
Comprehensive Profiling of Paper Mulberry (Broussonetia papyrifera) Crotonylome Reveals the Significance of Lysine Crotonylation in Young Leaves

Abstract
:1. Introduction
2. Results and Discussion
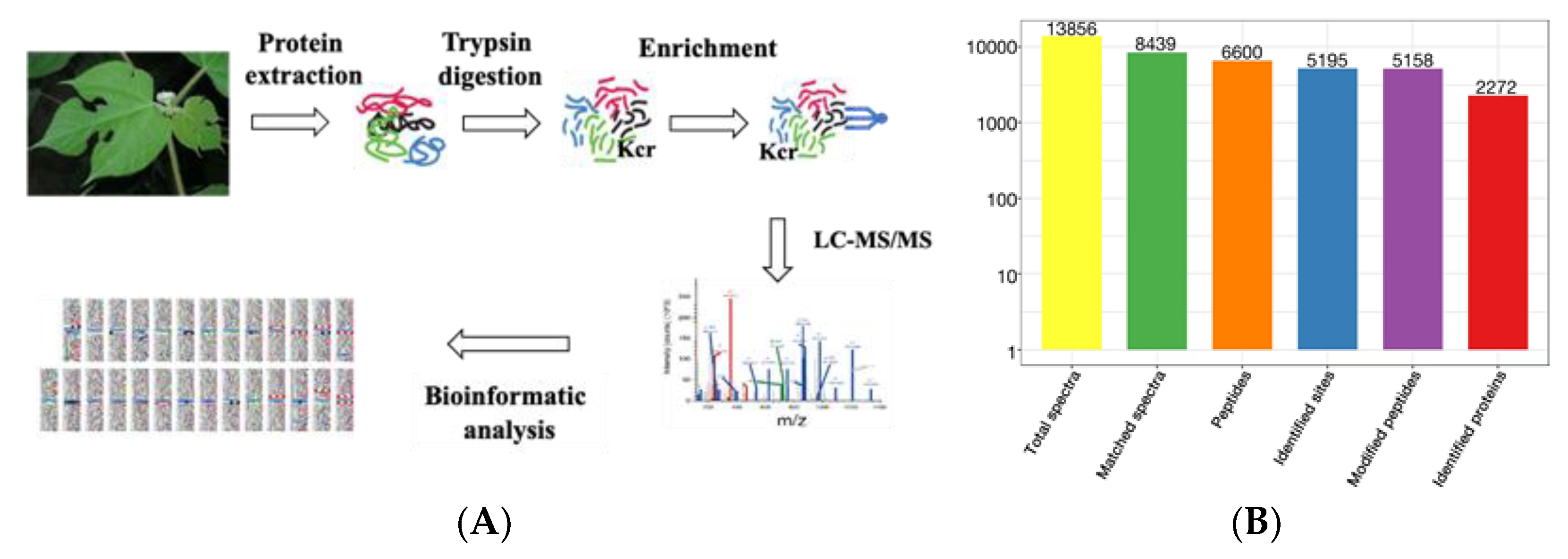
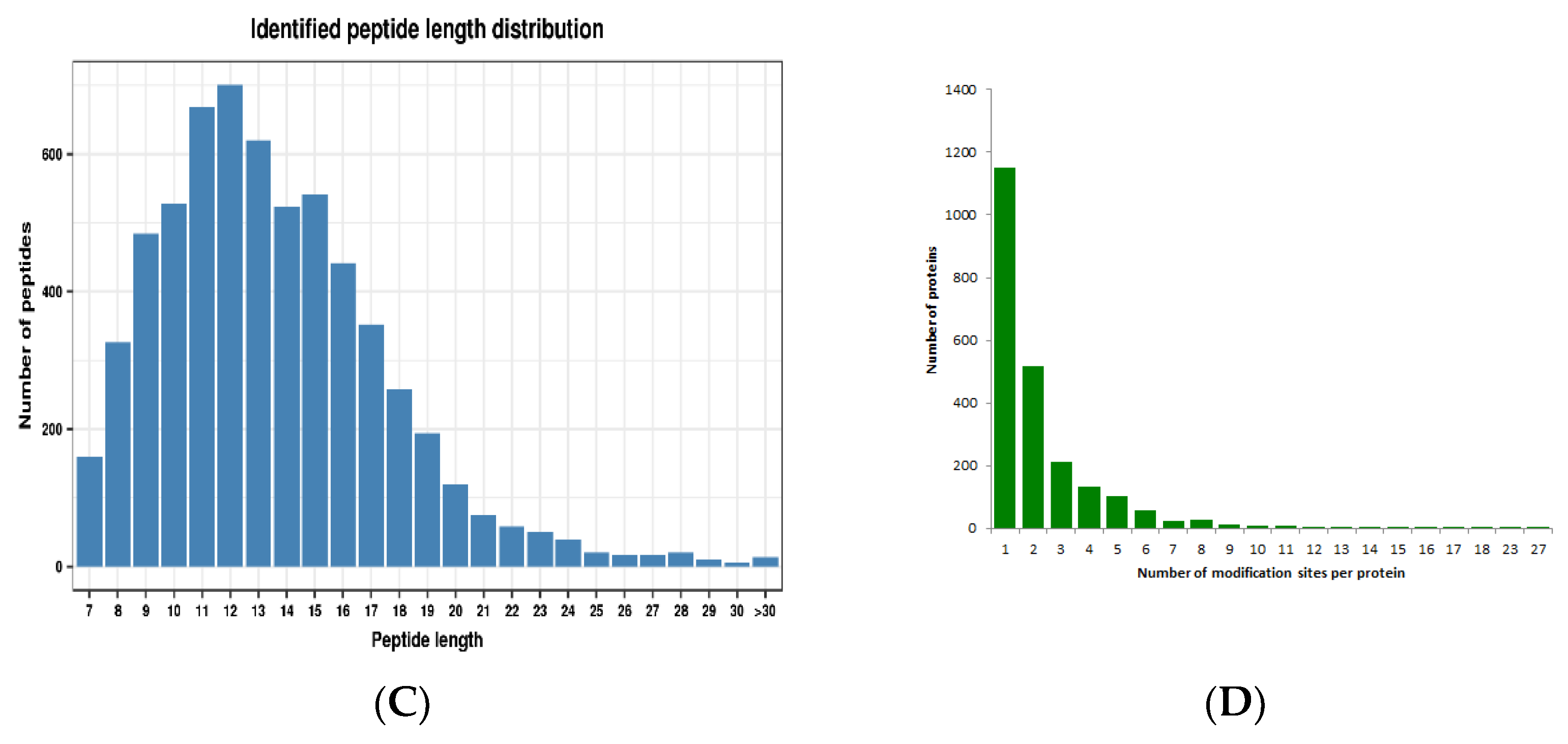
2.1. Profiling the Crotonylsites and Crotonylproteins on Paper Mulberry Young Leaf Tissues
2.2. Analysis of Crotonylated Lysine Motifs
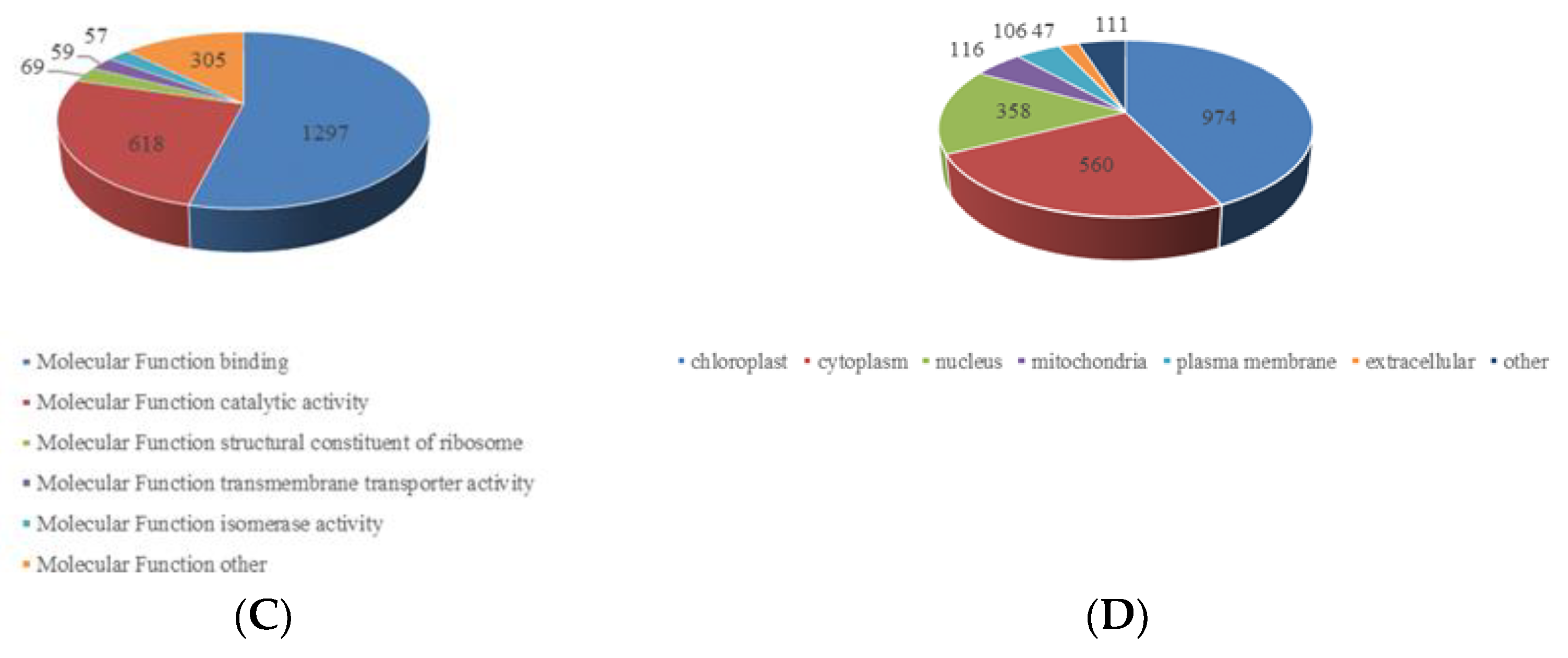
2.3. Functional Characterization of Lysine Crotonylome
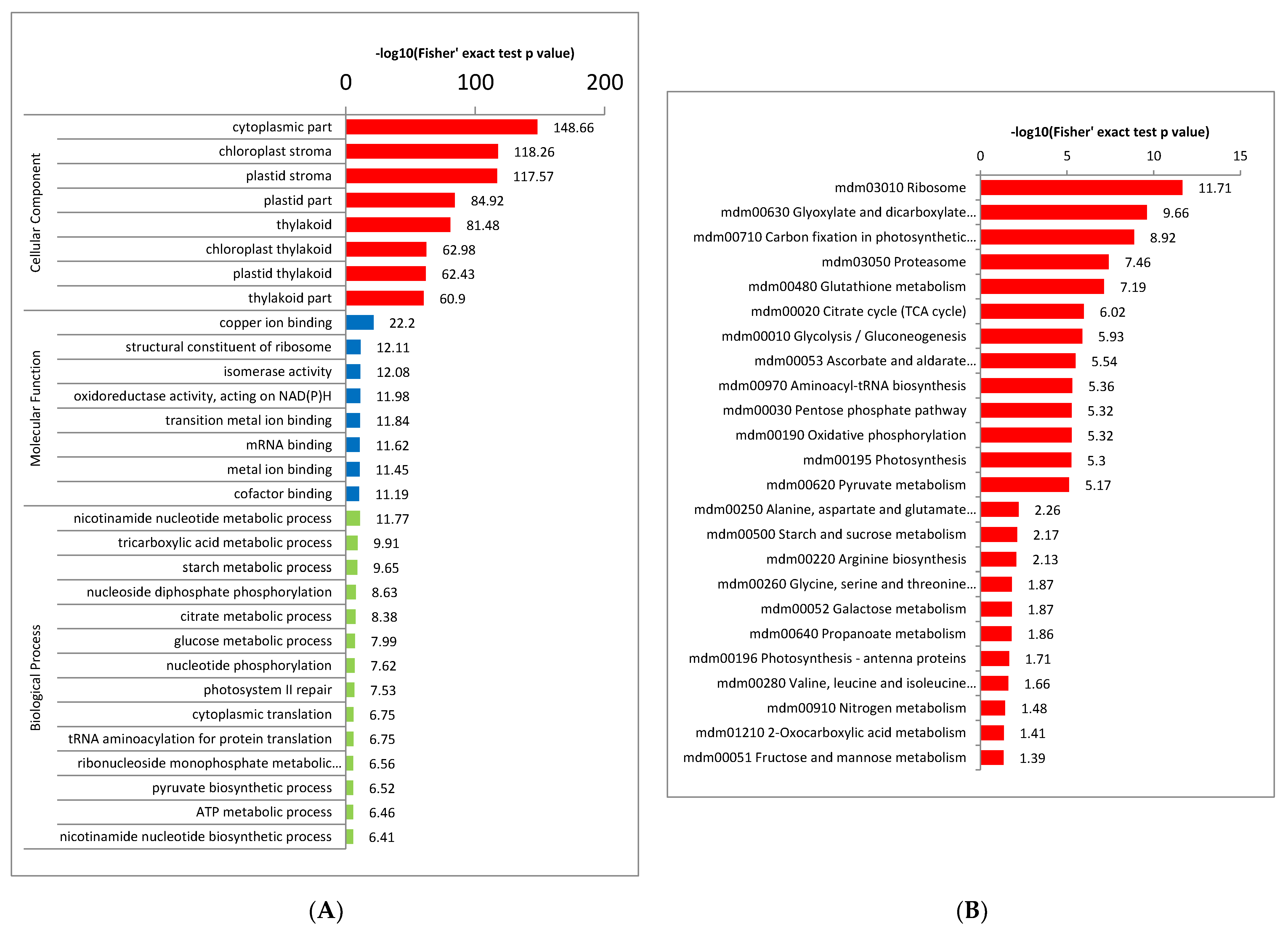
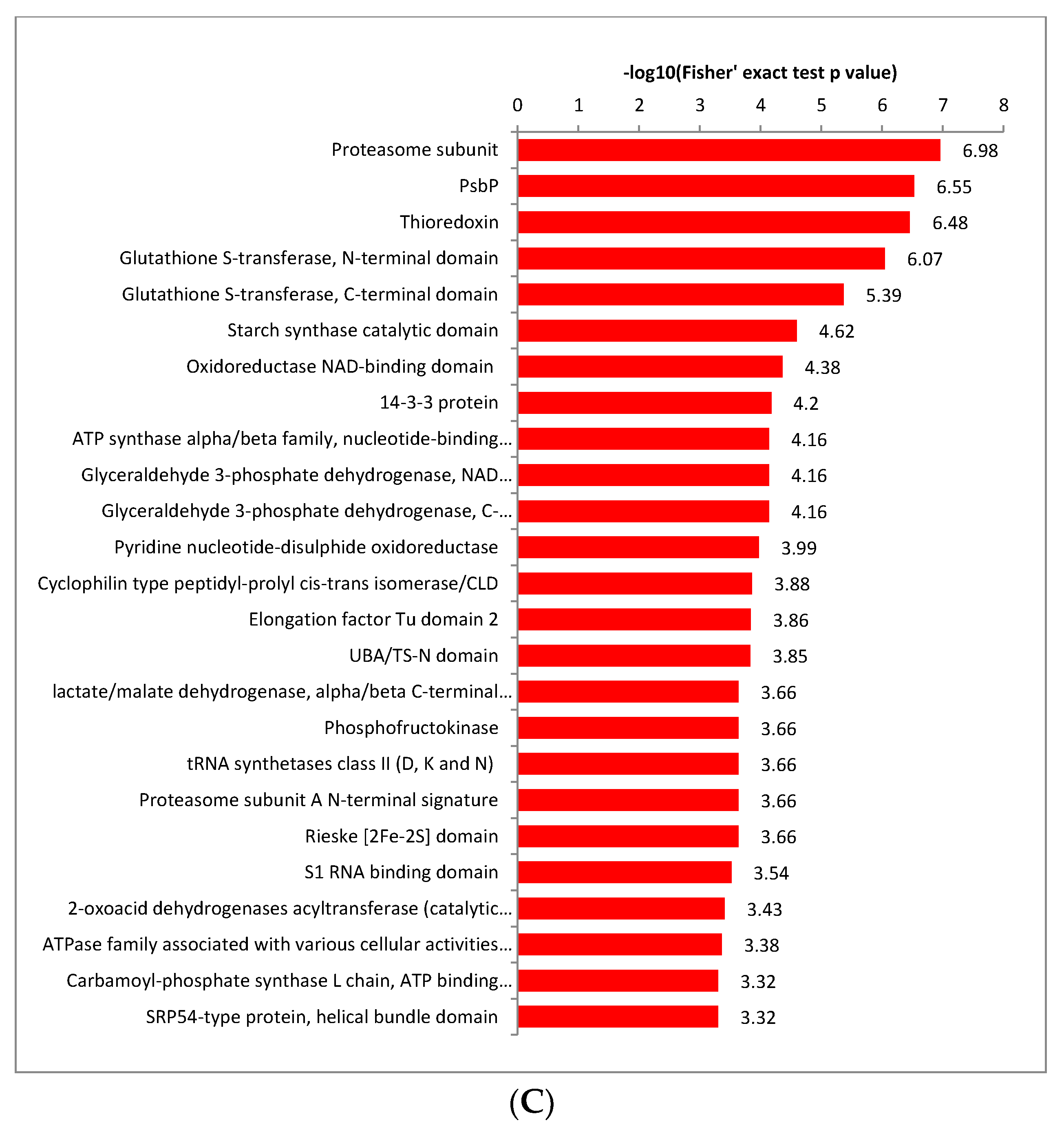
2.4. Functional Enrichment Analysis of Crotonylated Proteins
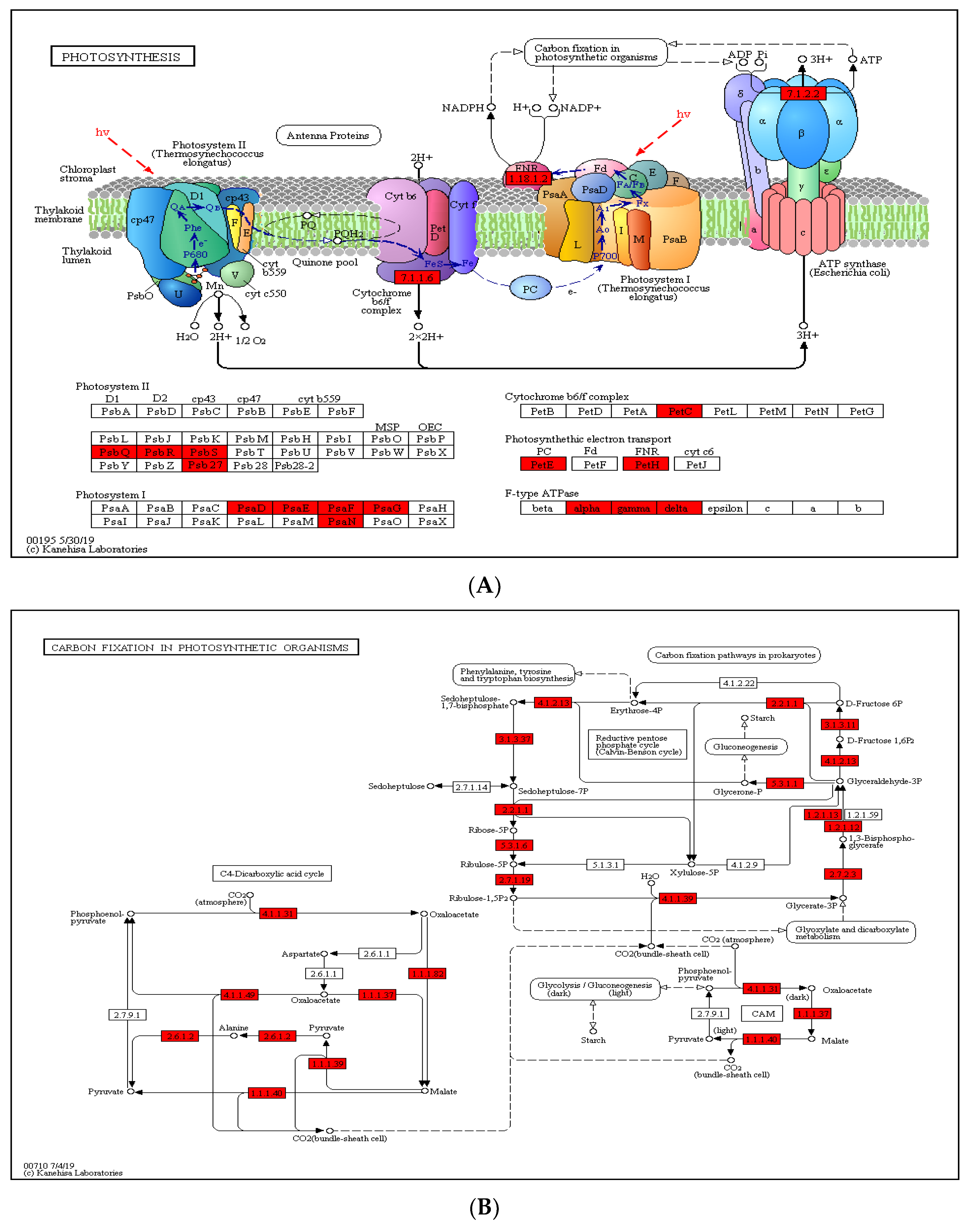
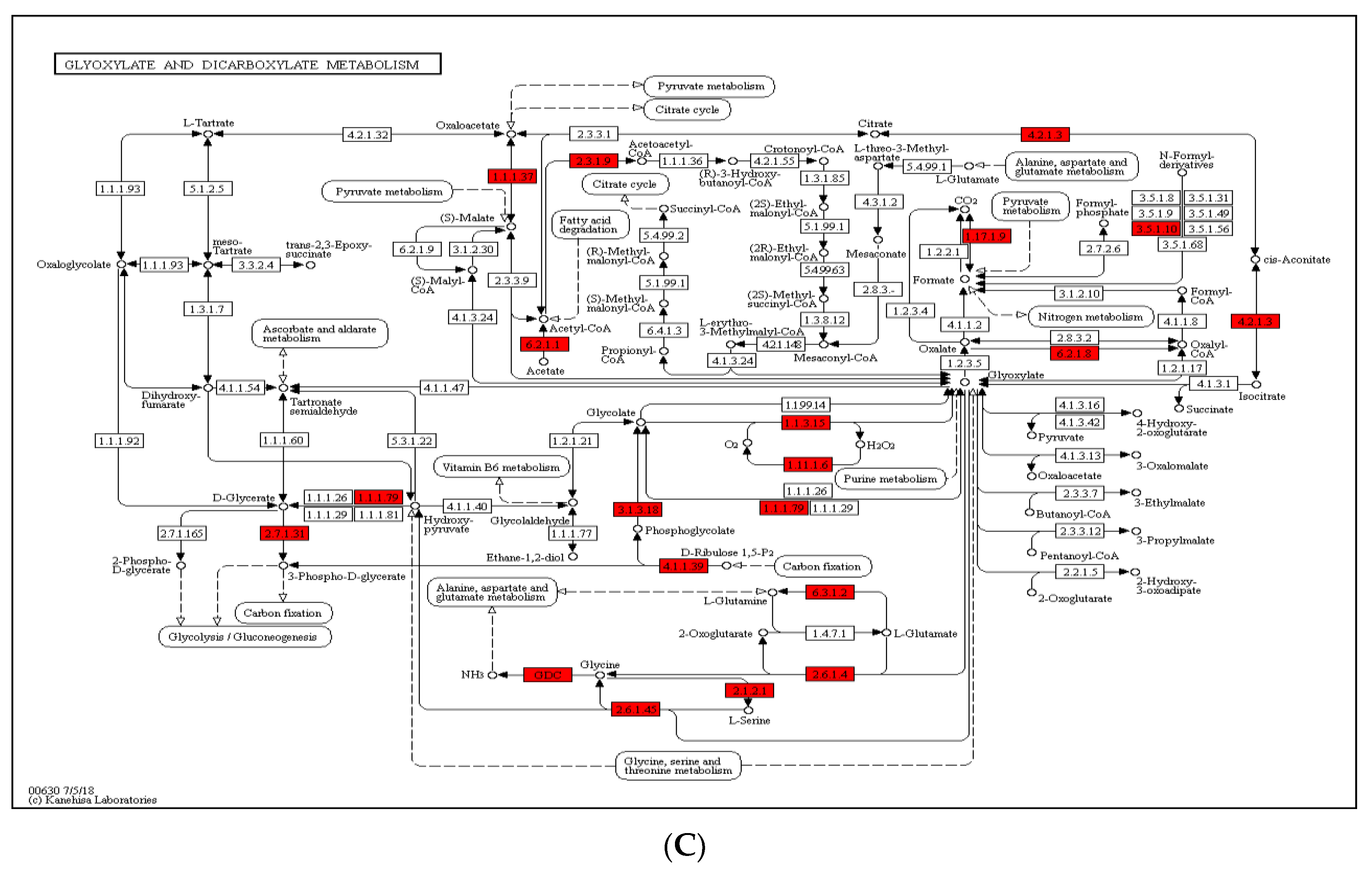
2.5. Crotonylated Proteins Involved in Photosynthesis and Carbon Fixation
2.6. Interactive Networks among Crotonylated Proteins in Paper Mulberry
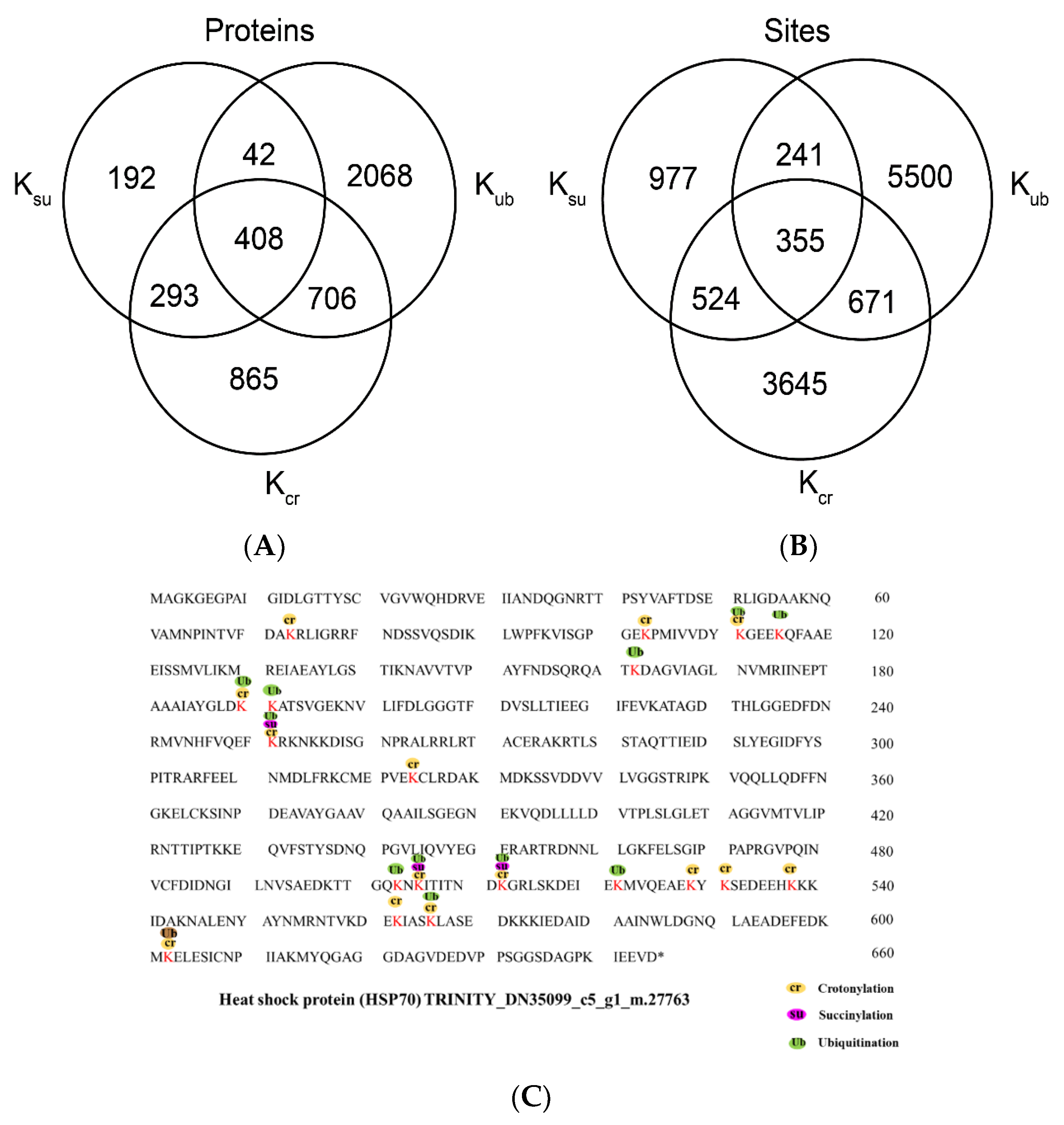
2.7. Overlap between Lysine Crotonylation, Succinylation and Ubiquitylation in Paper Mulberry
3. Materials and Methods
3.1. Collection and Preparation of Plant Materials
3.2. Protein Extraction
3.3. Trypsin Digestion
3.4. Panantibody-Based PTM Enrichment
3.5. LC-MS/MS Analysis
3.6. Database Search
3.7. Bioinformatics Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Zhang, Z.; Tan, M.; Xie, Z.; Dai, L.; Chen, Y.; Zhao, Y. Identification of lysine succinylation as a new post-translational modi-fication. Nat. Chem. Biol. 2011, 7, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Christopher, T.; Tsodikova, S.; Gregory, J. Protein Posttranslational Modifications: The Chemistry of Proteome Diversifications. Angew. Chem. Int. Ed. 2005, 44, 7342–7372. [Google Scholar]
