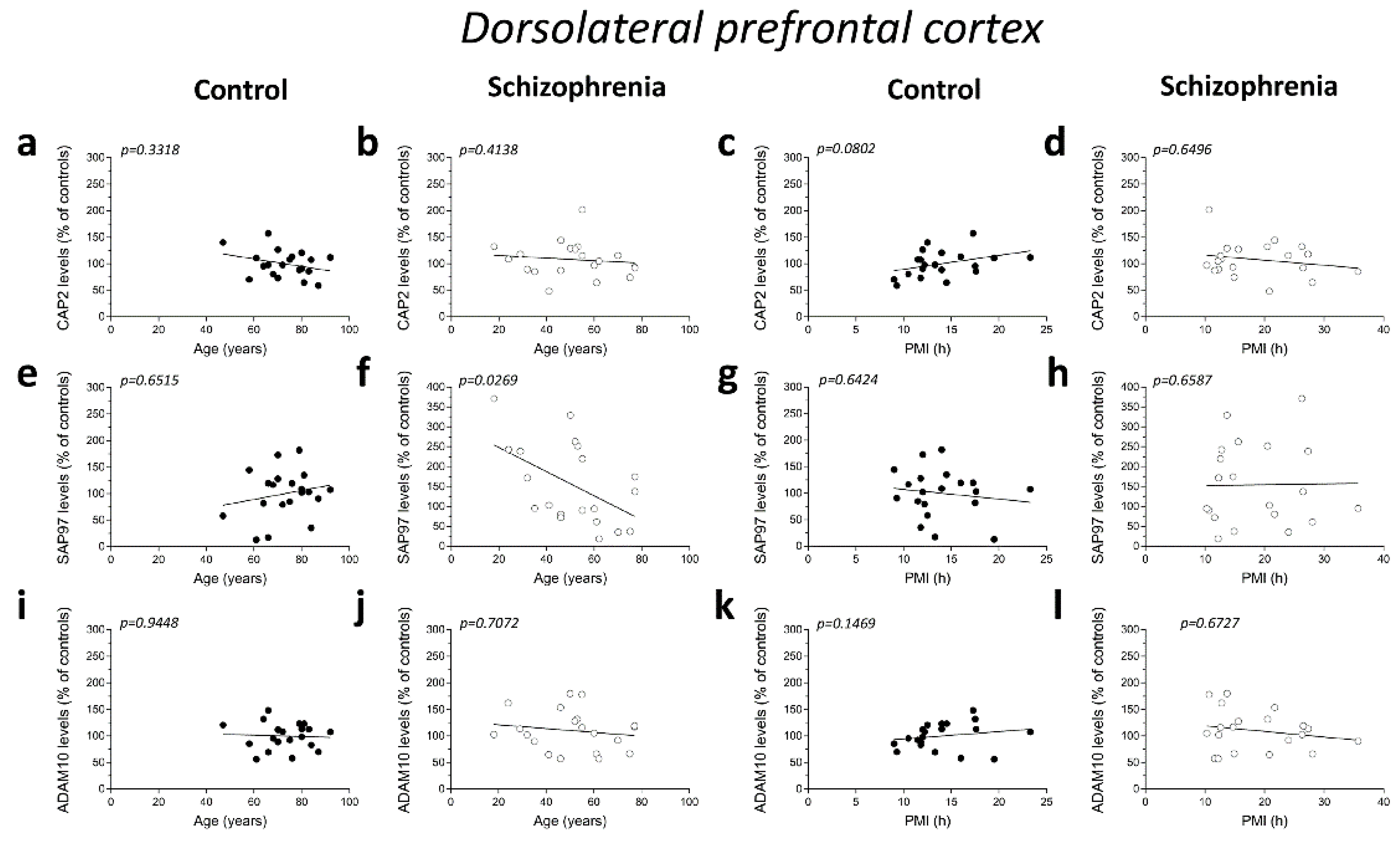
2.4. Correlation Analysis of CAP2, SAP97, and ADAM10 Protein Levels with Age and PMI in the Post-Mortem Dorsolateral Prefrontal Cortex and Hippocampus of Schizophrenia Patients
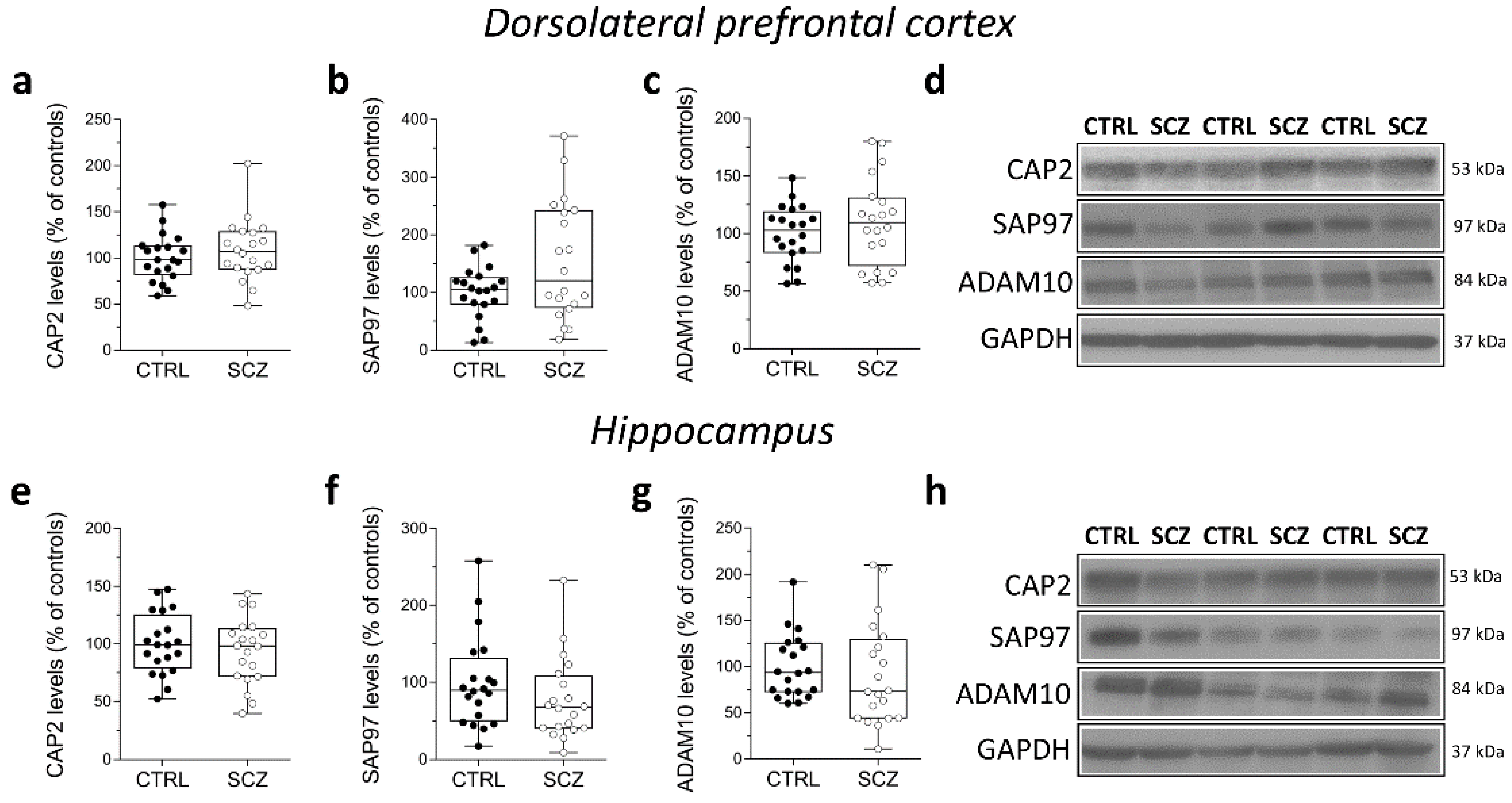
2.5. Analysis of Protein Expression of CAP2, SAP97, and ADAM10 in the Post-Mortem Dorsolateral Prefrontal Cortex and Hippocampus of SCZ Patients
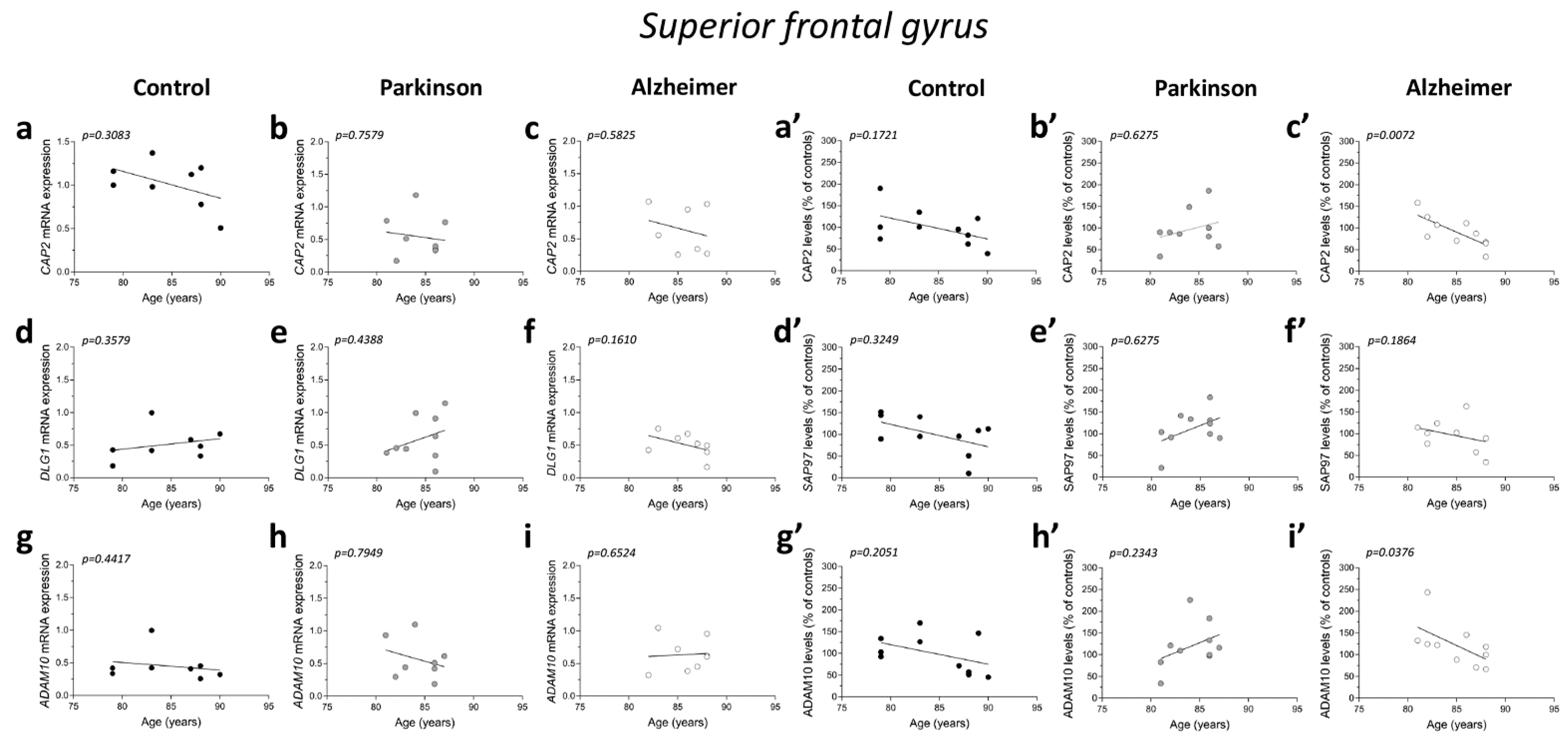
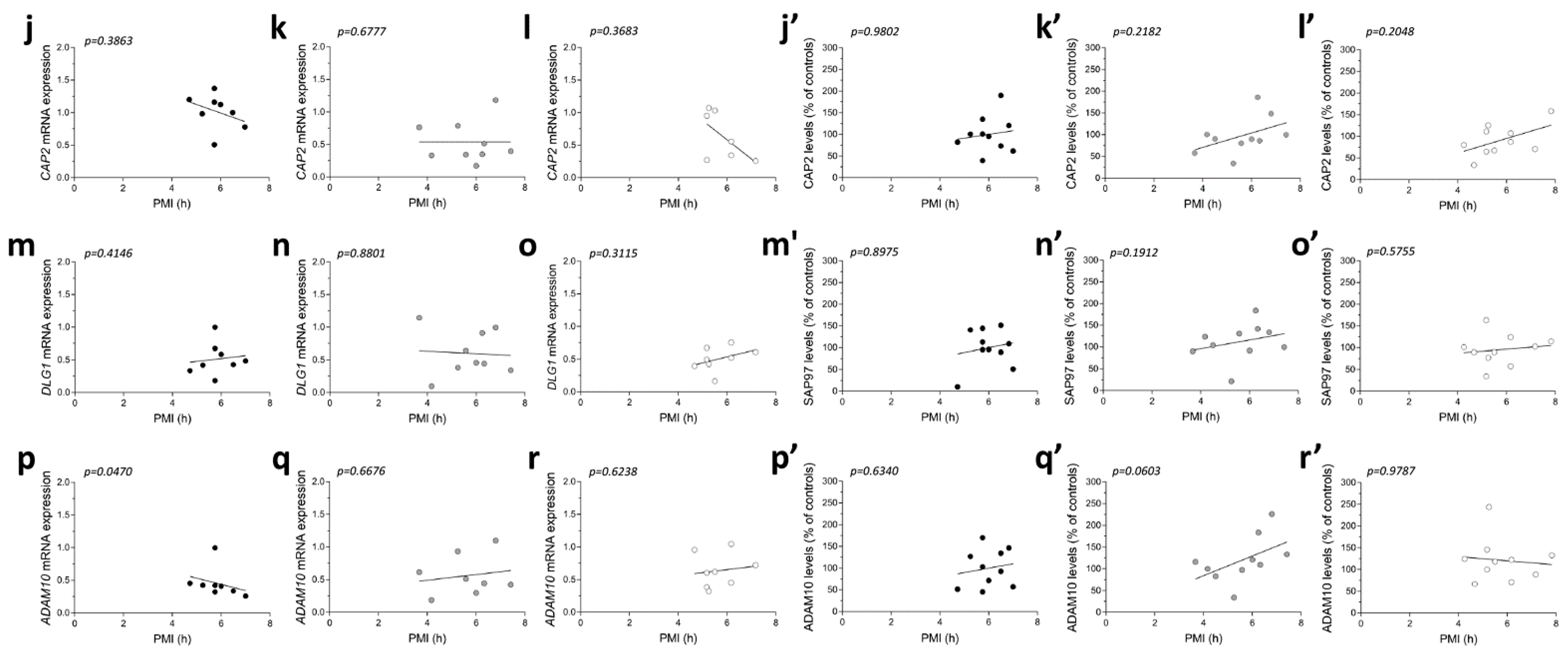
2.6. Correlation Analysis of CAP2, SAP97, and ADAM10 mRNA and Protein Expression with Age and PMI in the Post-Mortem Superior Frontal Gyrus of Alzheimer’s and Parkinson’s Disease Patients
2.7. Analysis of CAP2, DLG1, and ADAM10 Transcript Levels in the Post-Mortem Superior Frontal Gyrus of Alzheimer’s and Parkinson’s Disease Patients
2.8. Analysis of Protein Expression of CAP2, SAP97, and ADAM10 in the Post-Mortem Superior Frontal Gyrus of Alzheimer’s and Parkinson’s Disease Patients
3. Discussion
4. Materials and Methods
4.1. Neuronal Cultures Preparation and Immunocytochemistry
4.2. Human Post-Mortem Tissue Collection
4.3. RNA Extraction and Quantitative RT-PCR Analysis
4.4. Western Blotting
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, T.; Lauriello, J. Schizophrenia: An Overview, Focus. Am. Psychiatr. Publ. 2016, 14, 300–307. [Google Scholar]
- Wang, S.H.; Hsiao, P.C.; Yeh, L.L.; Liu, C.M.; Liu, C.C.; Hwang, T.J.; Hsieh, M.H.; Chien, Y.L.; Lin, Y.T.; Chandler, S.D.; et al. Polygenic risk for schizophrenia and neurocognitive performance in patients with schizophrenia. Genes Brain Behav. 2018, 17, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.A.; Levitt, P. Schizophrenia as a disorder of neurodevelopment. Annu. Rev. Neurosci. 2002, 25, 409–432. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.M.; Bhavsar, V.; Tripoli, G.; Howes, O. 30 Years on: How the Neurodevelopmental Hypothesis of Schizophrenia Morphed into the Developmental Risk Factor Model of Psychosis. Schizophr. Bull. 2017, 43, 1190–1196. [Google Scholar] [CrossRef]
- Rujescu, D. Schizophrenia genes: On the matter of their convergence. Curr. Top. Behav. Neurosci. 2012, 12, 429–440. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.L.; Alhassan, J.; Newman, J.T.; Richard, M.; Gu, H.; Kelly, R.M.; Sampson, A.R.; Fish, K.N.; Penzes, P.; Wills, Z.P.; et al. Selective Loss of Smaller Spines in Schizophrenia. Am. J. Psychiatry 2017, 174, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Kaizuka, T.; Takumi, T. Postsynaptic density proteins and their involvement in neurodevelopmental disorders. J. Biochem. 2018, 163, 447–455. [Google Scholar] [CrossRef]
