
Differential Upregulation and Functional Activity of S1PR1 in Human Peripheral Blood Basophils of Atopic Patients

 , , , , , and
, , , , , and 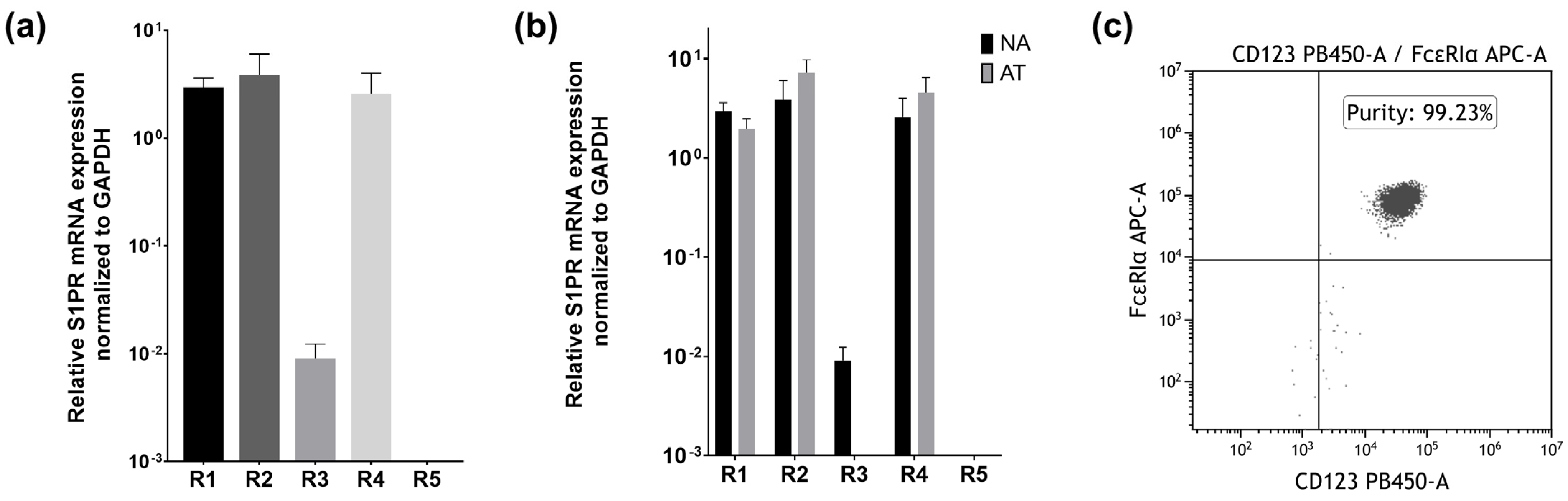{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Basophils Express S1PR1, S1PR2, S1PR3, and S1PR4 on the mRNA Level
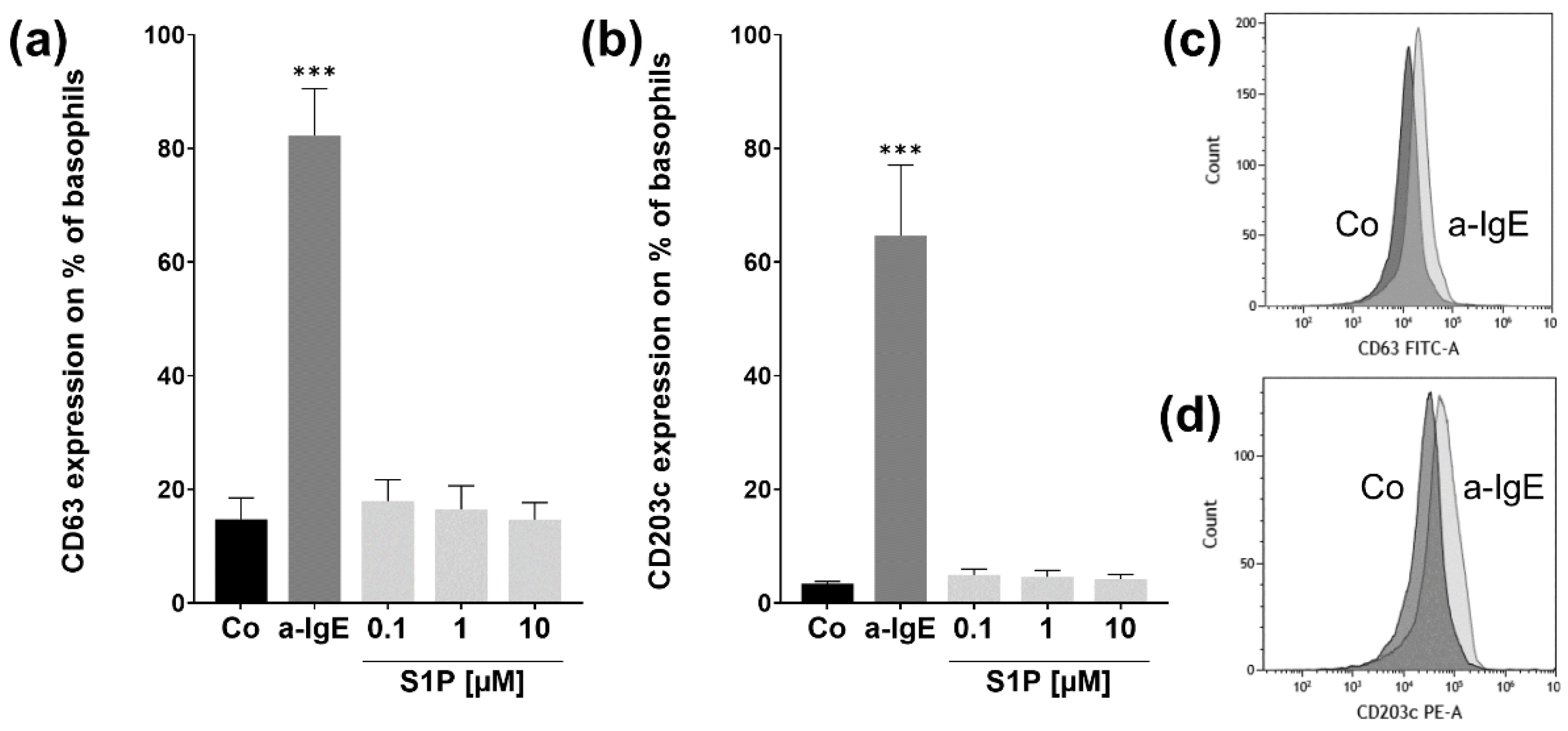
2.2. S1P Does Not Influence Basophil CD63 or CD203c Expression in Basophils from NA Patients
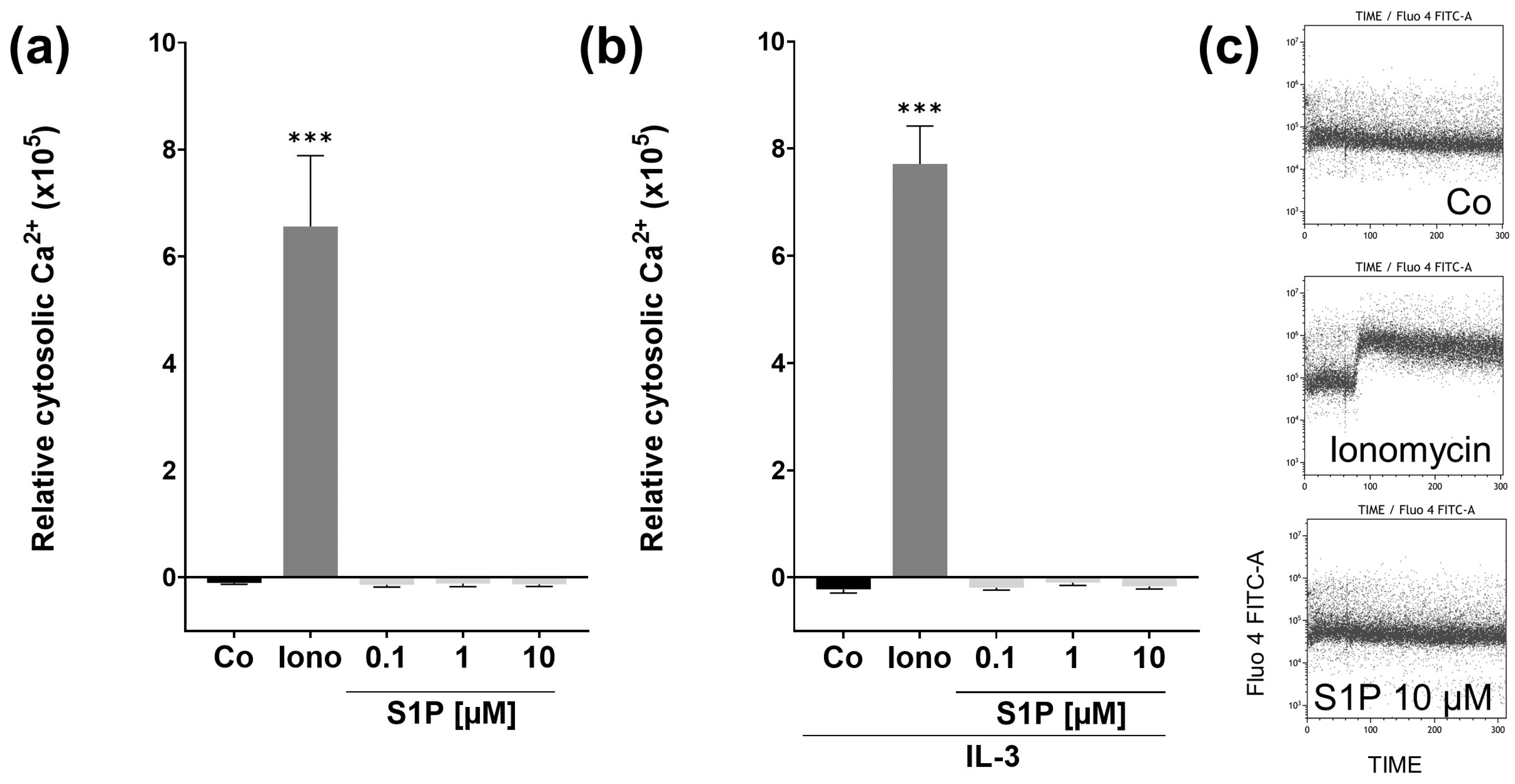
2.3. S1P Does Not Affect Short Term Cytosolic Ca2+ Levels in Basophils from NA Patients
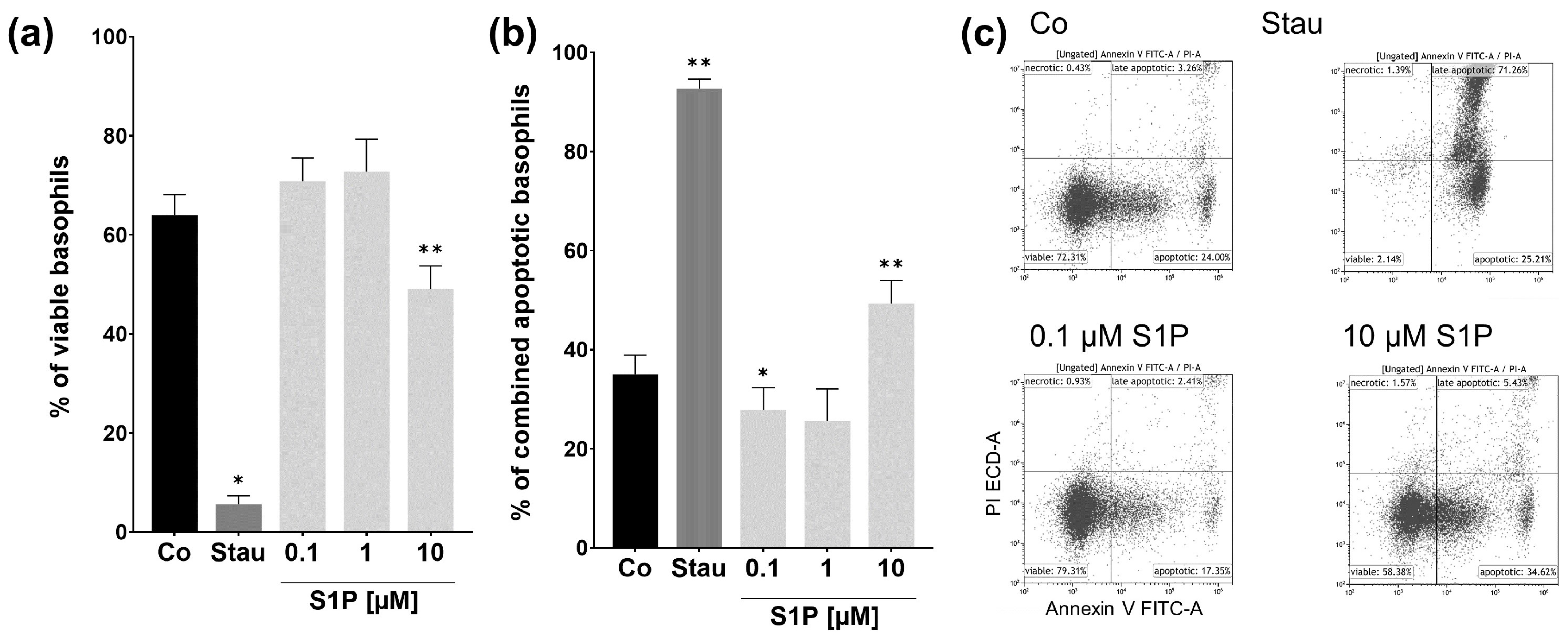
2.4. S1P Has Pro- and Antiapoptotic Effects on Basophils from NA Patients
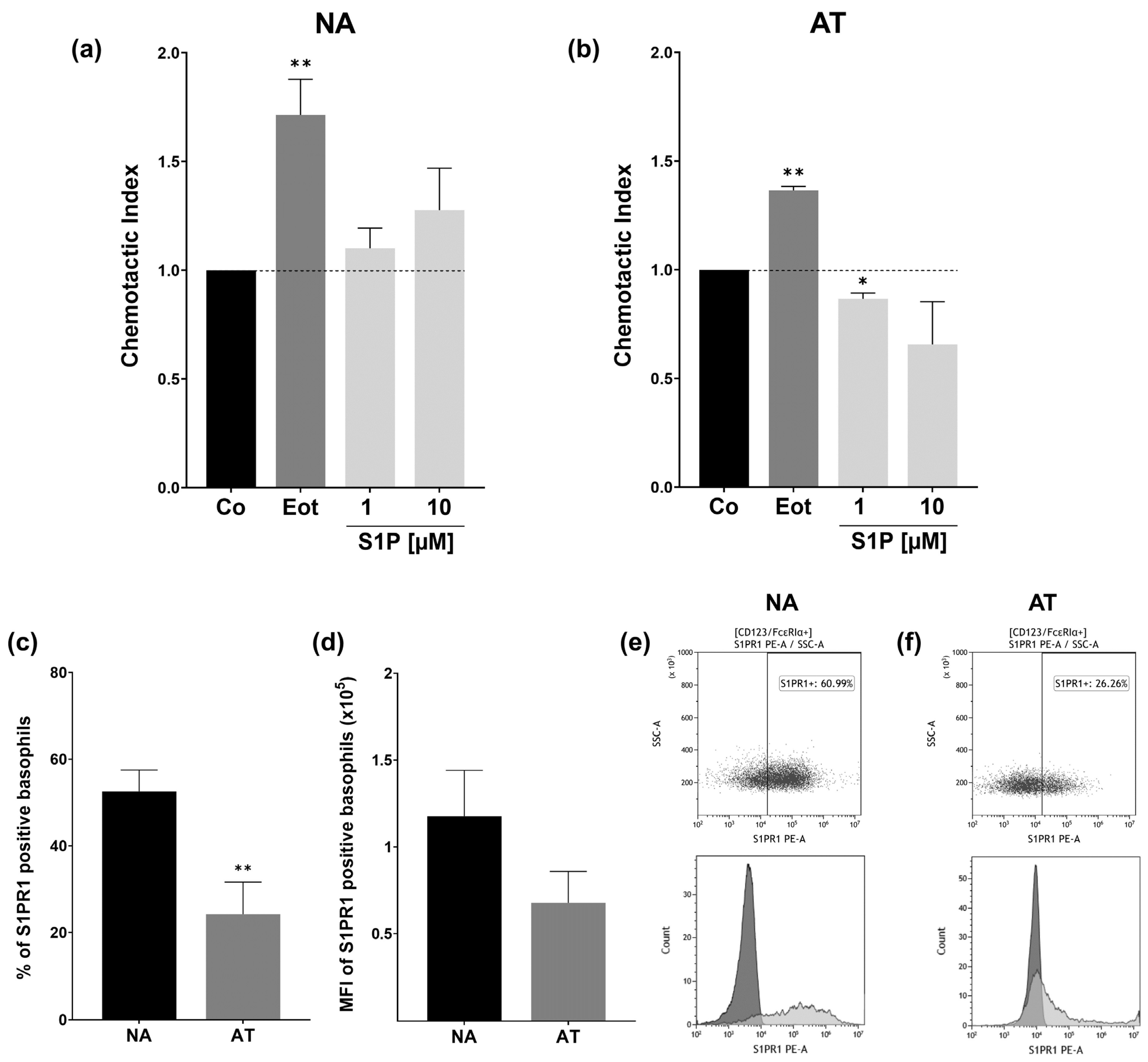
2.5. S1PR1 Protein Expression and Basophil Chemotaxis Differ in NA and AT Patients
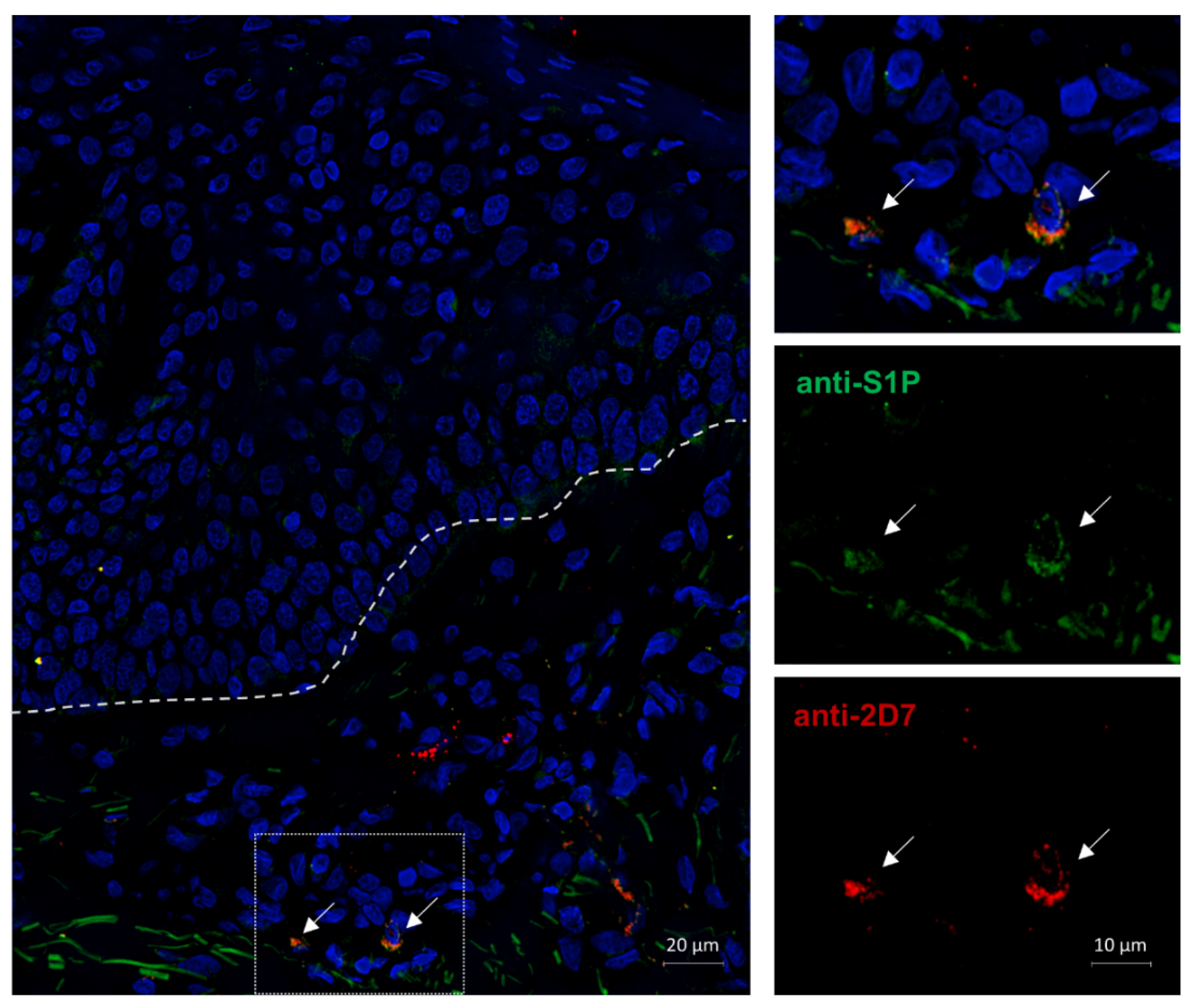
2.6. S1P Is Expressed Intracellularly in the Basophils of an AD Patient
3. Discussion
4. Materials and Methods
4.1. Collection of Blood Samples
4.2. Basophil Isolation
4.3. Quantitative Real-Time PCRs
4.4. Basophil Activation Test (BAT)
4.5. Calcium-Flux Experiments
4.6. Apoptosis Assay
4.7. Chemotaxis Assay
4.8. S1PR1 Protein Expression Analysis
4.9. Immunofluorescence Staining of AD Skin Sections
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APC | allophycocyanin | |
| AD | atopic dermatitis | |
| a-IgE | anti-IgE | |
| AT | atopic | |
| BALF | bronchoalveolar lavage fluid | |
| BAT | basophil activation test | |
| BSA | bovine serum albumin | |
| Ca | calcium | |
| CCR3 | chemokine receptor 3 | |
| CI | chemotactic index | |
| CO2 | carbon dioxide | |
| cDNA | complementary deoxyribonucleic acid | |
| EDTA | ethylenediaminetetraacetic acid | |
| FCS | fetal calf serum | |
| FMO | fluorescence minus one | |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase | |
| h | hours | |
| IgE | immunoglobulin E | |
| IL-3 | interleukin 3 | |
| IL-4 | interleukin 4 | |
| IL-10 | interleukin 10 | |
| IL-13 | interleukin 13 | |
| IL-31 | interleukin 31 | |
| LTC4 | leukotriene 4 | |
| MCP-4 | monocyte chemotactic protein 4 | |
| MFI | mean fluorescence intensity | |
| min | minutes | |
| mRNA | messenger ribonucleic acid | |
| NA | non-atopic | |
| NET | neutrophil extracellular traps | |
| NK | natural killer | |
| PB | pacific blue | |
| PC | polycarbonate | |
| PE | phycoerythrin | |
| PI | propidium iodide | |
| RNA | ribonucleic acid | |
| s | seconds | |
| S1P | sphingosine-1-phosphate | |
| S1PR | sphingosine-1-phosphate receptor | |
| SEM | standard error of the mean | |
| TALL | T cell acute lymphoblastic leukemia | |
| Th2 | T helper cell type 2 | |
| TSLP | thymic stromal lymphopoietin | |
| qRT-PCR | quantitative real-time polymerase chain reaction | |
