
Cyclophilin A Is Not Acetylated at Lysine-82 and Lysine-125 in Resting and Stimulated Platelets

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
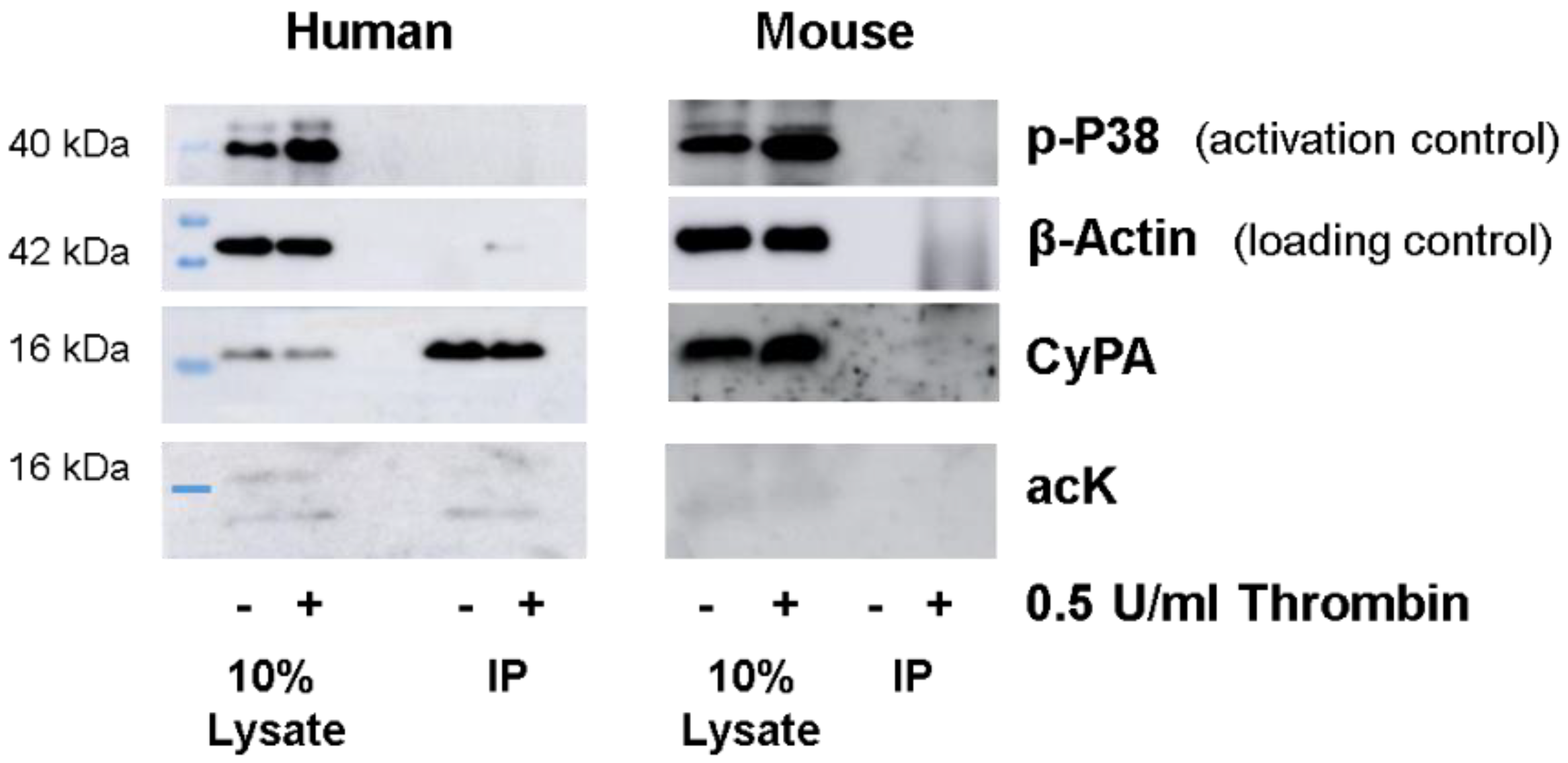
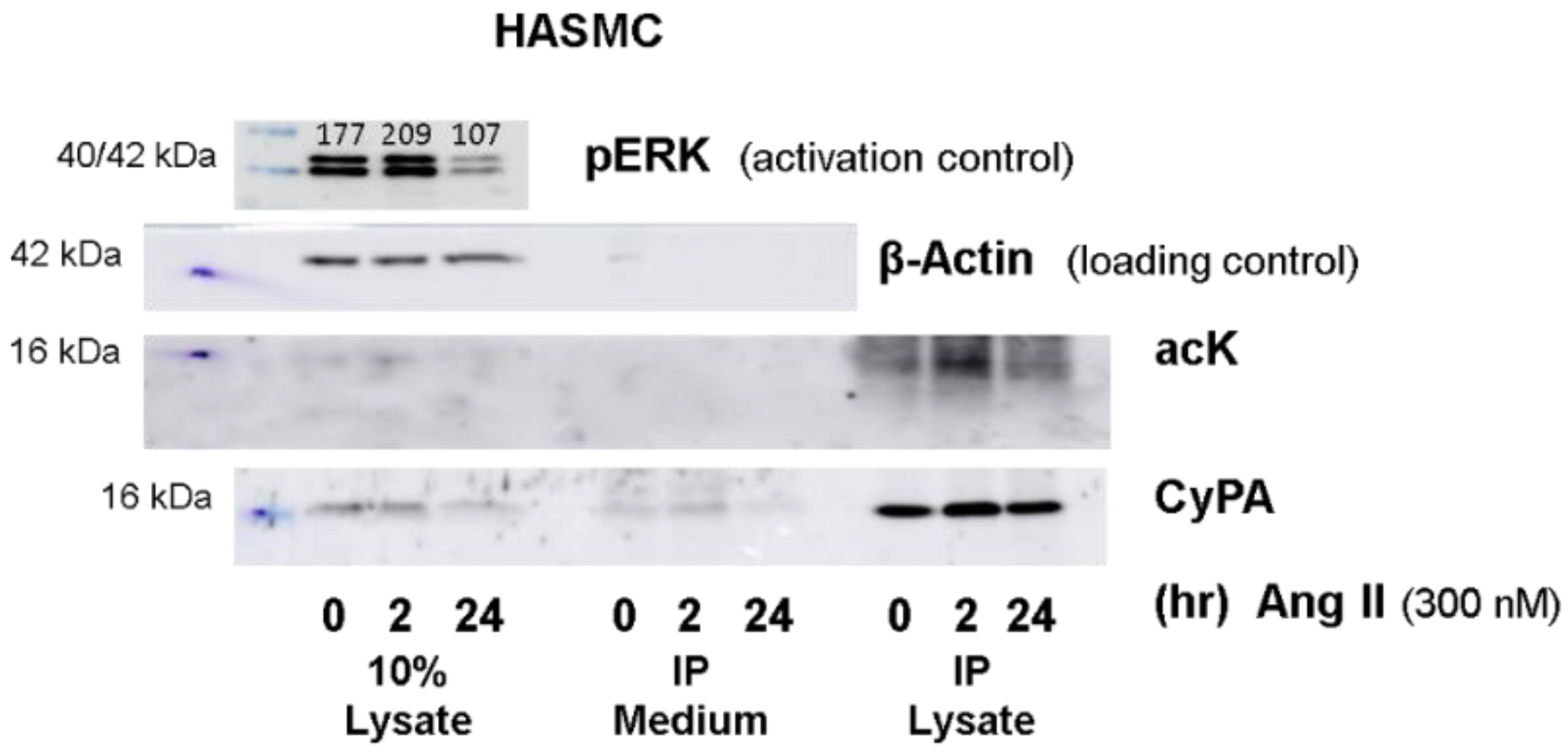
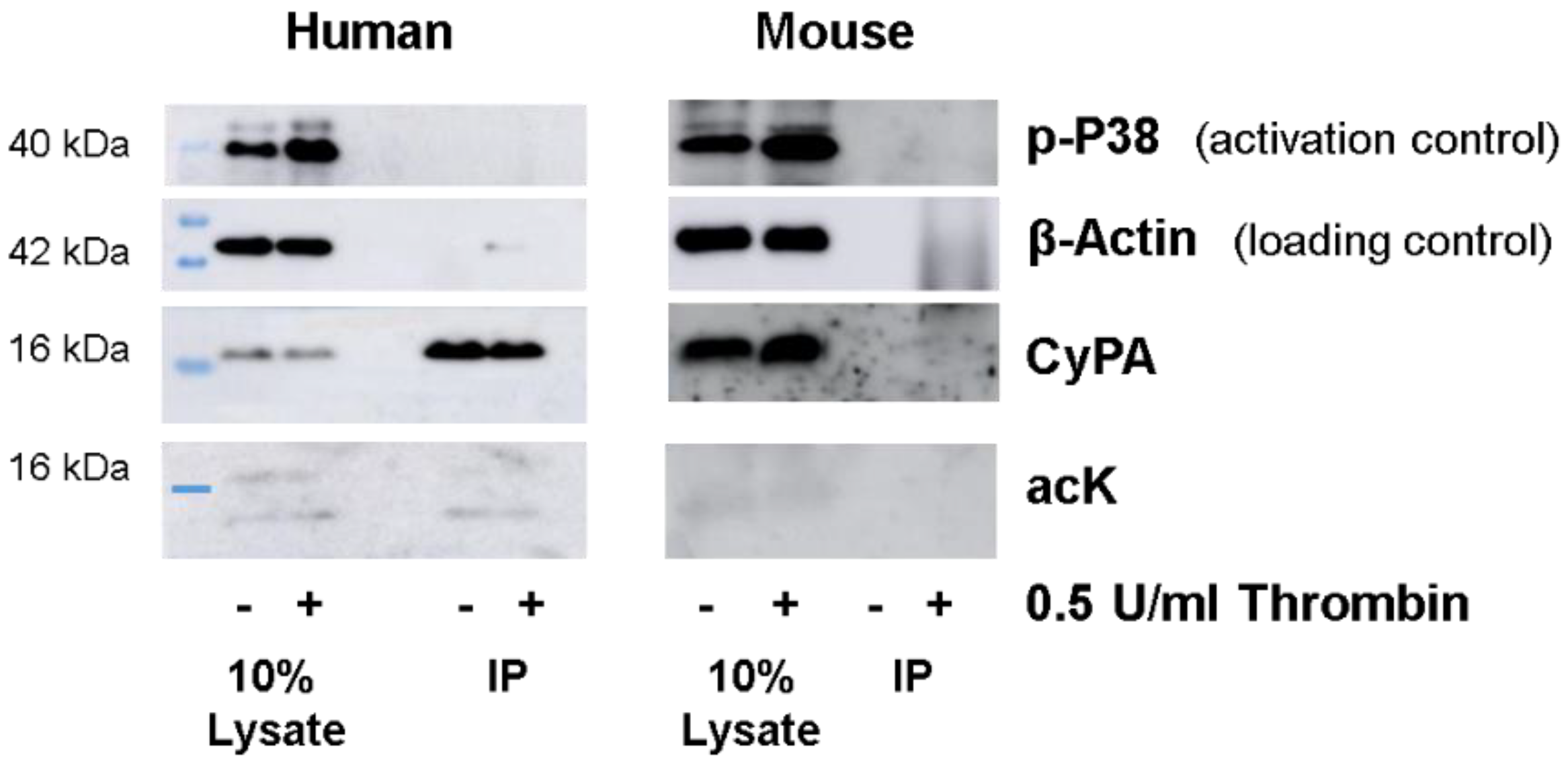
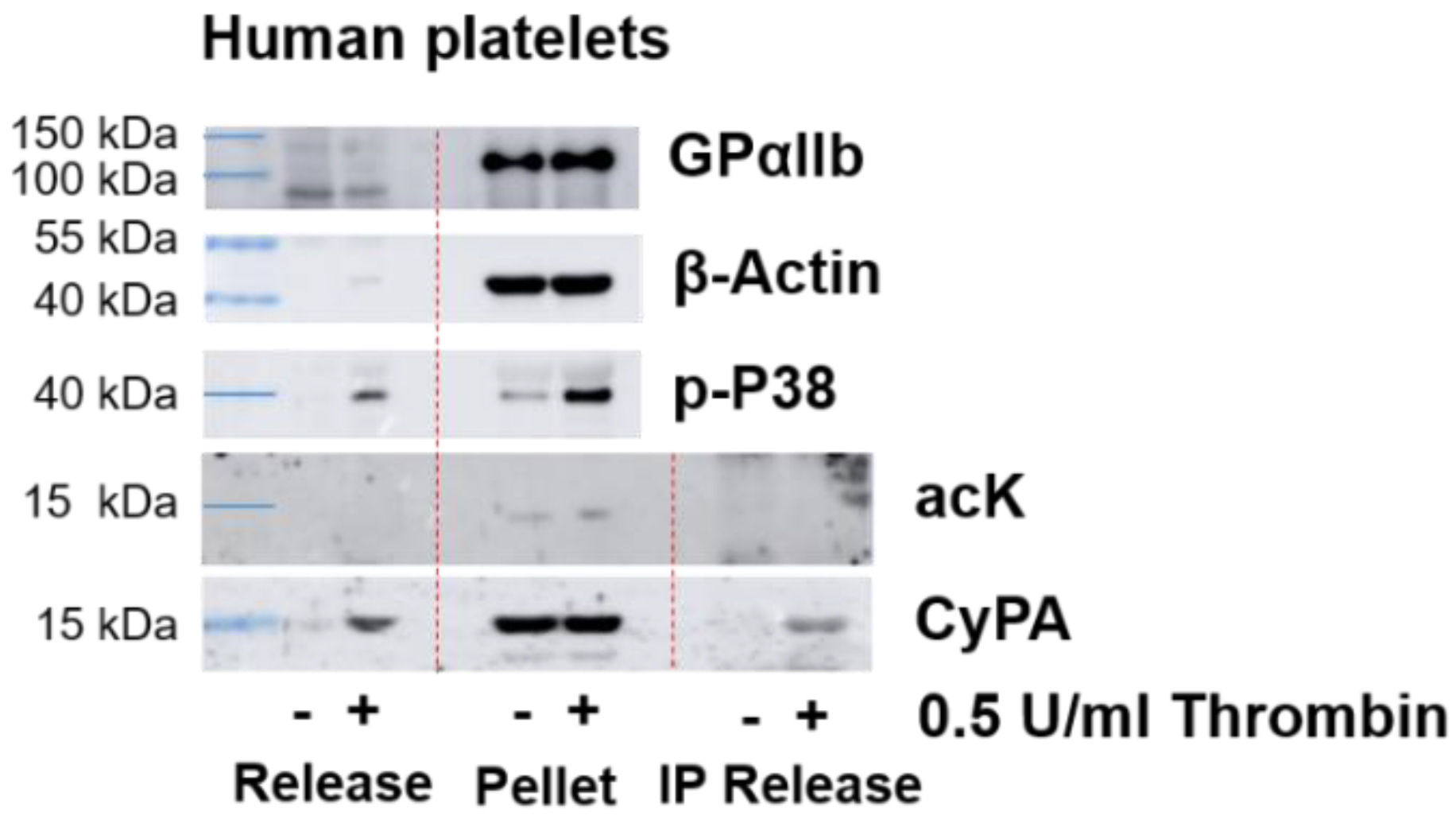
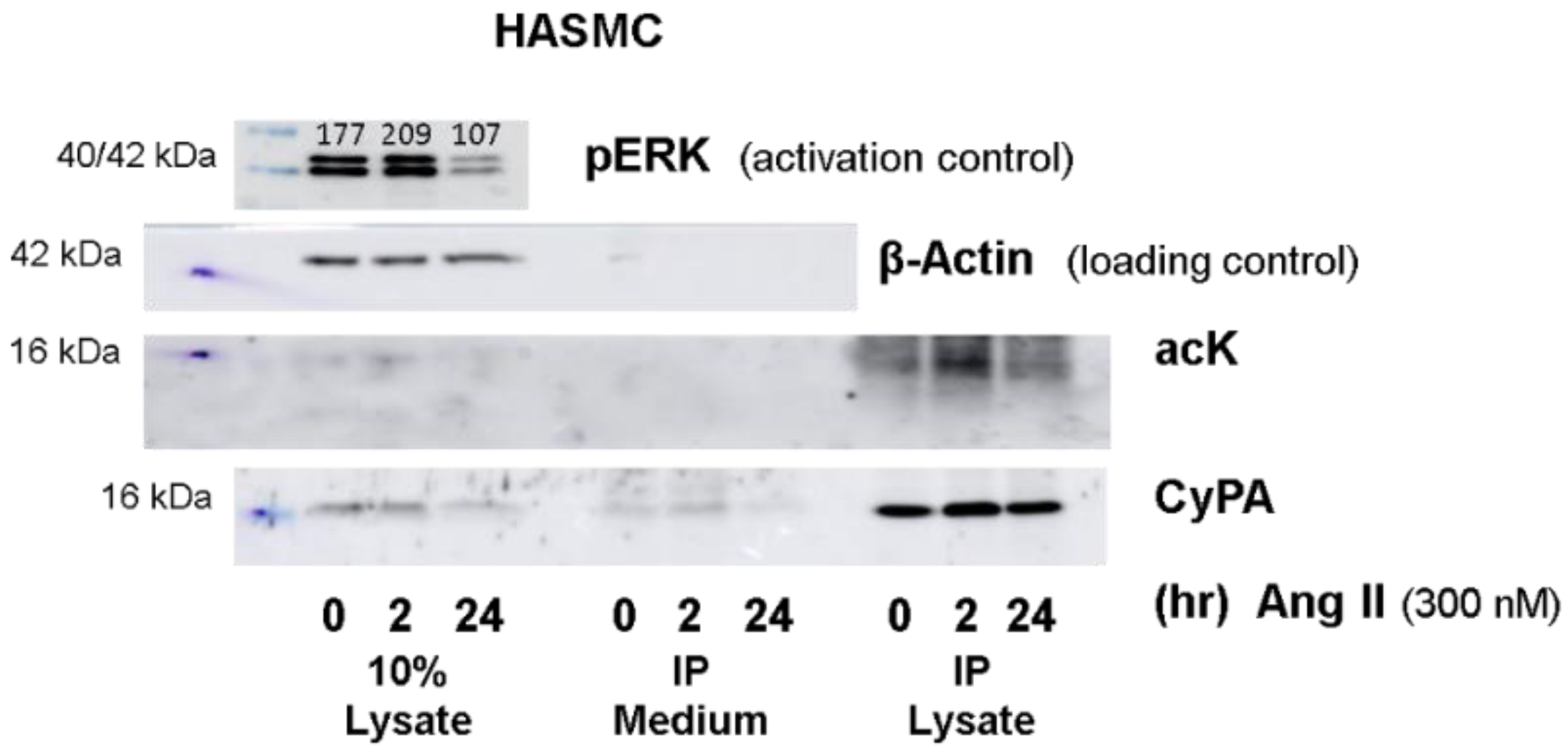
2.1. Western Blot Analyses of CyPA Acetylation in Platelets
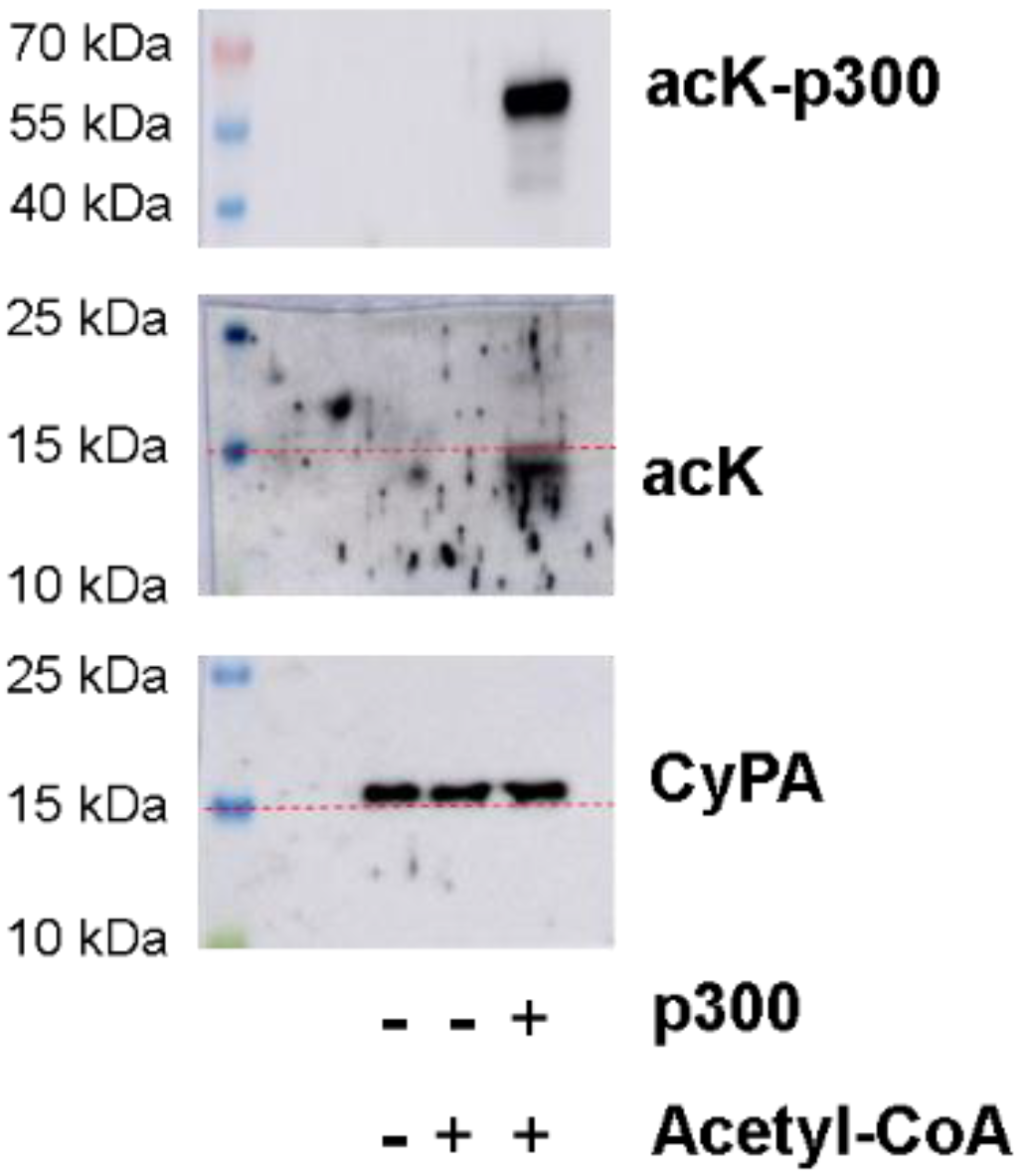
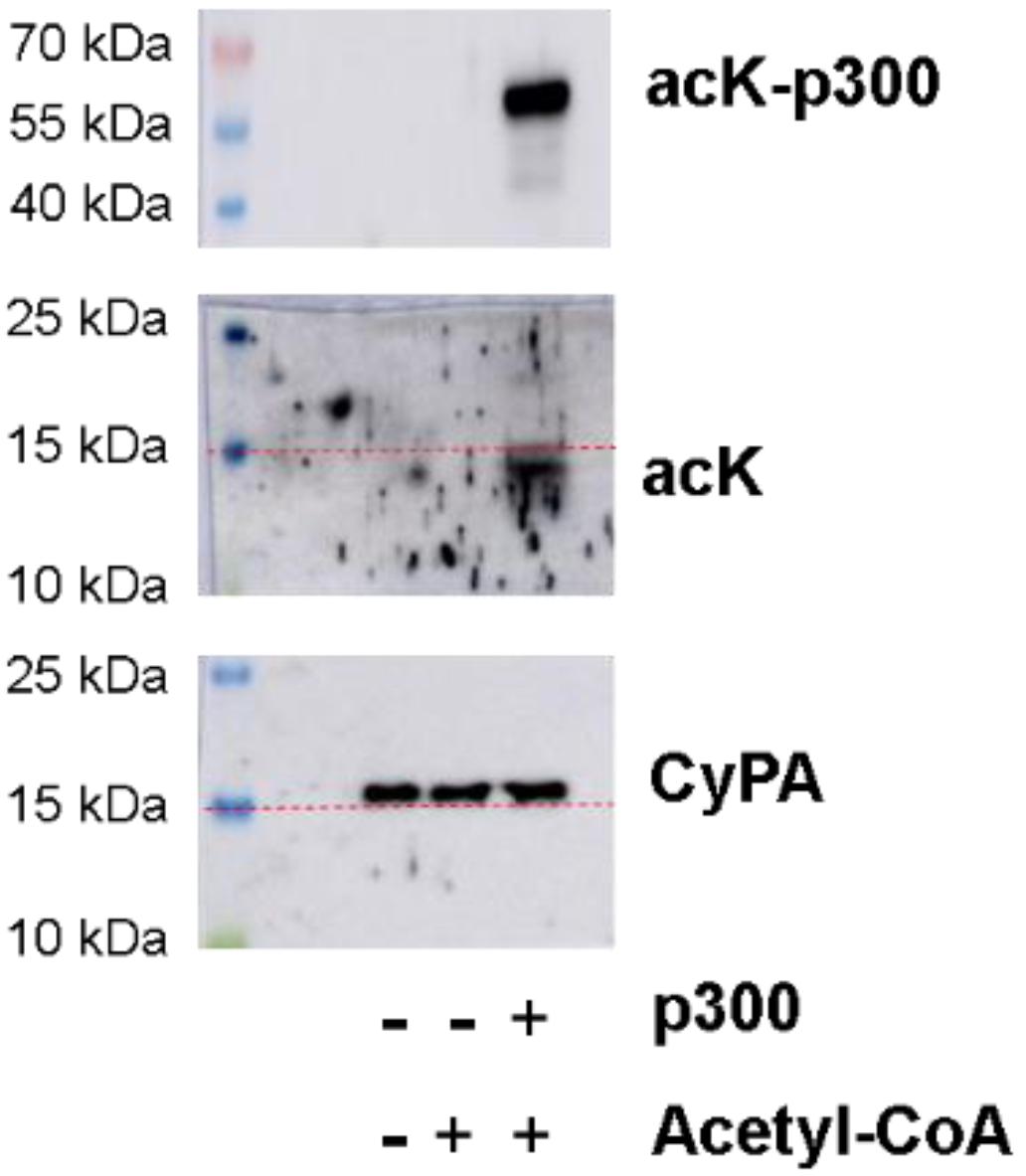
2.2. Recombinant CyPA Acetylation by p300
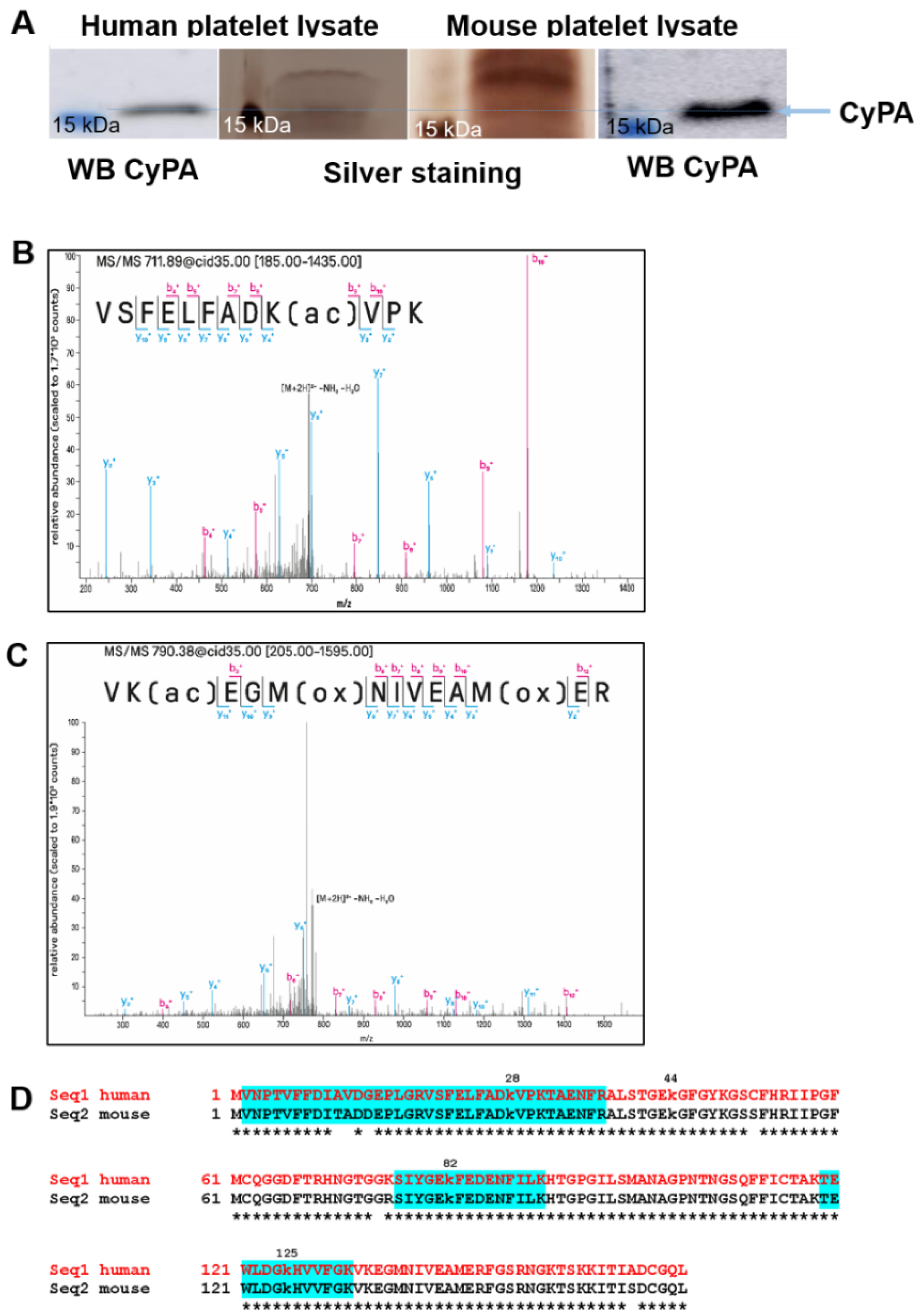
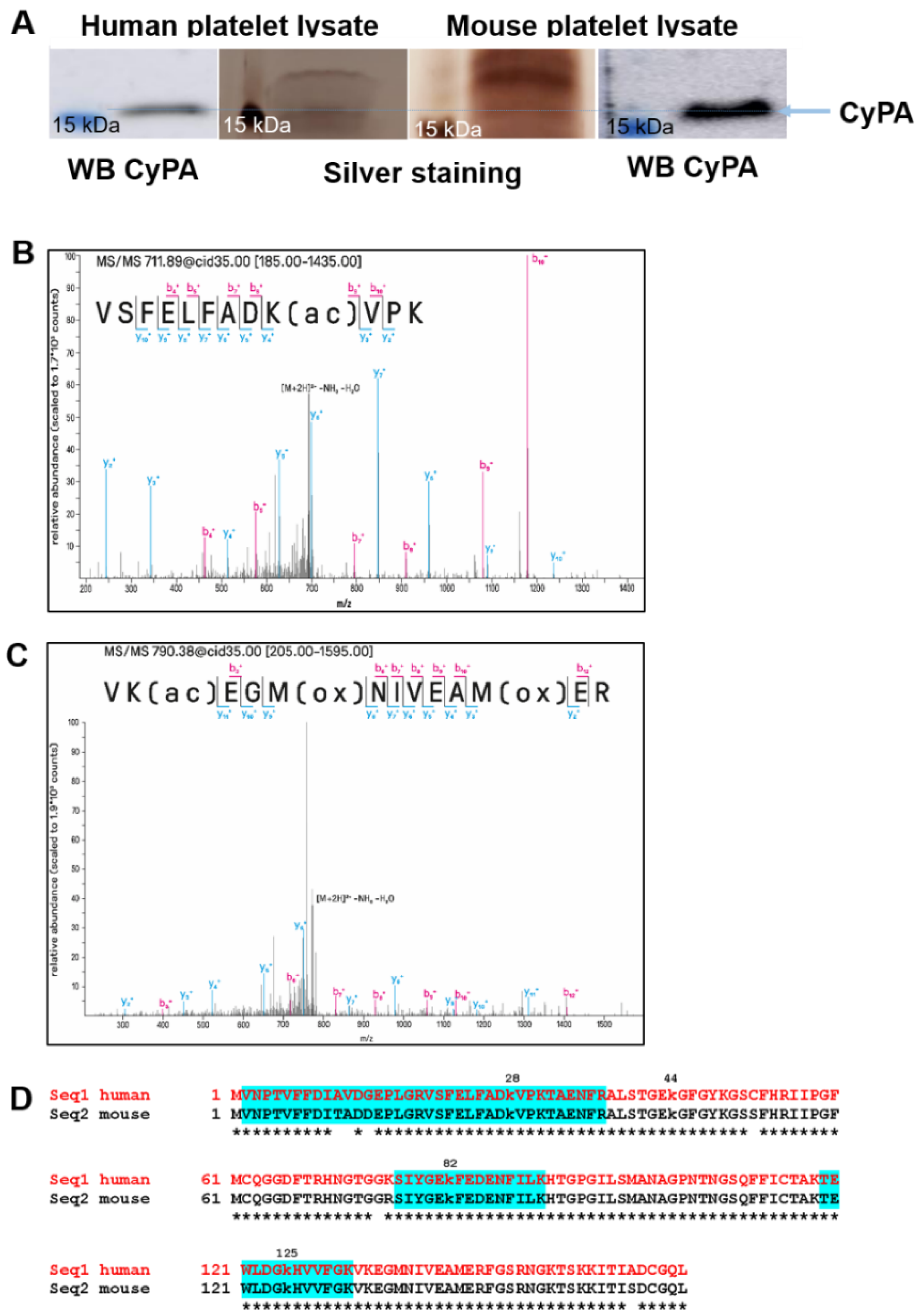
2.3. Mass Spectrometric Analyses of CyPA
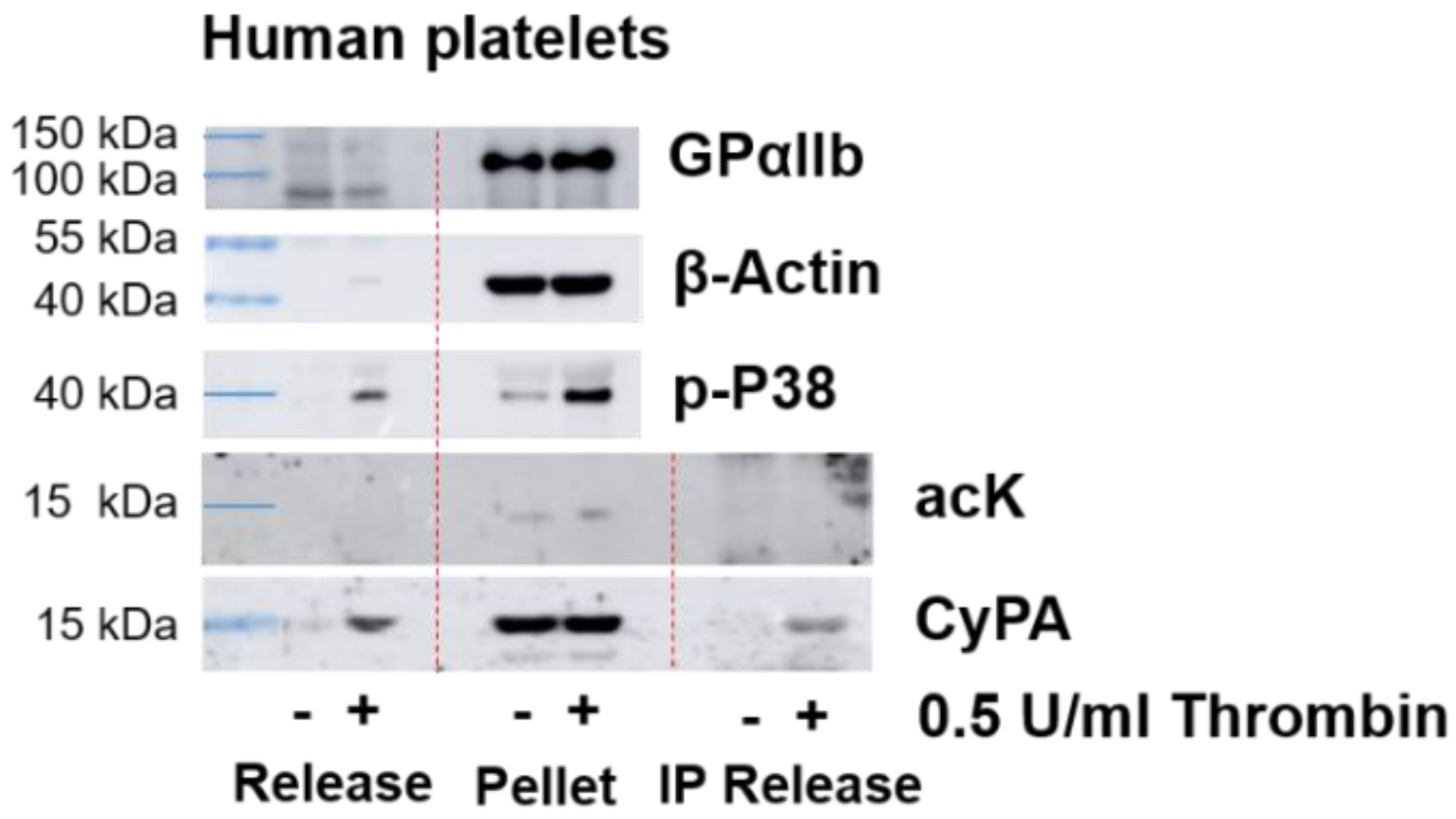
2.4. CyPA Release
3. Discussion
4. Materials and Methods
4.1. Platelet Preparation
4.2. Immunoprecipitation
4.3. In Vitro Acetylation Assay
4.4. Western Blot
4.5. Cultivation of Primary Aortic Smooth Muscle Cells (HASMCs)
4.6. Silver Staining
4.7. Processing of Gel Bands for LC–MS
4.8. Data-Dependent Acquisition
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Satoh, K.; Shimokawa, H.; Berk, B.C. Cyclophilin A—Promising New Target in Cardiovascular Therapy. Circ. J. 2010, 74, 2249–2256. [Google Scholar] [CrossRef] [Green Version]
- Franke, E.K.; Yuan, H.E.H.; Luban, J. Specific incorporation of cyclophilin A into HIV-1 virions. Nature 1994, 372, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Sowden, M.P.; Berk, B.C. Extracellular and Intracellular Cyclophilin A, Native and Post-Translationally Modified, Show Diverse and Specific Pathological