- Feng, Z.; Wu, X.; Zhang, M. Presynaptic bouton compartmentalization and postsynaptic density-mediated glutamate receptor clustering via phase separation. Neuropharmacology 2021, 193, 108622. [Google Scholar] [CrossRef]
- Sala, C.; Segal, M. Dendritic spines: The locus of structural and functional plasticity. Physiol. Rev. 2014, 94, 141–188. [Google Scholar] [CrossRef]
- Yang, G.; Pan, F.; Gan, W.B. Stably maintained dendritic spines are associated with lifelong memories. Nature 2009, 462, 920–924. [Google Scholar] [CrossRef] [Green Version]
- Bourne, J.; Harris, K.M. Do thin spines learn to be mushroom spines that remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Glausier, J.R.; Lewis, D.A. Dendritic spine pathology in schizophrenia. Neuroscience 2013, 251, 90–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, A.M.; Owens, D.C.; Moorhead, W.J.; Whalley, H.C.; Stanfield, A.C.; Hall, J.; Johnstone, E.C.; Lawrie, S.M. Longitudinal volume reductions in people at high genetic risk of schizophrenia as they develop psychosis. Biol. Psychiatry 2011, 69, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.S.; Pogue-Geile, M.F. Variation in fourteen brain structure volumes in schizophrenia: A comprehensive meta-analysis of 246 studies. Neurosci. Biobehav. Rev. 2019, 98, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J. The hippocampus in schizophrenia: A review of the neuropathological evidence and its pathophysiological implications. Psychopharmacology 2004, 174, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Tamminga, C.A.; Stan, A.D.; Wagner, A.D. The hippocampal formation in schizophrenia. Am. J. Psychiatry 2010, 167, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Bourne, J.N.; Harris, K.M. Balancing structure and function at hippocampal dendritic spines. Annu. Rev. Neurosci. 2008, 31, 47–67. [Google Scholar] [CrossRef] [Green Version]
- Dalva, M.B.; McClelland, A.C.; Kayser, M.S. Cell adhesion molecules: Signalling functions at the synapse. Nat. Rev. Neurosci. 2007, 8, 206–220. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, L.A.; Goda, Y. Actin in action: The interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci. 2008, 9, 344–356. [Google Scholar] [CrossRef]