References
- Siracusa, M.C.; Kim, B.S.; Spergel, J.M.; Artis, D. Basophils and allergic inflammation. J. Allergy Clin. Immunol. 2013, 132, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.T. Chapter 4 Basophils. In Advances in Immunology; Alt, F.W., Austen, K.F., Eds.; Academic Press: Amsterdam, The Netherlands, 2009; Volume 101, pp. 123–161. ISBN 9780123747938. [Google Scholar]
- Iype, J.; Fux, M. Basophils Orchestrating Eosinophils’ Chemotaxis and Function in Allergic Inflammation. Cells 2021, 10, 895. [Google Scholar] [CrossRef]
- Bernstein, D.I.; Schwartz, G.; Bernstein, J.A. Allergic Rhinitis: Mechanisms and Treatment. Immunol. Allergy Clin. N. Am. 2016, 36, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Reber, L.L.; Hernandez, J.D.; Galli, S.J. The pathophysiology of anaphylaxis. J. Allergy Clin. Immunol. 2017, 140, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, A.J.; Kon, O.M.; Smith, S.J.; Zeibecoglou, K.; Khan, L.; Barata, L.T.; McEuen, A.R.; Buckley, M.G.; Walls, A.F.; Meng, Q.; et al. Basophils, eosinophils, and mast cells in atopic and nonatopic asthma and in late-phase allergic reactions in the lung and skin. J. Allergy Clin. Immunol. 2000, 105, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F.; Mihm, M.C. Basophilic leukocytes in allergic contact dermatitis. J. Exp. Med. 1972, 135, 235–254. [Google Scholar] [CrossRef]
- Ito, Y.; Satoh, T.; Takayama, K.; Miyagishi, C.; Walls, A.F.; Yokozeki, H. Basophil recruitment and activation in inflammatory skin diseases. Allergy 2011, 66, 1107–1113. [Google Scholar] [CrossRef]
- Ishizaka, T.; de Bernardo, R.; Tomioka, H.; Lichtenstein, L.M.; Ishizaka, K. Identification of basophil granulocytes as a site of allergic histamine release. J. Immunol. 1972, 108, 1000–1008. [Google Scholar]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef]
- May, C.D. High spontaneous release of histamine in vitro from leukocytes of persons hypersensitive to food. J. Allergy Clin. Immunol. 1976, 58, 432–437. [Google Scholar] [CrossRef]
- MacGlashan, D.W.; Peters, S.P.; Warner, J.; Lichtenstein, L.M. Characteristics of human basophil sulfidopeptide leukotriene release: Releasability defined as the ability of the basophil to respond to dimeric cross-links. J. Immunol. 1986, 136, 2231–2239. [Google Scholar] [PubMed]
- MacGlashan, D.; White, J.M.; Huang, S.K.; Ono, S.J.; Schroeder, J.T.; Lichtenstein, L.M. Secretion of IL-4 from human basophils. The relationship between IL-4 mRNA and protein in resting and stimulated basophils. J. Immunol. 1994, 152, 3006–3016. [Google Scholar] [PubMed]
- Li, H.; Sim, T.C.; Alam, R. IL-13 released by and localized in human basophils. J. Immunol. 1996, 156, 4833–4838. [Google Scholar] [PubMed]