Roles in Diseases. Arter. Thromb. Vasc. Biol. 2018, 38, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, W.; Ge, H.; He, B. Pro-inflammatory activities induced by CyPA-EMMPRIN interaction in monocytes. Atherosclerosis 2010, 213, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Yang, Y. The roles of CyPA and CD147 in cardiac remodelling. Exp. Mol. Pathol. 2018, 104, 222–226. [Google Scholar] [CrossRef]
- Yurchenko, V.; Zybarth, G.; O’Connor, M.; Dai, W.W.; Franchin, G.; Hao, T.; Guo, H.; Hung, H.-C.; Toole, B.; Gallay, P.; et al. Active Site Residues of Cyclophilin A Are Crucial for Its Signaling Activity via CD147. J. Biol. Chem. 2002, 277, 22959–22965. [Google Scholar] [CrossRef] [Green Version]
- Seizer, P.; Gawaz, M.; May, A.E. Cyclophilin A and EMMPRIN (CD147) in cardiovascular diseases. Cardiovasc. Res. 2014, 102, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Lefrançais, E.; Ortiz-Muñoz, G.; Caudrillier, A.; Mallavia, B.; Liu, F.; Sayah, D.M.; Thornton, E.E.; Headley, M.B.; David, T.; Coughlin, S.R.; et al. The lung is a site of platelet biogenesis and a reservoir for haematopoietic progenitors. Nature 2017, 544, 105–109. [Google Scholar] [CrossRef]
- Ruggeri, Z.M. Platelets in atherothrombosis. Nature Med. 2002, 8, 1227–1234. [Google Scholar] [CrossRef]
- Arman, M.; Payne, H.; Ponomaryov, T.; Brill, A. Role of Platelets in Inflammation. In The Non-Thrombotic Role of Platelets in Health and Disease; Kerrigan, S.W., Moran, N., Eds.; IntechOpen: Rijeka, Croatia, 2015; Chapter 3. [Google Scholar]
