
Overview of Anti-Inflammatory and Anti-Nociceptive Effects of Polyphenols to Halt Osteoarthritis: From Preclinical Studies to New Clinical Insights
 , , ,
, , ,  ,
, 
Abstract
1. Introduction
Literature Search Strategy
2. OA Description: Focus on Inflammation and Pain
2.1. Molecular Signalling Pathways during Inflammation in Knee OA
2.2. Biological Basis of Pain in Knee OA
3. Traditional Therapeutic Strategies to Counteract Inflammation and Pain in OA: Pros and Cons
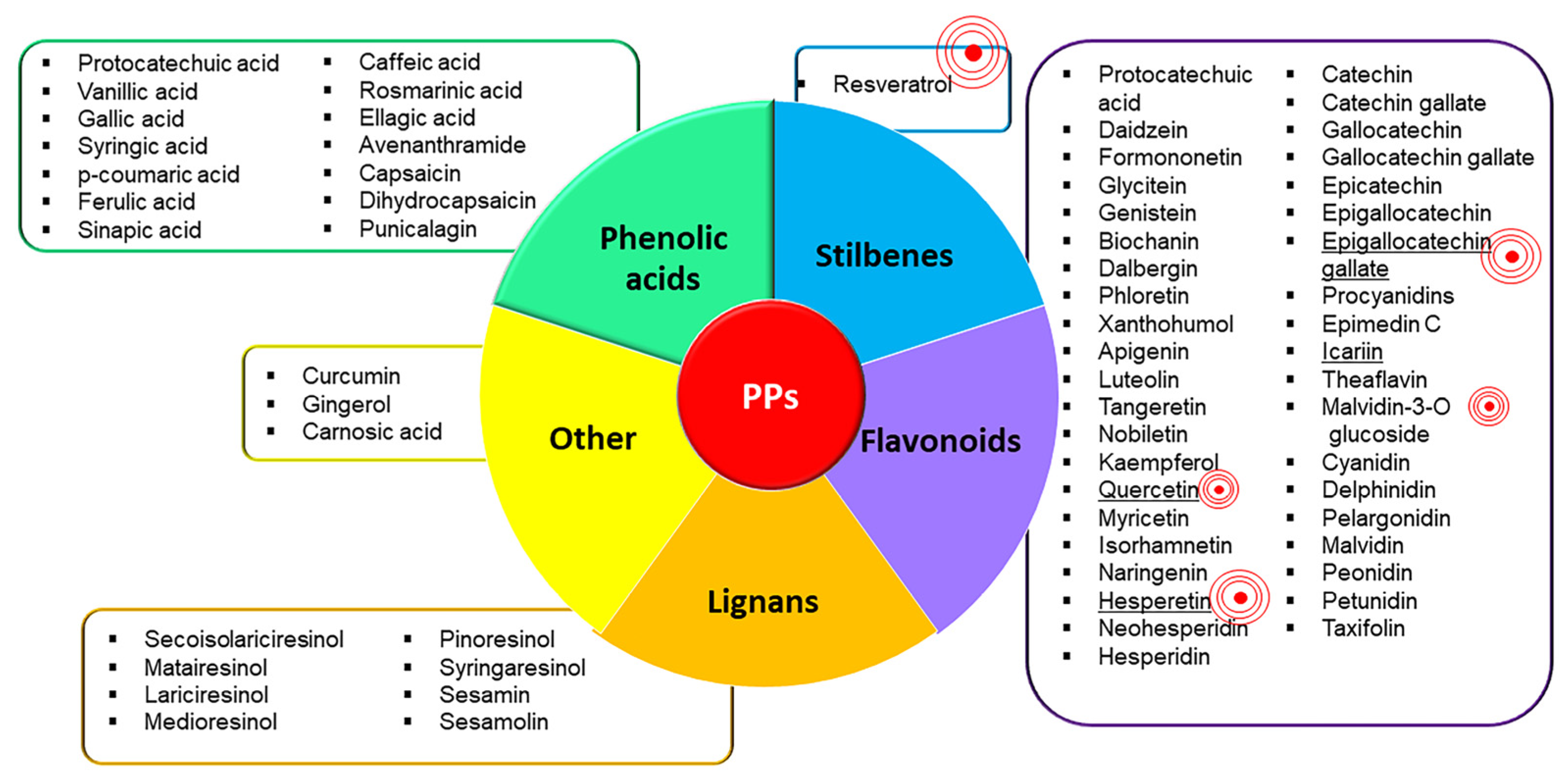
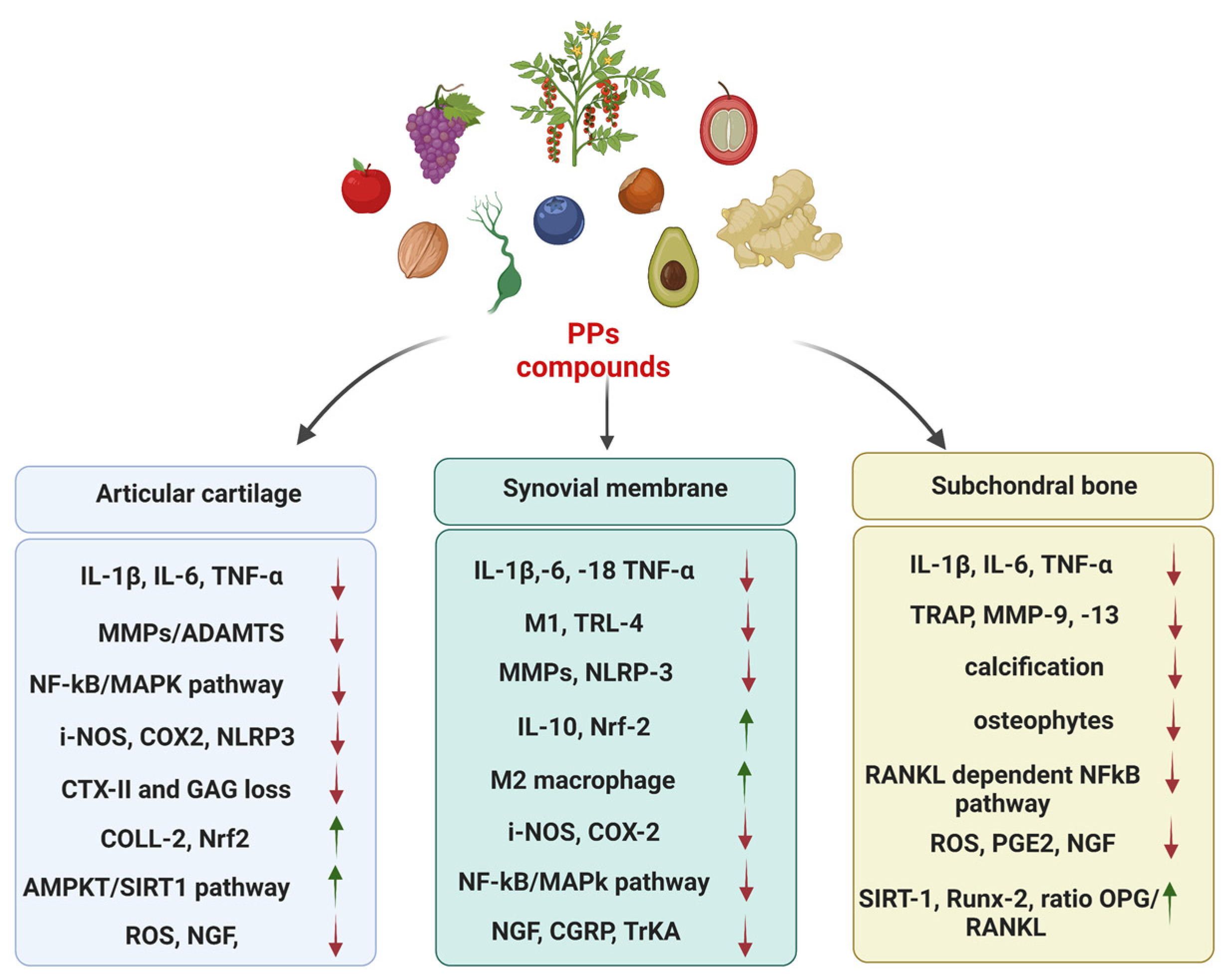
4. Efficacy and Safety of PPs in the Management of OA
Role of PPs in Modulating Inflammation and Pain: Focus on Preclinical In Vitro and In Vivo Studies
5. From Basic Research to Translational Applications of PPs
5.1. Clinical Studies on PPs for the Management of OA
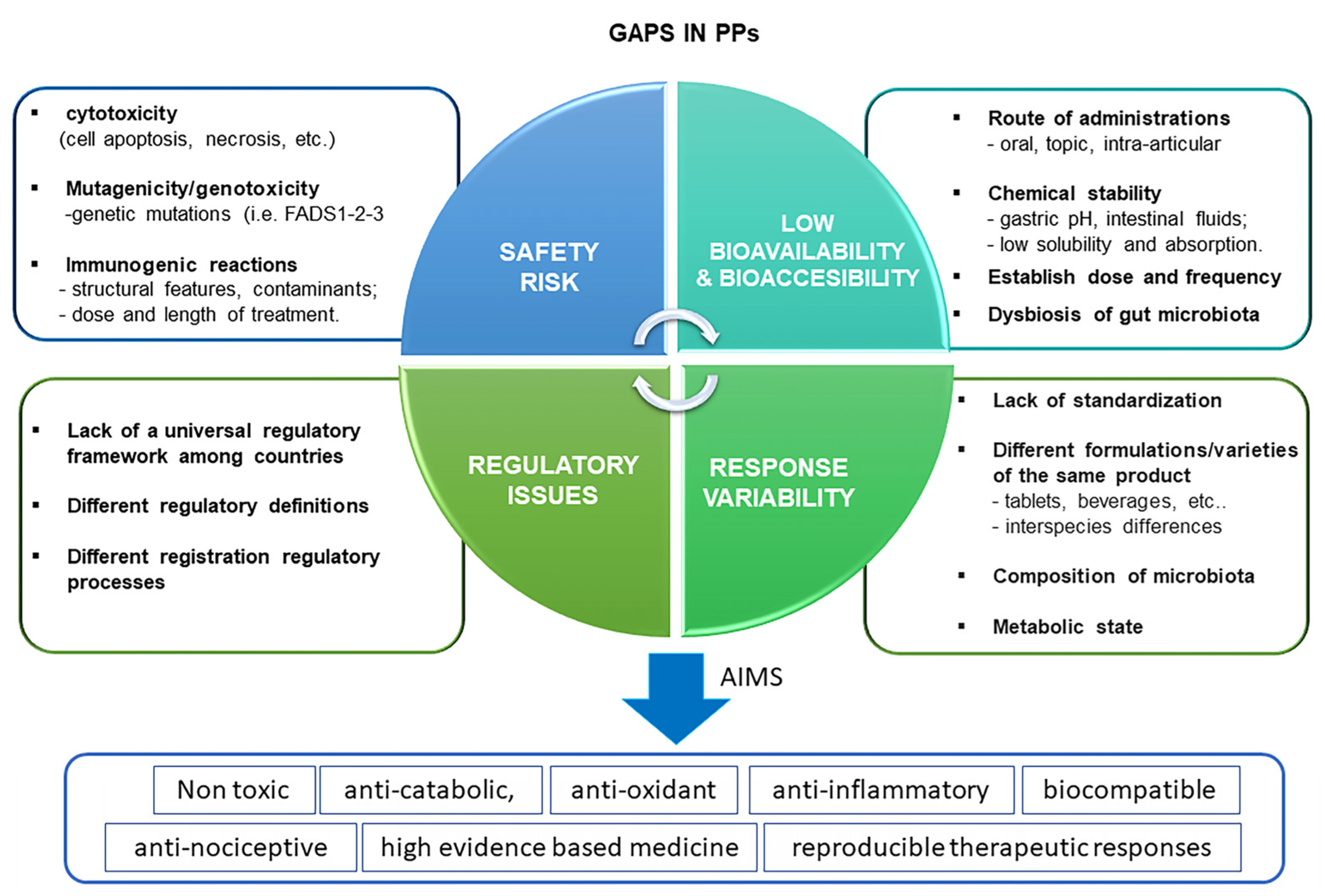
5.2. PPs: Research and Clinical Gaps in OA
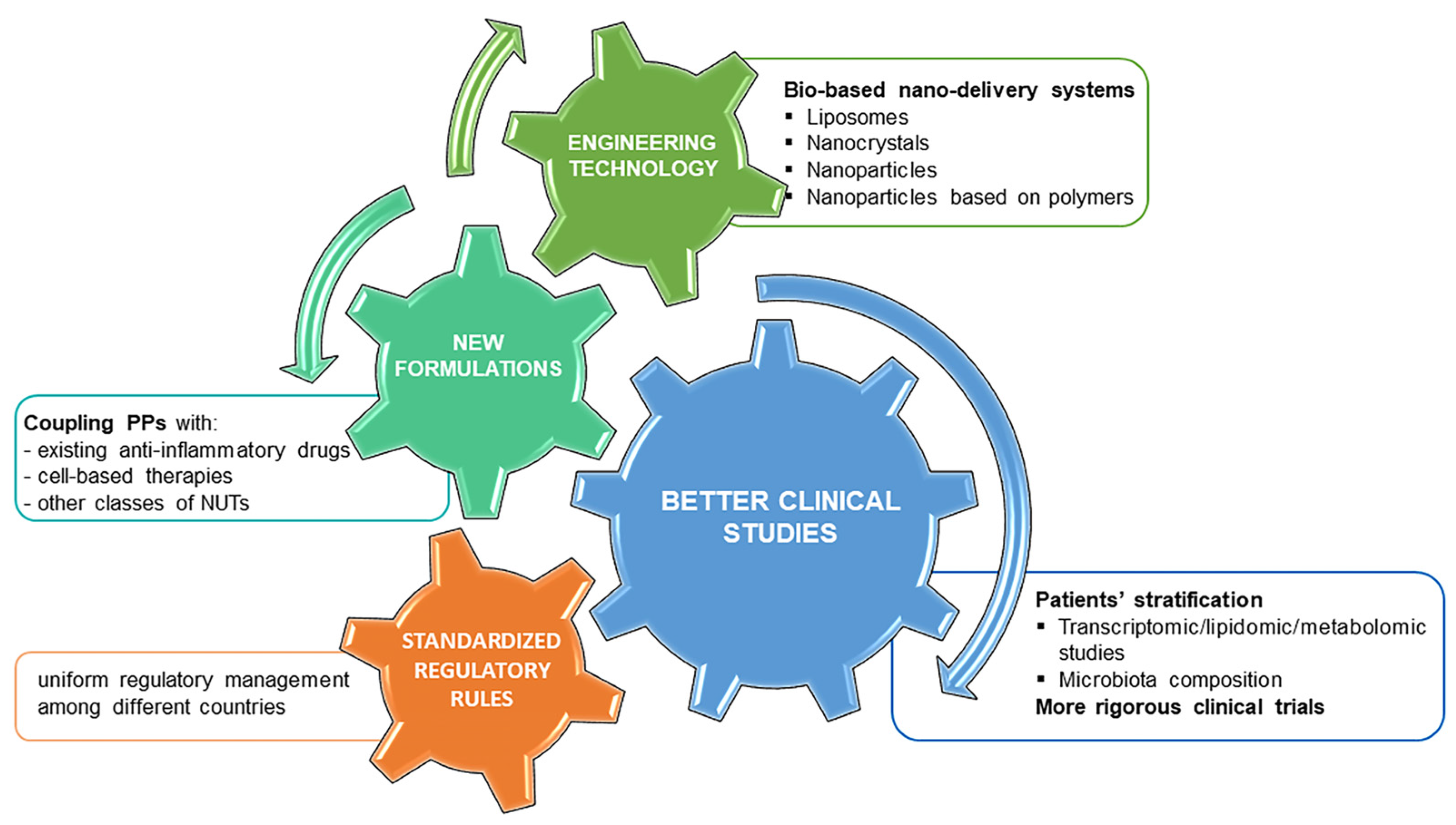
5.3. From Knowledge Gaps to New Opportunities: Perspectives of PPs in OA
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Acronym | Full-Extensive name |
| ACAN | Aggrecan |
| ADAMTs | a disintegrin and metalloproteinase with thrombospondin motifs |
| AGEs | advanced glycation end-products |
| Arg1 | arginase-1 |
| BCP | basic calcium phosphate |
| BDNF | brain-derived neurotrophic factor |
| BDMs | bioactive dietary molecules |
| BDNF | brain-derived neurotrophic factor |
| CCL | Chemokine (C-C motif) ligand |
| CFA | complete Freund’s adjuvant |
| CGRP | calcitonin gene-related peptide |
| ChPF | Chondroitin Polymerizing Factor |
| CHSY-1 | Chondroitin sulfate synthase 1 |
| COX-2 | cyclooxygenase-2 |
| CPPD | Calcium pyrophosphate dihydrate |
| CRP | C-reactive protein |
| CSPG | Chondroitin sulfate proteoglycans |
| CXCL | C-X-C Motif Chemokine Ligand |
| DAMPs | damage-associated molecular patterns |
| DEC | Decorin core protein |
| DRG | dorsal root ganglia |
| ECM | extracellular matrix |
| EFSA | European Food Safety Authority |
| ERK | extracellular signal regulated kinase |
| FMN | Formononetin |
| GABA | Gamma-aminobutyric acid |
| GAGs | Glycosamminoglycans |
| GSH | reduced glutathione |
| GSSG | oxidized disulfide form glutathione |
| GSTA4-4 | glutathione-s-tranferase A4-4 |
| H2O2 | produces hydrogen peroxide |
| HA | hyaluronic acid |
| HMGB1 | high-mobility group box-1 |
| HNE | 4-hydroxynonenal |
| HNE | 4-hydroxynonenal |
| IA | intra-articular |
| IL | Interleukin |
| IL1-R | interleukin-1 receptor |
| iNOS | inducible NO synthase |
| JNK | c-Jun NH2-terminal kinase |
| KOOS | Knee injury and Osteoarthritis Outcome Score |
| LGI | low-grade inflammation |
| MAPK | mitogen-activated protein kinase |
| MCP-1, CCL2 | monocyte chemoattractant protein-1 |
| MIA | mono-iodoacetate |
| miR | miRNA |
| MMPs | matrix metalloproteinases |
| MWT | mechanical withdrawal threshold |
| NE | nor-epinephrine |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NGF | nerve growth factor |
| NK | neurokinin |
| NLRP3 | nucleotide-binding oligomerization domain-like receptor protein 3 |
| NLRs | NOD-like receptors |
| NMDA | N-methyl-D-aspartate |
| NO | nitric oxide |
| NOX | NADPH Oxidase |
| NSAIDs | non-steroidal anti-inflammatory drugs |
| OA | osteoarthritis |
| OSM | oncostatin M |
| PG | Proteoglycans |
| PGE2 | prostaglandin E2 |
| PI3K/ AKT | phosphoinositide 3-kinase/protein kinase B |
| PLGA | poly lactic-co-glycolic acid |
| PPs | polyphenols |
| PRP | Platelet-rich plasma |
| PRRs | pattern recognition receptors |
| PTGS2 | prostaglandin-endoperoxide synthase 2 |
| PWL | paw withdrawal latency |
| PWT | paw withdrawal threshold |
| RAGEs | receptor for advanced glycosylation end products |
| ReHo | Regional homogeneity |
| ROM | Range of motion |
| ROS | Reactive oxygen species |
| Rsv | Resveratol |
| Runx-2 | runt-related transcription factor 2 |
| SASP | senescence-associated secretory phenotype |
| Sox-9 | SRY-box transcription factor 9 |
| SP | substance P |
| TGF- β | transforming growth factor β |
| TBHP | tert-butylhydroperoxide |
| TKA | total knee arthroplasty |
| TLR | toll-like receptors |
| TNF-α | tumor necrosis factor-α |
| TRIF | TIRdomain-containing adaptor-inducing interferon-β |
| TrK | tropomyosin receptor kinase |
| TRP | Transient receptor potential |
| TrpV1 | Transient receptor potential cation channel subfamily V member 1 |
| TWL | thermal withdrawal latency |
| VAS | Visual Analogh scale |
| VEGF | vascular endothelial growth factor |
| WOMAC | Western Ontario and McMaster Universities Arthritis Index |
| XT | Xylosyltransferase |
| Ym1 | chitinase 3-like protein 3 |
References
- Cross, M.; Smith, E.; Hoy, D.; Nolte, S.; Ackerman, I.; Fransen, M.; Bridgett, L.; Williams, S.; Guillemin, F.; Hill, C.L.; et al. The global burden of hip and knee osteoarthritis: Estimates from the global burden of disease 2010 study. Ann. Rheum. Dis. 2014, 73, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Van Spil, W.E.; Kubassova, O.; Boesen, M.; Bay Jensen, A.C.; Mobasheri, A. Osteoarthritis phenotypes and novel therapeutic targets. Biochem. Pharmacol. 2019, 165, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Conaghan, P.G.; Cook, A.D.; Hamilton, J.A.; Tak, P.P. Therapeutic options for targeting inflammatory osteoarthritis pain. Nat. Rev. Rheumatol. 2019, 15, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Castrogiovanni, P.; Trovato, F.M.; Loreto, C.; Nsir, H.; Szychlinska, M.A.; Musumeci, M. Nutraceutical Supplements in the Management and Prevention of Osteoarthritis. Int. J. Mol. Sci. 2016, 17, 2042. [Google Scholar] [CrossRef] [PubMed]
- Larussa, T.; Imeneo, M.; Luzza, F. Potential role of nutraceutical compounds in inflammatory bowel disease. World J. Gastroenterol. 2017, 23, 2483–2492. [Google Scholar] [CrossRef]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary Anti-Aging Polyphenols and Potential Mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Tayab, M.A.; Islam, M.N.; Chowdhury, K.A.A.; Tasnim, F.M. Targeting neuroinflammation by polyphenols: A promising therapeutic approach against inflammation-associated depression. Biomed. Pharmacother. 2022, 147, 112668. [Google Scholar]
- Iqubal, A.; Ahmed, M.; Iqubal, M.K.; Pottoo, F.; Haque, S.E. Polyphenols as Potential Therapeutics for Pain and Inflammation in Spinal Cord Injury. Curr. Mol. Pharmacol. 2021, 14, 714–730. [Google Scholar] [CrossRef]
- D’Adamo, S.; Cetrullo, S.; Panichi, V.; Mariani, E.; Flamigni, F.; Borzì, R.M.B. Nutraceutical Activity in Osteoarthritis Biology: A Focus on the Nutrigenomic Role. Cells 2020, 9, 1232. [Google Scholar] [CrossRef]
- Liu, X.; Machado, G.C.; Eyles, J.P.; Ravi, V.; Hunter, D.J. Dietary supplements for treating osteoarthritis: A systematic review and meta-analysis. Br. J. Sports Med. 2018, 52, 167–175. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Ebata, T.; Yokota, S.; Takahashi, D.; Endo, T.; Matsumae, G.; Shimizu, T.; Kadoya, K.; Iwasaki, N. Low-Grade Inflammation in the Pathogenesis of Osteoarthritis: Cellular and Molecular Mechanisms and Strategies for Future Therapeutic Intervention. Biomedicines 2022, 10, 1109. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Lopez, E.; Coras, R.; Torres, A.; Lane, N.E.; Guma, M. Synovial inflammation in osteoarthritis progression. Nat. Rev. Rheumatol. 2022, 18, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zuo, Z.; Kuang, Y. An Emerging Target in the Battle against Osteoarthritis: Macrophage Polarization. Int. J. Mol. Sci. 2020, 21, 8513. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Yu, Y.; Zhuang, Q.; Wang, L.; Zhan, B.; Du, S.; Liu, Y.; Huang, J.; Hao, J.; Zhu, X. Bone erosion in inflammatory arthritis is attenuated by Trichinella spiralis through inhibiting M1 monocyte/macrophage polarization. iScience 2022, 25, 103979. [Google Scholar] [CrossRef] [PubMed]
- Frevert, C.W.; Felgenhauer, J.; Wygrecka, M.; Nastase, M.V.; Schaefer, L. Danger-Associated Molecular Patterns Derived From the Extracellular Matrix Provide Temporal Control of Innate Immunity. J. Histochem. Cytochem. 2018, 66, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Sofat, N. Analysing the role of endogenous matrix molecules in the development of osteoarthritis. Int. J. Exp. Pathol. 2009, 90, 463–479. [Google Scholar] [CrossRef]
- Sokolove, J.; Lepus, C.M. Role of inflammation in the pathogenesis of osteoarthritis: Latest findings and interpretations. Ther. Adv. Musculoskelet. Dis. 2013, 5, 77–94. [Google Scholar] [CrossRef]
- Schelbergen, R.F.; Blom, A.B.; van den Bosch, M.H.; Sloetjes, A.; Abdollahi-Roodsaz, S.; Schreurs, W.; Mort, J.S.; Vogl, T.; Roth, J.; van den Berg, W.B.; et al. Alarmins S100A8 and S100A9 elicit a catabolic effect in human osteoarthritic chondrocytes that is dependent on Toll-like receptor 4. Arthritis Rheum. 2012, 64, 1477–1487. [Google Scholar] [CrossRef]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef]
- Yang, S.; Kim, J.; Ryu, J.H.; Oh, H.; Chun, C.H.; Kim, B.J.; Min, B.H.; Chun, J.S. Hypoxia-inducible factor-2alpha is a catabolic regulator of osteoarthritic cartilage destruction. Nat. Med. 2010, 16, 687–693. [Google Scholar] [CrossRef]
- Shen, J.; Abu-Amer, Y.; O’Keefe, R.J.; McAlinden, A. Inflammation and epigenetic regulation in osteoarthritis. Connect. Tissue Res. 2017, 58, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Grigolo, B.; Roseti, L.; Fiorini, M.; Facchini, A. Enhanced lipid peroxidation in synoviocytes from patients with osteoarthritis. J. Rheumatol. 2003, 30, 345–347. [Google Scholar] [PubMed]
- Boehme, K.A.; Rolauffs, B. Onset and Progression of Human Osteoarthritis-Can Growth Factors, Inflammatory Cytokines, or Differential miRNA Expression Concomitantly Induce Proliferation, ECM Degradation, and Inflammation in Articular Cartilage? Int. J. Mol. Sci. 2018, 19, 2282. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, M.J.; Ferrandiz, M.L. Relevance of Nrf2 and heme oxygenase-1 in articular diseases. Free Radic. Biol. Med. 2020, 157, 83–93. [Google Scholar] [CrossRef]
- Liu, F.C.; Wang, C.C.; Lu, J.W.; Lee, C.H.; Chen, S.C.; Ho, Y.J.; Peng, Y.J. Chondroprotective Effects of Genistein against Osteoarthritis Induced Joint Inflammation. Nutrients 2019, 11, 1180. [Google Scholar] [CrossRef]
- Abusarah, J.; Benabdoune, H.; Shi, Q.; Lussier, B.; Martel-Pelletier, J.; Malo, M.; Fernandes, J.C.; de Souza, F.P.; Fahmi, H.; Benderdour, M. Elucidating the Role of Protandim and 6-Gingerol in Protection Against Osteoarthritis. J. Cell Biochem. 2017, 118, 1003–1013. [Google Scholar] [CrossRef]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-kappaB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Roberti, A.; Chaffey, L.E.; Greaves, D.R. NF-kappaB Signaling and Inflammation-Drug Repurposing to Treat Inflammatory Disorders? Biology 2022, 11, 372. [Google Scholar] [CrossRef]
