
A Central Role for TRPM4 in Ca2+-Signal Amplification and Vasoconstriction


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
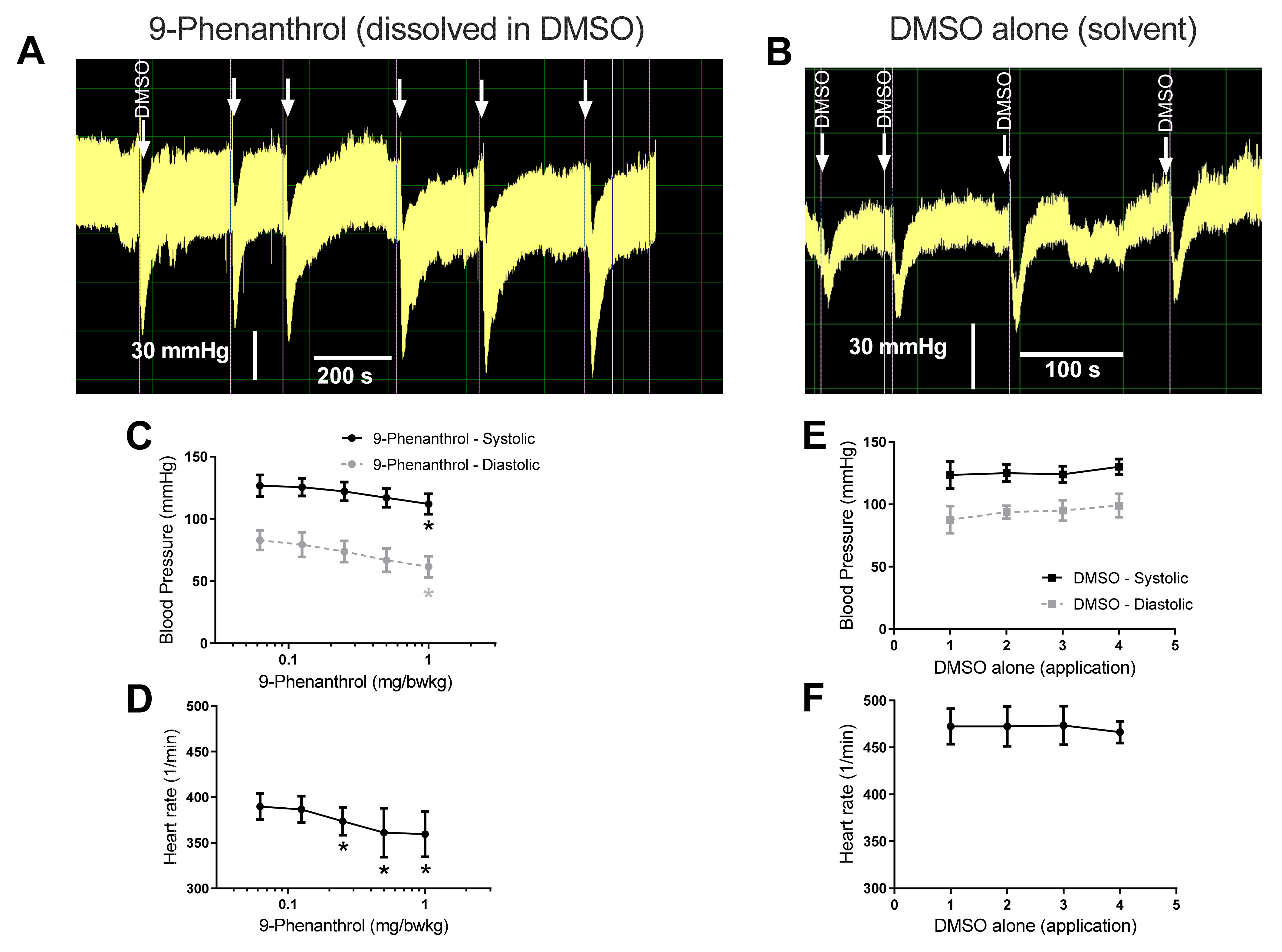
2.1. Pharmacological Inhibition of TRPM4 Results in a Dose-Dependent Decrease in Blood Pressure
2.2. TRPM4 Inhibition Abolishes Myogenic Response to Pressure Elevation in Skeletal Muscle Arteries
2.3. TRPM4 Inhibition Completely Antagonized Norepinephrine Evoked Vasoconstriction
2.4. TRPM4 Inhibition Completely Antagonized Vasoconstriction Caused by High Extracellular KCl Concentration-Evoked Depolarization of the Smooth Muscle Cells
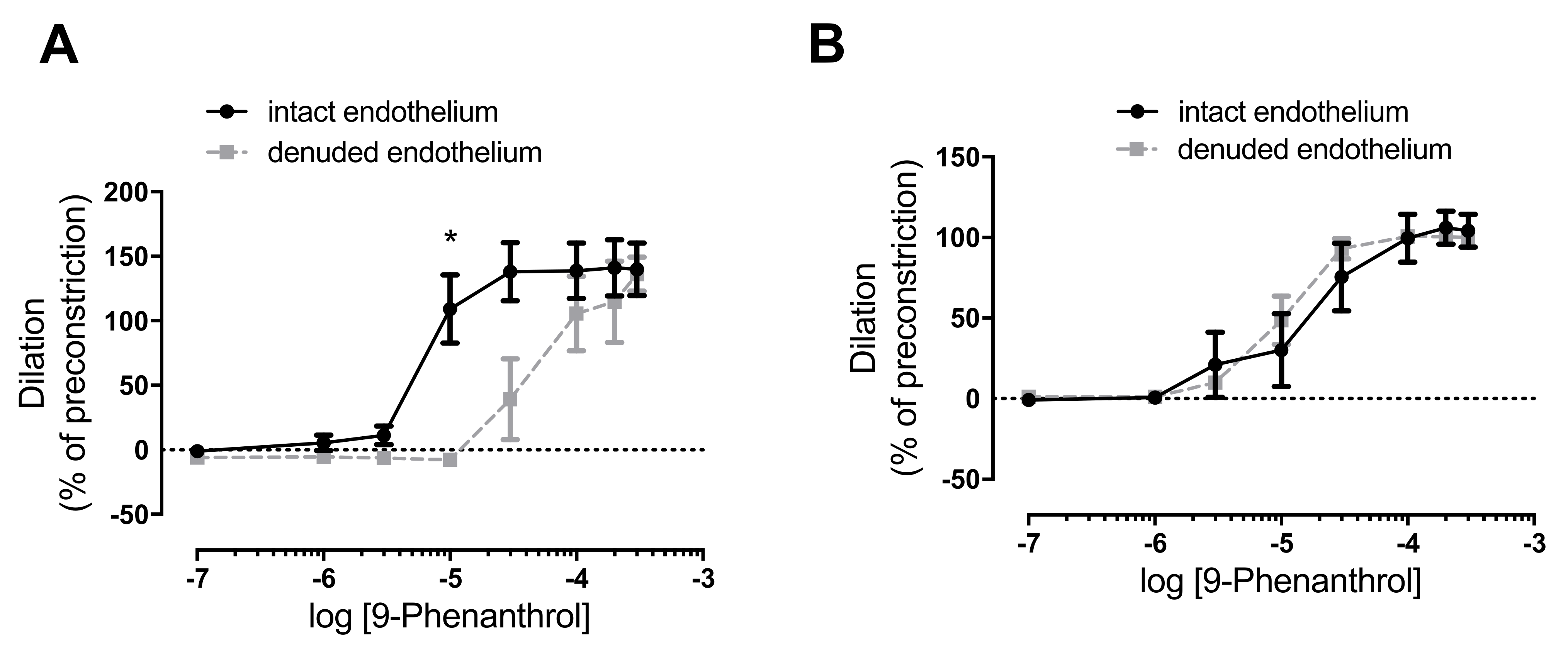
2.5. TRPM4 Inhibition Completely Antagonized Norepinephrine-Evoked Vasoconstriction in the Absence of Endothelium
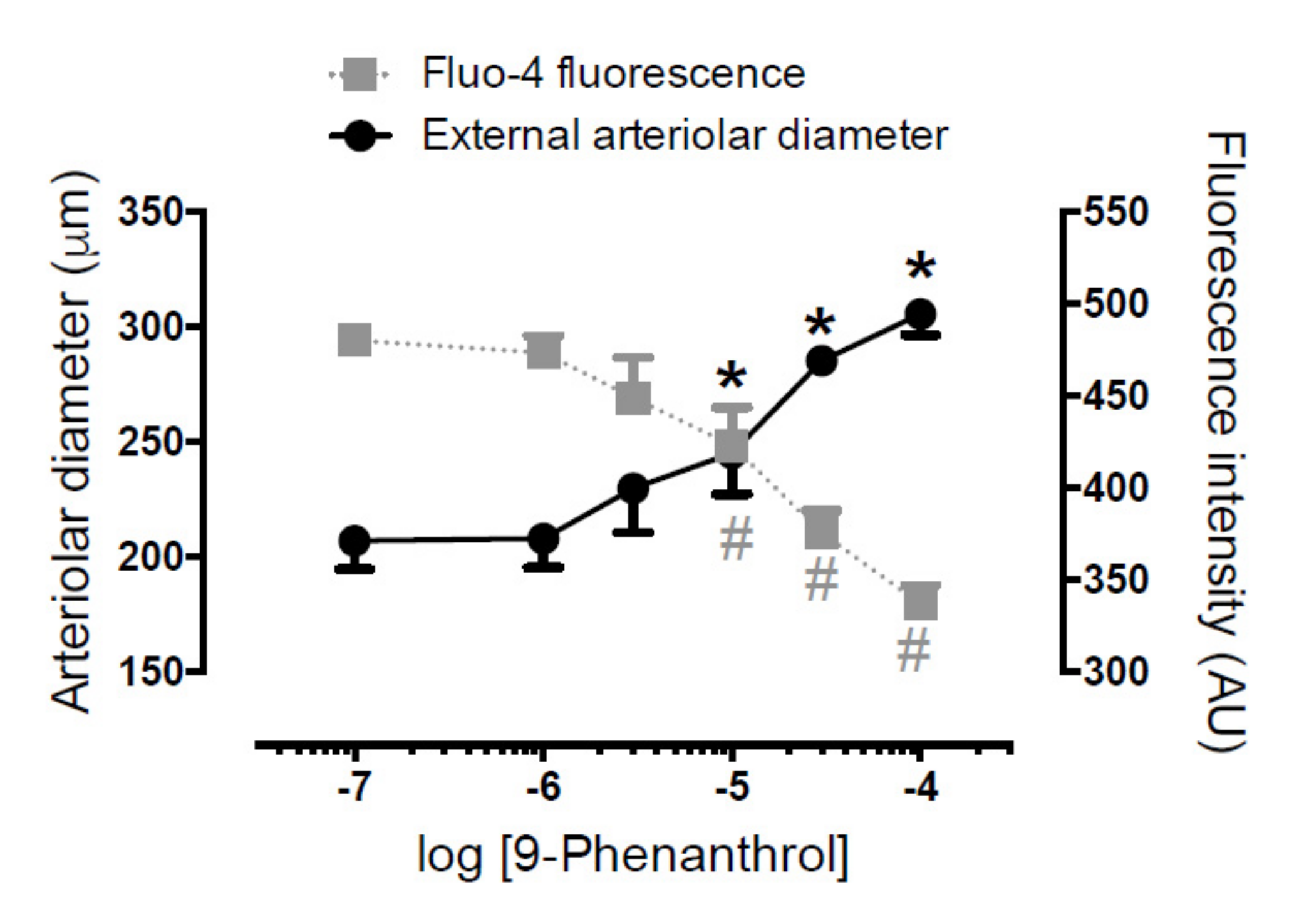
2.6. TRPM4 Inhibition Antagonized Norepinephrine-Evoked Vasoconstriction on Skeletal Muscle Arteries by Decreasing Intracellular Ca2+ Concentration in Vascular Smooth Muscle Cells
2.7. Immunohistochemistry Confirmed Expression of TRPM4 in Arteries Used in the Functional Assays
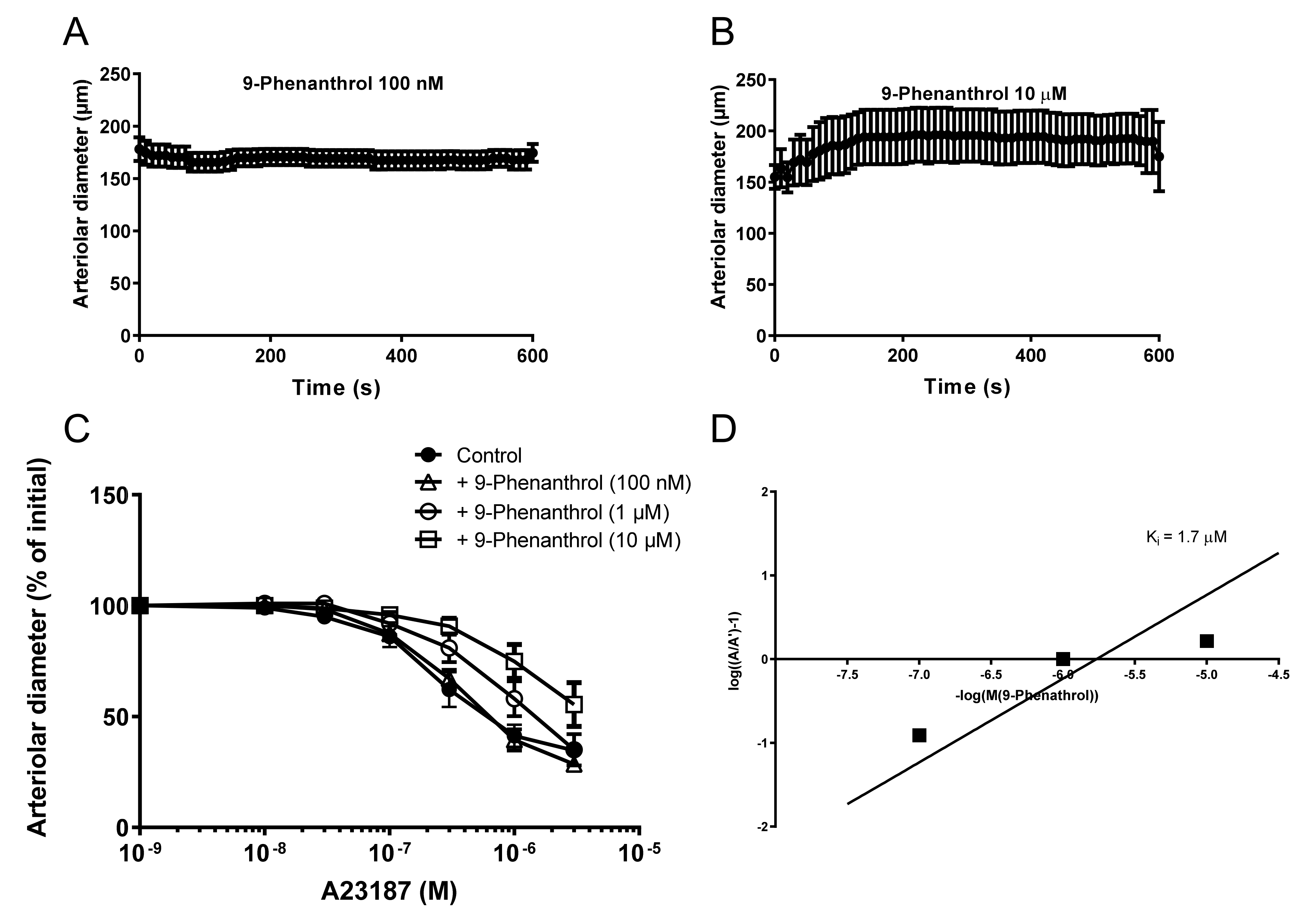
2.8. The Ca2+ Ionophore A23187 Acts as a TRPM4 Agonist Which Is Competitively Antagonized by 9-Phenanthrol in Skeletal Muscle Arteries
3. Discussion
4. Materials and Methods
4.1. Animal Housing and Experimental Protocol Approval
4.2. Invasive Measurement of Blood Pressure
4.3. Pressure Myography of Arteries
4.4. Wire Myography of Arteries
4.5. Immunohistochemistry
