
Anti-Obesity Efficacy of Pediococcus acidilactici MNL5 in Canorhabditis elegans Gut Model

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of Cholesterol-Lowering LAB
2.2. Antibiotic Resistance
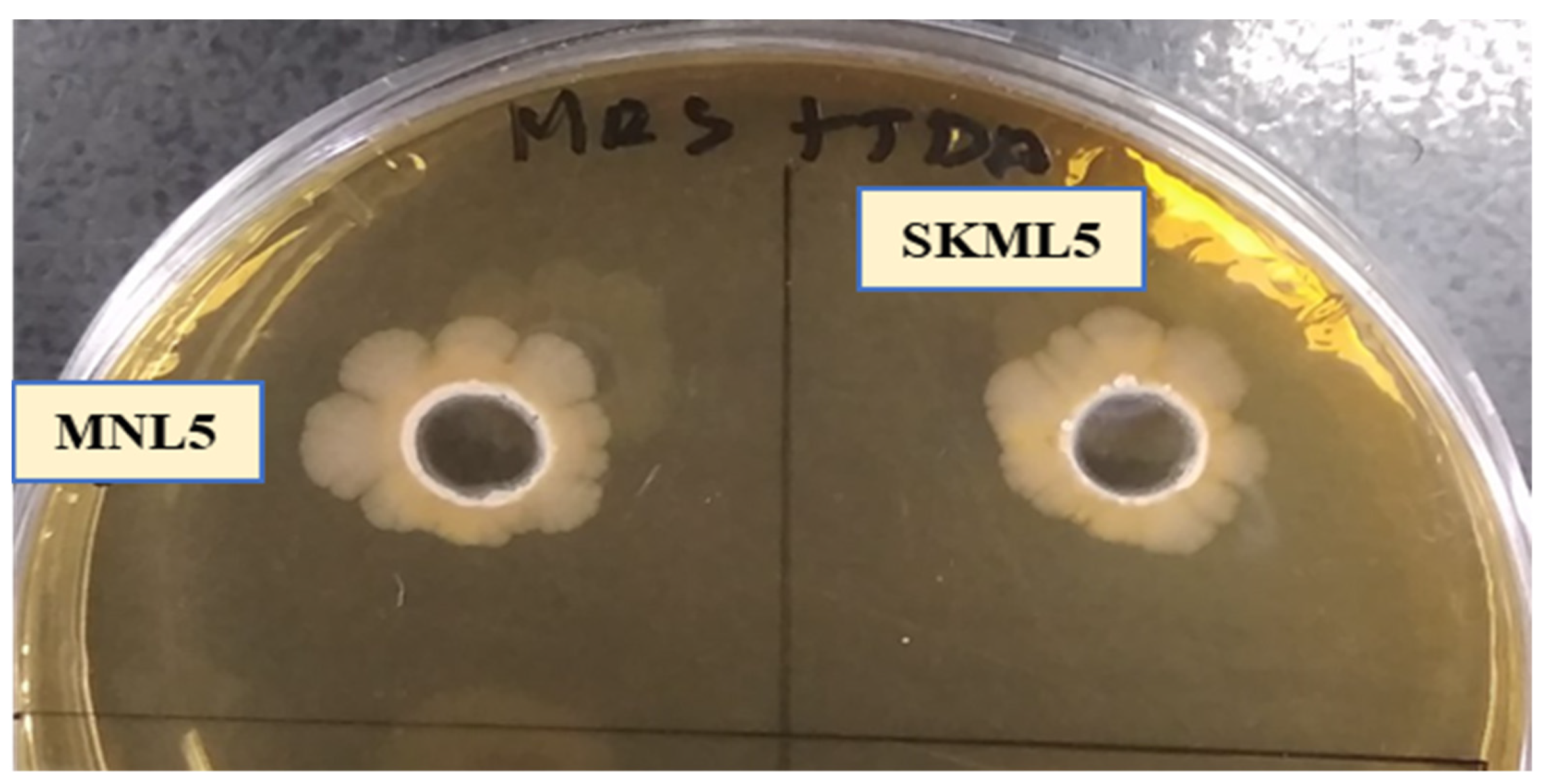
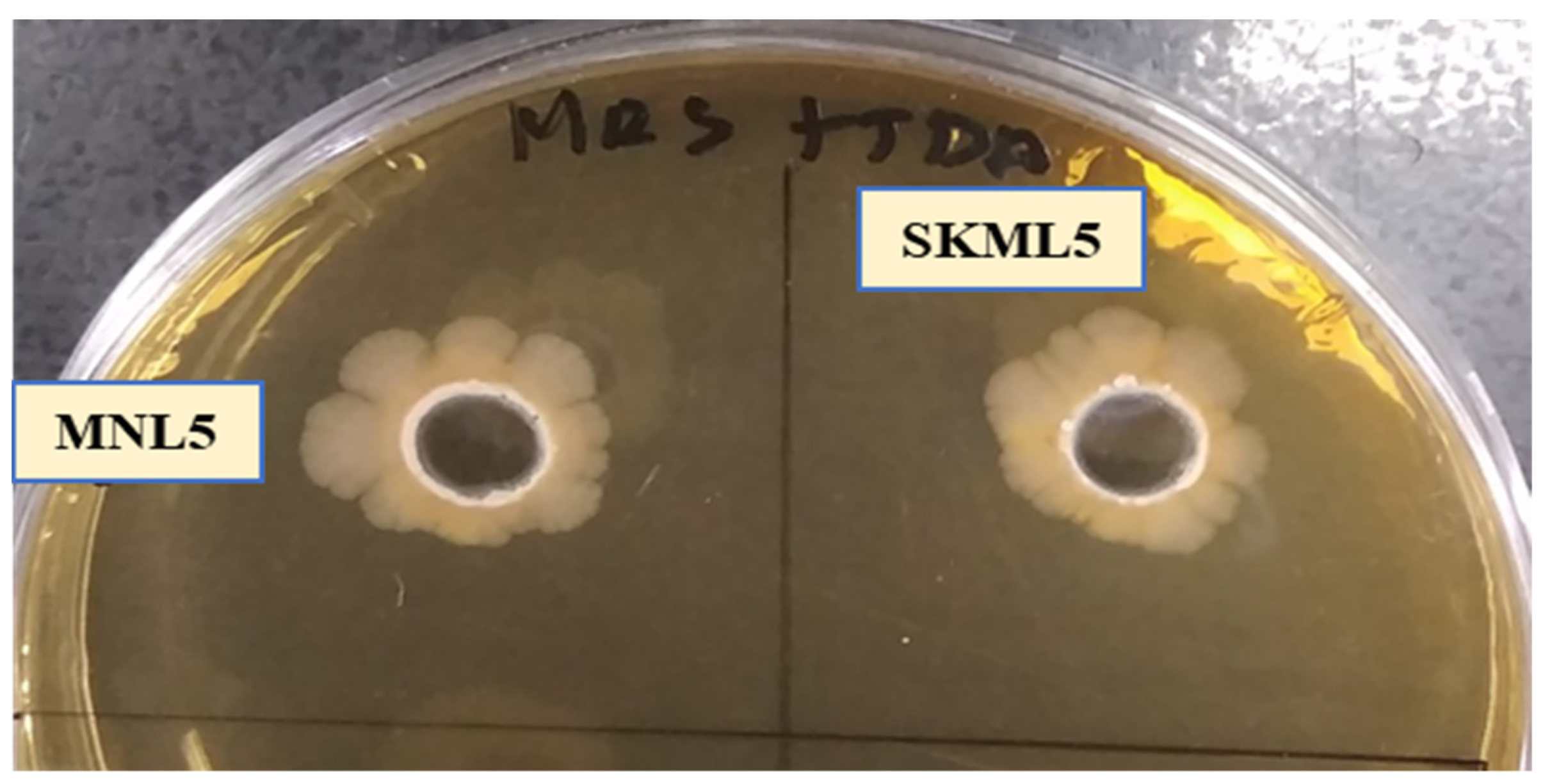
2.3. Antimicrobial Activity
2.4. Hemolytic Activity
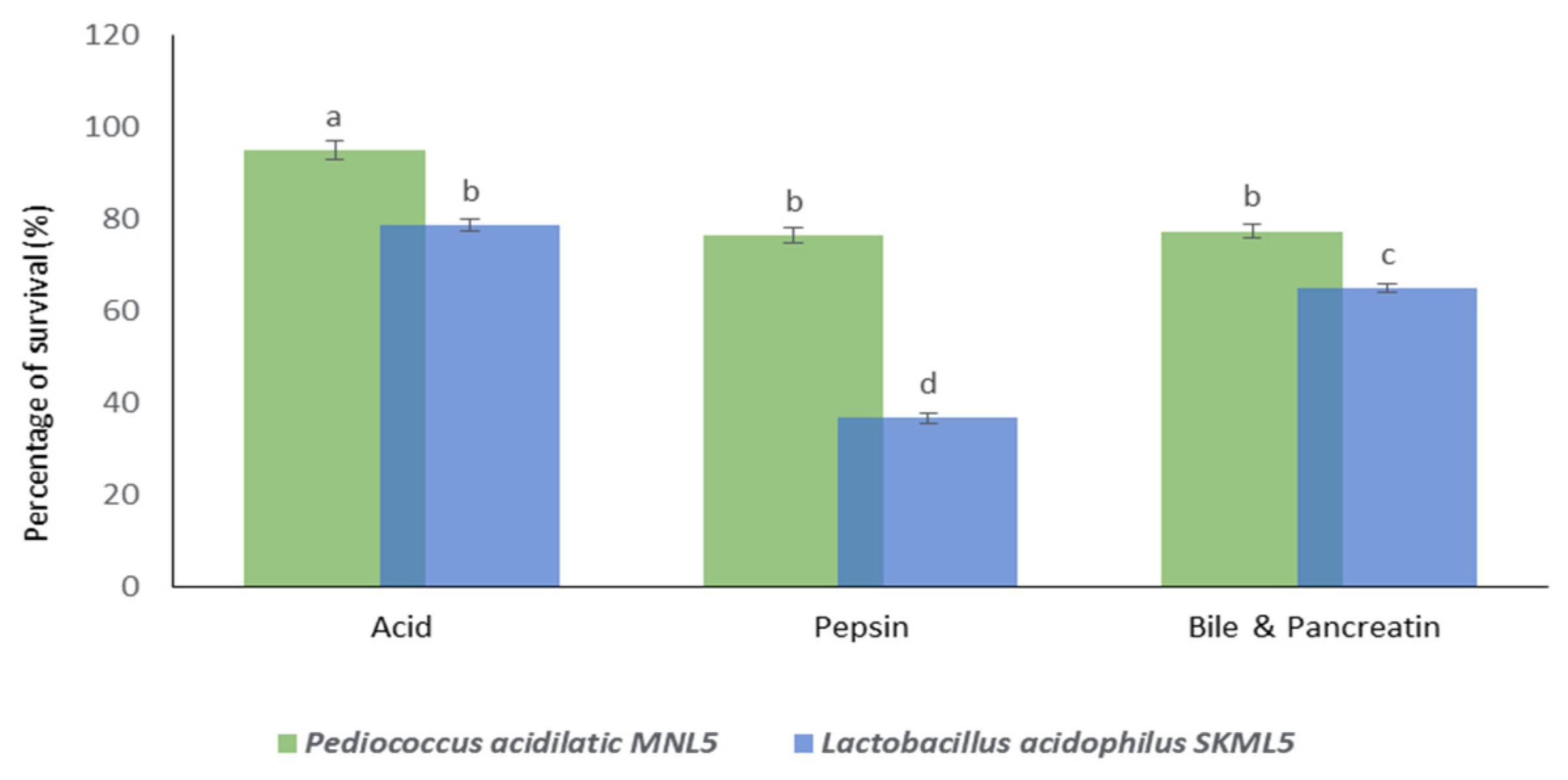
2.5. Survival under In Vitro Gastrointestinal Conditions
Tolerance to Low pH, Pepsin and Bile Salts and Pancreatin
2.6. Chemotaxis Assays
2.7. Worm Size Measurement
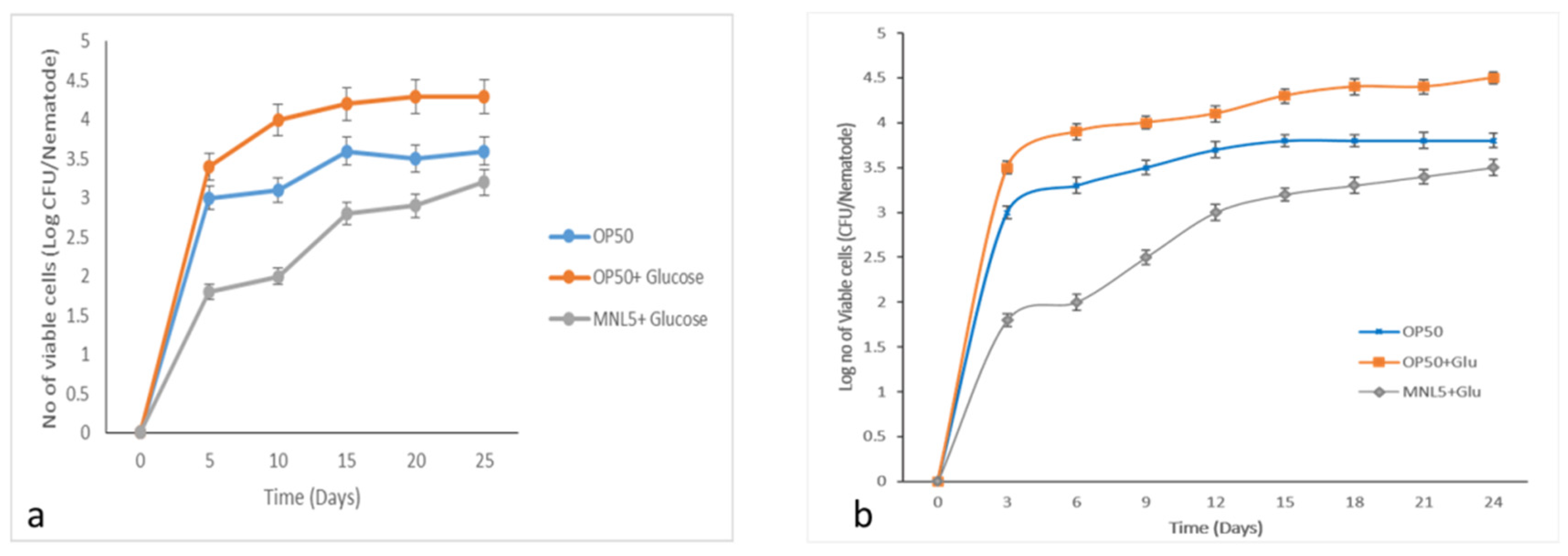
2.8. C. elegans Gut Colonization Ability
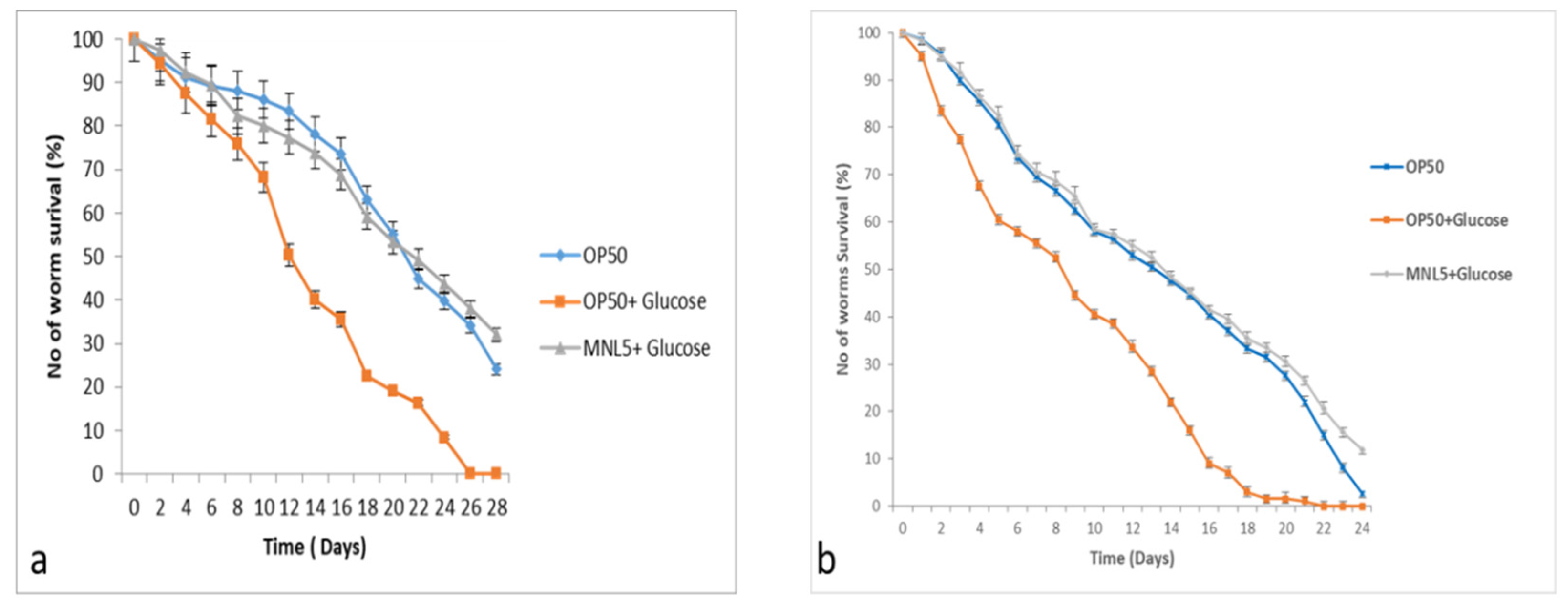
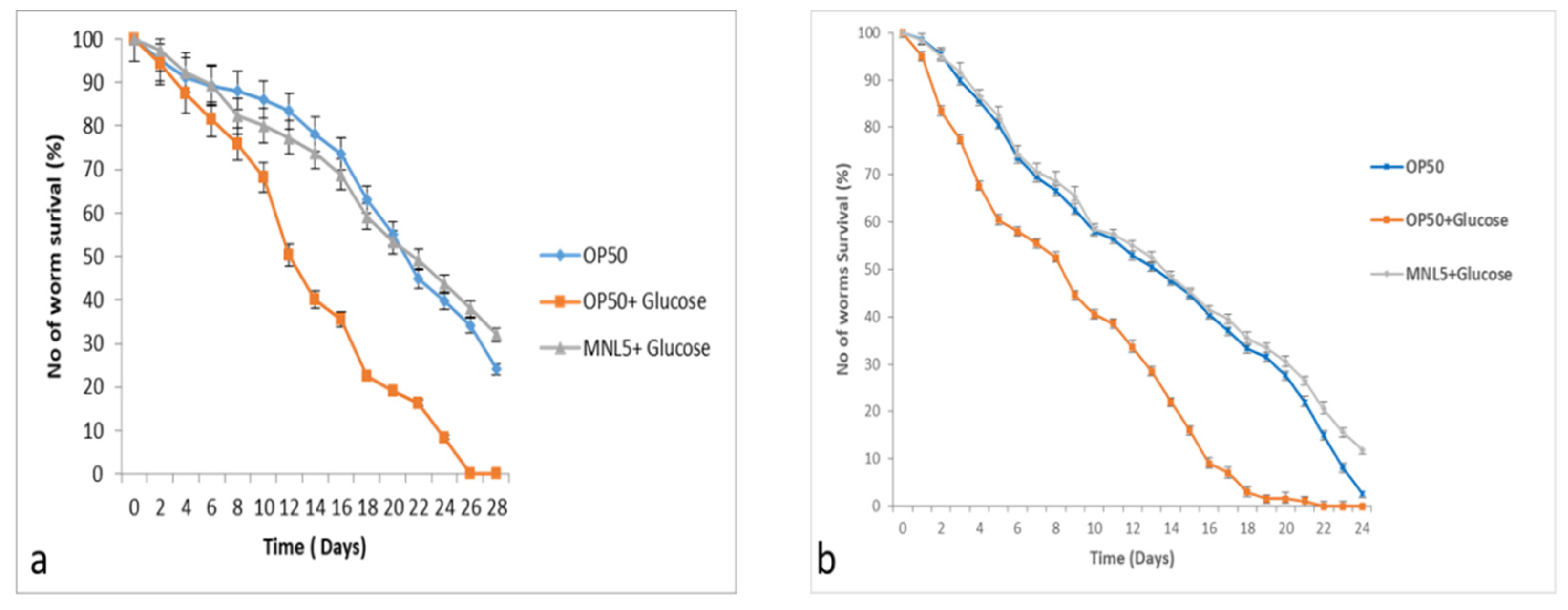
2.9. C. elegans Life Span Assay
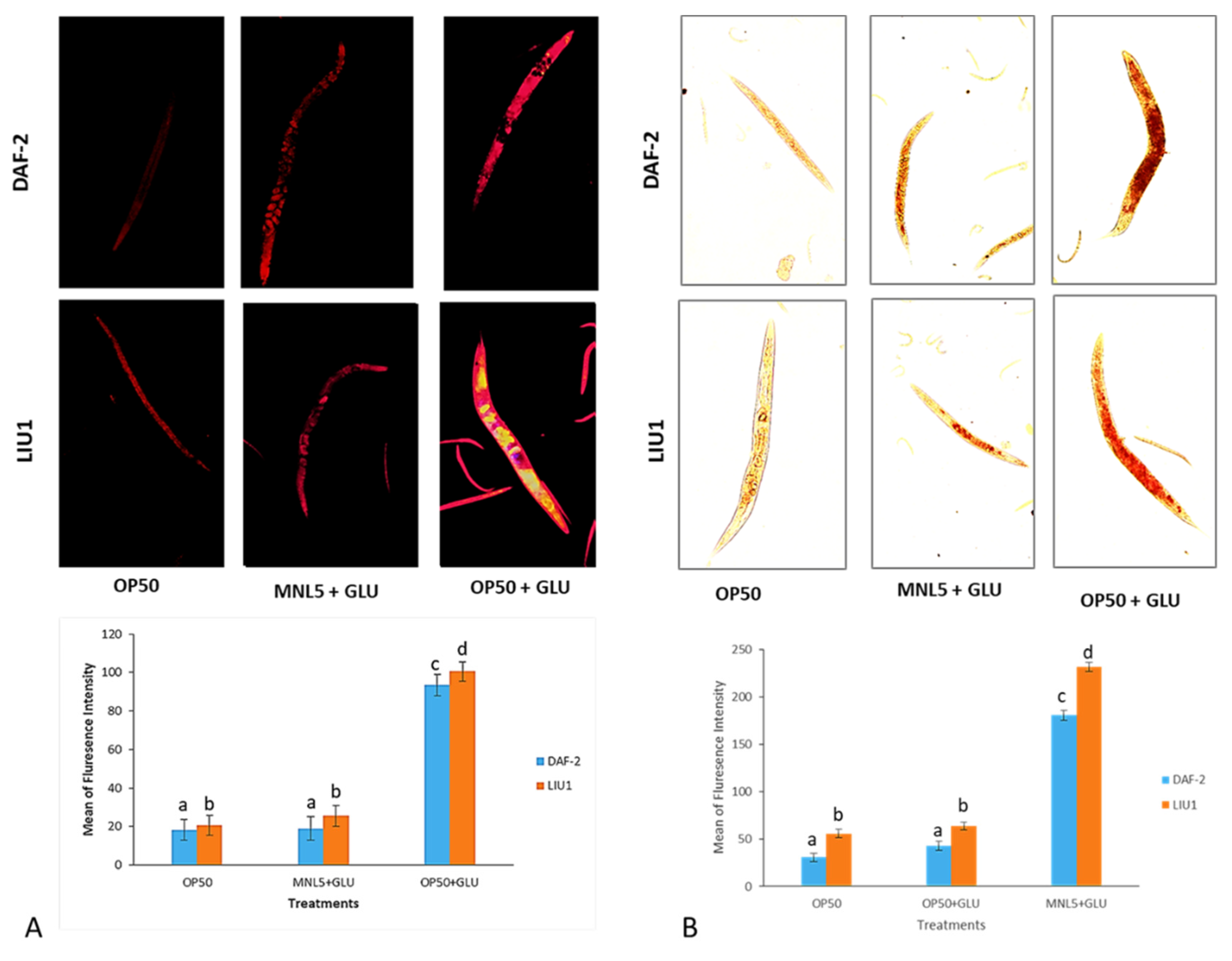
2.10. Lipid Determination by Microscope Analysis
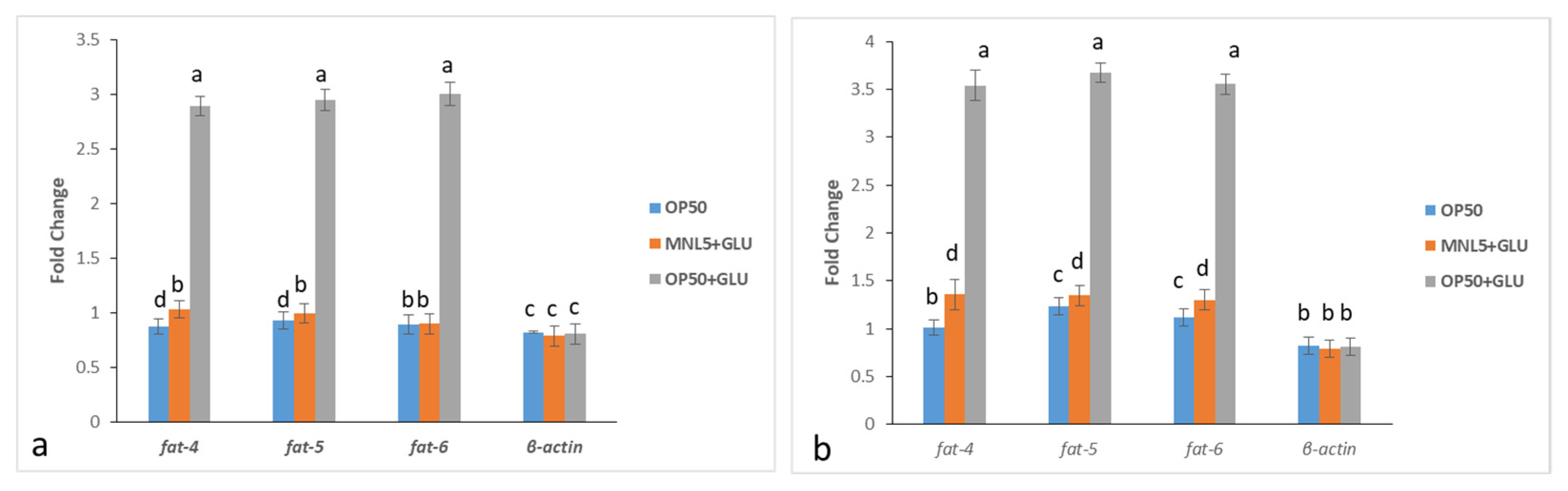
2.11. Transcriptomic Analysis in C. elegans
3. Discussion
4. Materials and Methods
4.1. Isolation and Characterization of Cholesterol Reducing LAB
4.2. BSH Activityand Cholesterol Assimilation
4.3. Molecular Identification of LAB
4.4. Safety Aspect of LAB
4.4.1. Antibiotic Susceptibility
4.4.2. Antibacterial Activity
4.4.3. Hemolytic Activity
4.5. Gastrointestinal Survival under In Vitro Conditions
4.5.1. Tolerance to Low pH
4.5.2. Tolerance to Pepsin
4.5.3. Resistance of Pancreatin
4.6. In Vivo Studies
4.6.1. Maintain the C. elegans
4.6.2. C. elegans Synchronizing
4.6.3. Chemotaxis Assay
4.6.4. Establishment of Glucose Diet with LAB
4.6.5. Bacterial Colonization Assay
4.6.6. Lifespan Assay
4.6.7. Body Size Measurements
4.6.8. Nile Red and Oil Red O (ORO) Staining Methods
4.6.9. Fluorescence Quantification
4.6.10. RNA Extraction and Primer Deign
4.6.11. Quantitative PCR Analyses
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization. The State of Food and Agriculture 2001. No. 33; Food and Agriculture Organization: Quebec City, QC, Canada, 2001. [Google Scholar]
- Fasoli, S.; Marzotto, M.; Rizzotti, L.; Rossi, F.; Dellaglio, F.; Torriani, S. Bacterial composition of commercial probiotic products as evaluated by PCR-DGGE analysis. Int. J. Food Microbiol. 2003, 82, 59–70. [Google Scholar] [CrossRef]