- Rigoglou, S.; Papavassiliou, A.G. The NF-kappaB signalling pathway in osteoarthritis. Int. J. Biochem. Cell Biol. 2013, 45, 2580–2584. [Google Scholar] [CrossRef]
- Roman-Blas, J.A.; Jimenez, S.A. NF-kappaB as a potential therapeutic target in osteoarthritis and rheumatoid arthritis. Osteoarthr. Cartil. 2006, 14, 839–848. [Google Scholar] [CrossRef]
- Xie, J.; Lin, J.; Wei, M.; Teng, Y.; He, Q.; Yang, G.; Yang, X. Sustained Akt signaling in articular chondrocytes causes osteoarthritis via oxidative stress-induced senescence in mice. Bone Res. 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Penas, C.; Navarro, X. Epigenetic Modifications Associated to Neuroinflammation and Neuropathic Pain After Neural Trauma. Front. Cell. Neurosci. 2018, 12, 158. [Google Scholar] [CrossRef]
- den Hollander, W.; Meulenbelt, I. DNA Methylation in Osteoarthritis. Curr. Genomics 2015, 16, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulos, P.K.; Lambrou, G.I. The Involvement of MicroRNAs in Osteoarthritis and Recent Developments: A Narrative Review. Mediterr. J. Rheumatol. 2018, 29, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Zhang, F.; Yao, H.; Li, H.; Tuan, R.S. Histone Modifications and Chondrocyte Fate: Regulation and Therapeutic Implications. Front. Cell Dev. Biol. 2021, 9, 626708. [Google Scholar] [CrossRef]
- Deng, Z.; Li, Y.; Liu, H.; Xiao, S.; Li, L.; Tian, J.; Cheng, C.; Zhang, G.; Zhang, F. The role of sirtuin 1 and its activator, resveratrol in osteoarthritis. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Oppenheimer, H.; Gabay, O.; Meir, H.; Haze, A.; Kandel, L.; Liebergall, M.; Gagarina, V.; Lee, E.J.; Dvir-Ginzberg, M. 75-kd sirtuin 1 blocks tumor necrosis factor alpha-mediated apoptosis in human osteoarthritic chondrocytes. Arthritis Rheum. 2012, 64, 718–728. [Google Scholar] [CrossRef]
- Dvir-Ginzberg, M.; Gagarina, V.; Lee, E.J.; Booth, R.; Gabay, O.; Hall, D.J. Tumor necrosis factor alpha-mediated cleavage and inactivation of SirT1 in human osteoarthritic chondrocytes. Arthritis Rheum. 2011, 63, 2363–2373. [Google Scholar] [CrossRef]
- Takayama, K.; Ishida, K.; Matsushita, T.; Fujita, N.; Hayashi, S.; Sasaki, K.; Tei, K.; Kubo, S.; Matsumoto, T.; Fujoka, H. SIRT1 regulation of apoptosis of human chondrocytes. Arthritis Rheum. 2009, 60, 2731–2740. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, W.; Wu, P.; Deng, Z.; Zeng, C.; Li, H.; Yang, T.; Lei, G. The expression of SIRT1 in articular cartilage of patients with knee osteoarthritis and its correlation with disease severity. J. Orthop. Surg. Res. 2016, 11, 144. [Google Scholar] [CrossRef]
- Greene, M.A.; Loeser, R.F. Aging-related inflammation in osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1966–1971. [Google Scholar] [CrossRef] [PubMed]
- Chisari, E.; Wouthuyzen-Bakker, M.; Friedrich, A.W.; Parvizi, J. The relation between the gut microbiome and osteoarthritis: A systematic review of literature. PLoS ONE 2021, 16, e0261353. [Google Scholar] [CrossRef] [PubMed]
- Neogi, T. The epidemiology and impact of pain in osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, T.W.; Felson, D.T. Mechanisms of Osteoarthritis (OA) Pain. Curr. Osteoporos. Rep. 2018, 16, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Gronblad, M.; Liesi, P.; Korkala, O.; Karaharju, E.; Polak, J. Innervation of human bone periosteum by peptidergic nerves. Anat. Rec. 1984, 209, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Mapp, P.I. Innervation of the synovium. Ann. Rheum. Dis. 1995, 54, 398–403. [Google Scholar] [CrossRef]
- Ashraf, S.; Wibberley, H.; Mapp, P.I.; Hill, R.; Wilson, D.; Walsh, D.A. Increased vascular penetration and nerve growth in the meniscus: A potential source of pain in osteoarthritis. Ann. Rheum. Dis. 2011, 70, 523–529. [Google Scholar] [CrossRef]
- Hirasawa, Y.; Okajima, S.; Ohta, M.; Tokioka, T. Nerve distribution to the human knee joint: Anatomical and immunohistochemical study. Int. Orthop. 2000, 24, 1–4. [Google Scholar] [CrossRef]
- Suri, S.; Gill, S.E.; Massena de Camin, S.; Wilson, D.; McWilliams, D.F.; Walsh, D.A. Neurovascular invasion at the osteochondral junction and in osteophytes in osteoarthritis. Ann. Rheum. Dis. 2007, 66, 1423–1428. [Google Scholar] [CrossRef]
- Nees, T.A.; Rosshirt, N.; Zhang, J.A.; Reiner, T.; Sorbi, R.; Tripel, E.; Walker, T.; Schiltenwolf, M.; Hagmann, S.; Moradi, B. Synovial Cytokines Significantly Correlate with Osteoarthritis-Related Knee Pain and Disability: Inflammatory Mediators of Potential Clinical Relevance. J. Clin. Med. 2019, 8, 1343. [Google Scholar] [CrossRef]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef]
- Takayama, Y.; Derouiche, S.; Maruyama, K.; Tominaga, M. Emerging Perspectives on Pain Management by Modulation of TRP Channels and ANO1. Int. J. Mol. Sci. 2019, 20, 3411. [Google Scholar] [CrossRef]
- Yam, M.F.; Loh, Y.C.; Tan, C.S.; Khadijah Adam, S.; Abdul Manan, N.; Basir, R. General Pathways of Pain Sensation and the Major Neurotransmitters Involved in Pain Regulation. Int. J. Mol. Sci. 2018, 19, 2164. [Google Scholar] [CrossRef]
- Liu, Y.; Peng, H.; Meng, Z.; Wei, M. Correlation of IL-17 Level in Synovia and Severity of Knee Osteoarthritis. Med. Sci. Monit. 2015, 21, 1732–1736. [Google Scholar]
- Pan, F.; Tian, J.; Cicuttini, F.; Jones, G. Prospective Association Between Inflammatory Markers and Knee Cartilage Volume Loss and Pain Trajectory. Pain Ther. 2022, 11, 107–119. [Google Scholar] [CrossRef]
- Vincent, T.L. Peripheral pain mechanisms in osteoarthritis. Pain 2020, 161 (Suppl. S1), S138–S146. [Google Scholar] [CrossRef]
- Hirose, M.; Kuroda, Y.; Murata, E. NGF/TrkA Signaling as a Therapeutic Target for Pain. Pain Pract. 2016, 16, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, Y.; Liu, R.; Li, W.; Hua, B.; Bao, Y. Wnt Signaling Pathways: A Role in Pain Processing. Neuromolecular Med. 2022, 24, 233–249. [Google Scholar] [CrossRef]
- Ji, R.R.; Gereau, R.W.T.; Malcangio, M.; Strichartz, G.R. MAP kinase and pain. Brain Res. Rev. 2009, 60, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A narrative review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Lories, R.J.; Monteagudo, S. Review Article: Is Wnt Signaling an Attractive Target for the Treatment of Osteoarthritis? Rheumatol. Ther. 2020, 7, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Houweling, P.; Kulkarni, R.N.; Baldock, P.A. Neuronal control of bone and muscle. Bone 2015, 80, 95–100. [Google Scholar] [CrossRef]
- Mlost, J.; Wasik, A.; Michaluk, J.T.; Antkiewicz-Michaluk, L.; Starowicz, K. Changes in Monoaminergic Neurotransmission in an Animal Model of Osteoarthritis: The Role of Endocannabinoid Signaling. Front. Mol. Neurosci. 2018, 11, 466. [Google Scholar] [CrossRef] [PubMed]
- Larson, C.M.; Wilcox, G.L.; Fairbanks, C.A. The Study of Pain in Rats and Mice. Comp. Med. 2019, 69, 555–570. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Dubner, R. Interactions between the immune and nervous systems in pain. Nat. Med. 2010, 16, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.S.; Xu, B.X.; Fan, K.J.; Li, Y.W.; Wu, J.; Wang, T.Y. Dexamethasone-Loaded Thermosensitive Hydrogel Suppresses Inflammation and Pain in Collagen-Induced Arthritis Rats. Drug Des. Devel. Ther. 2020, 14, 4101–4113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Lu, S.; Yu, A.; Fu, Z. Increased pain in response to mechanical or thermal stimulation in a rat model of incision-induced pain with nicotine dependence and withdrawal. Exp. Ther. Med. 2013, 5, 1063–1066. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gregory, N.S.; Harris, A.L.; Robinson, C.R.; Dougherty, P.M.; Fuchs, P.N.; Sluka, K.A. An overview of animal models of pain: Disease models and outcome measures. J. Pain 2013, 14, 1255–1269. [Google Scholar]
- Piel, M.J.; Kroin, J.S.; van Wijnen, A.J.; Kc, R.; Im, H.J. Pain assessment in animal models of osteoarthritis. Gene 2014, 537, 184–188. [Google Scholar] [CrossRef]
- Deng, Z.H.; Zeng, C.; Yang, Y.; Li, Y.S.; Wei, J.; Yang, T.; Li, H.; Lei, G.H. Topical diclofenac therapy for osteoarthritis: A meta-analysis of randomized controlled trials. Clin. Rheumatol. 2016, 35, 1253–1261. [Google Scholar] [CrossRef]
- da Costa, B.R.; Reichenbach, S.; Keller, N.; Nartey, L.; Wandel, S.; Jüni, P.; Trelle, S. Effectiveness of non-steroidal anti-inflammatory drugs for the treatment of pain in knee and hip osteoarthritis: A network meta-analysis. Lancet 2017, 390, e21–e33. [Google Scholar] [CrossRef] [PubMed]
- Burian, M.; Geisslinger, G. COX-dependent mechanisms involved in the antinociceptive action of NSAIDs at central and peripheral sites. Pharmacol. Ther. 2005, 107, 139–154. [Google Scholar] [CrossRef]
- Mukherjee, D.; Nissen, S.E.; Topol, E.J. Risk of cardiovascular events associated with selective COX-2 inhibitors. JAMA 2001, 286, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Honvo, G.; Leclercq, V.; Geerinck, A.; Thomas, T.; Veronese, N.; Charles, A.; Rabenda, V.; Beaudart, C.; Cooper, C.; Reginster, J.Y.; et al. Safety of Topical Non-steroidal Anti-Inflammatory Drugs in Osteoarthritis: Outcomes of a Systematic Review and Meta-Analysis. Drugs Aging 2019, 36, 45–64. [Google Scholar] [CrossRef] [PubMed]
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef]
- Osani, M.C.; Lohmander, L.S.; Bannuru, R.R. Is There Any Role for Opioids in the Management of Knee and Hip Osteoarthritis? A Systematic Review and Meta-Analysis. Arthritis Care Res. 2021, 73, 1413–1424. [Google Scholar] [CrossRef]
- Abdel Shaheed, C.; Awal, W.; Zhang, G.; Gilbert, S.E.; Gallacher, D.; McLachlan, A.; Day, R.O.; Ferreira, G.E.; Jones, C.M.P.; Ahedi, H.; et al. Efficacy, safety, and dose-dependence of the analgesic effects of opioid therapy for people with osteoarthritis: Systematic review and meta-analysis. Med. J. Aust. 2022, 216, 305–311. [Google Scholar] [CrossRef]
- Presti, D.; Scott, J.E. Hyaluronan-mediated protective effect against cell damage caused by enzymatically produced hydroxyl (OH.) radicals is dependent on hyaluronan molecular mass. Cell Biochem. Funct. 1994, 12, 281–288. [Google Scholar] [CrossRef]
- Julovi, S.M.; Yasuda, T.; Shimizu, M.; Hiramitsu, T.; Nakamura, T. Inhibition of interleukin-1beta-stimulated production of matrix metalloproteinases by hyaluronan via CD44 in human articular cartilage. Arthritis Rheum. 2004, 50, 516–525. [Google Scholar] [CrossRef]
- Moreland, L.W. Intra-articular hyaluronan (hyaluronic acid) and hylans for the treatment of osteoarthritis: Mechanisms of action. Arthritis Res. Ther. 2003, 5, 54–67. [Google Scholar] [CrossRef]