- Martin, C.; Yi, Z. Mechanisms of epigenetic inheritance. Curr. Opin. Cell Biol. 2007, 19, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Ruthenburg, A.J.; Li, H.; Patel, D.J.; Allis, C.D. Multivalent engagement of chromatin modifications by linked binding modules. Nat. Rev. Mol. Cell Biol. 2007, 8, 983–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruizandres, O.; Sanchezniño, M.; Cannataortiz, P.; Ruizortega, M.; Egido, J.; Ortiz, A.; Sanz, A.B. Histone lysine crotonylation during acute kidney injury in mice. Dis. Models Mech. 2016, 9, 633–645. [Google Scholar]
- Tan, M.; Luo, H.; Lee, S.; Jin, F.; Yang, J.S.; Montellier, E.; Buchou, T.; Cheng, Z.; Rousseaux, S.; Rajagopal, N.; et al. Identification of 67 Histone Marks and Histone Lysine Crotonylation as a New Type of Histone Modification. Cell 2011, 146, 1016–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Liu, X.; Li, F.; Li, W.; Zhang, J.; Xiao, Z.; Shen, L.; Li, Y.; Wang, F.; Yang, J. First comprehensive proteome analysis of lysine crotonylation in seedling leaves of Nicotiana tabacum. Sci. Rep. 2017, 7, 3013. [Google Scholar] [CrossRef] [PubMed]
- Ntorla, A.; Burgoyne, J.R. The Regulation and Function of Histone Crotonylation. Front. Cell Dev. Biol. 2021, 9, 729. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.; Liu, S.; Sun, X.; Wang, X.; Xu, P.; Chen, H.; Yang, J. Global Lysine Crotonylation Profiling of Mouse Liver. Proteomics 2020, 20, 2000049. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, Z.; Liang, W.; Liu, M. Global Proteomic Analysis of Lysine Crotonylation in the Plant Pathogen Botrytis cinerea. Front. Microbiol. 2020, 11, 564350. [Google Scholar] [CrossRef]