- Raap, U.; Gehring, M.; Kleiner, S.; Rüdrich, U.; Eiz-Vesper, B.; Haas, H.; Kapp, A.; Gibbs, B.F. Human basophils are a source of -and are differentially activated by-IL-31. Clin. Exp. Allergy 2017, 47, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Mukai, K.; Tsujimura, Y.; Ishiwata, K.; Kawano, Y.; Minegishi, Y.; Watanabe, N.; Karasuyama, H. Basophils are essential initiators of a novel type of chronic allergic inflammation. Blood 2007, 110, 913–920. [Google Scholar] [CrossRef]
- Kumar, A.; Rani, L.; Mhaske, S.T.; Pote, S.T.; Behera, S.; Mishra, G.C.; Wani, M.R. IL-3 Receptor Expression on Activated Human Th Cells Is Regulated by IL-4, and IL-3 Synergizes with IL-4 to Enhance Th2 Cell Differentiation. J. Immunol. 2020, 204, 819–831. [Google Scholar] [CrossRef]
- Varricchi, G.; Poto, R.; Marone, G.; Schroeder, J.T. IL-3 in the development and function of basophils. Semin. Immunol. 2021, 54, 101510. [Google Scholar] [CrossRef]
- Vilariño, N.; Miura, K.; MacGlashan, D.W. Acute IL-3 priming up-regulates the stimulus-induced Raf-1-Mek-Erk cascade independently of IL-3-induced activation of Erk. J. Immunol. 2005, 175, 3006–3014. [Google Scholar] [CrossRef]
- Bonam, S.R.; Chauvin, C.; Mathew, M.J.; Bayry, J. IFN-γ Induces PD-L1 Expression in Primed Human Basophils. Cells 2022, 11, 801. [Google Scholar] [CrossRef]
- Tsujimura, Y.; Obata, K.; Mukai, K.; Shindou, H.; Yoshida, M.; Nishikado, H.; Kawano, Y.; Minegishi, Y.; Shimizu, T.; Karasuyama, H. Basophils play a pivotal role in immunoglobulin-G-mediated but not immunoglobulin-E-mediated systemic anaphylaxis. Immunity 2008, 28, 581–589. [Google Scholar] [CrossRef]
- Chen, K.; Xu, W.; Wilson, M.; He, B.; Miller, N.W.; Bengtén, E.; Edholm, E.-S.; Santini, P.A.; Rath, P.; Chiu, A.; et al. Immunoglobulin D enhances immune surveillance by activating antimicrobial, proinflammatory and B cell–stimulating programs in basophils. Nat. Immunol. 2009, 10, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, M.C.; Saenz, S.A.; Hill, D.A.; Kim, B.S.; Headley, M.B.; Doering, T.A.; Wherry, E.J.; Jessup, H.K.; Siegel, L.A.; Kambayashi, T.; et al. TSLP promotes interleukin-3-independent basophil haematopoiesis and type 2 inflammation. Nature 2011, 477, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.; Coward, W.R.; Pritchard, D.I.; Hewitt, C.R.A. Basophils express a type 2 cytokine profile on exposure to proteases from helminths and house dust mites. J. Leukoc. Biol. 2003, 73, 165–171. [Google Scholar] [CrossRef]