- Hotulainen, P.; Hoogenraad, C.C. Actin in dendritic spines: Connecting dynamics to function. J. Cell Biol. 2010, 189, 619–629. [Google Scholar] [CrossRef] [Green Version]
- McGeachie, A.B.; Cingolani, L.A.; Goda, Y. Stabilising influence: Integrins in regulation of synaptic plasticity. Neurosci. Res. 2011, 70, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudenko, G. Dynamic Control of Synaptic Adhesion and Organizing Molecules in Synaptic Plasticity. Neural. Plast. 2017, 2017, 6526151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelucchi, S.; Vandermeulen, L.; Pizzamiglio, L.; Aksan, B.; Yan, J.; Konietzny, A.; Bonomi, E.; Borroni, B.; Padovani, A.; Rust, M.B.; et al. Cyclase-associated protein 2 dimerization regulates cofilin in synaptic plasticity and Alzheimer’s disease. Brain Commun. 2020, 2, fcaa086. [Google Scholar] [CrossRef] [PubMed]
- Marcello, E.; Gardoni, F.; Mauceri, D.; Romorini, S.; Jeromin, A.; Epis, R.; Borroni, B.; Cattabeni, F.; Sala, C.; Padovani, A.; et al. Synapse-associated protein-97 mediates alpha-secretase ADAM10 trafficking and promotes its activity. J. Neurosci. 2007, 27, 1682–1691. [Google Scholar] [CrossRef] [Green Version]
- Elias, G.M.; Nicoll, R.A. Synaptic trafficking of glutamate receptors by MAGUK scaffolding proteins. Trends Cell Biol. 2007, 17, 343–352. [Google Scholar] [CrossRef]
- Marcello, E.; Saraceno, C.; Musardo, S.; Vara, H.; de la Fuente, A.G.; Pelucchi, S.; Di Marino, D.; Borroni, B.; Tramontano, A.; Perez-Otano, I.; et al. Endocytosis of synaptic ADAM10 in neuronal plasticity and Alzheimer’s disease. J. Clin. Investig. 2013, 123, 2523–2538. [Google Scholar] [CrossRef]
- Lepeta, K.; Lourenco, M.V.; Schweitzer, B.C.; Martino Adami, P.V.; Banerjee, P.; Catuara-Solarz, S.; de La Fuente Revenga, M.; Guillem, A.M.; Haidar, M.; Ijomone, O.M.; et al. Synaptopathies: Synaptic dysfunction in neurological disorders—A review from students to students. J. Neurochem. 2016, 138, 785–805. [Google Scholar] [CrossRef]
- Taoufik, E.; Kouroupi, G.; Zygogianni, O.; Matsas, R. Synaptic dysfunction in neurodegenerative and neurodevelopmental diseases: An overview of induced pluripotent stem-cell-based disease models. Open Biol. 2018, 8, 180138. [Google Scholar] [CrossRef] [Green Version]