- Kwon, O.K.; Kim, S.J.; Lee, S. First profiling of lysine crotonylation of myofilament proteins and ribosomal proteins in zebrafish embryos. Sci. Rep. 2018, 8, 3652. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, Y.; Apaliya, M.T.; Zheng, X.; Serwah, B.N.A.; Zhang, X.; Zhang, H. The Response of Rhodotorula mucilaginosa to Patulin Based on Lysine Crotonylation. Front. Microbiol. 2018, 9, 2025. [Google Scholar] [CrossRef]
- Xu, M.; Luo, J.; Li, Y.; Shen, L.; Zhang, X.; Yu, J.; Guo, Z.; Wu, J.; Chi, Y.; Yang, J. First comprehensive proteomics analysis of lysine crotonylation in leaves of peanut (Arachis hypogaea L.). Proteomics 2021, 21, e2000156. [Google Scholar] [CrossRef]
- Bao, C.; Song, C.; Liu, Y.; Yang, Y.; Cui, Z. Large-scale lysine crotonylation analysis reveals its potential role in spermiogenesis in the Chinese mitten crab Eriocheir sinensis. J. Proteom. 2020, 226, 103891. [Google Scholar] [CrossRef]
- Liu, S.; Xue, C.; Fang, Y.; Chen, G.; Peng, X.; Zhou, Y.; Chen, C.; Liu, G.; Gu, M.; Wang, K.; et al. Global Involvement of Lysine Crotonylation in Protein Modification and Transcription Regulation in Rice. Mol. Cell. Proteom. 2018, 17, 1922–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.-P.; Cheng, K.-W.; Chao, J.; Wu, J.; Wang, M. Tyrosinase inhibitors from paper mulberry (Broussonetia papyrifera). Food Chem. 2008, 106, 529–535. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Liu, C.-Q.; Li, P.-P.; Wang, J.-Z.; Xing, D.; Wang, B.-L. Photosynthetic characteristics involved in adaptability to Karst soil and alien invasion of paper mulberry (Broussonetia papyrifera (L.) Vent.) in comparison with mulberry (Morus alba L.). Photosynthetica 2009, 47, 155–160. [Google Scholar] [CrossRef]