- Wu, Y.Z.; Huang, H.T.; Ho, C.J.; Shih, C.L.; Chen, C.H.; Cheng, T.L.; Wang, Y.C.; Lin, S.Y. Molecular Weight of Hyaluronic Acid Has Major Influence on Its Efficacy and Safety for Viscosupplementation in Hip Osteoarthritis: A Systematic Review and Meta-Analysis. Cartilage 2021, 13, 169S–184S. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, A.; Di Matteo, B.; Tentoni, F.P.; Selleri, F.; Cenacchi, A.; Kon, E.; Filardo, G. Platelet-Rich Plasma Versus Hyaluronic Acid Injections for the Treatment of Knee Osteoarthritis: Results at 5 Years of a Double-Blind, Randomized Controlled Trial. Am. J. Sports Med. 2019, 47, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Dragoo, J.L.; Danial, C.M.; Braun, H.J.; Pouliot, M.A.; Kim, H.J. The chondrotoxicity of single-dose corticosteroids. Knee Surg. Sports Traumatol. Arthrosc. 2012, 20, 1809–1814. [Google Scholar] [CrossRef] [PubMed]
- Elksnins-Finogejevs, A.; Vidal, L.; Peredistijs, A. Intra-articular platelet-rich plasma vs. corticosteroids in the treatment of moderate knee osteoarthritis: A single-center prospective randomized controlled study with a 1-year follow up. J. Orthop. Surg. Res. 2020, 15, 257. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomas-Barberan, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Milagro, F.I.; Riezu-Boj, J.I.; Martinez, J.A. Epigenetic signatures underlying inflammation: An interplay of nutrition, physical activity, metabolic diseases, and environmental factors for personalized nutrition. Inflamm. Res. 2021, 70, 29–49. [Google Scholar] [CrossRef]
- Liu, L.; Gu, H.; Liu, H.; Jiao, Y.; Li, K.; Zhao, Y.; An, L.; Yang, J. Protective effect of resveratrol against IL-1beta-induced inflammatory response on human osteoarthritic chondrocytes partly via the TLR4/MyD88/NF-kappaB signaling pathway: An “in vitro study”. Int. J. Mol. Sci. 2014, 15, 6925–6940. [Google Scholar] [CrossRef]
- Gu, H.; Jiao, Y.; Yu, X.; Li, X.; Wang, W.; Ding, L.; Liu, L. Resveratrol inhibits the IL-1beta-induced expression of MMP-13 and IL-6 in human articular chondrocytes via TLR4/MyD88-dependent and -independent signaling cascades. Int. J. Mol. Med. 2017, 39, 734–740. [Google Scholar] [CrossRef]
- Xu, X.; Liu, X.; Yang, Y.; He, J.; Jiang, M.; Huang, Y.; Liu, X.; Liu, L.; Gu, H. Resveratrol Exerts Anti-Osteoarthritic Effect by Inhibiting TLR4/NF-kappaB Signaling Pathway via the TLR4/Akt/FoxO1 Axis in IL-1beta-Stimulated SW1353 Cells. Drug Des. Devel. Ther. 2020, 14, 2079–2090. [Google Scholar] [CrossRef]
- Liu, F.C.; Hung, L.F.; Wu, W.L.; Chang, D.M.; Huang, C.Y.; Lai, J.H.; Ho, L.J. Chondroprotective effects and mechanisms of resveratrol in advanced glycation end products-stimulated chondrocytes. Arthritis Res. Ther. 2010, 12, R167. [Google Scholar] [CrossRef] [PubMed]
- Csaki, C.; Keshishzadeh, N.; Fischer, K.; Shakibaei, M. Regulation of inflammation signalling by resveratrol in human chondrocytes in vitro. Biochem. Pharmacol. 2008, 75, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Braun, H.J.; Dragoo, J.L. The effect of resveratrol on normal and osteoarthritic chondrocyte metabolism. Bone Joint. Res. 2014, 3, 51–59. [Google Scholar] [CrossRef]
- Liang, C.; Xing, H.; Wang, C.; Xu, X.; Hao, Y.; Qiu, B. Resveratrol protection against IL-1beta-induced chondrocyte damage via the SIRT1/FOXO1 signaling pathway. J. Orthop. Surg. Res. 2022, 17, 406. [Google Scholar] [CrossRef]
- Li, W.; Cai, L.; Zhang, Y.; Cui, L.; Shen, G. Intra-articular resveratrol injection prevents osteoarthritis progression in a mouse model by activating SIRT1 and thereby silencing HIF-2alpha. J. Orthop. Res. 2015, 33, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Zhang, W.; Cui, S.Y.; Fan, J.B.; Zhu, X.H.; Liu, W. Identification and validation of key long non-coding RNAs in resveratrol protect against IL-1beta-treated chondrocytes via integrated bioinformatic analysis. J. Orthop. Surg. Res. 2021, 16, 421. [Google Scholar] [CrossRef] [PubMed]
- Limagne, E.; Lancon, A.; Delmas, D.; Cherkaoui-Malki, M.; Latruffe, N. Resveratrol Interferes with IL1-beta-Induced Pro-Inflammatory Paracrine Interaction between Primary Chondrocytes and Macrophages. Nutrients 2016, 8, 280. [Google Scholar] [CrossRef] [PubMed]
- Siard, M.H.; McMurry, K.E.; Adams, A.A. Effects of polyphenols including curcuminoids, resveratrol, quercetin, pterostilbene, and hydroxypterostilbene on lymphocyte pro-inflammatory cytokine production of senior horses in vitro. Vet. Immunol. Immunopathol. 2016, 173, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Cao, G.; Ba, X.; Jiang, H. Epigallocatechin-3-O-gallate promotes extracellular matrix and inhibits inflammation in IL-1beta stimulated chondrocytes by the PTEN/miRNA-29b pathway. Pharm. Biol. 2022, 60, 589–599. [Google Scholar] [CrossRef]
- Akhtar, N.; Haqqi, T.M. Epigallocatechin-3-gallate suppresses the global interleukin-1beta-induced inflammatory response in human chondrocytes. Arthritis Res. Ther. 2011, 13, R93. [Google Scholar] [CrossRef]
- Rasheed, Z.; Rasheed, N.; Al-Shobaili, H.A. Epigallocatechin-3-O-gallate up-regulates microRNA-199a-3p expression by down-regulating the expression of cyclooxygenase-2 in stimulated human osteoarthritis chondrocytes. J. Cell Mol. Med. 2016, 20, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Ahmed, S.; Malemud, C.J.; Goldberg, V.M.; Haqqi, T.M. Epigallocatechin-3-gallate selectively inhibits interleukin-1beta-induced activation of mitogen activated protein kinase subgroup c-Jun N-terminal kinase in human osteoarthritis chondrocytes. J. Orthop. Res. 2003, 21, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, F.; Sfriso, P.; Scanu, A.; Fiocco, U.; Spinella, P.; Punzi, L. Epigallocatechin-3-gallate reduces inflammation induced by calcium pyrophosphate crystals in vitro. Front. Pharmacol. 2013, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Chen, Z.; Pengcheng, L.; Zhang, S.; Wang, X. Quercetin attenuates oxidative stress-induced apoptosis via SIRT1/AMPK-mediated inhibition of ER stress in rat chondrocytes and prevents the progression of osteoarthritis in a rat model. J. Cell Physiol. 2019, 234, 18192–18205. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Tang, Y.; Lu, H.; Qi, Y.; Li, G.; He, H.; Lu, F.; Yang, Y.; Sun, H. Quercetin Alleviates Osteoarthritis Progression in Rats by Suppressing Inflammation and Apoptosis via Inhibition of IRAK1/NLRP3 Signaling. J. Inflamm. Res. 2021, 14, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.P.; Xie, W.P.; Bi, Y.F.; Wang, B.A.; Song, H.B.; Wang, S.L.; Bi, R.X. Quercetin suppresses apoptosis of chondrocytes induced by IL-1beta via inactivation of p38 MAPK signaling pathway. Exp. Ther. Med. 2021, 21, 468. [Google Scholar] [CrossRef]
- Hu, Y.; Gui, Z.; Zhou, Y.; Xia, L.; Lin, K.; Xu, Y. Quercetin alleviates rat osteoarthritis by inhibiting inflammation and apoptosis of chondrocytes, modulating synovial macrophages polarization to M2 macrophages. Free Radic. Biol. Med. 2019, 145, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Fu, C.; Yan, Z.; Wu, Y.; Zhan, J.; Lou, Z.; Liao, X.; Pan, J. The protective effect of hesperetin in osteoarthritis: An in vitro and in vivo study. Food Funct. 2020, 11, 2654–2666. [Google Scholar] [CrossRef]
- Ouyang, Z.; Tan, T.; Liu, C.; Duan, J.; Wang, W.; Guo, X.; Zhang, Q.; Li, Z.; Huang, Q.; Dou, P.; et al. Targeted delivery of hesperetin to cartilage attenuates osteoarthritis by bimodal imaging with Gd2(CO3)3@PDA nanoparticles via TLR-2/NF-kappaB/Akt signaling. Biomaterials 2019, 205, 50–63. [Google Scholar] [CrossRef]
- Fu, Z.; Chen, Z.; Xie, Q.; Lei, H.; Xiang, S. Hesperidin protects against IL-1beta-induced inflammation in human osteoarthritis chondrocytes. Exp. Ther. Med. 2018, 16, 3721–3727. [Google Scholar]
- Zu, Y.; Mu, Y.; Li, Q.; Zhang, S.T.; Yan, H.J. Icariin alleviates osteoarthritis by inhibiting NLRP3-mediated pyroptosis. J. Orthop. Surg. Res. 2019, 14, 307. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Y.; Li, H.; Jia, L.; Wang, J.; Liang, S.; Cai, A.; Tan, X.; Wang, L.; Wang, X.; et al. Verification of pain-related neuromodulation mechanisms of icariin in knee osteoarthritis. Biomed. Pharmacother. 2021, 144, 112259. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, L.; Wang, S.; Wang, L.; Wang, X.; Xu, H.; Li, X.; Ye, H. Confirmation of inhibiting TLR4/Myd88/NFkB signalling pathway by Duhuo Jisheng decoction on osteoarthritis: A network pharmacology approach-integrated experimental study. Front. Pharmacol. 2022, 12, 784822. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Rong, X.F.; Li, R.H.; Wu, X.Y. Icariin inhibits MMP1, MMP3 and MMP13 expression through MAPK pathways in IL1betastimulated SW1353 chondrosarcoma cells. Mol. Med. Rep. 2017, 15, 2853–2858. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, T.; Cao, B.R.; Luan, F.Y.; Liu, R.X.; Yin, H.R.; Wang, W.B. Icariin possesses chondroprotective efficacy in a rat model of dexamethasone-induced cartilage injury through the activation of miR-206 targeting of cathepsin K. Int. J. Mol. Med. 2018, 41, 1039–1047. [Google Scholar] [CrossRef]
- Luo, H.; Zhang, R. Icariin enhances cell survival in lipopolysaccharide-induced synoviocytes by suppressing ferroptosis via the Xc-/GPX4 axis. Exp. Ther. Med. 2021, 21, 72. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, C.; Zhao, S.; Xu, H.; Peng, Z.; Zhang, B.; Cai, W.; Mo, Y.; Zhao, W. Evaluation of the effectiveness and safety of icariin in the treatment of knee osteoarthritis: A protocol for a systematic review and meta-analysis. Medicine 2021, 17, e28277. [Google Scholar] [CrossRef]
- Li, J.; Luo, M.; Wang, S.; Jin, G.; Han, Z.; Ma, Y.; Tang, G.; Guo, Z. Icariin Ameliorates Lower Back Pain in Rats via Suppressing the Secretion of Cytokine-Induced Neutrophil Chemoatractant-1. Biomed Res Int. 2020, 2020, 4670604. [Google Scholar] [CrossRef]
- Schwager, J.; Richard, N.; Fowler, A.; Seifert, N.; Raederstorff, D. Carnosol and Related Substances Modulate Chemokine and Cytokine Production in Macrophages and Chondrocytes. Molecules 2016, 21, 465. [Google Scholar] [CrossRef]
- Sun, W.; Xie, W.; Huang, D.; Cui, Y.; Yue, J.; He, Q.; Jiang, L.; Xiong, J.; Sun, W.; Yi, Q. Caffeic acid phenethyl ester attenuates osteoarthritis progression by activating NRF2/HO1 and inhibiting the NFkappaB signaling pathway. Int. J. Mol. Med. 2022, 50, 1–14. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, J.; Li, S.; Wu, Y.; Yu, C.; Ni, L.; Xiao, J.; Shao, Z.; Zhu, H.; Wang, J.; et al. Tangeretin suppresses osteoarthritis progression via the Nrf2/NF-kappaB and MAPK/NF-kappaB signaling pathways. Phytomedicine 2022, 98, 153928. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wu, D.; Huang, L.; Jiang, C.; Pan, T.; Kang, X.; Pan, J. Nobiletin Inhibits IL-1beta-Induced Inflammation in Chondrocytes via Suppression of NF-kappaB Signaling and Attenuates Osteoarthritis in Mice. Front. Pharmacol. 2019, 10, 570. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Guo, L.; Tian, F.D.; An, N.; Luo, L.; Hao, R.H.; Wang, B.; Zhou, Z.H. Naringenin regulates production of matrix metalloproteinases in the knee-joint and primary cultured articular chondrocytes and alleviates pain in rat osteoarthritis model. Braz. J. Med. Biol. Res. 2017, 50, e5714. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Jin, Z.; Ren, W.; Lu, M.; Hou, M.; Zhou, Q.; Wang, W.; Yang, H.; Zou, J. Theaflavin-3,3’-Digallate Protects Cartilage from Degradation by Modulating Inflammation and Antioxidant Pathways. Oxid. Med. Cell. Longev. 2022, 2022, 3047425. [Google Scholar] [CrossRef] [PubMed]