- Vogel, S.; Bodenstein, R.; Chen, Q.; Feil, S.; Feil, R.; Rheinlaender, J.; Schäffer, T.E.; Bohn, E.; Frick, J.S.; Borst, O.; et al. Platelet-derived HMGB1 is a critical mediator of thrombosis. J. Clin. Investig. 2015, 125, 4638–4654. [Google Scholar] [CrossRef] [Green Version]
- Elvers, M.; Herrmann, A.; Seizer, P.; Münzer, P.; Beck, S.; Schönberger, T.; Borst, O.; Martin-Romero, F.J.; Lang, F.; May, A.E.; et al. Intracellular cyclophilin A is an important Ca(2+) regulator in platelets and critically involved in arterial thrombus formation. Blood 2012, 120, 1317–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Soe, N.N.; Sowden, M.; Xu, Y.; Modjeski, K.; Baskaran, P.; Kim, Y.; Smolock, E.M.; Morrell, C.N.; Berk, B.C. Cyclophilin A is an important mediator of platelet function by regulating integrin αIIbβ3 bidirectional signalling. Thromb. Haemost. 2014, 111, 873–882. [Google Scholar] [PubMed]
- Coppinger, J.A.; Cagney, G.; Toomey, S.; Kislinger, T.; Belton, O.; McRedmond, J.P.; Cahill, D.J.; Emili, A.; Fitzgerald, D.J.; Maguire, P.B. Characterization of the proteins released from activated platelets leads to localization of novel platelet proteins in human atherosclerotic lesions. Blood 2004, 103, 2096–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.S.; Su, S.L.; Kuo, C.L.; Huang, C.S.; Tseng, W.M.; Lin, S.S.; Liu, C.S. Cyclophilin A: A Predictive Biomarker of Carotid Stenosis in Cerebral Ischemic Stroke. Curr. Neurovasc. Res. 2018, 15, 111–119. [Google Scholar] [CrossRef]