- Chelliah, R.; Choi, J.G.; Hwang, S.B.; Park, B.J.; Daliri, E.B.M.; Kim, S.H.; Wei, S.; Ramakrishnan, S.R.; Oh, D.H. In vitro and in vivo defensive effect of probiotic LAB against Pseudomonas aeruginosa using Caenorhabditis elegans model. Virulence 2018, 9, 1489–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khusro, A.; Aarti, C.; Dusthackeer, A.; Agastian, P. Anti-tubercular and probiotic properties of coagulase-negative staphylococci isolated from Koozh, a traditional fermented food of South India. Microb. Pathog. 2018, 114, 239–250. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Shi, J.; Zhu, J.; Shao, D.; Huang, Q.; Yang, H.; Jin, M. Capacity of lactic acid bacteria in immunity enhancement and cancer prevention. Appl. Microbiol. Biotechnol. 2017, 101, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.A.; Chang, H.C. Cholesterol-lowering effects of a putative probiotic strain Lactobacillus plantarum EM isolated from kimchi. LWT—Food Sci. Technol. 2015, 62, 210–217. [Google Scholar] [CrossRef]
- Miremadi, F.; Ayyash, M.; Sherkat, F.; Stojanovska, L. Cholesterol reduction mechanisms and fatty acid composition of cellular membranes of probiotic Lactobacilli and Bifidobacteria. J. Funct. Foods 2014, 9, 295–305. [Google Scholar] [CrossRef]
- Pinto, M.G.V.; Franz, C.M.; Schillinger, U.; Holzapfel, W.H. Lactobacillus spp. within vitro probiotic properties from human faeces and traditional fermented products. Int. J. Food Microbiol. 2006, 109, 205–214. [Google Scholar] [CrossRef]
- Heo, W.; Lee, E.S.; Cho, H.T.; Kim, J.H.; Lee, J.H.; Yoon, S.M.; Kim, Y.J. Lactobacillus plantarum LRCC 5273 isolated from Kimchi ameliorates diet-induced hypercholesterolemia in C57BL/6 mice. Biosci. Biotechnol. Biochem. 2018, 82, 1964–1972. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.C.; Chan, H.Y.; Wann, S.Y.; Lin, F.M.; Lee, F.L.; Liao, C.C. Isolated Pediococcus Acidilactici 05b0111 and Method of Producing Exopolysaccharide. U.S. Patent US9873899B2, 19 November 2015. [Google Scholar]
