
Production and Biochemical Characterization of Dimeric Recombinant Gremlin-1

 , , ,
, , ,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
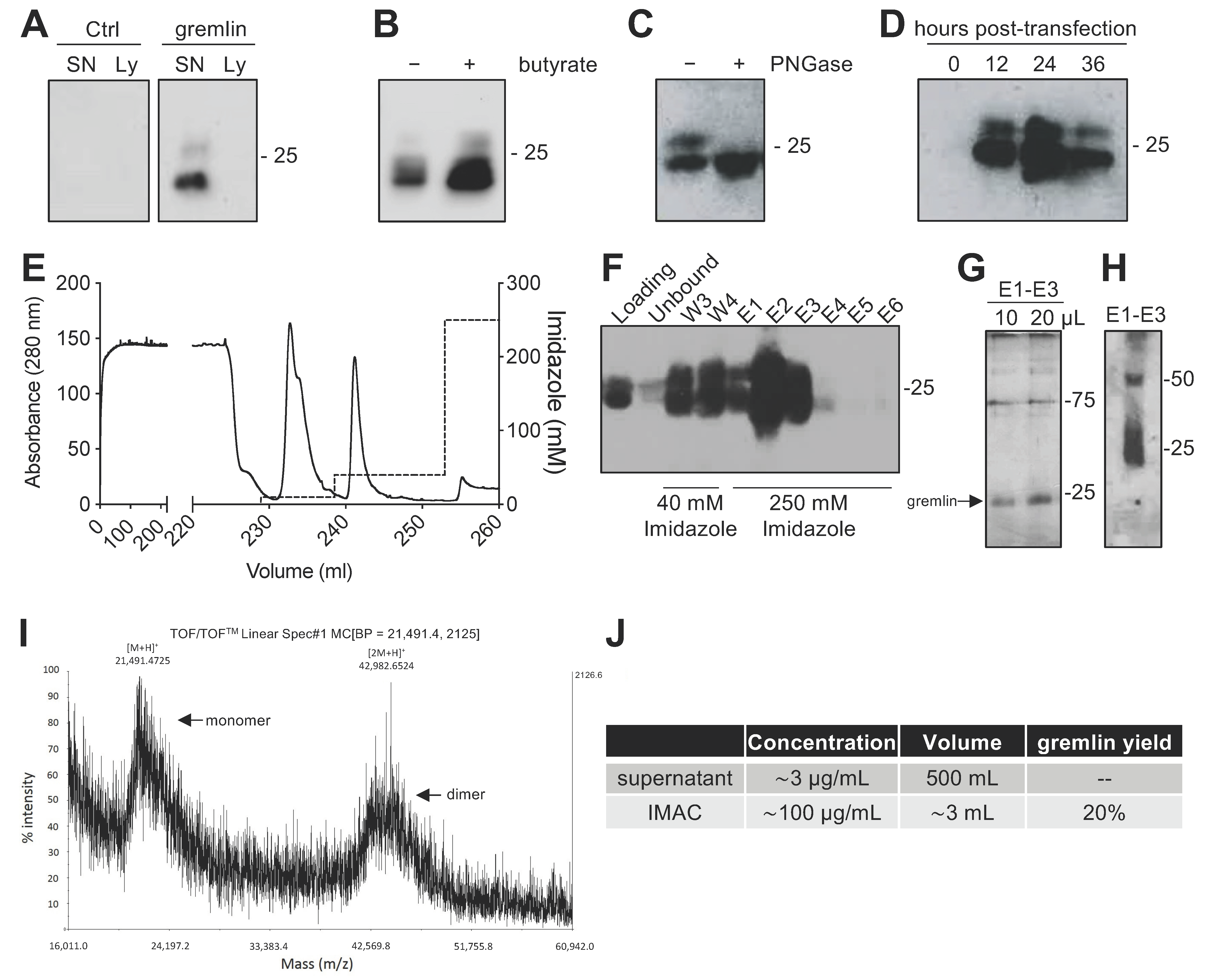
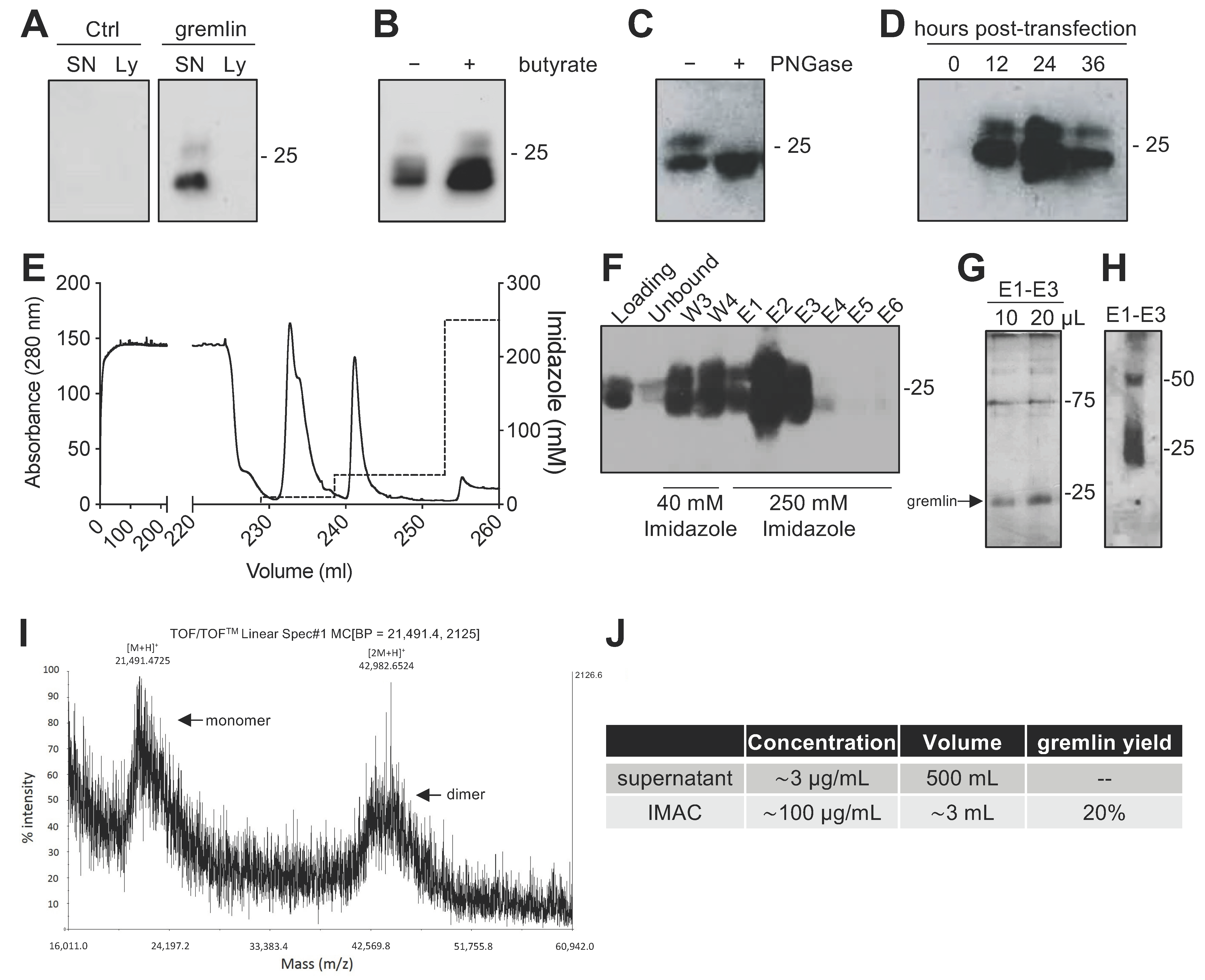
2.1. Expression of Recombinant Gremlin in HEK293T Cells
2.2. Expression of Recombinant Gremlin in HEK293T Cells Followed by Purification via IMAC Yields Gremlin Both in the Monomeric and Dimeric States
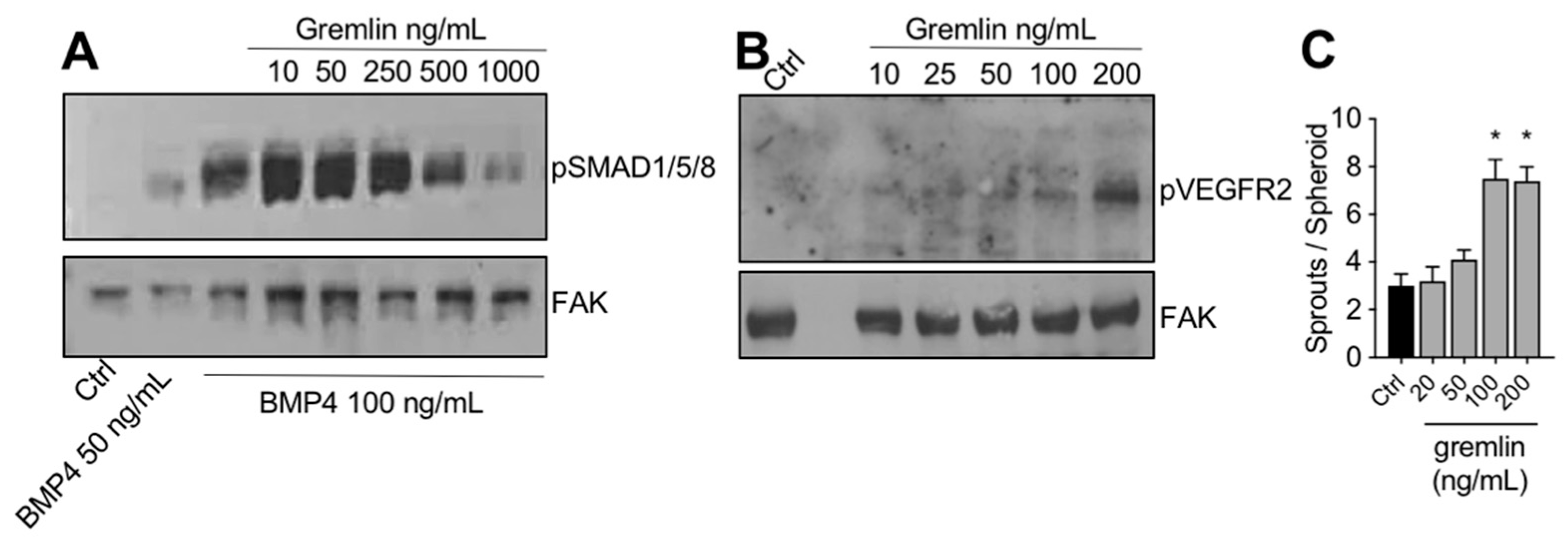
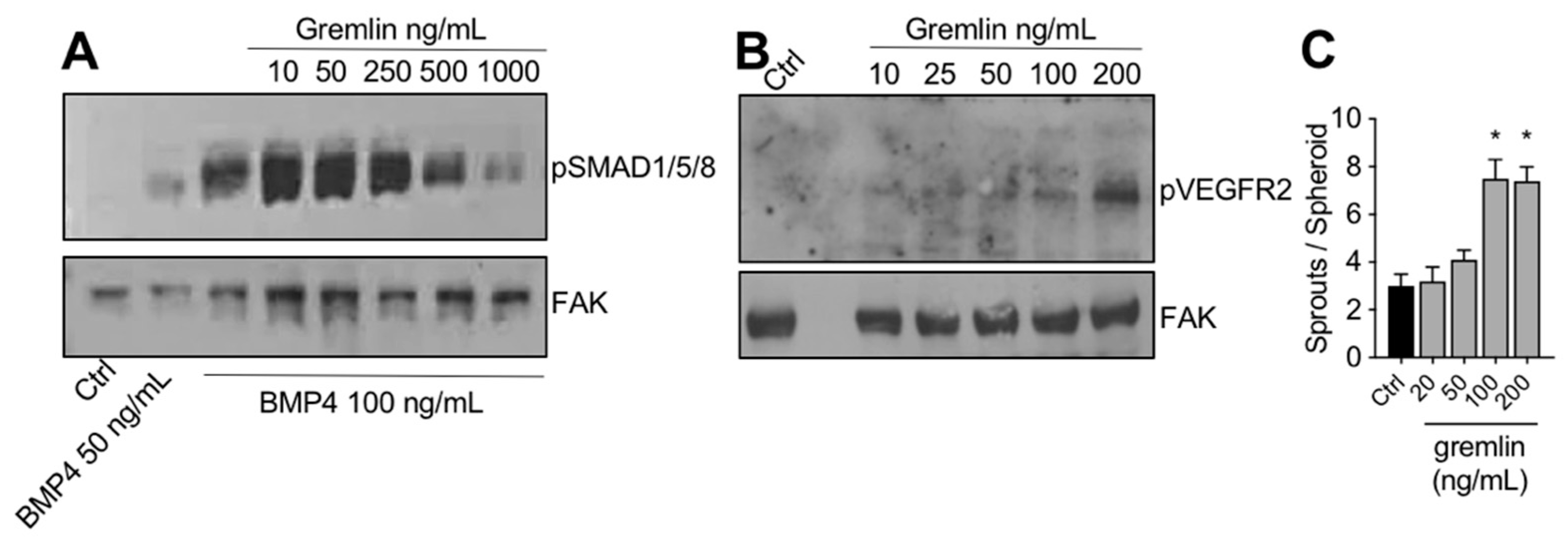
2.3. IMAC-Purified Gremlin Exhibits Full BMP Antagonist Activity and Only Mild VEGFR2 Activation Capacity
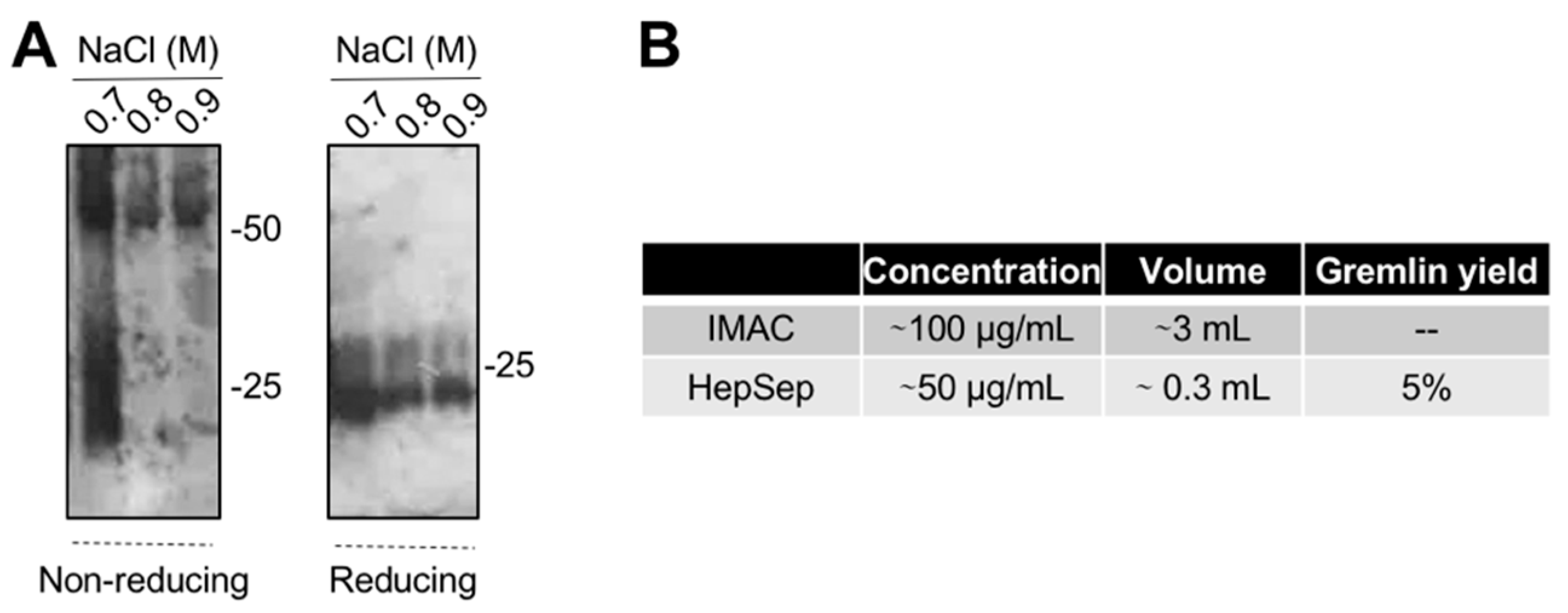
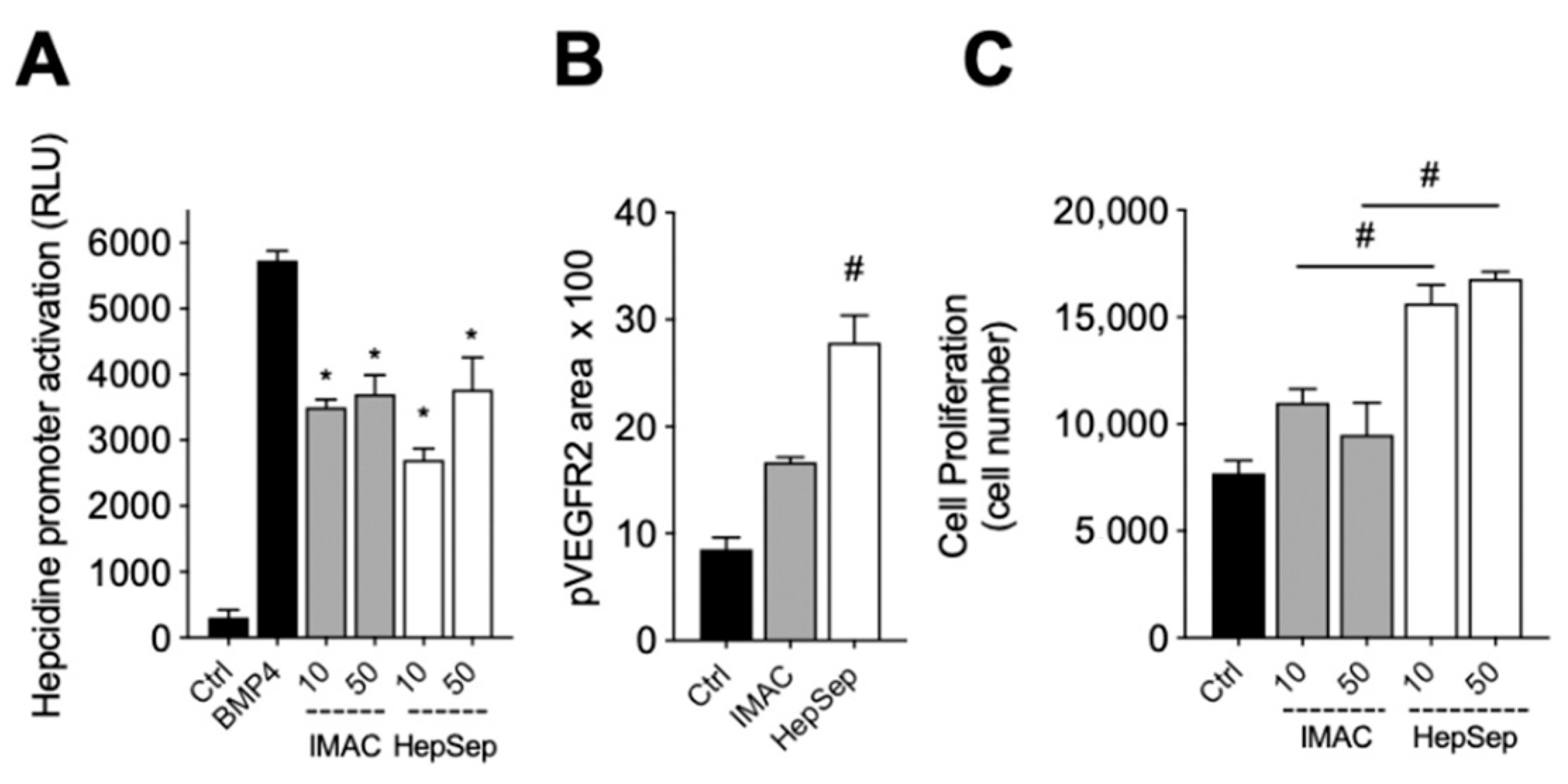
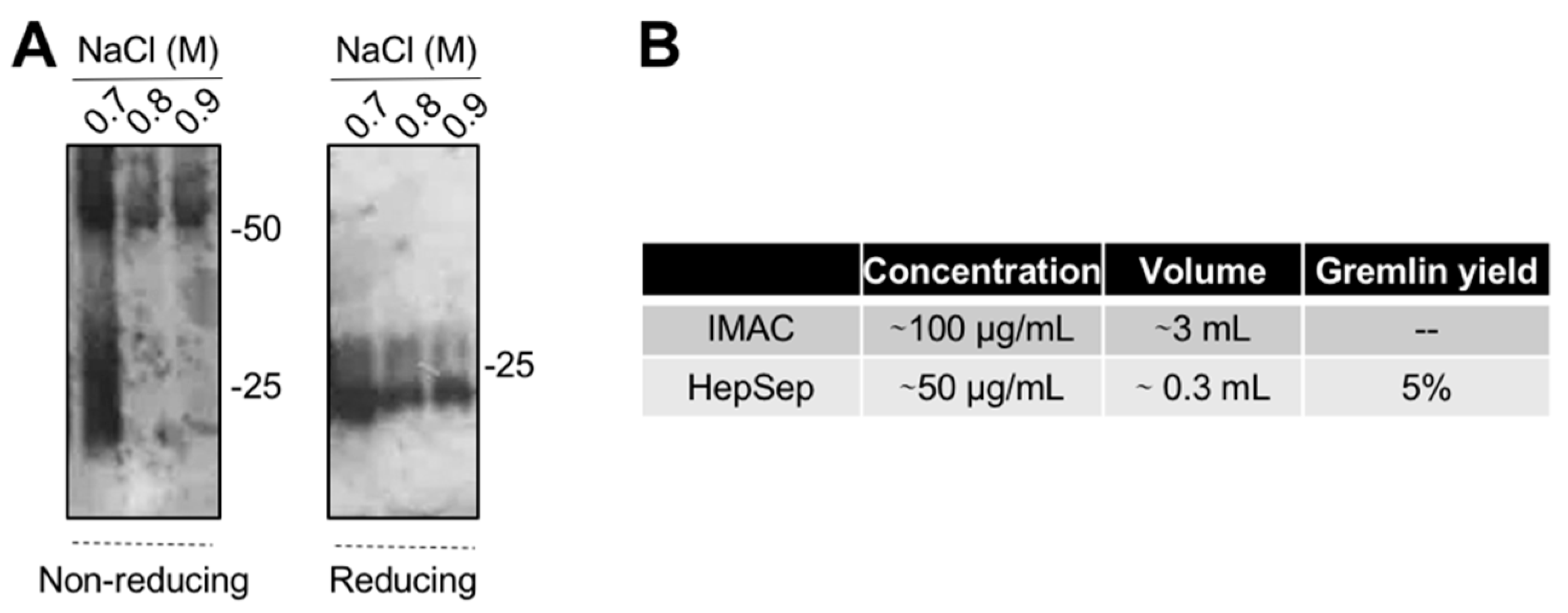
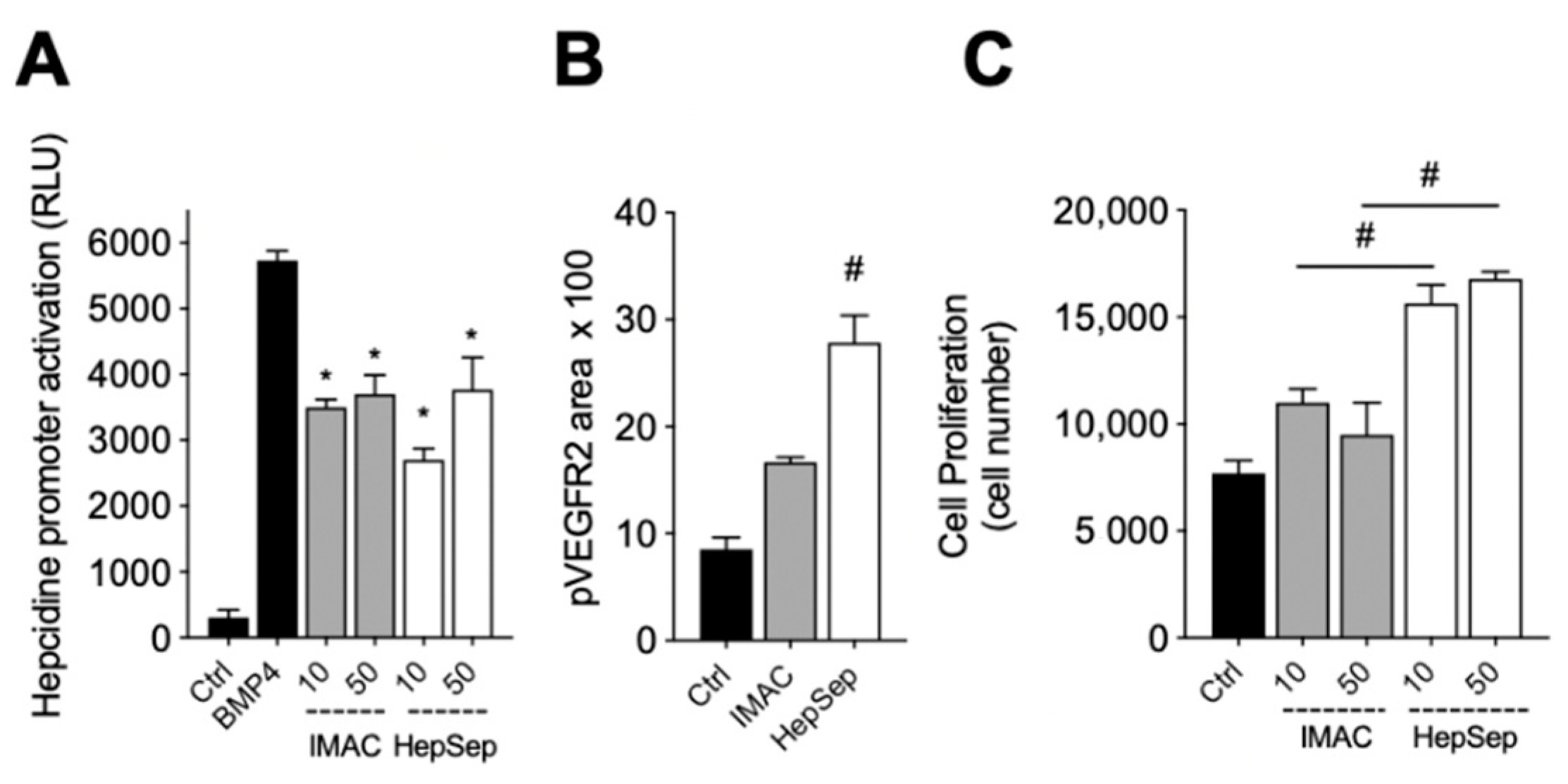
2.4. Purification of Fully Active, Dimeric Gremlin by Heparin-Affinity Chromatography
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Generation of DNA Constructs
4.3. Expression in HEK293T Cells and Purification of Recombinant Gremlin
4.4. Gel Analyses and Western Blotting
4.5. Matrix-Assisted Laser Desorption/Ionization (MALDI) Time-Of-Flight (TOF)/TOF Mass Spectrometry (MS) (MALDI-TOF/TOF–MS) Analyses
4.6. Analysis of SMAD and VEGFR2 Activation