- Yan, J.; Zang, X.; Chen, R.; Yuan, W.; Gong, J.; Wang, C.; Li, Y. The clinical implications of increased cyclophilin A levels in patients with acute coronary syndromes. Clin. Chim. Acta Int. J. Clin. Chem. 2012, 413, 691–695. [Google Scholar] [CrossRef]
- Rajh Hamza Al-Kraity, W.; Al-Dujaili, A. Assessment of cyclophilin-A level in women with heart disease after menopause. Res. J. Pharm. Technol. 2017, 10, 1675–1678. [Google Scholar] [CrossRef]
- Satoh, K.; Fukumoto, Y.; Sugimura, K.; Miura, Y.; Aoki, T.; Nochioka, K.; Tatebe, S.; Miyamichi-Yamamoto, S.; Shimizu, T.; Osaki, S.; et al. Plasma cyclophilin A is a novel biomarker for coronary artery disease. Circ. J. Off. J. Jpn. Circ. Soc. 2012, 77, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-H.; Chang, C.-C.; Kuo, C.-L.; Huang, C.-S.; Lin, C.-S.; Liu, C.-S. Decrease in Plasma Cyclophilin A Concentration at 1 Month after Myocardial Infarction Predicts Better Left Ventricular Performance and Synchronicity at 6 Months: A Pilot Study in Patients with ST Elevation Myocardial Infarction. Int. J. Biol. Sci. 2015, 11, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Sowden, M.; Berk, B.C. Extracellular Cyclophilin A, Especially Acetylated, Causes Pulmonary Hypertension by Stimulating Endothelial Apoptosis, Redox Stress, and Inflammation. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1138–1146. [Google Scholar] [CrossRef] [Green Version]