- Pingitore, E.; Jimenez, M.E.; Dallagnol, A.; Belfiore, C.; Fontana, C.; Fontana, P.; Plumed-Ferrer, C. Screening and characterization of potential probiotic and starter bacteria for plant fermentations. LWT—Food Sci. Technol. 2016, 71, 288–294. [Google Scholar] [CrossRef]
- Ortu, S.; Felis, G.E.; Marzotto, M.; Deriu, A.; Molicotti, P.; Sechi, L.A.; Zanetti, S. Identification and functional characterization of Lactobacillus strains isolated from milk and Gioddu, a traditional Sardinian fermented milk. Int. Dairy J. 2017, 35, 1312–1320. [Google Scholar] [CrossRef]
- Ashrafi, K. Obesity and the regulation of fat metabolism. WormBook Online Rev. C Elegans Biol. 2007, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tan, X.R.; Li, S.J.; Zhang, X.X. LncRNA NEAT1 promotes hepatic lipid accumulation via regulating miR-146a-5p/ROCK1 in nonalcoholic fatty liver disease. Life Sci. 2019, 235, 116829. [Google Scholar] [CrossRef] [PubMed]
- Belguesmia, Y.; Alard, J.; Mendil, R.; Ravallec, R.; Grangette, C.; Drider, D.; Cudennec, B. In vitro probiotic properties of selected lactobacilli and multi-strain consortium on immune function gut barrier strengthening and gut hormone secretion. J. Funct. Foods 2019, 57, 382–391. [Google Scholar] [CrossRef]
- World Health Organization. Report of a Joint FAO/OIE/WHO Expert Consultation on Antimicrobial Use in Aquaculture and Antimicrobial Resistance, Seoul, Korea, 13–16 June.2006; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Bhagat, D.; Raina, N.; Kumar, A.; Katoch, M.; Khajuria, Y.; Slathia, P.S.; Sharma, P. Probiotic properties of a phytase producing Pediococcus acidilactici strain SMVDUDB2 isolated from traditional fermented cheese product, Kalarei. Sci. Rep. 2020, 10, 1926. [Google Scholar] [CrossRef]
- Bartkiene, E.; Mozuriene, E.; Lele, V.; Zokaityte, E.; Gruzauskas, R.; Jakobsone, I.; Bartkevics, V. Changes of bioactive compounds in barley industry by-products during submerged and solid state fermentation with antimicrobial Pediococcus acidilactici strain LUHS29. Food Sci. Nutr. 2020, 8, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Noda, M.; Sugiyama, M.; Ahmad, A.; Kaur, B. Production of Functional Buttermilk and Soymilk Using Pediococcus acidilactici BD16 (alaD+). Molecules 2021, 26, 4671. [Google Scholar] [CrossRef]