- Patella, V.; Florio, G.; Petraroli, A.; Marone, G. HIV-1 gp120 induces IL-4 and IL-13 release from human Fc epsilon RI+ cells through interaction with the VH3 region of IgE. J. Immunol. 2000, 164, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Schramm, G.; Mohrs, K.; Wodrich, M.; Doenhoff, M.J.; Pearce, E.J.; Haas, H.; Mohrs, M. Cutting edge: IPSE/α-1, a glycoprotein from Schistosoma mansoni eggs, induces IgE-dependent, antigen-independent IL-4 production by murine basophils in vivo. J. Immunol. 2007, 178, 6023–6027. [Google Scholar] [CrossRef] [PubMed]
- Sarfati, M.; Wakahara, K.; Chapuy, L.; Delespesse, G. Mutual Interaction of Basophils and T Cells in Chronic Inflammatory Diseases. Front. Immunol. 2015, 6, 399. [Google Scholar] [CrossRef] [PubMed]
- Rivellese, F.; Suurmond, J.; de Paulis, A.; Marone, G.; Huizinga, T.W.J.; Toes, R.E.M. IgE and IL-33-mediated triggering of human basophils inhibits TLR4-induced monocyte activation. Eur. J. Immunol. 2014, 44, 3045–3055. [Google Scholar] [CrossRef]
- Egawa, M.; Mukai, K.; Yoshikawa, S.; Iki, M.; Mukaida, N.; Kawano, Y.; Minegishi, Y.; Karasuyama, H. Inflammatory monocytes recruited to allergic skin acquire an anti-inflammatory M2 phenotype via basophil-derived interleukin-4. Immunity 2013, 38, 570–580. [Google Scholar] [CrossRef]
- Kleiner, S.; Rüdrich, U.; Gehring, M.; Loser, K.; Eiz-Vesper, B.; Noubissi Nzeteu, G.A.; Patsinakidis, N.; Meyer, N.H.; Gibbs, B.F.; Raap, U. Human basophils release the anti-inflammatory cytokine IL-10 following stimulation with α-melanocyte-stimulating hormone. J. Allergy Clin. Immunol. 2021, 147, 1521–1523.e3. [Google Scholar] [CrossRef]
- Tsai, H.-C.; Han, M.H. Sphingosine-1-Phosphate (S1P) and S1P Signaling Pathway: Therapeutic Targets in Autoimmunity and Inflammation. Drugs 2016, 76, 1067–1079. [Google Scholar] [CrossRef]
- Gray, N.; Limberg, M.M.; Bräuer, A.U.; Raap, U. Novel functions of S1P in chronic itchy and inflammatory skin diseases. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Perales, A.; Escribese, M.M.; Garrido-Arandia, M.; Obeso, D.; Izquierdo-Alvarez, E.; Tome-Amat, J.; Barber, D. The Role of Sphingolipids in Allergic Disorders. Front. Allergy 2021, 2, 675557. [Google Scholar] [CrossRef] [PubMed]
- Calise, S.; Blescia, S.; Cencetti, F.; Bernacchioni, C.; Donati, C.; Bruni, P. Sphingosine 1-phosphate stimulates proliferation and migration of satellite cells: Role of S1P receptors. Biochim. Biophys. Acta 2012, 1823, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Takuwa, Y.; Du, W.; Qi, X.; Okamoto, Y.; Takuwa, N.; Yoshioka, K. Roles of sphingosine-1-phosphate signaling in angiogenesis. World J. Biol. Chem. 2010, 1, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Cuvillier, O.; Edsall, L.C.; Kohama, T.; Menzeleev, R.; Olah, Z.; Olivera, A.; Pirianov, G.; Thomas, D.M.; Tu, Z.; et al. Sphingosine-1-phosphate in cell growth and cell death. Ann. N. Y. Acad. Sci. 1998, 845, 11–18. [Google Scholar] [CrossRef]