- Nagpal, U.M.K.; Bankar, A.V.; Pawar, N.J.; Kapadnis, B.P.; Zinjarde, S.S. Equilibrium and Kinetic Studies on Biosorption of Heavy Metals by Leaf Powder of Paper Mulberry (Broussonetia papyrifera). Water Air Soil Pollut. 2010, 215, 177–188. [Google Scholar] [CrossRef]
- Peng, X.; Liu, H.; Chen, P.; Tang, F.; Hu, Y.; Wang, F.; Pi, Z.; Zhao, M.; Chen, N.; Chen, H.; et al. A Chromosome-Scale Genome Assembly of Paper Mulberry (Broussonetia papyrifera) Provides New Insights into Its Forage and Papermaking Usage. Mol. Plant 2019, 12, 661–677. [Google Scholar] [CrossRef]
- He, D.; Wang, Q.; Li, M.; Damaris, R.N.; Yi, X.; Cheng, Z.; Yang, P. Global Proteome Analyses of Lysine Acetylation and Suc-cinylation Reveal the Widespread Involvement of both Modification in Metabolism in the Embryo of Germinating Rice Seed. J. Proteome Res. 2016, 15, 879–890. [Google Scholar] [CrossRef]
- Qiu, W.; Sun, B.; Xiao, X.; Xu, Z.; Jia, J.; Chou, K. iKcr-PseEns: Identify lysine crotonylation sites in histone proteins with pseudo components and ensemble classifier. Genomics 2018, 110, 239–246. [Google Scholar] [CrossRef]
- Sun, J.; Qiu, C.; Qian, W.; Wang, Y.; Sun, L.; Li, Y.; Ding, Z. Ammonium triggered the response mechanism of lysine crotonylome in tea plants. BMC Genom. 2019, 20, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Yuan, C.; Li, H.; Chen, K.; Lu, L.; Shen, C.; Zheng, X. A qualitative proteome-wide lysine crotonylation profiling of papaya (Carica papaya L.). Sci. Rep. 2018, 8, 8230. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xu, Q.; Liu, Y.; Yu, Y.; Cheng, Z.-Y.; Zhao, Y.; Zhou, D.-X. Dynamics and functional interplay of histone lysine butyrylation, crotonylation, and acetylation in rice under starvation and submergence. Genome Biol. 2018, 19, 144. [Google Scholar] [CrossRef] [PubMed]
- Vellai, T.; Vida, G. The origin of eukaryotes: The difference between prokaryotic and eukaryotic cells. Proc. R. Soc. B Boil. Sci. 1999, 266, 1571–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clauss, M.; Motel, A.; Lichtenthaler, H.K. Studies on the Plant Methylcrotonyl-CoA Carboxylase 1. J. Plant Physiol. 1993, 141, 508–511. [Google Scholar] [CrossRef]
- Smalle, J.; Kurepa, J.; Yang, P.; Emborg, T.J.; Babiychuk, E.; Kushnir, S.; Vierstra, R.D. The Pleiotropic Role of the 26S Proteasome Subunit RPN10 in Arabidopsis Growth and Development Supports a Substrate-Specific Function in Abscisic Acid Signaling. Plant Cell 2003, 15, 965–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, C.V.; Rey, P. Plant thioredoxins are key actors in the oxidative stress response. Trends Plant Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Howe, C.J.; Fearnley, I.M.; Walker, J.E.; Dyer, T.A.; Gray, J.C. Nucleotide sequences of the genes for the alpha, beta and epsilon subunits of wheat chloroplast ATP synthase. Plant Mol. Biol. 1985, 4, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.M. Effects of water deficit on photosynthetic capacity. Physiol. Plant. 1987, 71, 142–149. [Google Scholar] [CrossRef]