- Chuntakaruk, H.; Kongtawelert, P.; Pothacaroen, P. Chondroprotective effects of purple corn anthocyanins on advanced glycation end products induction through suppression of NFkB and MAPK signalling. Sci. Rep. 2021, 11, 1895. [Google Scholar] [CrossRef]
- Jiang, C.; Sun, Z.M.; Hu, J.N.; Jin, Y.; Guo, Q.; Xu, J.J.; Chen, Z.X.; Jiang, R.H.; Wu, Y.S. Cyanidin ameliorates the progression of osteoarthritis via the Sirt6/NF-kappaB axis in vitro and in vivo. Food Funct. 2019, 10, 5873–5885. [Google Scholar] [CrossRef]
- Lin, Z.; Lin, C.; Fu, C.; Lu, H.; Jin, H.; Chen, Q.; Pan, J. The protective effect of Ellagic acid (EA) in osteoarthritis: An in vitro and in vivo study. Biomed. Pharmacother. 2020, 125, 109845. [Google Scholar] [CrossRef]
- Du, K.; Fang, X.; Li, Z. Ferulic acid suppresses interleukin-1beta-induced degeneration of chondrocytes isolated from patients with osteoarthritis through the SIRT1/AMPK/PGC-1alpha signaling pathway. Immun. Inflamm. Dis. 2021, 9, 710–720. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, Z.; You, S.; Zhang, H.; Tao, Z.; Wang, Q.; Chen, H.; Wu, Y. Fisetin inhibits IL-1beta-induced inflammatory response in human osteoarthritis chondrocytes through activating SIRT1 and attenuates the progression of osteoarthritis in mice. Int. Immunopharmacol. 2017, 45, 135–147. [Google Scholar] [CrossRef]
- Wang, K.; Chen, X.; Chen, Y.; Sheng, S.; Huang, Z. Grape seed procyanidins suppress the apoptosis and senescence of chondrocytes and ameliorates osteoarthritis via the DPP4-Sirt1 pathway. Food Funct. 2020, 11, 10493–10505. [Google Scholar] [CrossRef]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J. Cell. Mol. Med. 2020, 24, 10855–10865. [Google Scholar] [CrossRef] [PubMed]
- Ziadlou, R.; Barbero, A.; Martin, I.; Wang, X.; Qin, L.; Alini, M.; Grad, S. Anti-Inflammatory and Chondroprotective Effects of Vanillic Acid and Epimedin C in Human Osteoarthritic Chondrocytes. Biomolecules 2020, 10, 932. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Tang, J.L.; Bao, J.P.; Hu, P.F.; Shi, Z.L.; Wu, L.D. Anti-arthritic effects of chlorogenic acid in interleukin-1beta-induced rabbit chondrocytes and a rabbit osteoarthritis model. Int. Immunopharmacol. 2011, 11, 23–28. [Google Scholar] [CrossRef]
- Wu, D.Q.; Zhong, H.M.; Ding, Q.H.; Ba, L. Protective effects of biochanin A on articular cartilage: In vitro and in vivo studies. BMC Complement. Altern. Med. 2014, 14, 444. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Zhang, Y.; Fang, J.; Jin, Y. Network Pharmacology-Based Prediction and Verification of the Potential Targets of Pinoresinol Diglucoside for OA Treatment. Evid.-Based Complementary Altern. Med. 2022, 2022, 9733742. [Google Scholar] [CrossRef] [PubMed]
- Srisuthtayanont, W.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Effects of sesamin on chondroitin sulfate proteoglycan synthesis induced by interleukin-1beta in human chondrocytes. BMC Complement. Altern. Med. 2017, 17, 286. [Google Scholar] [CrossRef]
- Li, X.; Lin, J.; Ding, X.; Xuan, J.; Hu, Z.; Wu, D.; Zhu, X.; Feng, Z.; Ni, W.; Wu, A. The protective effect of sinapic acid in osteoarthritis: In vitro and in vivo studies. J. Cell. Mol. Med. 2019, 23, 1940–1950. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Huff, T.W.; Liu, J.; Yuan, T.; Wei, Z.; Qin, J. Alleviation of Cartilage Destruction by Sinapic Acid in Experimental Osteoarthritis. Biomed. Res. Int. 2019, 2019, 5689613. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Hong, H.; Wang, N.; Chen, J.; Lu, S.; Zhang, H.; Zhang, X.; Bei, C. Xanthohumol suppresses inflammation in chondrocytes and ameliorates osteoarthritis in mice. Biomed. Pharmacother. 2021, 137, 111238. [Google Scholar] [CrossRef]
- Pan, X.; Chen, T.; Zhang, Z.; Chen, X.; Chen, C.; Chen, L.; Wang, X.; Ying, X. Activation of Nrf2/HO-1 signal with Myricetin for attenuating ECM degradation in human chondrocytes and ameliorating the murine osteoarthritis. Int. Immunopharmacol. 2019, 75, 105742. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, L.; Liu, Y.; Huang, C.; Xia, W.; Zhou, H.; Zhou, Z.; Zhou, X. Luteolin Protects Chondrocytes from H2O2-Induced Oxidative Injury and Attenuates Osteoarthritis Progression by Activating AMPK-Nrf2 Signaling. Oxid. Med. Cell. Longev. 2022, 2022, 5635797. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Park, Y.J.; Song, M.G.; Kim, D.R.; Zada, S.; Kim, D.H. Cytoprotective Effects of Delphinidin for Human Chondrocytes against Oxidative Stress through Activation of Autophagy. Antioxidants 2020, 9, 83. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, R.; Zheng, T.; Chen, Z.; Ji, G.; Peng, F.; Wang, W. Xanthohumol Attenuated Inflammation and ECM Degradation by Mediating HO-1/C/EBPbeta Pathway in Osteoarthritis Chondrocytes. Front. Pharmacol. 2021, 12, 680585. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xi, Y.; Mao, Z.; Chu, X.; Zhang, R.; Ma, X.; Ni, B.; Cheng, H.; You, H. Vanillic acid attenuates cartilage degeneration by regulating the MAPK and PI3K/AKT/NF-kappaB pathways. Eur. J. Pharmacol. 2019, 859, 172481. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Jin, G.J.; Xiong, Y.; Hu, P.F.; Bao, J.P.; Wu, L.D. Rosmarinic acid down-regulates NO and PGE2 expression via MAPK pathway in rat chondrocytes. J. Cell. Mol. Med. 2018, 22, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Kang, S.H.; Jeong, S.J.; Kim, S.H.; Ko, H.S.; Kim, S.H. Inhibition of c-Jun N-terminal kinase and nuclear factor kappa B pathways mediates fisetin-exerted anti-inflammatory activity in lipopolysccharide-treated RAW264.7 cells. Immunopharmacol. Immunotoxicol. 2012, 34, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Gomis-Ruth, F.X.; Maskos, K.; Betz, M.; Betz, M.; Bergner, A.; Huber, R.; Suzuki, K.; Yoshida, N.; Nagase, H.; Brew, K.; et al. Mechanism of inhibition of the human matrix metalloproteinase stromelysin-1 by TIMP-1. Nature 1997, 389, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yang, H.; Li, D.; Wu, X.; Han, Q. Punicalagin attenuates osteoarthritis progression via regulating Foxo1/Prg4/HIF3alpha axis. Bone 2021, 152, 116070. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Zhang, Y.; Tang, L.; Ji, Y.; Yan, C.; Zhang, X. Protective effects of quercetin against inflammation and oxidative stress in a rabbit model of knee osteoarthritis. Drug Dev. Res. 2019, 80, 360–367. [Google Scholar] [CrossRef]
- Mok, S.W.; Fu, S.C.; Cheuk, Y.C.; Chu, I.M.; Chan, K.M.; Qin, L.; Yung, S.H.; Kevin Ho, K.W. Intra-Articular Delivery of Quercetin Using Thermosensitive Hydrogel Attenuate Cartilage Degradation in an Osteoarthritis Rat Model. Cartilage 2020, 11, 490–499. [Google Scholar] [CrossRef]
- Britti, D.; Crupi, R.; Impellizzeri, D.; Gugliandolo, E.; Fusco, R.; Schievano, C.; Morittu, V.M.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. A novel composite formulation of palmitoylethanolamide and quercetin decreases inflammation and relieves pain in inflammatory and osteoarthritic pain models. BMC Vet. Res. 2017, 13, 229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Leong, D.J.; Xu, L.; He, Z.; Wang, A.; Navati, M.; Kim, S.J.; Hirsh, D.M.; Hardin, J.A.; Cobelli, N.J.; et al. Curcumin slows osteoarthritis progression and relieves osteoarthritis-associated pain symptoms in a post-traumatic osteoarthritis mouse model. Arthritis Res. Ther. 2016, 18, 128. [Google Scholar] [CrossRef] [PubMed]
- Yabas, M.; Orhan, C.; Er, B.; Tuzcu, M.; Durmus, A.S.; Ozercan, I.H.; Sahin, N.; Bhanuse, P.; Morde, A.A.; Padigaru, M.; et al. A Next Generation Formulation of Curcumin Ameliorates Experimentally Induced Osteoarthritis in Rats via Regulation of Inflammatory Mediators. Front. Immunol. 2021, 12, 609629. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Shi, K.; Chen, G.; Shen, Y.; Pan, T. Malvidin attenuates pain and inflammation in rats with osteoarthritis by suppressing NF-kappaB signaling pathway. Inflamm. Res. 2017, 66, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Sheu, S.Y.; Chen, W.S.; Sun, J.S.; Lin, F.H.; Wu, T. Biological characterization of oxidized hyaluronic acid/resveratrol hydrogel for cartilage tissue engineering. J. Biomed. Mater. Res. A 2013, 101, 3457–3466. [Google Scholar] [CrossRef]
- Shakibaei, M.; John, T.; Seifarth, C.; Mobasheri, A. Resveratrol inhibits IL-1 beta-induced stimulation of caspase-3 and cleavage of PARP in human articular chondrocytes in vitro. Ann. N. Y. Acad. Sci. 2007, 1095, 554–563. [Google Scholar] [CrossRef]
- Wendling, D.; Abbas, W.; Godfrin-Valnet, M.; Guillot, X.; Khan, K.A.; Cedoz, J.P.; Baud, L.; Prati, C.; Herbein, G. Resveratrol, a sirtuin 1 activator, increases IL-6 production by peripheral blood mononuclear cells of patients with knee osteoarthritis. Clin. Epigenetics 2013, 5, 10. [Google Scholar] [CrossRef]
- Xiong, F.; Qin, Z.; Chen, H.; Lan, Q.; Wang, Z.; Lan, N.; Yang, Y.; Zheng, L.; Zhao, J.; Kai, D. pH-responsive and hyaluronic acid-functionalized metal-organic frameworks for therapy of osteoarthritis. J. Nanobiotechnol. 2020, 18, 139. [Google Scholar] [CrossRef]
- Wen, L.; Qu, T.B.; Zhai, K.; Ding, J.; Hai, Y.; Zhou, J.L. Gallic acid can play a chondroprotective role against AGE-induced osteoarthritis progression. J. Orthop. Sci. 2015, 20, 734–741. [Google Scholar] [CrossRef]
- Huang, X.; You, Y.; Xi, Y.; Ni, B.; Chu, X.; Zhang, R.; You, H. p-Coumaric Acid Attenuates IL-1beta-Induced Inflammatory Responses and Cellular Senescence in Rat Chondrocytes. Inflammation 2020, 43, 619–628. [Google Scholar] [CrossRef]
- Chen, M.P.; Yang, S.H.; Chou, C.H.; Yang, K.C.; Wu, C.C.; Cheng, Y.H.; Lin, F.H. The chondroprotective effects of ferulic acid on hydrogen peroxide-stimulated chondrocytes: Inhibition of hydrogen peroxide-induced pro-inflammatory cytokines and metalloproteinase gene expression at the mRNA level. Inflamm. Res. 2010, 59, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Pan, Q.; Mao, Z.; Zhang, R.; Ma, X.; Xi, Y.; You, H. Sinapic Acid Inhibits the IL-1beta-Induced Inflammation via MAPK Downregulation in Rat Chondrocytes. Inflammation 2018, 41, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xi, Y.; Pan, Q.; Mao, Z.; Zhang, R.; Ma, X.; You, H. Caffeic acid protects against IL-1beta-induced inflammatory responses and cartilage degradation in articular chondrocytes. Biomed. Pharmacother. 2018, 107, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Zhang, Y.; Dai, B.L.; Ma, Y.J.; Zhang, Q.; Wang, Y.; Yang, H. Chlorogenic acid prevents inflammatory responses in IL1betastimulated human SW1353 chondrocytes, a model for osteoarthritis. Mol. Med. Rep. 2017, 16, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Wu, L.D. Chlorogenic acid suppresses interleukin-1beta-induced inflammatory mediators in human chondrocytes. Int. J. Clin. Exp. Pathol. 2014, 7, 8797–8801. [Google Scholar]