4.7. Luciferase Reporter Gene Assay
4.8. Ventral Plasma Membrane Preparation
4.9. HUVEC Proliferation
4.10. Endothelial Cell Sprouting Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Topol, L.; Modi, W.; Koochekpour, S.; Blair, D. DRM/GREMLIN (CKTSF1B1) maps to human chromosome 15 and is highly expressed in adult and fetal brain. Cytogenet. Cell Genet. 2000, 89, 79–84. [Google Scholar] [CrossRef]
- Topol, L.Z.; Bardot, B.; Zhang, Q.; Resau, J.; Huillard, E.; Marx, M.; Calothy, G.; Blair, D.G. Biosynthesis, Post-translation Modification, and Functional Characterization of Drm/Gremlin. J. Biol. Chem. 2000, 275, 8785–8793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stabile, H.; Mitola, S.M.F.; Moroni, E.; Belleri, M.; Nicoli, S.; Coltrini, D.; Peri, F.; Pessi, A.; Orsatti, L.; Talamo, F.; et al. Bone morphogenic protein antagonist Drm/gremlin is a novel proangiogenic factor. Blood 2007, 109, 1834–1840. [Google Scholar] [CrossRef] [Green Version]
- Avsian-Kretchmer, O.; Hsueh, A.J.W. Comparative Genomic Analysis of the Eight-Membered Ring Cystine Knot-Containing Bone Morphogenetic Protein Antagonists. Mol. Endocrinol. 2004, 18, 1–12. [Google Scholar] [CrossRef]