- Kumar, A.; Saba, J.D. Regulation of Immune Cell Migration by Sphingosine-1-Phosphate. Cell. Mol. Biol. OMICS 2015, 61, 121. [Google Scholar]
- Aarthi, J.J.; Darendeliler, M.A.; Pushparaj, P.N. Dissecting the role of the S1P/S1PR axis in health and disease. J. Dent. Res. 2011, 90, 841–854. [Google Scholar] [CrossRef]
- Matloubian, M.; Lo, C.G.; Cinamon, G.; Lesneski, M.J.; Xu, Y.; Brinkmann, V.; Allende, M.L.; Proia, R.L.; Cyster, J.G. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 2004, 427, 355–360. [Google Scholar] [CrossRef]
- Vora, K.A.; Nichols, E.; Porter, G.; Cui, Y.; Keohane, C.A.; Hajdu, R.; Hale, J.; Neway, W.; Zaller, D.; Mandala, S. Sphingosine 1-phosphate receptor agonist FTY720-phosphate causes marginal zone B cell displacement. J. Leukoc. Biol. 2005, 78, 471–480. [Google Scholar] [CrossRef]
- Allende, M.L.; Zhou, D.; Kalkofen, D.N.; Benhamed, S.; Tuymetova, G.; Borowski, C.; Bendelac, A.; Proia, R.L. S1P1 receptor expression regulates emergence of NKT cells in peripheral tissues. FASEB J. 2008, 22, 307–315. [Google Scholar] [CrossRef]
- Messias, C.V.; Santana-Van-Vliet, E.; Lemos, J.P.; Moreira, O.C.; Cotta-de-Almeida, V.; Savino, W.; Mendes-da-Cruz, D.A. Sphingosine-1-Phosphate Induces Dose-Dependent Chemotaxis or Fugetaxis of T-ALL Blasts through S1P1 Activation. PLoS ONE 2016, 11, e0148137. [Google Scholar] [CrossRef]
- Jolly, P.S.; Rosenfeldt, H.M.; Milstien, S.; Spiegel, S. The roles of sphingosine-1-phosphate in asthma. Mol. Immunol. 2002, 38, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Ammit, A.J.; Hastie, A.T.; Edsall, L.C.; Hoffman, R.K.; Amrani, Y.; Krymskaya, V.P.; Kane, S.A.; Peters, S.P.; Penn, R.B.; Spiegel, S.; et al. Sphingosine 1-phosphate modulates human airway smooth muscle cell functions that promote inflammation and airway remodeling in asthma. FASEB J. 2001, 15, 1212–1214. [Google Scholar] [CrossRef] [PubMed]
- Oskeritzian, C.A.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate in allergic responses, asthma and anaphylaxis. Pharmacol. Ther. 2007, 115, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Chiba, Y.; Suzuki, K.; Kurihara, E.; Uechi, M.; Sakai, H.; Misawa, M. Sphingosine-1-phosphate aggravates antigen-induced airway inflammation in mice. Open Respir. Med. J. 2010, 4, 82–85. [Google Scholar] [CrossRef]
- Price, M.M.; Oskeritzian, C.A.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate synthesis and functions in mast cells. Future Lipidol. 2008, 3, 665–674. [Google Scholar] [CrossRef][Green Version]
- Yatomi, Y.; Igarashi, Y.; Yang, L.; Hisano, N.; Qi, R.; Asazuma, N.; Satoh, K.; Ozaki, Y.; Kume, S. Sphingosine 1-phosphate, a bioactive sphingolipid abundantly stored in platelets, is a normal constituent of human plasma and serum. J. Biochem. 1997, 121, 969–973. [Google Scholar] [CrossRef]