- Huh, J.E.; Seo, D.M.; Baek, Y.H.; Choi, D.Y.; Park, D.S.; Lee, J.D. Biphasic positive effect of formononetin on metabolic activity of human normal and osteoarthritic subchondral osteoblasts. Int. Immunopharmacol. 2010, 10, 500–507. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, Q.; Guo, P.; Huang, Y.; Ye, Z.; Hu, J. Antichondrocyte apoptosis effect of genistein in treating inflammationinduced osteoarthritis. Mol. Med. Rep. 2020, 22, 2032–2042. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.S.; Cho, I.A.; Kang, K.R.; You, J.S.; Yu, S.J.; Lee, G.J.; Seo, Y.S.; Kim, C.S.; Kim, D.K.; Kim, S.G.; et al. Biochanin-A antagonizes the interleukin-1beta-induced catabolic inflammation through the modulation of NFkappaB cellular signaling in primary rat chondrocytes. Biochem. Biophys. Res. Commun. 2016, 477, 723–730. [Google Scholar] [CrossRef]
- Zheng, W.; Chen, C.; Zhang, C.; Cai, L.; Chen, H. The protective effect of phloretin in osteoarthritis: An in vitro and in vivo study. Food Funct. 2018, 9, 263–278. [Google Scholar] [CrossRef]
- Cho, C.; Kang, L.J.; Jang, D.; Jeon, J.; Lee, H.; Choi, S.; Han, S.J.; Oh, E.; Nam, J.; Kim, C.S.; et al. Cirsium japonicum var. maackii and apigenin block Hif-2alpha-induced osteoarthritic cartilage destruction. J. Cell. Mol. Med. 2019, 23, 5369–5379. [Google Scholar] [CrossRef]
- Park, J.S.; Kim, D.K.; Shin, H.D.; Lee, H.J.; Jo, H.S.; Jeong, J.H.; Choi, Y.L.; Lee, C.J.; Hwang, S.C. Apigenin Regulates Interleukin-1beta-Induced Production of Matrix Metalloproteinase Both in the Knee Joint of Rat and in Primary Cultured Articular Chondrocytes. Biomol. Ther. 2016, 24, 163–170. [Google Scholar] [CrossRef]
- Xue, J.; Ye, J.; Xia, Z.; Cheng, B. Effect of luteolin on apoptosis, MAPK and JNK signaling pathways in guinea pig chondrocyte with osteoarthritis. Cell. Mol. Biol. 2019, 65, 91–95. [Google Scholar] [CrossRef]
- Kang, B.J.; Ryu, J.; Lee, C.J.; Hwang, S.C. Luteolin Inhibits the Activity, Secretion and Gene Expression of MMP-3 in Cultured Articular Chondrocytes and Production of MMP-3 in the Rat Knee. Biomol. Ther. 2014, 22, 239–245. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, L.; Zheng, Y.; Liu, W.; Xu, Y. Kaempferol attenuates the effects of XIST/miR-130a/STAT3 on inflammation and extracellular matrix degradation in osteoarthritis. Future Med. Chem. 2021, 13, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Ye, G.; Huang, B. Kaempferol Alleviates the Interleukin-1beta-Induced Inflammation in Rat Osteoarthritis Chondrocytes via Suppression of NF-kappaB. Med. Sci. Monit. 2017, 23, 3925–3931. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.S.; Tseng, C.Y.; Lee, C.H.; Su, S.L.; Lee, H.S. Effects of (-)-epigallocatechin-3-gallate on cyclooxygenase 2, PGE(2), and IL-8 expression induced by IL-1beta in human synovial fibroblasts. Rheumatol. Int. 2010, 30, 1197–1203. [Google Scholar] [CrossRef]
- Li, J.; Wu, R.; Qin, X.; Liu, D.; Lin, F.; Feng, Q. Isorhamnetin inhibits IL1betainduced expression of inflammatory mediators in human chondrocytes. Mol. Med. Rep. 2017, 16, 4253–4258. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Qian, Y.; Chen, C.; Feng, F.; Pan, L.; Yang, L.; Wang, C. Hesperetin Exhibits Anti-Inflammatory Effects on Chondrocytes via the AMPK Pathway to Attenuate Anterior Cruciate Ligament Transection-Induced Osteoarthritis. Front. Pharmacol. 2021, 12, 735087. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Ding, H.; Zhuang, C.; Fan, W. Effects of Hesperidin on H2O2-Treated Chondrocytes and Cartilage in a Rat Osteoarthritis Model. Med. Sci. Monit. 2018, 24, 9177–9186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mi, B.; Lv, H.; Liu, J.; Xiong, Y.; Hu, L.; Xue, H.; Panayi, A.C.; Liu, G.; Zhou, W. Shared KEGG pathways of icariin-targeted genes and osteoarthritis. J. Cell Biochem. 2018, 120, 7741–7750. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, Y.; Chen, N.; Yang, L. Icariin Regulates Cellular Functions and Gene Expression of Osteoarthritis Patient-Derived Human Fibroblast-Like Synoviocytes. Int. J. Mol. Sci. 2017, 18, 2656. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, A.; Chen, D.; Haqqi, T.M. Delphinidin inhibits IL-1beta-induced activation of NF-kappaB by modulating the phosphorylation of IRAK-1(Ser376) in human articular chondrocytes. Rheumatology 2013, 52, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Song, W.H.; Lee, G.; Kim, H.S.; Park, D.; Huh, Y.H.; Ryu, J.H. Avenanthramide C as a novel candidate to alleviate osteoarthritic pathogenesis. BMB Rep. 2021, 54, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Csaki, C.; Nebrich, S.; Mobasheri, A. Resveratrol suppresses interleukin-1beta-induced inflammatory signaling and apoptosis in human articular chondrocytes: Potential for use as a novel nutraceutical for the treatment of osteoarthritis. Biochem. Pharmacol. 2008, 76, 1426–1439. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Mobasheri, A.; Buhrmann, C. Curcumin synergizes with resveratrol to stimulate the MAPK signaling pathway in human articular chondrocytes in vitro. Genes Nutr. 2011, 6, 171–179. [Google Scholar] [CrossRef]
- Csaki, C.; Mobasheri, A.; Shakibaei, M. Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: Inhibition of IL-1beta-induced NF-kappaB-mediated inflammation and apoptosis. Arthritis Res. Ther. 2009, 11, R165. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, D.; Tang, J. Identification of the Resveratrol Potential Targets in the Treatment of Osteoarthritis. Evid.-Based Complementary Altern. Med. 2021, 2021, 9911286. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Zhang, W.; Cui, Z.M.; Cui, S.Y.; Fan, J.B.; Zhu, X.H.; Liu, W. Resveratrol alleviates the interleukin-1beta-induced chondrocytes injury through the NF-kappaB signaling pathway. J. Orthop. Surg. Res. 2020, 15, 424. [Google Scholar] [CrossRef]
- Ma, P.; Yue, L.; Yang, H.; Fan, Y.; Bai, J.; Li, S.; Yuan, J.; Zhang, Z.; Yao, C.; Lin, M.; et al. Chondroprotective and anti-inflammatory effects of amurensin H by regulating TLR4/Syk/NF-kappaB signals. J. Cell. Mol. Med. 2020, 24, 1958–1968. [Google Scholar] [CrossRef]
- Dave, M.; Attur, M.; Palmer, G.; Al-Mussawir, H.E.; Kennish, L.; Patel, J.; Abramson, S.B. The antioxidant resveratrol protects against chondrocyte apoptosis via effects on mitochondrial polarization and ATP production. Arthritis Rheum. 2008, 58, 2786–2797. [Google Scholar] [CrossRef]
- Wongwichai, T.; Teeyakasem, P.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Anthocyanins and metabolites from purple rice inhibit IL-1beta-induced matrix metalloproteinases expression in human articular chondrocytes through the NF-kappaB and ERK/MAPK pathway. Biomed. Pharmacother. 2019, 112, 108610. [Google Scholar] [CrossRef] [PubMed]
- Garbacki, N.; Angenot, L.; Bassleer, C.; Damas, J.; Tits, M. Effects of prodelphinidins isolated from Ribes nigrum on chondrocyte metabolism and COX activity. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2002, 365, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Anbazhagan, A.N.; Akhtar, N.; Ramamurthy, S.; Voss, F.R.; Haqqi, T.M. Green tea polyphenol epigallocatechin-3-gallate inhibits advanced glycation end product-induced expression of tumor necrosis factor-alpha and matrix metalloproteinase-13 in human chondrocytes. Arthritis Res. Ther. 2009, 11, R71. [Google Scholar] [CrossRef]
- Singh, R.; Ahmed, S.; Islam, N.; Goldberg, V.M.; Haqqi, T.M. Epigallocatechin-3-gallate inhibits interleukin-1beta-induced expression of nitric oxide synthase and production of nitric oxide in human chondrocytes: Suppression of nuclear factor kappaB activation by degradation of the inhibitor of nuclear factor kappaB. Arthritis Rheum. 2002, 46, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Rahman, A.; Hasnain, A.; Lalonde, M.; Goldberg, V.M.; Haqqi, T.M. Green tea polyphenol epigallocatechin-3-gallate inhibits the IL-1 beta-induced activity and expression of cyclooxygenase-2 and nitric oxide synthase-2 in human chondrocytes. Free Radic. Biol. Med. 2002, 33, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Ishitobi, H.; Sanada, Y.; Kato, Y.; Ikuta, Y.; Shibata, S.; Yamasaki, S.; Lotz, M.K.; Matsubara, K.; Miyaki, S.; Adachi, N. Carnosic acid attenuates cartilage degeneration through induction of heme oxygenase-1 in human articular chondrocytes. Eur. J. Pharmacol. 2018, 830, 1–8. [Google Scholar] [CrossRef]
- Phitak, T.; Pothacharoen, P.; Settakorn, J.; Poompimol, W.; Caterson, B.; Kongtawelert, P. Chondroprotective and anti-inflammatory effects of sesamin. Phytochemistry 2012, 80, 77–88. [Google Scholar] [CrossRef]
- Estakhri, F.; Reza Panjehshahin, M.; Tanideh, N.; Gheisari, R.; Azarpira, N.; Gholijani, N. Efficacy of Combination Therapy with Apigenin and Synovial Membrane-Derived Mesenchymal Stem Cells on Knee Joint Osteoarthritis in a Rat Model. Iran, J. Med. Sci. 2021, 46, 383–394. [Google Scholar]
- Estakhri, F.; Panjehshahin, M.R.; Tanideh, N.; Gheisari, R.; Mahmoodzadeh, A.; Azarpira, N.; Gholijani, N. The effect of kaempferol and apigenin on allogenic synovial membrane-derived stem cells therapy in knee osteoarthritic male rats. Knee 2020, 27, 817–832. [Google Scholar] [CrossRef]
- Kalff, K.M.; El Mouedden, M.; van Egmond, J.; Veening, J.; Joosten, L.; Scheffer, G.J.; Meert, T.; Vissers, K. Pre-treatment with capsaicin in a rat osteoarthritis model reduces the symptoms of pain and bone damage induced by monosodium iodoacetate. Eur. J. Pharmacol. 2010, 641, 108–113. [Google Scholar] [CrossRef]
- Huang, H.T.; Cheng, T.L.; Yang, C.D.; Chang, C.F.; Ho, C.J.; Chuang, S.C.; Li, J.Y.; Huang, S.H.; Lin, Y.S.; Shen, H.Y.; et al. Intra-Articular Injection of (-)-Epigallocatechin 3-Gallate (EGCG) Ameliorates Cartilage Degeneration in Guinea Pigs with Spontaneous Osteoarthritis. Antioxidants 2021, 10, 178. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.T.; Cheng, T.L.; Ho, C.J.; Huang, H.H.; Lu, C.C.; Chuang, S.C.; Li, J.Y.; Lee, T.C.; Chen, S.T.; Lin, Y.S.; et al. Intra-Articular Injection of (-)-Epigallocatechin 3-Gallate to Attenuate Articular Cartilage Degeneration by Enhancing Autophagy in a Post-Traumatic Osteoarthritis Rat Model. Antioxidants 2020, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xu, P. The articular cartilage preservative effects of genistein in an experimental model of knees osteoarthritis. Appl. Physiol. Nutr. Metab. 2021, 46, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fu, B.; Chen, X.; Chen, D.; Yang, H. Protocatechuic acid attenuates anterior cruciate ligament transection-induced osteoarthritis by suppressing osteoclastogenesis. Exp. Ther. Med. 2020, 19, 232–240. [Google Scholar] [CrossRef]