- Penzes, P.; Cahill, M.E.; Jones, K.A.; VanLeeuwen, J.E.; Woolfrey, K.M. Dendritic spine pathology in neuropsychiatric disorders. Nat. Neurosci. 2011, 14, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Shirao, T.; Gonzalez-Billault, C. Actin filaments and microtubules in dendritic spines. J. Neurochem. 2013, 126, 155–164. [Google Scholar] [CrossRef]
- Begre, S.; Koenig, T. Cerebral disconnectivity: An early event in schizophrenia. Neuroscientist 2008, 14, 19–45. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Lindenberg, A. From maps to mechanisms through neuroimaging of schizophrenia. Nature 2010, 468, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ursini, G.; Romer, A.L.; Knodt, A.R.; Mezeivtch, K.; Xiao, E.; Pergola, G.; Blasi, G.; Straub, R.E.; Callicott, J.H.; et al. Schizophrenia polygenic risk score predicts mnemonic hippocampal activity. Brain 2018, 141, 1218–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampino, A.; Di Carlo, P.; Fazio, L.; Ursini, G.; Pergola, G.; De Virgilio, C.; Gadaleta, G.; Giordano, G.M.; Bertolino, A.; Blasi, G. Association of functional genetic variation in PP2A with prefrontal working memory processing. Behav. Brain Res. 2017, 316, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zeppillo, T.; Schulmann, A.; Macciardi, F.; Hjelm, B.E.; Focking, M.; Sequeira, P.A.; Guella, I.; Cotter, D.; Bunney, W.E.; Limon, A.; et al. Functional impairment of cortical AMPA receptors in schizophrenia. Schizophr. Res. 2020, in press. [CrossRef] [PubMed]
- Uezato, A.; Kimura-Sato, J.; Yamamoto, N.; Iijima, Y.; Kunugi, H.; Nishikawa, T. Further evidence for a male-selective genetic association of synapse-associated protein 97 (SAP97) gene with schizophrenia. Behav. Brain Funct. 2012, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uezato, A.; Yamamoto, N.; Jitoku, D.; Haramo, E.; Hiraaki, E.; Iwayama, Y.; Toyota, T.; Umino, M.; Umino, A.; Iwata, Y.; et al. Genetic and molecular risk factors within the newly identified primate-specific exon of the SAP97/DLG1 gene in the 3q29 schizophrenia-associated locus. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2017, 174, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liang, C.; Lv, D.; Yin, J.; Luo, X.; Fu, J.; Yan, H.; Zhou, X.; Dai, Z.; Zhu, D.; et al. Association of the Synapse-Associated Protein 97 (SAP97) Gene Polymorphism with Neurocognitive Function in Schizophrenic Patients. Front. Psychiatry 2018, 9, 458. [Google Scholar] [CrossRef]