- Seizer, P.; Ungern-Sternberg, S.N.I.v.; Schönberger, T.; Borst, O.; Münzer, P.; Schmidt, E.-M.; Mack, A.F.; Heinzmann, D.; Chatterjee, M.; Langer, H.; et al. Extracellular Cyclophilin A Activates Platelets Via EMMPRIN (CD147) and PI3K/Akt Signaling, Which Promotes Platelet Adhesion and Thrombus Formation In Vitro and In Vivo. Arter. Thromb. Vasc. Biol. 2015, 35, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Soe, N.N.; Sowden, M.; Baskaran, P.; Kim, Y.; Nigro, P.; Smolock, E.M.; Berk, B.C. Acetylation of cyclophilin A is required for its secretion and vascular cell activation. Cardiovasc. Res. 2014, 101, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Morozov, Y.A.; Khromykh, L.M.; Dementieva, I.I.; Charnaia, M.A.; Kulikova, N.L.; Kazansky, D.B. Recombinant Human Cyclophilin A In Vitro Inhibits the Formation of Fibrin Clot. ActaNaturae 2012, 4, 98–101. [Google Scholar]
- Rath, D.; von Ungern-Sternberg, S.; Heinzmann, D.; Sigle, M.; Monzien, M.; Horstmann, K.; Schaeffeler, E.; Winter, S.; Müller, K.; Groga-Bada, P.; et al. Platelet surface expression of cyclophilin A is associated with increased mortality in patients with symptomatic coronary artery disease. J. Thromb. Haemost. JTH 2020, 18, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Tao, Y.; Li, M.; Che, T.; Qu, J. Protein acetylation and deacetylation: An important regulatory modification in gene transcription (Review). Exp. Therapeutic Med. 2020, 20, 2923–2940. [Google Scholar] [CrossRef]
- Aslan, J.E.; Rigg, R.A.; Nowak, M.S.; Loren, C.P.; Baker-Groberg, S.M.; Pang, J.; David, L.L.; McCarty, O.J. Lysine acetyltransfer supports platelet function. J. Thromb. Haemost. JTH 2015, 13, 1908–1917. [Google Scholar] [CrossRef]