- Albano, H.; Todorov, S.D.; van Reenen, C.A.; Hogg, T.; Dicks, L.M.; Teixeira, P. Characterization of two bacteriocins produced by Pediococcus acidilactici isolated from “Alheira”, a fermented sausage traditionally produced in Portugal. Int. J. Food Microbiol. 2007, 116, 239–247. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Martín, A.; Benito, M.J.; Hernández, A.; Casquete, R.; de Guia Córdoba, M. Application of Lactobacillus fermentum HL57 and Pediococcus acidilactici SP979 as potential probiotics in the manufacture of traditional Iberian dry-fermented sausages. Food Microbiol. 2011, 28, 839–847. [Google Scholar] [CrossRef]
- Rahmdel, S.; Shekarforoush, S.S.; Hosseinzadeh, S.; Torriani, S.; Gatto, V. Antimicrobial spectrum activity of bacteriocinogenic Staphylococcus strains isolated from goat and sheep milk. J. Dairy Sci. 2019, 102, 2928–2940. [Google Scholar] [CrossRef] [Green Version]
- Barathikannan, K.; Chelliah, R.; Rubab, M.; Daliri, E.B.-M.; Elahi, F.; Kim, D.-H.; Agastian, P.; Oh, S.-Y.; Oh, D.H. Gut Microbiome Modulation Based on Probiotic Application for Anti-Obesity: A Review on Efficacy and Validation. Microorganisms 2019, 7, 456. [Google Scholar] [CrossRef] [Green Version]
- Khalesi, S.; Bellissimo, N.; Vandelanotte, C.; Williams, S.; Stanley, D.; Irwin, C. A review of probiotic supplementation in healthy adults: Helpful or hype? Eur. J. Clin. Nutr. 2019, 73, 24. [Google Scholar] [CrossRef] [PubMed]
- Park, D.Y.; Ahn, Y.T.; Park, S.H.; Huh, C.S.; Yoo, S.R.; Yu, R.; Sung, M.K.; McGregor, R.A.; Choi, M.S. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PLoS ONE 2013, 8, e59470. [Google Scholar] [CrossRef] [Green Version]
- Irazoqui, J.E.; Urbach, J.M.; Ausubel, F.M. Evolution of host innate defence: Insights from C. elegans and primitive invertebrates. Nat. Rev. Immunol. 2010, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skora, S.; Mende, F.; Zimmer, M. Energy scarcity promotes a brain-wide sleep state modulated by insulin signaling in C. elegans. Cell Rep. 2018, 22, 953–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revtovich, A.V.; Lee, R.; Kirienko, N.V. Interplay between mitochondria and diet mediates pathogen and stress resistance in Caenorhabditis elegans. PLoS Genet. 