- Cornic, G.; Briantais, J.-M. Partitioning of photosynthetic electron flow between CO2 and O2 reduction in a C3 leaf (Phaseolus vulgaris L.) at different CO2 concentrations and during drought stress. Planta 1991, 183, 178–184. [Google Scholar] [CrossRef]
- Price, A.H.; Hendry, G.A.F. Iron-catalysed oxygen radical formation and its possible contribution to drought damage in nine native grasses and three cereals. Plant Cell Environ. 1991, 14, 477–484. [Google Scholar] [CrossRef]
- Zhang, X.; Glaser, E. Interaction of plant mitochondrial and chloroplast signal peptides with the Hsp70 molecular chaperone. Trends Plant Sci. 2002, 7, 14–21. [Google Scholar] [CrossRef]
- Qiu, W.; Sun, B.; Xiao, X.; Xu, Z.; Chou, K. iPTM-mLys: Identifying multiple lysine PTM sites and their different types. Bioinformatics 2016, 32, 3116–3123. [Google Scholar] [CrossRef] [Green Version]
- Sung, D.-Y.; Kaplan, F.; Guy, C.L. Plant Hsp70 molecular chaperones: Protein structure, gene family, expression and function. Physiol. Plant. 2001, 113, 443–451. [Google Scholar] [CrossRef]
- Zhou, X.; Song, N.; Li, D.; Li, X.; Liu, W. Systematic Analysis of the Lysine Crotonylome and Multiple Posttranslational Modification Analysis (Acetylation, Succinylation, and Crotonylation) in Candida albicans. Msystems 2021, 6, e01316-20. [Google Scholar] [CrossRef]
- Velazquez, J.M.; Lindquist, S. hsp70: Nuclear concentration during environmental stress and cytoplasmic storage during recovery. Cell 1984, 36, 655–662. [Google Scholar] [CrossRef]