- Guan, K.L.; Yu, W.; Lin, Y.; Xiong, Y.; Zhao, S. Generation of acetyllysine antibodies and affinity enrichment of acetylated peptides. Nature Protoc. 2010, 5, 1583–1595. [Google Scholar] [CrossRef] [Green Version]
- Lammers, M.; Neumann, H.; Chin, J.W.; James, L.C. Acetylation regulates Cyclophilin A catalysis, immunosuppression and HIV isomerization. Nature Chem. Biol. 2010, 6, 331–337. [Google Scholar] [CrossRef]
- Chevalier, F.; Depagne, J.; Hem, S.; Chevillard, S.; Bensimon, J.; Bertrand, P.; Lebeau, J. Accumulation of cyclophilin A isoforms in conditioned medium of irradiated breast cancer cells. Proteomics 2012, 12, 1756–1766. [Google Scholar] [CrossRef]
- Tribouillard-Tanvier, D.; Carroll, J.A.; Moore, R.A.; Striebel, J.F.; Chesebro, B. Role of cyclophilin A from brains of prion-infected mice in stimulation of cytokine release by microglia and astroglia in vitro. J. Biol. Chem. 2012, 287, 4628–4639. [Google Scholar] [CrossRef] [Green Version]
- Coppinger, J.A.; Maguire, P.B. Insights into the platelet releasate. Curr. Pharm. Des. 2007, 13, 2640–2646. [Google Scholar] [CrossRef]
- Maynard, D.M.; Heijnen, H.F.; Horne, M.K.; White, J.G.; Gahl, W.A. Proteomic analysis of platelet alpha-granules using mass spectrometry. J. Thromb. Haemost. JTH 2007, 5, 1945–1955. [Google Scholar] [CrossRef]
- Lundby, A.; Lage, K.; Weinert, B.T.; Bekker-Jensen, D.B.; Secher, A.; Skovgaard, T.; Kelstrup, C.D.; Dmytriyev, A.; Choudhary, C.; Lundby, C.; et al. Proteomic analysis of lysine acetylation sites in rat tissues reveals organ specificity and subcellular patterns. Cell Rep. 2012, 2, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Valentine, S.J.; Plasencia, M.D.; Trimpin, S.; Naylor, S.; Clemmer, D.E. Mapping the human plasma proteome by SCX-LC-IMS-MS. J. Am. Soc. Mass Spectrom. 