- Xiong, Y.; Hla, T. S1P control of endothelial integrity. Curr. Top. Microbiol. Immunol. 2014, 378, 85–105. [Google Scholar] [CrossRef]
- Sakai, T.; Herrmann, N.; Maintz, L.; Nümm, T.J.; Welchowski, T.; Claus, R.A.; Gräler, M.H.; Bieber, T. Serum sphingosine-1-phosphate is elevated in atopic dermatitis and associated with severity. Allergy 2021, 76, 2592–2595. [Google Scholar] [CrossRef]
- Masuda-Kuroki, K.; Di Nardo, A. Sphingosine 1-Phosphate Signaling at the Skin Barrier Interface. Biology 2022, 11, 809. [Google Scholar] [CrossRef]
- Roviezzo, F.; Del Galdo, F.; Abbate, G.; Bucci, M.; D’Agostino, B.; Antunes, E.; de Dominicis, G.; Parente, L.; Rossi, F.; Cirino, G.; et al. Human eosinophil chemotaxis and selective in vivo recruitment by sphingosine 1-phosphate. Proc. Natl. Acad. Sci. USA 2004, 101, 11170–11175. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, M.; Costello, R.W.; Belmonte, K.E.; Gendy, S.S.; Walsh, M.-T. Neutrophil sphingosine 1-phosphate and lysophosphatidic acid receptors in pneumonia. Am. J. Respir. Cell Mol. Biol. 2006, 34, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Oskeritzian, C.A.; Alvarez, S.E.; Hait, N.C.; Price, M.M.; Milstien, S.; Spiegel, S. Distinct roles of sphingosine kinases 1 and 2 in human mast-cell functions. Blood 2008, 111, 4193–4200. [Google Scholar] [CrossRef]
- Oskeritzian, C.A.; Price, M.M.; Hait, N.C.; Kapitonov, D.; Falanga, Y.T.; Morales, J.K.; Ryan, J.J.; Milstien, S.; Spiegel, S. Essential roles of sphingosine-1-phosphate receptor 2 in human mast cell activation, anaphylaxis, and pulmonary edema. J. Exp. Med. 2010, 207, 465–474. [Google Scholar] [CrossRef]
- Aoki, M.; Aoki, H.; Ramanathan, R.; Hait, N.C.; Takabe, K. Sphingosine-1-Phosphate Signaling in Immune Cells and Inflammation: Roles and Therapeutic Potential. Mediat. Inflamm. 2016, 2016, 8606878. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, L.; Chang, N.; Hou, L.; Zhou, X.; Dong, C.; Liu, F.; Yang, L.; Li, L. Neutrophil recruitment mediated by sphingosine 1-phosphate (S1P)/S1P receptors during chronic liver injury. Cell. Immunol. 2021, 359, 104243. [Google Scholar] [CrossRef] [PubMed]
- Jolly, P.S.; Bektas, M.; Olivera, A.; Gonzalez-Espinosa, C.; Proia, R.L.; Rivera, J.; Milstien, S.; Spiegel, S. Transactivation of sphingosine-1-phosphate receptors by FcepsilonRI triggering is required for normal mast cell degranulation and chemotaxis. J. Exp. Med. 2004, 199, 959–970. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, L.; Chang, N.; Hou, L.; Zhou, X.; Yang, L.; Li, L. Neutrophils undergo switch of apoptosis to NETosis during murine fatty liver injury via S1P receptor 2 signaling. Cell Death Dis. 2020, 11, 379. [Google Scholar] [CrossRef]