2019, 15, e1008011. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, L.; Zhang, L.; Wang, W.; Wei, S.; Wang, J.; Che, H.; Zhang, Y. Effects of sugars and lipids on the growth and development of Caenorhabditis elegans. BioRxiv 2019, 615807. [Google Scholar] [CrossRef]
- Martorell, P.; Llopis, S.; Gonzalez, N.; Chenoll, E.; Lopez-Carreras, N.; Aleixandre, A.; Genovés, S. Probiotic strain Bifidobacterium animalis subsp. lactis CECT 8145 reduces fat content and modulates lipid metabolism and antioxidant response in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 3462–3472. [Google Scholar]
- Jones, K.T.; Ashrafi, K. Caenorhabditis elegans as an emerging model for studying the basic biology of obesity. Dis. Models Mech. 2009, 2, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Chambers, D.B.; Androschuk, A.; Rosenfelt, C.; Langer, S.; Harding, M.; Bolduc, F.V. Insulin signaling is acutely required for long-term memory in Drosophila. Front. Neural Circuits 2015, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.L.; Lum, K.M.; Lara-Gonzalez, P.; Ogasawara, D.; Cognetta, A.B.; To, A.; Parsons, W.H.; Simon, G.M.; Desai, A.; Petrascheck, M.; et al. Pharmacological convergence reveals a lipid pathway that regulates C. elegans lifespan. Nat. Chem. Biol. 2019, 15, 453–462. [Google Scholar] [CrossRef]
- Aranaz, P.; Navarro-Herrera, D.; Zabala, M.; Romo-Hualde, A.; López-Yoldi, M.; Vizmanos, J.L.; Milagro, F.I.; González-Navarro, C.J. Phenolic compounds reduce the fat content in Caenorhabditis elegans by affecting lipogenesis, lipolysis, and different stress responses. Pharmaceuticals 2020, 13, 355. [Google Scholar] [CrossRef] [PubMed]
- Wahlby, C.; Lee Conery, A.; Bray, M.A.; Kamentsky, L.; Larkins-Ford, J.; Sokolnicki, K.L.; Veneskey, M.; Michaels, K.; Carpenter, A.E.; O’Rourke, E.J. High- and low-throughput scoring of fat mass and body fat distribution in C. elegans. Methods 2014, 68, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannock, G.W.; Dashkevicz, M.P.; Feighner, S.D. Lactobacilli and bile salt hydrolase in the murine intestinal tract. Appl. Environ. Microbiol. 1989, 55, 1848–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudel, L.L.; Morris, M.D. Determination of cholesterol using o-phthalaldehyde. J. Lipid Res. 1973, 14, 364–366. [Google Scholar] [CrossRef]