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Earley, S. Vanilloid and Melastatin Transient Receptor Potential Channels in Vascular Smooth Muscle. Microcirculation 2010, 17, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapham, D.E.; Julius, D.; Montell, C.; Schultz, G. International Union of Pharmacology. XLIX. Nomenclature and struc-ture-function relationships of transient receptor potential channels. Pharmacol. Rev. 2005, 57, 427–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earley, S.; Straub, S.V.; Brayden, J.E. Protein kinase C regulates vascular myogenic tone through activation of TRPM4. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2613–H2622. [Google Scholar] [CrossRef] [PubMed]
- Earley, S.; Waldron, B.J.; Brayden, J.E. Critical role for transient receptor potential channel TRPM4 in myogenic constriction of cerebral arteries. Circ. Res. 2004, 95, 922–929. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Baylie, R.L.; Tavares, M.J.; E Brayden, J. TRPM4 Channels Couple Purinergic Receptor Mechanoactivation and Myogenic Tone Development in Cerebral Parenchymal Arterioles. Br. J. Pharmacol. 2014, 34, 1706–1714. [Google Scholar] [CrossRef] [Green Version]
- Reading, S.A.; Brayden, J.E. Central Role of TRPM4 Channels in Cerebral Blood Flow Regulation. Stroke 2007, 38, 2322–2328. [Google Scholar] [CrossRef] [Green Version]
- Launay, P.; Fleig, A.; Perraud, A.L.; Scharenberg, A.M.; Penner, R.; Kinet, J.P. TRPM4 is a Ca2+-activated nonselective cation channel mediating cell membrane depolarization. Cell 2002, 109, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Mahieu, F.; Prenen, J.; Janssens, A.; Owsianik, G.; Vennekens, R.; Voets, T. The Ca2+-activated cation channel TRPM4 is regulated by phosphatidylinositol 4,5-biphosphate. EMBO J. 2006, 25, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Okawa, H.; Wang, Y.; Liman, E.R. Phosphatidylinositol 4,5-Bisphosphate Rescues TRPM4 Channels from Desensitization. J. Biol. Chem. 2005, 280, 39185–39192. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Prenen, J.; Tang, J.; Wang, C.; Owsianik, G.; Janssens, A.; Voets, T.; Zhu, M.X. Regulation of the Ca2+ sensitivity of the nonselective cation channel TRPM4. J. Biol. Chem. 2005, 280, 6423–6433. [Google Scholar] [CrossRef] [Green Version]
- Launay, P.; Cheng, H.; Srivatsan, S.; Penner, R.; Fleig, A.; Kinet, J.P. TRPM4 regulates calcium oscillations after T cell activation. Science 2004, 306, 1374–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Beck, A.; Launay, P.; Gross, S.A.; Stokes, A.J.; Kinet, J.P.; Fleig, A.; Penner, R. TRPM4 controls insulin secretion in pancreatic β-cells. Cell Calcium 2007, 41, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinamard, R.; Bouvagnet, P.; Hof, T.; Liu, H.; Simard, C.; Sallé, L. TRPM4 in cardiac electrical activity. Cardiovasc. Res. 2015, 108, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abriel, H.; Syam, N.; Sottas, V.; Amarouch, M.Y.; Rougier, J.-S. TRPM4 channels in the cardiovascular system: Physiology, pathophysiology, and pharmacology. Biochem. Pharmacol. 2012, 84, 873–881. [Google Scholar] [CrossRef]