- Bryan, A.M.; Del Poeta, M. Sphingosine-1-phosphate receptors and innate immunity. Cell. Microbiol. 2018, 20, e12836. [Google Scholar] [CrossRef]
- MacGlashan, D. Expression of CD203c and CD63 in human basophils: Relationship to differential regulation of piecemeal and anaphylactic degranulation processes. Clin. Exp. Allergy 2010, 40, 1365–1377. [Google Scholar] [CrossRef]
- Saraste, A. Morphologic criteria and detection of apoptosis. Herz 1999, 24, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Bonnaud, S.; Niaudet, C.; Legoux, F.; Corre, I.; Delpon, G.; Saulquin, X.; Fuks, Z.; Gaugler, M.-H.; Kolesnick, R.; Paris, F. Sphingosine-1-phosphate activates the AKT pathway to protect small intestines from radiation-induced endothelial apoptosis. Cancer Res. 2010, 70, 9905–9915. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.S.; Teegarden, D. Sphingosine-1-phosphate inhibition of apoptosis requires mitogen-activated protein kinase phosphatase-1 in mouse fibroblast C3H10T 1/2 cells. J. Nutr. 2003, 133, 3343–3349. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hagen, N.; van Veldhoven, P.P.; Proia, R.L.; Park, H.; Merrill, A.H.; van Echten-Deckert, G. Subcellular origin of sphingosine 1-phosphate is essential for its toxic effect in lyase-deficient neurons. J. Biol. Chem. 2009, 284, 11346–11353. [Google Scholar] [CrossRef] [PubMed]
- Shute, J. Basophil migration and chemotaxis. Clin. Exp. Allergy 1992, 22, 321–323. [Google Scholar] [CrossRef]
- Uguccioni, M.; Mackay, C.R.; Ochensberger, B.; Loetscher, P.; Rhis, S.; LaRosa, G.J.; Rao, P.; Ponath, P.D.; Baggiolini, M.; Dahinden, C.A. High expression of the chemokine receptor CCR3 in human blood basophils. Role in activation by eotaxin, MCP-4, and other chemokines. J. Clin. Investig. 1997, 100, 1137–1143. [Google Scholar] [CrossRef]
- Mommert, S.; Kleiner, S.; Gehring, M.; Eiz-Vesper, B.; Stark, H.; Gutzmer, R.; Werfel, T.; Raap, U. Human basophil chemotaxis and activation are regulated via the histamine H4 receptor. Allergy 2016, 71, 1264–1273. [Google Scholar] [CrossRef]
- Custovic, A. Epidemiology of Allergic Diseases. In Middleton’s Allergy Essentials; O’Hehir, R.E., Holgate, S.T., Sheikh, A., Eds.; Elsevier: Edinburgh, UK, 2017; pp. 51–72. ISBN 9780323375795. [Google Scholar]
- Yunginger, J.W.; Ahlstedt, S.; Eggleston, P.A.; Homburger, H.A.; Nelson, H.S.; Ownby, D.R.; Platts-Mills, T.A.; Sampson, H.A.; Sicherer, S.H.; Weinstein, A.M.; et al. Quantitative IgE antibody assays in allergic diseases. J. Allergy Clin. Immunol. 2000, 105, 1077–1084. [Google Scholar] [CrossRef]