- Xiong, W.; Lan, Q.; Liang, X.; Zhao, J.; Huang, H.; Zhan, Y.; Qin, Z.; Jiang, X.; Zheng, L. Cartilage-targeting poly(ethylene glycol) (PEG)-formononetin (FMN) nanodrug for the treatment of osteoarthritis. J. Nanobiotechnol. 2021, 19, 197. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, J.; Zhao, D.; Wang, C.; Zhang, Y.; Wang, Y.; Li, T. Therapeutic effect and mechanism of action of quercetin in a rat model of osteoarthritis. J. Int. Med. Res. 2020, 48, 300060519873461. [Google Scholar] [CrossRef]
- Heydari Nasrabadi, M.; Parsivand, M.; Mohammadi, N.; Asghari Moghaddam, N. Comparison of Elaeagnus angustifolia L. extract and quercetin on mouse model of knee osteoarthritis. J. Ayurveda Integr. Med. 2022, 13, 100529. [Google Scholar] [CrossRef]
- Qiu, L.; Luo, Y.; Chen, X. Quercetin attenuates mitochondrial dysfunction and biogenesis via upregulated AMPK/SIRT1 signaling pathway in OA rats. Biomed. Pharmacother. 2018, 103, 1585–1591. [Google Scholar] [CrossRef]
- Permatasari, D.A.; Karliana, D.; Iskandarsyah, I.; Arsianti, A.; Bahtiar, A. Quercetin prevent proteoglycan destruction by inhibits matrix metalloproteinase-9, matrix metalloproteinase-13, a disintegrin and metalloproteinase with thrombospondin motifs-5 expressions on osteoarthritis model rats. J. Adv. Pharm. Technol. Res. 2019, 10, 2–8. [Google Scholar]
- Zhou, Z.; Deng, Z.; Liu, Y.; Zheng, Y.; Yang, S.; Lu, W.; Xiao, D.; Zhu, W. Protective Effect of SIRT1 Activator on the Knee with Osteoarthritis. Front. Physiol. 2021, 12, 661852. [Google Scholar] [CrossRef]
- Wang, Z.M.; Chen, Y.C.; Wang, D.P. Resveratrol, a natural antioxidant, protects monosodium iodoacetate-induced osteoarthritic pain in rats. Biomed. Pharmacother. 2016, 83, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.J.; Liu, C.C.; Chen, L.M.; Wu, G.H.; Wang, X.P. Reduction of SIRT1 Mediates Monosodium Iodoacetate-Induced Osteoarthritic Pain by Upregulating p53 Expression in Rats. Pain Physician 2021, 24, E1025–E1035. [Google Scholar] [PubMed]
- Long, Z.; Xiang, W.; Li, J.; Yang, T.; Yu, G. Exploring the Mechanism of Resveratrol in Reducing the Soft Tissue Damage of Osteoarthritis Based on Network Pharmacology and Experimental Pharmacology. Evid.-Based Complementary Altern. Med. 2021, 2021, 9931957. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, X.; Yu, X.; Liu, X.; Xu, X.; He, J.; Gu, H.; Liu, L. Oral Administration of Resveratrol Alleviates Osteoarthritis Pathology in C57BL/6J Mice Model Induced by a High-Fat Diet. Mediat. Inflamm. 2017, 2017, 7659023. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Xu, Z.; Zhao, X.; Yuan, M.; Pan, L.; Lu, T.; Du, A.; Qin, L. In Vivo Effect of Resveratrol-Cellulose Aerogel Drug Delivery System to Relieve Inflammation on Sports Osteoarthritis. Gels 2022, 8, 544. [Google Scholar] [CrossRef]
- Ma, Z.; Huang, Z.; Zhang, L.; Li, X.; Xu, B.; Xiao, Y.; Shi, X.; Zhang, H.; Liao, T.; Wang, P. Vanillic Acid Reduces Pain-Related Behavior in Knee Osteoarthritis Rats Through the Inhibition of NLRP3 Inflammasome-Related Synovitis. Front. Pharmacol. 2020, 11, 599022. [Google Scholar] [CrossRef]
- Shep, D.; Khanwelkar, C.; Gade, P.; Karad, S. Efficacy and safety of combination of curcuminoid complex and diclofenac versus diclofenac in knee osteoarthritis: A randomized trial. Medicine 2020, 99, e19723. [Google Scholar] [CrossRef]
- Belcaro, G.; Cesarone, M.R.; Dugall, M.; Pellegrini, L.; Ledda, A.; Grossi, M.G.; Togni, S.; Appendino, G. Efficacy and safety of Meriva(R), a curcumin-phosphatidylcholine complex, during extended administration in osteoarthritis patients. Altern. Med. Rev. 2010, 15, 337–344. [Google Scholar]
- Marouf, B.H. Effect of Resveratrol on Serum Levels of Type II Collagen and Aggrecan in Patients with Knee Osteoarthritis: A Pilot Clinical Study. Biomed. Res. Int. 2021, 2021, 3668568. [Google Scholar] [CrossRef]
- Marouf, B.H.; Hussain, S.A.; Ali, Z.S.; Ahmmad, R.S. Resveratrol Supplementation Reduces Pain and Inflammation in Knee Osteoarthritis Patients Treated with Meloxicam: A Randomized Placebo-Controlled Study. J. Med. Food 2018, 21, 1253–1259. [Google Scholar] [CrossRef]
- Matsuno, H.; Nakamura, H.; Katayama, K.; Hayashi, S.; Kano, S.; Yudoh, K.; Kiso, Y. Effects of an oral administration of glucosamine-chondroitin-quercetin glucoside on the synovial fluid properties in patients with osteoarthritis and rheumatoid arthritis. Biosci. Biotechnol. Biochem. 2009, 73, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, N.; Saito, K.; Maeda, A.; Kitagawa, Y.; Kiso, Y.; Watanabe, K.; Tomonaga, A.; Nagaoka, I.; Yamaguchi, H. Effect of a dietary supplement containing glucosamine hydrochloride, chondroitin sulfate and quercetin glycosides on symptomatic knee osteoarthritis: A randomized, double-blind, placebo-controlled study. J. Sci. Food Agric. 2012, 92, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Persson, M.S.M.; Stocks, J.; Sarmanova, A.; Fernandes, G.; Walsh, D.A.; Doherty, M.; Zhang, W. Individual responses to topical ibuprofen gel or capsaicin cream for painful knee osteoarthritis: A series of n-of-1 trials. Rheumatology 2021, 60, 2231–2237. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.M.; Ervin, J.; Nezzer, J.; Nieves, Y.; Guedes, K.; Burges, R.; Hanson, P.D.; Campbell, J.N. Randomized, Double-Blind, Placebo-Controlled Trial of Intraarticular Trans-Capsaicin for Pain Associated With Osteoarthritis of the Knee. Arthritis Rheumatol. 2019, 71, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Farid, R.; Mirfeizi, Z.; Mirheidari, M.; Hatef, M.R.; Mirheidari, M.; Mansouri, H.; Esmaelli, H.; Bentley, G.; Lu, Y.; Foo, Y.; et al. Pycnogenol supplementation reduces pain and stiffness and improves physical function in adults with knee osteoarthritis. Nutr. Res. 2007, 27, 692–697. [Google Scholar] [CrossRef]
- Belcaro, G.; Cesarone, M.R.; Errichi, S.; Zulli, C.; Errichi, B.M.; Vinciguerra, G.; Ledda, A.; Di Renzo, A.; Stuard, S.; Dugall, M.; et al. Variations in C-reactive protein, plasma free radicals and fibrinogen values in patients with osteoarthritis treated with Pycnogenol. Redox Rep. 2008, 13, 271–276. [Google Scholar] [CrossRef]
- Ofosu, F.K.; Daliri, E.B.M.; Elahi, F.; Chelliah, R.; Lee, B.H.; Oh, D.H. New Insights on the Use of Polyphenols as Natural Preservatives and Their Emerging Safety Concerns. Front. Sustain. Food Syst. 2020, 4, 525810. [Google Scholar] [CrossRef]
- Ochi, M.; Uchio, Y.; Tobita, M.; Kuriwaka, M. Current concepts in tissue engineering technique for repair of cartilage defect. Artif. Organs 2001, 25, 172–179. [Google Scholar] [CrossRef]
- Rompicherla, N.C.; Joshi, P.; Shetty, A.; Sudhakar, K.; Amin, H.I.M.; Mishra, Y.; Mishra, V.; Albutti, A.; Alhumeed, N. Design, Formulation, and Evaluation of Aloe vera Gel-Based Capsaicin Transemulgel for Osteoarthritis. Pharmaceutics 2022, 14, 1812. [Google Scholar] [CrossRef]
- Gutierres, V.O.; Campos, M.L.; Arcaro, C.A.; Assis, R.P.; Baldan-Cimatti, H.M.; Peccinini, R.G.; Paula-Gomes, S.; Kettelhut, I.C.; Baviera, A.M.; Brunetti, I.L. Curcumin Pharmacokinetic and Pharmacodynamic Evidences in Streptozotocin-Diabetic Rats Support the Antidiabetic Activity to Be via Metabolite(s). Evid.-Based Complementary Altern. Med. 2015, 2015, 678218. [Google Scholar] [CrossRef]
- Azzolini, M.; La Spina, M.; Mattarei, A.; Paradisi, C.; Zoratti, M.; Biasutto, L. Pharmacokinetics and tissue distribution of pterostilbene in the rat. Mol. Nutr. Food Res. 2014, 58, 2122–2132. [Google Scholar] [CrossRef] [PubMed]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z.; Thompson, T.N.; McCormick, D.L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother. Pharmacol. 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Horio, F.; Osawa, T. Absorption and metabolism of cyanidin 3-O-beta-D-glucoside in rats. FEBS Lett. 1999, 449, 179–182. [Google Scholar] [CrossRef] [PubMed]
- de Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef]
- Smoliga, J.M.; Blanchard, O. Enhancing the delivery of resveratrol in humans: If low bioavailability is the problem, what is the solution? Molecules 2014, 19, 17154–17172. [Google Scholar] [CrossRef]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef]
- Eseberri, I.; Trepiana, J.; Leniz, A.; Gómez-García, I.; Carr-Ugarte, H.; González, M.; Portillo, M.P. Variability in the Beneficial Effects of Phenolic Compounds: A Review. Nutrients 2022, 14, 1925. [Google Scholar] [CrossRef]
- Kellgren, J.H.; Lawrence, J.S. Radiological assessment of osteoarthrosis. Ann. Rheum. Dis. 1957, 16, 494–502. [Google Scholar] [CrossRef]
- Helal, N.A.; Eassa, H.A.; Amer, A.M.; Eltokhy, M.A.; Edafiogho, I.; Nounou, M.I. Nutraceuticals’ Novel Formulations: The Good, the Bad, the Unknown and Patents Involved. Recent Pat. Drug Deliv. Formul. 2019, 13, 105–156. [Google Scholar] [CrossRef]
- Santini, A.; Novellino, E. Nutraceuticals—Shedding light on the grey area between pharmaceuticals and food. Expert Rev. Clin. Pharmacol. 2018, 11, 545–547. [Google Scholar] [CrossRef]
- Sirse, M. Effect of Dietary Polyphenols on Osteoarthritis-Molecular Mechanisms. Life 2022, 12, 436. [Google Scholar] [CrossRef]
- Puttasiddaiah, R.; Lakshminarayana, R.; Somashekar, N.L.; Gupta, V.K.; Inbaraj, B.S.; Usmani, Z.; Raghavendra, V.; Sridhar, K.; Sharma, M. Advances in Nanofabrication Technology for Nutraceuticals: New Insights and Future Trends. Bioengineering 2022, 9, 478. [Google Scholar] [CrossRef]
- Jayusman, P.A.; Nasruddin, N.S.; Mahamad Apandi, N.I.; Ibrahim, N.; Budin, S.B. Therapeutic Potential of Polyphenol and Nanoparticles Mediated Delivery in Periodontal Inflammation: A Review of Current Trends and Future Perspectives. Front. Pharmacol. 2022, 13, 847702. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.; Krithica, N.; Madhan, B.; Sehgal, P.K. Formulation and evaluation of quercetin polycaprolactone microspheres for the treatment of rheumatoid arthritis. J. Pharm. Sci. 2011, 100, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Ronholm, J.; Tian, Y.; Sethi, B.; Cao, X. Sterilization techniques for biodegradable scaffolds in tissue engineering applications. J. Tissue Eng. 2016, 7, 2041731416648810. [Google Scholar] [CrossRef] [PubMed]
- Maudens, P.; Jordan, O.; Allemann, E. Recent advances in intra-articular drug delivery systems for osteoarthritis therapy. Drug Discov. Today 2018, 23, 1761–1775. [Google Scholar] [CrossRef]