- Alastalo, T.-P.; Hellesuo, M.; Sandqvist, A.; Hietakangas, V.; Kallio, M.; Sistonen, L. Formation of nuclear stress granules involves HSF2 and coincides with the nucleolar localization of Hsp70. J. Cell Sci. 2003, 116, 3557–3570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghaie, P.; Tafreshi, S.A.H. Central role of 70-kDa heat shock protein in adaptation of plants to drought stress. Cell Stress Chaperones 2020, 25, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive Expression Profile Analysis of the Arabidopsis Hsp70 Gene Family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-L.; Zhu, J.-H.; Zhang, Q.-Q.; Cai, Y.-B. Molecular characterization of an ethephon-induced Hsp70 involved in high and low-temperature responses in Hevea brasiliensis. Plant Physiol. Biochem. 2009, 47, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Bai, H.; He, L.; Huang, Q.; Liu, Q. Proteome-wide and lysine crotonylation profiling reveals the importance of crotonylation in chrysanthemum (Dendranthema grandiforum) under low-temperature. BMC Genom. 2020, 22, 51. [Google Scholar] [CrossRef]
- Martinez-Sena, T.; Luongo, G.; Sanjuan-Herráez, D.; Castell, J.V.; Vento, M.; Quintás, G.; Kuligowski, J. Monitoring of system conditioning after blank injections in untargeted UPLC-MS metabolomic analysis. Sci. Rep. 2019, 9, 9822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wixon, J.; Kell, D. The Kyoto Encyclopedia of Genes and Genomes—KEGG. Yeast 2000, 1, 48–55. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.C.; Petrey, D.; Garzón, J.I.; Deng, L.; Honig, B. PrePPI: A structure-informed database of protein–protein interactions. Nucleic Acids Res. 2012, 41, D828–D833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.; Grant, C.E.; Noble, W.S.; Bailey, T.L. MoMo: Discovery of statistically significant post-translational modification motifs. Bioinformatics 2018, 35, 2774–2782. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lysine Crotonylation | Number of Identified Proteins | Number of Identified Sites | Reference |