- Salo, P.M.; Arbes, S.J.; Jaramillo, R.; Calatroni, A.; Weir, C.H.; Sever, M.L.; Hoppin, J.A.; Rose, K.M.; Liu, A.H.; Gergen, P.J.; et al. Prevalence of allergic sensitization in the United States: Results from the National Health and Nutrition Examination Survey (NHANES) 2005–2006. J. Allergy Clin. Immunol. 2014, 134, 350–359. [Google Scholar] [CrossRef]
- Sugita, K.; Kabashima, K.; Sakabe, J.-I.; Yoshiki, R.; Tanizaki, H.; Tokura, Y. FTY720 regulates bone marrow egress of eosinophils and modulates late-phase skin reaction in mice. Am. J. Pathol. 2010, 177, 1881–1887. [Google Scholar] [CrossRef]
- Finley, A.; Chen, Z.; Esposito, E.; Cuzzocrea, S.; Sabbadini, R.; Salvemini, D. Sphingosine 1-phosphate mediates hyperalgesia via a neutrophil-dependent mechanism. PLoS ONE 2013, 8, e55255. [Google Scholar] [CrossRef] [PubMed]
- Corbett, A.H. Post-transcriptional regulation of gene expression and human disease. Curr. Opin. Cell Biol. 2018, 52, 96–104. [Google Scholar] [CrossRef]
- Mali, S.S.; Bautista, D.M. Basophils add fuel to the flame of eczema itch. Cell 2021, 184, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, Y.; Mogi, K.; Takahashi, K.; Miyake, K.; Yoshikawa, S.; Karasuyama, H. Skin-infiltrating basophils promote atopic dermatitis-like inflammation via IL-4 production in mice. Allergy 2020, 75, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
- Braakman, S.; Perkumas, K.; Overby, D.; Woodward, D.; Stamer, D. Sphingosine-1-Phosphate signaling in cultured human trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1995. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gray, N.; Limberg, M.M.; Wiebe, D.; Weihrauch, T.; Langner, A.; Brandt, N.; Bräuer, A.U.; Raap, U. Differential Upregulation and Functional Activity of S1PR1 in Human Peripheral Blood Basophils of Atopic Patients. Int. J. Mol. Sci. 2022, 23, 16117. https://doi.org/10.3390/ijms232416117
Gray N, Limberg MM, Wiebe D, Weihrauch T, Langner A, Brandt N, Bräuer AU, Raap U. Differential Upregulation and Functional Activity of S1PR1 in Human Peripheral Blood Basophils of Atopic Patients. International Journal of Molecular Sciences. 2022; 23(24):16117. https://doi.org/10.3390/ijms232416117
Chicago/Turabian StyleGray, Natalie, Maren M. Limberg, Daniela Wiebe, Tobias Weihrauch, Anna Langner, Nicola Brandt, Anja U. Bräuer, and Ulrike Raap. 2022. "Differential Upregulation and Functional Activity of S1PR1 in Human Peripheral Blood Basophils of Atopic Patients" International Journal of Molecular Sciences 23, no. 24: 16117. https://doi.org/10.3390/ijms232416117
APA StyleGray, N., Limberg, M. M., Wiebe, D., Weihrauch, T., Langner, A., Brandt, N., Bräuer, A. U., & Raap, U. (2022). Differential Upregulation and Functional Activity of S1PR1 in Human Peripheral Blood Basophils of Atopic Patients. International Journal of Molecular Sciences, 23(24), 16117. https://doi.org/10.3390/ijms232416117