2007, 18, 1249–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatham, M.H.; Cole, C.; Scullion, P.; Wilkie, R.; Westwood, N.J.; Stark, L.A.; Hay, R.T. A Proteomic Approach to Analyze the Aspirin-mediated Lysine Acetylome. Mol. Cell. Proteom. MCP 2017, 16, 310–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, C.J.; Zhang, J.; He, Y.; Lee, Y.M.; Chen, S.; Lim, T.K.; Ng, S.; Shen, H.M.; Lin, Q. Mapping sites of aspirin-induced acetylations in live cells by quantitative acid-cleavable activity-based protein profiling (QA-ABPP). Sci. Rep. 2015, 5, 7896. [Google Scholar] [CrossRef] [Green Version]
- Bateman, L.A.; Zaro, B.W.; Miller, S.M.; Pratt, M.R. An Alkyne–Aspirin Chemical Reporter for the Detection of Aspirin-Dependent Protein Modification in Living Cells. J. Am. Chem. Soc. 2013, 135, 14568–14573. [Google Scholar] [CrossRef] [PubMed]
- Coppinger, J.A.; O’Connor, R.; Wynne, K.; Flanagan, M.; Sullivan, M.; Maguire, P.B.; Fitzgerald, D.J.; Cagney, G. Moderation of the platelet releasate response by aspirin. Blood 2007, 109, 4786–4792. [Google Scholar] [CrossRef]
- Suzuki, J.; Jin, Z.-G.; Meoli, D.F.; Matoba, T.; Berk, B.C. Cyclophilin A Is Secreted by a Vesicular Pathway in Vascular Smooth Muscle Cells. Circ. Res. 2006, 98, 811–817. [Google Scholar] [CrossRef] [Green Version]