|---|---|---|---|
| Paper mulberry (Broussonetia papyrifera) | 2272 | 5159 | This study |
| Papaya (Carica papaya L.) | 2120 | 5995 | [23] |
| Peanut (Arachis hypogaea L.) | 2508 | 6051 | [13] |
| Tobacco (Nicotiana tabacum) | 637 | 2044 | [7] |
| Rice (Oryza sativa L. japonica) | 690 | 1265 | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Chen, C. Comprehensive Profiling of Paper Mulberry (Broussonetia papyrifera) Crotonylome Reveals the Significance of Lysine Crotonylation in Young Leaves. Int. J. Mol. Sci. 2022, 23, 1173. https://doi.org/10.3390/ijms23031173
Dong Y, Chen C. Comprehensive Profiling of Paper Mulberry (Broussonetia papyrifera) Crotonylome Reveals the Significance of Lysine Crotonylation in Young Leaves. International Journal of Molecular Sciences. 2022; 23(3):1173. https://doi.org/10.3390/ijms23031173
Chicago/Turabian StyleDong, Yibo, and Chao Chen. 2022. "Comprehensive Profiling of Paper Mulberry (Broussonetia papyrifera) Crotonylome Reveals the Significance of Lysine Crotonylation in Young Leaves" International Journal of Molecular Sciences 23, no. 3: 1173. https://doi.org/10.3390/ijms23031173
APA StyleDong, Y., & Chen, C. (2022). Comprehensive Profiling of Paper Mulberry (Broussonetia papyrifera) Crotonylome Reveals the Significance of Lysine Crotonylation in Young Leaves. International Journal of Molecular Sciences, 23(3), 1173. https://doi.org/10.3390/ijms23031173