- Kisonaite, M.; Wang, X.; Hyvönen, M. Structure of Gremlin-1 and analysis of its interaction with BMP-2. Biochem. J. 2016, 473, 1593–1604. [Google Scholar] [CrossRef] [Green Version]
- Grillo, E.; Ravelli, C.; Corsini, M.; Ballmer-Hofer, K.; Zammataro, L.; Oreste, P.; Zoppetti, G.; Tobia, C.; Ronca, R.; Presta, M.; et al. Monomeric gremlin is a novel vascular endothelial growth factor receptor-2 antagonist. Oncotarget 2016, 7, 35353–35368. [Google Scholar] [CrossRef] [Green Version]
- Michos, O.; Panman, L.; Vintersten, K.; Beier, K.; Zeller, R.; Zuniga, A. Gremlin-mediated BMP antagonism induces the epithelial-mesenchymal feedback signaling controlling metanephric kidney and limb organogenesis. Development 2004, 131, 3401–3410. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Smid, M.; Iaria, J.; Salvatori, D.C.F.; van Dam, H.; Zhu, H.J.; Martens, J.W.M.; Dijke, P.T. Cancer-associated fibroblast-derived Gremlin 1 promotes breast cancer progression. Breast Cancer Res. 2019, 21, 109. [Google Scholar] [CrossRef] [Green Version]
- Koli, K.; Myllãrniemi, L.M.; Vuorinen, K.; Salmenkivi, K.; Ryynänen, M.J.; Kinnula, V.L.; Keski-Oja, J. Bone Morphogenetic Protein-4 Inhibitor Gremlin Is Overexpressed in Idiopathic Pulmonary Fibrosis. Am. J. Pathol. 