- Das, P.; Mukherjee, S.; Sen, R. Antimicrobial potential of a lipopeptide biosurfactant derived from a marine Bacillus circulans. J. Appl. Microbiol. 2008, 104, 1675–1684. [Google Scholar] [CrossRef]
- Aslam, S.N.; Stevenson, P.C.; Kokubun, T.; Hall, D.R. Antibacterial and antifungal activity of cicerfuran and related 2-arylbenzofurans and stilbenes. Microbiol. Res. 2009, 164, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Manhar, A.K.; Bashir, Y.; Saikia, D.; Nath, D.; Gupta, K.; Konwar, B.K.; Kumar, R.; Namsa, N.D.; Mandal, M. Cellulolytic potential of probiotic Bacillus Subtilis AMS6 isolated from traditional fermented soybean (Churpi): An in-vitro study with regards to application as an animal feed additive. Microbiol. Res. 2016, 186, 62–70. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [Green Version]
- Oh, A.; Daliri, E.B.M.; Oh, D.H. Screening for potential probiotic bacteria from Korean fermented soybean paste: In vitro and Caenorhabditis elegans model testing. LWT—Food Sci. Technol. 2018, 88, 132–138. [Google Scholar] [CrossRef]
- Wong, A.; Boutis, P.; Hekimi, S. Mutations in the clk-1 gene of Caenorhabditis elegans affect developmental and behavioral timing. Genetics 1995, 139, 1247–1259. [Google Scholar] [CrossRef]
- Margie, O.; Palmer, C.; Chin-Sang, I.C. C. elegans chemotaxis assay. J. Vis. Exp. 2013, 74, 50069. [Google Scholar]
- Antunes, C.A.; Clark, L.; Wanuske, M.T.; Hacker, E.; Ott, L.; Simpson-Louredo, L.; de Luna, M.D.G.; Hirata, R., Jr.; Mattos-Guaraldi, A.L.; Hodgkin, J.; et al. Caenorhabditis elegans star formation and negative chemotaxis induced by infection with corynebacteria. Microbiology 2016, 162, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Bito, T.; Matsunaga, Y.; Yabuta, Y.; Kawano, T.; Watanabe, F. Vitamin B12 deficiency in Caenorhabditis elegans results in loss of fertility, extended life cycle, and reduced lifespan. FEBS Open Bio 2013, 3, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolentseva, O.; Gusarov, I.; Gautier, L.; Shamovsky, I.; DeFrancesco, A.S.; Losick, R.; Nudler, E. Mechanism of biofilm-mediated stress resistance and lifespan extension in C. elegans. Sci. Rep. 2017, 7, 7137. [Google Scholar] [CrossRef] [PubMed]