- Vennekens, R.; Nilius, B. Insights into TRPM4 Function, Regulation and Physiological Role. In Transient Receptor Potential (TRP) Channels; Springer: Berlin/Heidelberg, Germany, 2007; pp. 269–285. [Google Scholar] [CrossRef]
- Garland, C.J.; Smirnov, S.; Bagher, P.; Lim, C.S.; Huang, C.Y.; Mitchell, R.; Stanley, C.; Pinkney, A.; Dora, K. TRPM4 inhibitor 9-phenanthrol activates endothelial cell intermediate conductance calcium-activated potassium channels in rat isolated mesenteric artery. Br. J. Pharmacol. 2014, 172, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.-Q.; Ban, T.; Liu, Z.-Y.; Lou, J.; Tang, L.-L.; Wang, J.-X.; Chu, W.-F.; Zhao, D.; Song, B.-L.; Zhang, Z.-R. Transient Receptor Potential Melastatin 4 (TRPM4) Contributes to High Salt Diet-Mediated Early-Stage Endothelial Injury. Cell. Physiol. Biochem. 2017, 41, 835–848. [Google Scholar] [CrossRef] [Green Version]
- Gonzales, A.L.; Garcia, Z.I.; Amberg, G.C.; Earley, S. Pharmacological inhibition of TRPM4 hyperpolarizes vascular smooth muscle. Am. J. Physiol. Cell Physiol. 2010, 299, C1195–C1202. [Google Scholar] [CrossRef] [Green Version]
- Grand, T.; Demion, M.; Norez, C.; Mettey, Y.; Launay, P.; Becq, F.; Bois, P.; Guinamard, R. 9-Phenanthrol inhibits human TRPM4 but not TRPM5 cationic channels. Br. J. Pharmacol. 2008, 153, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Guinamard, R.; Hof, T.; Del Negro, C.A. The TRPM4 channel inhibitor 9-phenanthrol. J. Cereb. Blood Flow Metab. 2014, 171, 1600–1613. [Google Scholar] [CrossRef] [Green Version]