2006, 169, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Blair, D.G.; Plisov, S.; Vasiliev, G.; Perantoni, A.O.; Chen, Q.; Athanasiou, M.; Wu, J.Y.; Oppenheim, J.J.; Yang, D. Cutting Edge: Bone Morphogenetic Protein Antagonists Drm/Gremlin and Dan Interact with Slits and Act as Negative Regulators of Monocyte Chemotaxis. J. Immunol. 2004, 173, 5914–5917. [Google Scholar] [CrossRef] [Green Version]
- Mitola, S.M.F.; Ravelli, C.; Moroni, E.; Salvi, V.; Leali, D.; Ballmer-Hofer, K.; Zammataro, L.; Presta, M. Gremlin is a novel agonist of the major proangiogenic receptor VEGFR2. Blood 2010, 116, 3677–3680. [Google Scholar] [CrossRef] [Green Version]
- Corsini, M.; Moroni, E.; Ravelli, C.; Andrés, G.; Grillo, E.; Ali, I.H.; Brazil, D.P.; Presta, M.; Mitola, S. Cyclic Adenosine Monophosphate-Response Element–Binding Protein Mediates the Proangiogenic or Proinflammatory Activity of Gremlin. Arter. Thromb. Vasc. Biol. 2014, 34, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoz, C.; Alique, M.; Díez, R.R.; Pato, J.; Keri, G.; Mezzano, S.; Egido, J.; Ruiz-Ortega, M. Gremlin regulates renal inflammation via the vascular endothelial growth factor receptor 2 pathway. J. Pathol. 