- Schölz, C.; Weinert, B.T.; Wagner, S.A.; Beli, P.; Miyake, Y.; Qi, J.; Jensen, L.J.; Streicher, W.; McCarthy, A.R.; Westwood, N.J.; et al. Acetylation site specificities of lysine deacetylase inhibitors in human cells. Nat. Biotechnol. 2015, 33, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, A.; Butt, E.; Hopper, C.P.; Loroch, S.; Bender, M.; Schulze, H.; Sickmann, A.; Vorlova, S.; Seizer, P.; Heinzmann, D.; et al. Cyclophilin A Is Not Acetylated at Lysine-82 and Lysine-125 in Resting and Stimulated Platelets. Int. J. Mol. Sci. 2022, 23, 1469. https://doi.org/10.3390/ijms23031469
Rosa A, Butt E, Hopper CP, Loroch S, Bender M, Schulze H, Sickmann A, Vorlova S, Seizer P, Heinzmann D, et al. Cyclophilin A Is Not Acetylated at Lysine-82 and Lysine-125 in Resting and Stimulated Platelets. International Journal of Molecular Sciences. 2022; 23(3):1469. https://doi.org/10.3390/ijms23031469
Chicago/Turabian StyleRosa, Annabelle, Elke Butt, Christopher P. Hopper, Stefan Loroch, Markus Bender, Harald Schulze, Albert Sickmann, Sandra Vorlova, Peter Seizer, David Heinzmann, and et al. 2022. "Cyclophilin A Is Not Acetylated at Lysine-82 and Lysine-125 in Resting and Stimulated Platelets" International Journal of Molecular Sciences 23, no. 3: 1469. https://doi.org/10.3390/ijms23031469
APA StyleRosa, A., Butt, E., Hopper, C. P., Loroch, S., Bender, M., Schulze, H., Sickmann, A., Vorlova, S., Seizer, P., Heinzmann, D., & Zernecke, A. (2022). Cyclophilin A Is Not Acetylated at Lysine-82 and Lysine-125 in Resting and Stimulated Platelets. International Journal of Molecular Sciences, 23(3), 1469. https://doi.org/10.3390/ijms23031469