- Barathikannan, K.; Venkatadri, B.; Khusro, A.; Al-Dhabi, N.A.; Agastian, P.; Arasu, M.V.; Kim, Y.O. Chemical analysis of Punica granatum fruit peel and it’s in vitro and in vivo biological properties. BMC Complementary Altern. Med. 2016, 16, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelliah, R.; Wei, S.; Park, B.J.; Kim, S.H.; Park, D.S.; Kim, S.H.; Hwan, K.S.; Oh, D.H. Novel motB as a potential predictive tool for identification of B. cereus, B. thuringiensis and differentiation from other Bacillus species by triplex real-time PCR. Microb. Pathog. 2017, 111, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Strain Name | Colour Morphology | Shape | Hemolytic Activity | BSH Activity | Cholesterol Assimilation (%) |
|---|---|---|---|---|---|---|
| 1 | MNL 1 | White/Smooth | cocci | NA | ++ | 25.3 ± 0.54 g |
| 2 | MNL 2 | White | rod | NA | + | 18.5 ± 0.41 h |
| 3 | MNL 3 | White/Round | rod | NA | - | 10.2 ± 0.39 h |
| 4 | MNL 4 | White/Smooth/Round | rod | NA | - | 11.7 ± 0.24 h |
| 5 | MNL 5 | White/Smooth | cocci | NA | +++ | 89.5 ± 1.09 a |
| 6 | MNL 6 | White | rod | NA | - | 25.5 ± 0.44 g |
| 7 | MNL 8 | White | cocci | NA | - | 23.4 ± 0.38 g |
| 8 | MNL 9 | White | cocci | NA | + | 31.5 ± 0.61 f |
| 9 | MNL 10 | White/Round | rod | NA | + | 24.5 ± 0.50 g |
| 10 | MNL 11 | White/Smooth/Round | short rod | NA | ++ | 47.5 ± 0.36 e |
| 11 | GSC 5 | White/Smooth | rod | NA | + | 31.0 ± 0.37 f |
| 12 | GSC 8 | White | rod | NA | - | 12.4 ± 0.47 h |
| 13 | GSC 10 | White | short rod | NA | + | 10.5 ± 0.39 h |
| 14 | TVL 11 | White/Smooth | short rod | NA | + | 14.6 ± 0.40 h |
| 15 | TVL 12 | White | short rod | NA | - | 12.4 ± 0.55 h |
| 16 | SKML 3 | White/Round | rod | NA | + | 22.5 ± 0.51 g |
| 17 | SKML 4 | White | rod | NA | - | 9.5 ± 0.64 h |
| 18 | SKML 5 | White/Smooth | cocci | NA | +++ | 77.1 ± 0.98 b |