2015, 236, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Mitola, S.; Moroni, E.; Ravelli, C.; Andres, G.; Belleri, M.; Presta, M. Angiopoietin-1 mediates the proangiogenic activity of the bone morphogenic protein antagonist Drm. Blood 2008, 112, 1154–1157. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.C.; Piouceau, L.; Cornwell, J.; Li, L.; McLoughlin, P. EXPRESS: Gremlin1 blocks vascular endothelial growth factor signalling in the pulmonary microvascular endothelium. Pulm. Circ. 2018, 10, 2045894018807205. [Google Scholar]
- Chiodelli, P.; Mitola, S.; Ravelli, C.; Oreste, P.; Rusnati, M.; Presta, M. Heparan Sulfate Proteoglycans Mediate the Angiogenic Activity of the Vascular Endothelial Growth Factor Receptor-2 Agonist Gremlin. Arter. Thromb. Vasc. Biol. 2011, 31, e116–e127. [Google Scholar] [CrossRef] [Green Version]
- Tatsinkam, A.J.; Mulloy, B.; Rider, C. Mapping the heparin-binding site of the BMP antagonist gremlin by site-directed mutagenesis based on predictive modelling. Biochem. J. 2015, 470, 53–64. [Google Scholar] [CrossRef]
- Dutton, L.R.; O’Neill, C.L.; Medina, R.J.; Brazil, D.P. No evidence of Gremlin1-mediated activation of VEGFR2 signaling in endothelial cells. J. Biol. Chem. 2019, 294, 18041–18045. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, X.; Song, Z.; Zhang, Y. Sodium butyrate enhances the acidic isoform content of recombinant human erythropoietin produced by Chinese hamster ovary cells. Biotechnol. Lett. 2014, 36, 907–911. [Google Scholar] [CrossRef]
- Grünberg, J.; Knogler, K.; Waibel, R.; Novak-Hofer, I. High-Yield Production of Recombinant Antibody Fragments in HEK-293 Cells Using Sodium Butyrate. Biotechniques 2003, 34, 968–972. [Google Scholar] [CrossRef]
- Hedjazifar, S.; Shahidi, R.K.; Hammarstedt, A.; Bonnet, L.; Church, C.; Boucher, J.; Blüher, M.; Smith, U. The Novel Adipokine Gremlin 1 Antagonizes Insulin Action and Is Increased in Type 2 Diabetes and NAFLD/NASH. Diabetes 2020, 69, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zeng, Q.-S.; Zou, M.; Zeng, J.; Nie, J.; Chen, D.; Gan, H.-T. Targeting Gremlin 1 Prevents Intestinal Fibrosis Progression by Inhibiting the Fatty Acid Oxidation of Fibroblast Cells. Front. Pharmacol. 2021, 12, 663774. [Google Scholar] [CrossRef]
- Cain, K.; Peters, S.; Hailu, H.; Sweeney, B.; Stephens, P.; Heads, J.; Sarkar, K.; Ventom, A.; Page, C.; Dickson, A. A CHO cell line engineered to express XBP1 and ERO1-Lalpha has increased levels of transient protein expression. Biotechnol. Prog. 2013, 29, 697–706. [Google Scholar] [CrossRef]