- Xu, X.; Wang, Y.; Zhou, X.; Yin, J.; Yu, H.; Wen, X.; Lv, D.; Zhu, D.; Xiong, S.; Yan, H.; et al. The genetic variations in SAP97 gene and the risk of schizophrenia in the Chinese Han population: A further study. Psychiatr. Genet. 2020, 30, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; He, B.; Lin, Z.; Wang, X.; Yin, J.; Luo, X.; Luo, S.; Liang, C.; Wen, X.; Xiong, S.; et al. SAP97 rs3915512 Polymorphism Affects the Neurocognition of Schizophrenic Patients: A Genetic Neuroimaging Study. Front. Genet. 2020, 11, 572414. [Google Scholar] [CrossRef]
- Xu, X.; Luo, S.; Wen, X.; Wang, X.; Yin, J.; Luo, X.; He, B.; Liang, C.; Xiong, S.; Zhu, D.; et al. Genetic Contribution of Synapse-Associated Protein 97 to Orbitofrontal-Striatal-Thalamic Circuitry Connectivity Changes in First-Episode Schizophrenia. Front. Psychiatry 2021, 12, 691007. [Google Scholar] [CrossRef] [PubMed]
- Toyooka, K.; Iritani, S.; Makifuchi, T.; Shirakawa, O.; Kitamura, N.; Maeda, K.; Nakamura, R.; Niizato, K.; Watanabe, M.; Kakita, A.; et al. Selective reduction of a PDZ protein, SAP-97, in the prefrontal cortex of patients with chronic schizophrenia. J. Neurochem. 2002, 83, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.C.; McCullumsmith, R.E.; Funk, A.J.; Haroutunian, V.; Meador-Woodruff, J.H. Evidence for abnormal forward trafficking of AMPA receptors in frontal cortex of elderly patients with schizophrenia. Neuropsychopharmacology 2010, 35, 2110–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dracheva, S.; McGurk, S.R.; Haroutunian, V. mRNA expression of AMPA receptors and AMPA receptor binding proteins in the cerebral cortex of elderly schizophrenics. J. Neurosci. Res. 2005, 79, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.F.; Mirza, G.; Sekhon, G.; Turnpenny, P.; Leroy, F.; Speleman, F.; Law, C.; van Regemorter, N.; Vamos, E.; Flinter, F.; et al. Delineation of two distinct 6p deletion syndromes. Hum. Genet. 1999, 104, 64–72. [Google Scholar] [CrossRef]
- Bremer, A.; Schoumans, J.; Nordenskjold, M.; Anderlid, B.M.; Giacobini, M. An interstitial deletion of 7.1 Mb in chromosome band 6p22.3 associated with developmental delay and dysmorphic features including heart defects, short neck, and eye abnormalities. Eur. J. Med. Genet. 2009, 52, 358–362. [Google Scholar] [CrossRef]