- Le Clanche, S.; Cheminel, T.; Rannou, F.; Bonnefont-Rousselot, D.; Borderie, D.; Charrueau, C. Use of Resveratrol Self-Emulsifying Systems in T/C28a2 Cell Line as Beneficial Effectors in Cellular Uptake and Protection Against Oxidative Stress-Mediated Death. Front. Pharmacol. 2018, 9, 538. [Google Scholar] [CrossRef]
- Cavalcanti, G.R.; Duarte, F.I.C.; Converti, A.; de Lima, A.A.N. Ferulic Acid Activity in Topical Formulations: Technological and Scientific Prospecting. Curr. Pharm. Des. 2021, 27, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A. Intersection of inflammation and herbal medicine in the treatment of osteoarthritis. Curr. Rheumatol. Rep. 2012, 14, 604–616. [Google Scholar] [CrossRef]
- Oskouie, M.N.; Aghili Moghaddam, N.S.; Butler, A.E.; Butler, A.E.; Zamani, P.; Sahebkar, A. Therapeutic use of curcumin-encapsulated and curcumin-primed exosomes. J. Cell Physiol. 2019, 234, 8182–8191. [Google Scholar] [CrossRef]
- Gleason, B.; Chisari, E.; Parvizi, J. Osteoarthritis Can Also Start in the Gut: The Gut-Joint Axis. Indian J. Orthop. 2022, 56, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Ramires, L.C.; Santos, G.S.; Ramires, R.P.; da Fonseca, L.F.; Jeyaraman, M.; Muthu, S.; Lana, A.V.; Azzini, G.; Smith, C.S.; Lana, J.F. The Association between Gut Microbiota and Osteoarthritis: Does the Disease Begin in the Gut? Int. J. Mol. Sci. 2022, 23, 1494. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Hong, W.; Qian, D.; Peng, F.; Li, H.; Liang, C.; Du, M.; Gu, J.; Mai, J.; Bai, B.; et al. Quercetin modulates the gut microbiota as well as the metabolome in a rat model of osteoarthritis. Bioengineered 2021, 12, 6240–6250. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule Tested | In Vitro Models | Doses | Main Effects | Specific Outcomes | Ref. |
|---|---|---|---|---|---|
| Genistein | IL-1β-treated human OA chondrocytes | 10 μM | Anti-inflammatory effect by stimulating Nrf2/HO1 signalling pathway |
| [25] |
| 6-gingerol | IL-1β-treated human OA chondrocytes | 1–2–5–10 μM | Anti-inflammatory and anti-oxidant effects due to NRF2/GSTA4-4 pathway |
| [26] |
| Resveratrol | IL-1β-treated human OA chondrocytes | 6.25–25–50–200 μM | Anti-inflammatory effect by targeting IL-1β dependent activation of TLR4/MyD88/NF-κB signaling pathway |
| [88] |
| Resveratrol | IL-1β-treated human OA chondrocytes | 6.25–12.5–25–50–100–200 μM | Inhibition of inflammation |
| [89] |
| Resveratrol | IL-1β-treated human chondrosarcoma cell line SW1353 | 50 μM | Anti-inflammatory effects by inhibition of TLR4/NF-κB signaling pathway |
| [90] |
| Resveratrol | Methylglyoxal-modified albumin (AGEs)-treated porcine chondrocytes | Pretreatment, 25–50–75–100 µg | Anti-inflammatory effect by inhibition of IKK-NF-KB and MAPK/AP- 1 signalling pathways |
| [91] |
| Resveratrol | IL-1β-treated human chondrocytes | 1–200 μM | Anti-inflammatory, anti-oxidative, and anti-apoptotic effects |
| [92] |
| Resveratrol | Normal and OA human chondrocytes | 1–10–25–50 µM | Modulation of epigenome (SIRT modulator) | ↑ SIRT1 | [93] |
| Resveratrol | IL-1β-treated human OA chondrocytes | 50 μM | ECM metabolism, autophagy, inflammation, apoptosis | ↑ SIRT1/FOXO1 signaling | [94] |
| Resveratrol | IL-1β-treated mouse OA chondrocytes | 2.3–23 µg/ml | SIRT1-dependent inhibition of inflammatory NF-kB-signaling |
| [95] |
| Resveratrol | IL-1β-treated human OA chondrocytes | Pretreatment, 25 μM | Modulator of epigenome |
| [96] |
| Resveratrol | Coculture model of human chondrocytes and macrophages | 10–25–50–100 μM | Inhibits the inflammatory amplification loop (IL-1β induced NF-κB and IL-6 in chondrocytes; IL-6 secreted activate STAT3 in macrophages; STAT3 positively regulate IL-6 secretion in chondrocytes). |
| [97] |
| Curcuminoids Resveratrol Quercetin Pterostilbene Hydroxypterostilbene | Equine lymphocytes | 10–20–40–80–160–320 μM | Decreased lymphocyte production of pro-inflammatory cytokines |
| [98] |
| Epigallocatechin 3-gallate | IL-1β treated chondrocytes | 20–50 μM | Anti-inflammatory effect by targeting PTEN/miRNA-29b pathway |
| [99] |
| Epigallocatechin-3-gallate | IL-1β -treated human chondrocytes | 10 to 100 uM | Anti-inflammatory response by inhibiting NF-κB and JNK-MAPK |
| [100] |
| Epigallocatechin-3-gallate | IL-1β-treated human OA chondrocytes | Pretreatment, 20–50 μM | Anti-inflammatory effects |
| [101] |
| Epigallocatechin-3-gallate | IL-1β-treated human OA chondrocytes | 100 μM | Inhibiting JNK |
| [102] |
| Epigallocatechin-3-gallate | Human fibroblast-like synoviocytes and THP-1 cells treated with CPP crystals in presence of methyl-β-cyclodextrin | 1–5–10 μM | Reduce the cytokine release induced by CPP crystals |
| [103] |
| Quercetin | Tert-butyl hydroperoxide (TBHP)-treated rat chondrocytes | 15–25–50–75–100 μM | Attenuate oxidative stress, ER stress, and associated apoptosis by activation of SIRT1/AMPK signalling |
| [104] |
| Quercetin | IL-1β-treated rat chondrocytes | Pretreatment, 8 µM | Anti-inflammatory and anti-apoptotic effects by suppressing IRAK1/NLRP3 signalling |
| [105] |
| Quercetin | IL-1β-treated articular cartilage human cells | 10–100 µM | Anti-inflammatory and anti-apoptotic effect by inhibiting p38 MAPK signalling pathway |
| [106] |
| Quercetin | IL-1β-treated rat chondrocytes | 2–4–8 μM | Anti-inflammatory, anti-catabolic, anti-apoptotic effect due to suppression of the Akt/NF-κB signalling pathway |
| [107] |
| Quercetin | RAW 264.7 cells | 8 μM | Promote M2 polarization and pro-chondrogenic microenvironment for chondrocytes |
| [107] |
| Quercetin | Coculture of RAW 264.7 cells and chondrocytes | 8 μM | Indirect chondrogenic effect mediated by monocytes |
| [107] |
| Hesperetin | IL-1β-treated chondrocytes | Anti-inflammatory effect by inhibition of NF-kB pathway |
| [108] | |
| Hesperetin | IL-1β-treated mice chondrocytes | Loaded in Gd2(CO3)3-based nanoparticles (NPs) | Anti-inflammatory and anti-apoptotic effects by inhibiting TLR-2/NF-κB/Akt signalling |
| [109] |
| Hesperidin | IL-1β-treated human OA chondrocytes | 2.0 mg/ml | Anti-inflammatory effect of NF-κB inhibition |
| [110] |
| Icariin | LPS-treated rat chondrocytes | 5–10–20 μM | Anti-inflammatory effect by inhibiting NLRP3 inflammasome-mediated caspase-1 signalling |
| [111] |
| Icariin | LPS-treated rat chondrocytes | 20 μM | Anti-inflammatory effect by inhibition of TLR4/Myd88/NF-κB pathway |
| [112] |
| Icariin | IL-1β-treated human SW1353 chondrosarcoma cells | 20 μM | Anti-inflammatory effect by inhibition of p38/ERK/JNK |
| [114] |
| Icariin | LPS-treated synoviocytes | 5–10 μM | Inhibition of ferroptosis during synovitis by the activation of the Xc-/GPX4 and NRF2 axis |
| [116] |
| Carnosol (CL), carnosic acid (CA), carnosic acid-12-methylether (CAME), 20-deoxocarnosol and abieta-8,11,13-triene-11,12,20-triol (ABTT) | LPS-treated RAW264.7 cells | 5–10–15 μM | Different anti-inflammatory potential |
| [118] |
| Carnosol (CL) | IL-1β-treated chondrosarcoma SW1353 cells | 6.25–12.5–25 μM | Anti-inflammatory and anti-catabolic effects |
| [118] |
| Carnosol (CL), carnosic acid-12-methylether (CAME), abieta-8,11,13-triene-11,12,20-triol (ABTT) | IL-1β-treated human articular chondrocytes from knee | 6.25–12.5 μM1.56–3.13 μM | Different anti-inflammatory potential |
| [118] |
| Caffeic acid phenethyl ester (CAPE) | IL-1β-treated human chondrocytes | 10–20 μM | Anti-inflammatory effects via inactivation of NFkβ signalling pathway due to activation of Nrf2/HO-1 signalling pathway |
| [119] |
| Tangeretin | IL-1β-treated mice chondrocytes | 5–10–20 μM | Anti-inflammatory effect by blocking NF-κB by activating Nrf2 and MAPK pathway |
| [120] |
| Nobiletin | IL-1β-treated mice chondrocytes | Pretreatment, 10–20–40 μM | Anti-inflammatory and anti-catabolic effects by inhibition of NF-kB pathway |
| [121] |
| Naringenin | IL-1β activated murine articular chondrocytes | 20–40 µM | Anti-inflammatory effect by inhibition of NF-kB pathway |
| [122] |
| Theaflavin-3-3′-digallate | IL-1β-treated rat chondrocytes | 20–40 μM | Anti-inflammatory effect by the PI3K/AKT/NF-κB and MAPK and Nrf2/HO-1 signaling pathway |
| [123] |
| Cyanidin | IL-1β-treated human OA chondrocytes | 12.5–25–50–100 μM | Anti-inflammatory effect by inhibiting NF-κB pathway |
| [125] |
| Ellagic acid | IL-1β-treated human chondrocytes | 50 μM | The anti-inflammatory effect through NF-κB inhibition |
| [126] |
| Ferulic acid | IL-1β-treated human OA chondrocyte | 1–5–10–20–20 μM | Anti-inflammatory effect by activating Sirt1/AMPK/PGC-1α signalling pathway. |
| [127] |
| Fisetin | IL-1β-treated human articular chondrocytes | Pre-treatment, 1–5–10–25–50 μM | Anti-inflammatory effect of SIRT-1 activation |
| [128] |
| Procyanidins | Chondrocytes | Attenuated apoptosis and senescence |
| [129] | |
| Curcumin | IL-1β-treated human chondrocytes | Exosomes derived from curcumin-treated MSC | Anti-inflammatory effect by targeting miR-124 and miR-143 |
| [130] |
| Vanillic Acid | IL-1β/TNF-α-treated human OA chondrocytes | 1 μM | Anti-inflammatory effect by targeting NF-κB signaling |
| [131] |
| Chlorogenic acid | IL-1β-treated rabbit chondrocytes | 5–10–20 μM | Anti-inflammatory effect by inhibition of NF-κB |
| [132] |
| Biochanin | IL-1β-treated rabbit chondrocytes | 5–25–50 μM | Anti-inflammatory effect via inhibition of IκBα/NF-κB activation |
| [133] |
| Sesamin | IL-1β- and/or OSM (long term) treated 3D human articular chondrocytes culture system | 0.25–0.5–1 μM | Anti-inflammatory effect and anti-catabolic effect |
| [135] |
| Sinapic acid | IL-1β-treated human OA chondrocytes | Pretreatment, 40–80–160–320 μM | Anti-inflammatory effects via activation of the Nrf2 signalling pathway |
| [136] |
| Sinapic acid | IL-1β-treated mice OA chondrocytes | 3–9–12 μM | Anti-inflammatory effects via activation of the Nrf2/HO-1 signalling pathway |
| [137] |
| Xanthohumol | IL-1β-treated rat chondrocytes | 10–25–50 μM | Anti-inflammatory effect by blocking the the activity of NF-κB via activating the Nrf2/HO-1 pathway |
| [138] |
| Myricetin | IL-1β-treated human chondrocytes | 5–10–15 μM | Anti-inflammatory effect by blocking NF-κB pathway and promoting PI3K/Akt mediated Nrf2/HO-1 axis |
| [139] |
| Luteolin | Murine chondrocyte | 20 µM | Anti-oxidant and anti-inflammatory effects by activating the AMPK/Nrf2 signalling pathway where AMPK acted as an upstream signal for Nrf2 |
| [140] |
| Delphinidin | H2O2-treated human chondrocytes | 40 μM | Anti-apoptotic effect by activating Nrf2 and NF-κB |
| [141] |
| Xanthohumol | IL-1β-treated rat chondrocytes | 5–20 μM | Anti-inflammatory effect by activating HO-1 signalling |
| [142] |
| AGEs-induced human articular chondrocytes | Pretreatment, 2.5–5–10 μM | Anti-inflammatory effect by targeting NF-kappaB and MAPK signalling |
| [143] |