- Borth, N.; Mattanovich, D.; Kunert, R.; Katinger, H. Effect of Increased Expression of Protein Disulfide Isomerase and Heavy Chain Binding Protein on Antibody Secretion in a Recombinant CHO Cell Line. Biotechnol. Prog. 2005, 21, 106–111. [Google Scholar] [CrossRef]
- Ravelli, C.; Grillo, E.; Corsini, M.; Coltrini, D.; Presta, M.; Mitola, S. β3 Integrin Promotes Long-Lasting Activation and Polarization of Vascular Endothelial Growth Factor Receptor 2 by Immobilized Ligand. Arter. Thromb. Vasc. Biol. 2015, 35, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Topol, L.Z.; Marx, M.; Laugier, D.; Bogdanova, N.N.; Boubnov, N.V.; A Clausen, P.; Calothy, G.; Blair, D.G. Identification of drm, a novel gene whose expression is suppressed in transformed cells and which can inhibit growth of normal but not transformed cells in culture. Mol. Cell. Biol. 1997, 17, 4801–4810. [Google Scholar] [CrossRef] [Green Version]
- Di Somma, M.; Schaafsma, W.; Grillo, E.; Vliora, M.; Dakou, E.; Corsini, M.; Ravelli, C.; Ronca, R.; Sakellariou, P.; Vanparijs, J.; et al. Natural Histogel-Based Bio-Scaffolds for Sustaining Angiogenesis in Beige Adipose Tissue. Cells 2019, 8, 1457. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitola, S.; Ravelli, C.; Corsini, M.; Gianoncelli, A.; Galvagni, F.; Ballmer-Hofer, K.; Presta, M.; Grillo, E. Production and Biochemical Characterization of Dimeric Recombinant Gremlin-1. Int. J. Mol. Sci. 2022, 23, 1151. https://doi.org/10.3390/ijms23031151
Mitola S, Ravelli C, Corsini M, Gianoncelli A, Galvagni F, Ballmer-Hofer K, Presta M, Grillo E. Production and Biochemical Characterization of Dimeric Recombinant Gremlin-1. International Journal of Molecular Sciences. 2022; 23(3):1151. https://doi.org/10.3390/ijms23031151
Chicago/Turabian StyleMitola, Stefania, Cosetta Ravelli, Michela Corsini, Alessandra Gianoncelli, Federico Galvagni, Kurt Ballmer-Hofer, Marco Presta, and Elisabetta Grillo. 2022. "Production and Biochemical Characterization of Dimeric Recombinant Gremlin-1" International Journal of Molecular Sciences 23, no. 3: 1151. https://doi.org/10.3390/ijms23031151
APA StyleMitola, S., Ravelli, C., Corsini, M., Gianoncelli, A., Galvagni, F., Ballmer-Hofer, K., Presta, M., & Grillo, E. (2022). Production and Biochemical Characterization of Dimeric Recombinant Gremlin-1. International Journal of Molecular Sciences, 23(3), 1151. https://doi.org/10.3390/ijms23031151