| 19 | SKML 6 | White/Smooth | rod | NA | ++ | 69.5 ± 0.78 c |
| 20 | SKML 8 | White/Smooth/Irregular | rod | NA | + | 53.43 ± 0.63 d |
| 21 | SKML 10 | White | cocci | NA | + | 47.56 ± 0.51 e |
| 22 | SKML 11 | White | short rod | NA | + | 45.21 ± 0.69 e |
| 23 | SKML 12 | White | cocci | NA | - | 25.80 ± 0.40 g |
| 24 | SKML 13 | White | short rod | NA | - | 22.4 ± 0.35 g |
| 25 | SK SPL 1 | White/Rough | short rod | NA | - | 21.4 ± 0.44 g |
| 26 | SK SPL 6 | White | cocci | NA | ++ | 57.4 ± 0.64 a |
| 27 | SK SPL7 | White/Smooth | cocci | NA | + | 42.5 ± 0.51 e |
| 28 | GSC 1 | White | cocci | NA | + | 38.4 ± 0.38 f |
| 29 | GSC 11 | White | short rod | NA | + | 32.5 ± 0.33 f |
| 30 | GSC 12 | White | cocci | NA | ++ | 41.5 ± 0.46 e |
| 31 | SKML 18 | White | short rod | NA | ++ | 57.8 ± 0.48 d |
| 32 | SKML 20 | White/Smooth | cocci | NA | ++ | 68.6 ± 0.64 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barathikannan, K.; Chelliah, R.; Elahi, F.; Tyagi, A.; Selvakumar, V.; Agastian, P.; Valan Arasu, M.; Oh, D.-H. Anti-Obesity Efficacy of Pediococcus acidilactici MNL5 in Canorhabditis elegans Gut Model. Int. J. Mol. Sci. 2022, 23, 1276. https://doi.org/10.3390/ijms23031276
Barathikannan K, Chelliah R, Elahi F, Tyagi A, Selvakumar V, Agastian P, Valan Arasu M, Oh D-H. Anti-Obesity Efficacy of Pediococcus acidilactici MNL5 in Canorhabditis elegans Gut Model. International Journal of Molecular Sciences. 2022; 23(3):1276. https://doi.org/10.3390/ijms23031276
Chicago/Turabian StyleBarathikannan, Kaliyan, Ramachandran Chelliah, Fazle Elahi, Akanksha Tyagi, Vijayalakshmi Selvakumar, Paul Agastian, Mariadhas Valan Arasu, and Deog-Hawn Oh. 2022. "Anti-Obesity Efficacy of Pediococcus acidilactici MNL5 in Canorhabditis elegans Gut Model" International Journal of Molecular Sciences 23, no. 3: 1276. https://doi.org/10.3390/ijms23031276
APA StyleBarathikannan, K., Chelliah, R., Elahi, F., Tyagi, A., Selvakumar, V., Agastian, P., Valan Arasu, M., & Oh, D.-H. (2022). Anti-Obesity Efficacy of Pediococcus acidilactici MNL5 in Canorhabditis elegans Gut Model. International Journal of Molecular Sciences, 23(3), 1276. https://doi.org/10.3390/ijms23031276