- Martins-de-Souza, D.; Maccarrone, G.; Wobrock, T.; Zerr, I.; Gormanns, P.; Reckow, S.; Falkai, P.; Schmitt, A.; Turck, C.W. Proteome analysis of the thalamus and cerebrospinal fluid reveals glycolysis dysfunction and potential biomarkers candidates for schizophrenia. J. Psychiatr. Res. 2010, 44, 1176–1189. [Google Scholar] [CrossRef]
- Wong, A.H.; Likhodi, O.; Trakalo, J.; Yusuf, M.; Sinha, A.; Pato, C.N.; Pato, M.T.; Van Tol, H.H.; Kennedy, J.L. Genetic and post-mortem mRNA analysis of the 14-3-3 genes that encode phosphoserine/threonine-binding regulatory proteins in schizophrenia and bipolar disorder. Schizophr. Res. 2005, 78, 137–146. [Google Scholar] [CrossRef]
- Hill, S.L.; Shao, L.; Beasley, C.L. Diminished levels of the chemokine fractalkine in post-mortem prefrontal cortex in schizophrenia but not bipolar disorder. World J. Biol. Psychiatry 2021, 22, 94–103. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Katsel, P.; Haroutunian, V.; Chelini, G.; Klengel, T.; Berretta, S. Molecular signature of extracellular matrix pathology in schizophrenia. Eur. J. Neurosci. 2021, 53, 3960–3987. [Google Scholar] [CrossRef]
- Tomasetti, C.; Dell’Aversano, C.; Iasevoli, F.; de Bartolomeis, A. Homer splice variants modulation within cortico-subcortical regions by dopamine D2 antagonists, a partial agonist, and an indirect agonist: Implication for glutamatergic postsynaptic density in antipsychotics action. Neuroscience 2007, 150, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, C.; Dell’Aversano, C.; Iasevoli, F.; Marmo, F.; de Bartolomeis, A. The acute and chronic effects of combined antipsychotic-mood stabilizing treatment on the expression of cortical and striatal postsynaptic density genes. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 184–197. [Google Scholar] [CrossRef] [PubMed]
- De Bartolomeis, A.; Prinzivalli, E.; Callovini, G.; D’Ambrosio, L.; Altavilla, B.; Avagliano, C.; Iasevoli, F. Treatment resistant schizophrenia and neurological soft signs may converge on the same pathology: Evidence from explanatory analysis on clinical, psychopathological, and cognitive variables. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 81, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Laursen, T.M. Causes of premature mortality in schizophrenia: A review of literature published in 2018. Curr. Opin. Psychiatry 2019, 32, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Olfson, M.; Gerhard, T.; Huang, C.; Crystal, S.; Stroup, T.S. Premature Mortality among Adults with Schizophrenia in the United States. JAMA Psychiatry 2015, 72, 1172–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcello, E.; Epis, R.; Saraceno, C.; Gardoni, F.; Borroni, B.; Cattabeni, F.; Padovani, A.; Di Luca, M. SAP97-mediated local trafficking is altered in Alzheimer disease patients’ hippocampus. Neurobiol. Aging 2012, 33, 422.e1–422.e10. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Xiong, S.; Zhou, X.; Chen, X.; Wang, X.; Liang, C.; Yin, J.; Luo, X.; Fu, J.; Wen, X.; et al. SAP97 polymorphisms associated with early onset Parkinson’s disease. Neurosci. Lett. 2020, 728, 134931. [Google Scholar] [CrossRef]
- Zhou, M.; Lin, Y.; Lu, L.; Zhang, Z.; Guo, W.; Peng, G.; Zhang, W.; Zhu, Z.; Wu, Z.; Mo, M.; et al. Association of ADAM10 gene variants with sporadic Parkinson’s disease in Chinese Han population. J. Gene Med. 2021, 23, e3319. [Google Scholar] [CrossRef]
- Fourie, C.; Kim, E.; Waldvogel, H.; Wong, J.M.; McGregor, A.; Faull, R.L.; Montgomery, J.M. Differential Changes in Postsynaptic Density Proteins in Postmortem Huntington’s Disease and Parkinson’s Disease Human Brains. J. Neurodegener. Dis. 2014, 2014, 938530. [Google Scholar] [CrossRef] [Green Version]
- Nash, J.E.; Johnston, T.H.; Collingridge, G.L.; Garner, C.C.; Brotchie, J.M. Subcellular redistribution of the synapse-associated proteins PSD-95 and SAP97 in animal models of Parkinson’s disease and L-DOPA-induced dyskinesia. FASEB J. 2005, 19, 583–585. [Google Scholar] [CrossRef]
- Piccoli, G.; Verpelli, C.; Tonna, N.; Romorini, S.; Alessio, M.; Nairn, A.C.; Bachi, A.; Sala, C. Proteomic analysis of activity-dependent synaptic plasticity in hippocampal neurons. J. Proteome Res. 2007, 6, 3203–3215. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braak, H.; Braak, E. Staging of Alzheimer’s disease-related neurofibrillary changes. Neurobiol. Aging 1995, 16, 271–278; discussion 278–284. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control | Schizophrenia | p-Value |
|---|---|---|---|
| Subjects (total number) | 20 | 20 | -- |
| Gender (M/F) | 16/4 | 12/8 | 0.301 a |
| Age (years, median (IQR)) | 73.50 (66.00–80.25) | 52.50 (39.50–61.25) | <0.001 b |
| PMI (hours, median (IQR)) | 12.90 (11.80–16.32) | 15.25 (12.52–24.58) | 0.020 c |
| pH (median, (IQR)) | 6.54 (6.49–6.63) | 6.50 (6.42–6.56) | 0.485 c |