- Guinamard, R.; Demion, M.; Magaud, C.; Potreau, D.; Bois, P. Functional Expression of the TRPM4 Cationic Current in Ventricular Cardiomyocytes from Spontaneously Hypertensive Rats. Hypertension 2006, 48, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Demion, M.; Bois, P.; Launay, P.; Guinamard, R. TRPM4, a Ca2+-activated nonselective cation channel in mouse sino-atrial node cells. Cardiovasc. Res. 2007, 73, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demion, M.; Thireau, J.; Gueffier, M.; Finan, A.; Khoueiry, Z.; Cassan, C.; Serafini, N.; Aimond, F.; Granier, M.; Pasquié, J.-L.; et al. Trpm4 Gene Invalidation Leads to Cardiac Hypertrophy and Electrophysiological Alterations. PLoS ONE 2014, 9, e115256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; El Zein, L.; Kruse, M.; Guinamard, R.; Beckmann, A.; Bozio, A.; Kurtbay, G.; Mégarbané, A.; Ohmert, I.; Blaysat, G.; et al. Gain-of-Function Mutations in TRPM4 Cause Autosomal Dominant Isolated Cardiac Conduction Disease. Circ. Cardiovasc. Genet. 2010, 3, 374–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burris, S.K.; Wang, Q.; Bulley, S.; Neeb, Z.P.; Jaggar, J.H. 9-Phenanthrol inhibits recombinant and arterial myocyte TMEM16A channels. Br. J. Pharmacol. 2015, 172, 2459–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulley, S.; Neeb, Z.P.; Burris, S.K.; Bannister, J.P.; Thomas-Gatewood, C.M.; Jangsangthong, W.; Jaggar, J.H. TMEM16A/ANO1 Channels Contribute to the Myogenic Response in Cerebral Arteries. Circ. Res. 2012, 111, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warshaw, D.M.; Mulvany, M.J.; Halpern, W. Mechanical and morphological properties of arterial resistance vessels in young and old spontaneously hypertensive rats. Circ. Res. 1979, 45, 250–259. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csípő, T.; Czikora, Á.; Fülöp, G.Á.; Gulyás, H.; Rutkai, I.; Tóth, E.P.; Pórszász, R.; Szalai, A.; Bölcskei, K.; Helyes, Z.; et al. A Central Role for TRPM4 in Ca2+-Signal Amplification and Vasoconstriction. Int. J. Mol. Sci. 2022, 23, 1465. https://doi.org/10.3390/ijms23031465
Csípő T, Czikora Á, Fülöp GÁ, Gulyás H, Rutkai I, Tóth EP, Pórszász R, Szalai A, Bölcskei K, Helyes Z, et al. A Central Role for TRPM4 in Ca2+-Signal Amplification and Vasoconstriction. International Journal of Molecular Sciences. 2022; 23(3):1465. https://doi.org/10.3390/ijms23031465
Chicago/Turabian StyleCsípő, Tamás, Ágnes Czikora, Gábor Á. Fülöp, Hajnalka Gulyás, Ibolya Rutkai, Enikő Pásztorné Tóth, Róbert Pórszász, Andrea Szalai, Kata Bölcskei, Zsuzsanna Helyes, and et al. 2022. "A Central Role for TRPM4 in Ca2+-Signal Amplification and Vasoconstriction" International Journal of Molecular Sciences 23, no. 3: 1465. https://doi.org/10.3390/ijms23031465
APA StyleCsípő, T., Czikora, Á., Fülöp, G. Á., Gulyás, H., Rutkai, I., Tóth, E. P., Pórszász, R., Szalai, A., Bölcskei, K., Helyes, Z., Pintér, E., Papp, Z., Ungvári, Z., & Tóth, A. (2022). A Central Role for TRPM4 in Ca2+-Signal Amplification and Vasoconstriction. International Journal of Molecular Sciences, 23(3), 1465. https://doi.org/10.3390/ijms23031465