| Characteristics | Control | Alzheimer’s Disease | Parkinson’s Disease | |||
|---|---|---|---|---|---|---|
| N | Median | N | Median | N | Median | |
| Subjects (total number) | 10 | 10 | 10 | |||
| Gender (M/F) | 10/0 | 10/0 | 10/0 | |||
| Age (years, median (IQR)) | 85.00 (79.00–88.25) | 85.50 (82.00–88.00) | 85.00 (81.75–86.00) | |||
| PMI (hours, median (IQR)) | 5.87 (5.62–6.58) | 5.37 (5.04–6.42) | 5.79 (4.42–6.45) | |||
| Amyloid (A/B/C/O) | 6/2/0/2 | 0/1/9/0 | 3/3/0/4 | |||
| Braak (0/I/II/III/IV/V/VI) | 0/0/3/5/2/0/0 | 0/0/0/0/0/7/3 | 0/0/3/4/0/0/0 | |||
| Braak LB (0/I/II/III/IV/V/VI) | 2/2/0/0/0/0/0 | 0/0/0/0/0/0/0 | 0/0/0/0/2/2/6 | |||
| Liquor pH (median (IQR)) | 6.37 (6.29–6.72) | 6.28 (6.13–6.49) | 6.33 (6.16–6.72) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Maio, A.; De Rosa, A.; Pelucchi, S.; Garofalo, M.; Marciano, B.; Nuzzo, T.; Gardoni, F.; Isidori, A.M.; Di Luca, M.; Errico, F.; et al. Analysis of mRNA and Protein Levels of CAP2, DLG1 and ADAM10 Genes in Post-Mortem Brain of Schizophrenia, Parkinson’s and Alzheimer’s Disease Patients. Int. J. Mol. Sci. 2022, 23, 1539. https://doi.org/10.3390/ijms23031539
Di Maio A, De Rosa A, Pelucchi S, Garofalo M, Marciano B, Nuzzo T, Gardoni F, Isidori AM, Di Luca M, Errico F, et al. Analysis of mRNA and Protein Levels of CAP2, DLG1 and ADAM10 Genes in Post-Mortem Brain of Schizophrenia, Parkinson’s and Alzheimer’s Disease Patients. International Journal of Molecular Sciences. 2022; 23(3):1539. https://doi.org/10.3390/ijms23031539
Chicago/Turabian StyleDi Maio, Anna, Arianna De Rosa, Silvia Pelucchi, Martina Garofalo, Benedetta Marciano, Tommaso Nuzzo, Fabrizio Gardoni, Andrea M. Isidori, Monica Di Luca, Francesco Errico, and et al. 2022. "Analysis of mRNA and Protein Levels of CAP2, DLG1 and ADAM10 Genes in Post-Mortem Brain of Schizophrenia, Parkinson’s and Alzheimer’s Disease Patients" International Journal of Molecular Sciences 23, no. 3: 1539. https://doi.org/10.3390/ijms23031539
APA StyleDi Maio, A., De Rosa, A., Pelucchi, S., Garofalo, M., Marciano, B., Nuzzo, T., Gardoni, F., Isidori, A. M., Di Luca, M., Errico, F., De Bartolomeis, A., Marcello, E., & Usiello, A. (2022). Analysis of mRNA and Protein Levels of CAP2, DLG1 and ADAM10 Genes in Post-Mortem Brain of Schizophrenia, Parkinson’s and Alzheimer’s Disease Patients. International Journal of Molecular Sciences, 23(3), 1539. https://doi.org/10.3390/ijms23031539