| Vanillic Acid | IL-1β-treated rat chondrocytes | 5–10–20 μg/mL | Anti-inflammatory and anti hypertrophic effect by inhibition of MAPK and PI3K/AKT/NF-κB pathways |
| [125] |
| Rosmarinic acid | IL-1β-treated rat articular chondrocytes | Pre-treatment, 10–50–100 μM | Anti-inflammatory effect by NF-κB and MAPK pathway inhibition |
| [144] |
| Fisetin | LPS-treated RAW264.7 mouse macrophages | Anti-inflammatory effect by JNK and NF-κB pathway inhibition |
| [145] | |
| Punicalagin | LPS-treated rat chondrocytes | Pretreatment, 20–50–100 μM | Anti-inflammatory effect activating Foxo1 and downstream Prg4/HIF3α axis and autophagy |
| [147] |
| Curcumin | IL-1β-treated human chondrocytes |
| Anti-inflammatory and chondroprotective effect |
| [151] |
| Resveratrol | Mice chondrocytes seeded on an oxidized hyaluronic acid hydrogel added with resveratrol | 0.001 μM | Anti-inflammatory and anti-catabolic effects |
| [154] |
| Resveratrol | LPS-treated Mice chondrocytes seeded on an oxidized hyaluronic acid hydrogel added with resveratrol | 0.001 μM | Anti-inflammatory and anti-catabolic effects |
| [154] |
| Resveratrol | IL-1β-treated human chondrocytes | 100 μM | Anti-inflammatory and anti-apoptotic effects |
| [155] |
| Resveratrol | IL-1β-treated human chondrocytes | 100 μM | Anti-inflammatory and anti-apoptotic effects |
| [155] |
| Resveratrol | PBMC of healthy donors | 1–5 μM | Increased inflammatory mediators |
| [156] |
| Protocatechuic acid alone or loaded into a delivery system (MOF@HA@PCA) | IL-1β-treated OA chondrocytes | 6 μM | Anti-inflammatory effect and anti-catabolic effect |
| [157] |
| Gallic acid | AGE-treated rabbit chondrocytes | Pretreatment | Prevented AGE-related changes |
| [158] |
| p-Coumaric Acid (PCA) | IL-1β-treated rat chondrocytes | Anti-inflammatory effect and cellular senescence by targeting MAPK, NF-κB pathways |
| [159] | |
| Ferulic acid | H2O2-stimulated porcine chondrocytes | Pretreatment, 40 μM | Inhibition of inflammatory response |
| [160] |
| Sinapic acid | IL-1β-treated rat chondrocytes | Anti-inflammatory effects via inhibition of the MAPK pathway |
| [161] | |
| Caffeic acid | IL-1β-treated rat chondrocytes | 5–10–20 μM | Anti-inflammatory effects via inactivation of NF-κB due to activation of the JNK pathway |
| [162] |
| Chlorogenic acid | IL-1β-treated human SW-1353 chondrocytes | 31.25–62.5–125–250–500–1000 μM | Anti-inflammatory and effect via inhibition of IκBα/NF-κB activation |
| [163] |
| Chlorogenic acid | IL-1β-treated human OA chondrocytes | 5–10–20 μM | Anti-inflammatory effect inhibiting the iNOS-NO signalling pathway |
| [164] |
| Formononetin | Human subchondral osteoblasts | 0.01–0.1–1–10–100 μM | Inhibition of inflammatory mediators |
| [165] |
| Genistein | IL-1β-treated OA chondrocytes | 25–50–100 μM | Inhibition of inflammatory mediators |
| [166] |
| Biochanin | IL-1β-treated rat chondrocytes | 5–7.5–10–15 μM | Anti-inflammatory, anti-catabolic, anti-oxidant responses via inhibition of NF-κB activation |
| [167] |
| Phloretin | IL-1β-treated human OA chondrocytes | 10–30–100 μM | Attenuates inflammatory responses via inhibition of PI3K/Akt, NF-κB activation |
| [168] |
| Apigenin | IL-1β-treated articular mice chondrocytes or IL-1β-treated cartilage explants | Pretreatment, 10–25–50 µM | Antioxidant and anti-inflammatory effect via Hif-2α regulation by JNK and NF-κB Pathways |
| [169] |
| Apigenin | IL-1β-treated rabbit articular chondrocytes | 1–10–50–100 µM | Anti-catabolic effect |
| [170] |
| Luteolin | Cartilage cells of OA guinea pigs | Anti-inflammatory effect |
| [171] | |
| Luteolin | IL-1β-treated rabbit articular chondrocytes | 1–10–50–100 µM | Inhibited IL-1β-induced cartilage degradation |
| [172] |
| Kaempferol | Osteoarthritis chondrocytes | Decreased proinflammatory cytokine production and ECM degradation |
| [173] | |
| Kaempferol | IL-1β-treated rat chondrocytes | 25–50–100–200 μM | Anti-inflammatory effect by inhibition of NF-kB pathway |
| [174] |
| Kaempferol | IL-1β-treated rat chondrocytes | Anti-inflammatory effect by inhibition of MApK-p38 pathway |
| [175] | |
| Epigallocatechin-3-gallate | Human chondrocytes | Anti-inflammatory effect |
| [176] | |
| Isorhamnetin | IL-1β-treated human OA chondrocytes | Pretreatment, 10–50–100 µM | Anti-inflammatory effect by inhibition of NF-kB pathway |
| [177] |
| Hesperetin | TNF-α-treated rat (SD) chondrocytes | 1–5–10–20–50–100 μM | Anti-inflammatory effect |
| [178] |
| Hesperidin | H2O2-treated rat chondrocytes | Pretreatment, 0.1 µM | Anti-oxidant and anti-inflammatory effects |
| [179] |
| Icariin | IL-1β-treated rat chondrocytes | Anti-inflammatory effect by NF-κB pathway inhibition |
| [180] | |
| Icariin | Human osteoarthritis fibroblast-like synoviocytes (OA-FLSs) | 0.1–0.5–1.5–10 μM | Anti-inflammatory effect |
| [181] |
| Delphinidin | IL-1β-treated human chondrocytes | Pretreatment, 10–50–100 μM | Anti-inflammatory effect by inhibiting NF-κB pathway |
| [182] |
| Avenanthramide | IL-1β-treated human chondrocytes | 50–100–200 μM | Anti-catabolic effect by modulating p38/JNK pathway |
| [183] |
| Resveratrol | IL-1β-treated human OA chondrocytes | 100 μM | Inhibits NFkB pathway |
| [184] |
| Curcumin Resveratrol | IL-1β-treated human OA chondrocytes | 10 μM | Anti-apoptotic effect by inhibition of MAPK/ERK pathway |
| [185] |
| IL-1β-treated human OA chondrocytes | 50 μM | Anti-inflammatory effects and anti-catabolic effect by inhibition of NF-κB |
| [186] |
| Resveratrol | Adult human OA chondrocytes | 10 μM | Anti-inflammatory effects |
| [187] |
| Resveratrol | IL-1β-treated human OA chondrocytes | 24–48 μM | Anti-inflammatory effects |
| [188] |
| Amurensin H (resveratrol dimer) | IL-1β-treated rat OA chondrocytes | 4–8 μmol/L | Anti-inflammatory effects |
| [189] |
| Resveratrol | IL-1β-treated human chondrocytes | Pretreatment, 1–10 μM | Anti-inflammatory and anti-catabolic effects |
| [190] |
| IL-1β-treated human chondrocytes | 6.25–12.5–25–50 μM | Anti-catabolic effect by inhibiting NFĸB, ERK, MAPK pathway |
| [191] |
| Prodelphinidins | Human chondrocytes | Anti-inflammatory effect |
| [192] | |
| Epigallocatechin-3-gallate | AGE-BSA-treated human chondrocytes | Pretreatment, 25–50–100–200 μM | Reduced AGE-induced inflammation by inhibiting the MAPK and NF-kB activation |
| [193] |
| Epigallocatechin-3-gallate | IL-1β-treated human OA chondrocytes | 1–100 μM | Inhibiting NF-kB/not through IKK but through the proteasome pathway |
| [194] |
| Epigallocatechin-3-gallate | IL-1β-treated human chondrocytes | Pretreatment, 100–200 μM | Anti-inflammatory effect |
| [195] |
| Carnosic acid (CA) | IL-1β-treated human chondrocytes | 1–50 μM | Prevented IL-1β-dependent decrease in HO-1 by increasing miR-140 |
| [196] |
| Carnosic acid (CA) | Mouse femoral head cartilage explants | 1–50 μM | Suppressed proteoglycan release |
| [196] |
| Sesamin | IL-1β-treated porcine cartilage explant culture systems | 0.25–0.5–1 μM | Anti-inflammatory and anti-catabolic effects |
| [197] |
| Molecule Tested | In Vivo OA Models | Doses and Duration Delivery Routes | Main Effects | Specific Outcomes | Ref. |
|---|---|---|---|---|---|
| Resveratrol | MIA-induced OA model in mice | 10, 100 µg administration after 1 and 4 weeks from OA induction IA injections | Prevention of OA progression |
| [95] |
| Quercetin (QUE) | MMTL-(DMM) induced OA in rats | 50–100 mg/kg QUE, once daily for 12 weeks Intraperitoneal injection | Reduced cartilage degeneration, apoptosis, ER stress |
| [104] |
| Quercetin | ACLT-induced OA in rats | 50–100 mg/kg once daily for 12 consecutive weeks Intraperitoneal injection | Anti-inflammatory and chondroprotective effects |
| [105] |
| Quercetin | MMT + AMTL-induced OA in rats | 8 μM once a week for 6 consecutive weeks IA injection | Anti-inflammatory and immunomodulatory effects |
| [107] |
| Hesperetin-Gd2(CO3)3@PDA-PEG-DWpeptide, (HGdPDW) Gd2(CO3)3-based nanoparticles (NPs) | ACLT-induced OA in mice | 5% (w/w) 3 times/week for 8 weeks IA injection | Alleviation of cartilage degeneration and apoptosis |
| [109] |
| Icariin | MIA-induced OA in rats | 20 μM 2 weeks post-MIA injection IA injection | Protective effect on OA degradation by inhibition ofNLRP3-mediated pyroptosis |
| [111] |
| Icariin | KOA rat model established by the Hulth method | 20 mg/kg, daily Intra-gastric administration | Protective effect on cartilage and subchondral bone degeneration; neuromodulation; analgesic effect |
| [112] |
| Caffeic acid phenethyl ester (CAPE) | DMM model in rats | 10 mg/kg/2 days for 8 weeks from DMM surgery Intraperitoneal injection | Delay of OA progression |
| [119] |
| Tangeretin | DMM-induced OA in mice | 10–20 mg/kg daily intragastric dose | Anti-degenerative and anti-inflammatory effects by blocking NF-κB by activating Nrf2 and MAPK signalling |
| [120] |
| Nobiletin | DMM-induced OA in mice | 20 mg/kg every 2 days for 8 weeks Intraperitoneal injection | Delay of OA progression |
| [121] |
| Naringenin | MIA-induced OA in rats | 20–40 mg/kg daily, two weeks after OA induction Intragastric delivery | Pain alleviation |
| [122] |
| Theaflavin-3,3′-Digallate | DMM model in rats | 4 mM, 2 weeks after OA induction every 2 days for 6 weeks IA injection | Protection for cartilage degradation |
| [123] |
| Ellagic acid | DMM mouse model | 40 mg/kg intragastric administration, every 2 days for 8 weeks | Prevent the progression of OA degeneration and inflammation |
| [126] |
| Fisetin | DMM-induced OA in mice | 20 mg/kg daily for 8 weeks after surgery Oral gavage | Reduction in synovitis and joint destruction |
| [128] |
| Curcumin | OA mouse model | exosomes derived from curcumin-treated MSCs | Inflammatory markers modulation |
| [130] |
| Pinoresinol diglucoside | ACLT-induced OA in rabbits | 2 mg/mL after 4 weeks from OA induction, once a week for 5 weeks IA injection | Cartilage-protecting effects |
| [134] |
| Sinapic acid | DMM model in mice | 10 mg/kg/day from DMM surgery Intragastric delivery | Prevention of OA progression by reducing inflammation and catabolism |
| [137] |
| Xanthohumol | DMM model in mice | 40 mg/kg XN in 0.5% carboxymethylcellulose (CMC) daily for 8 weeks ntragastric delivery | Safeguarding influence in OA advancement |
| [138] |
| Myricetin | DMM-induced OA in mice | 20 mg/kg myricetin in 0.5% carboxymethylcellulose (CMC), every two days for 8 weeks Intragastric administration | Reduction in joint destruction |
| [139] |
| Luteolin | DMM-induced OA in rats | 10 mg/kg/day in 0.5% carboxymethylcellulose (CMC) or vehicle alone (0.5% CMC) for 8 consecutive weeks Intra-gastric delivery | Prevention of cartilage damage |
| [140] |
| Punicalagin | ACLT-induced OA model in rats | 10 mg/kg/day Oral gavage | Alleviates the inflammatory injury and ECM degradation through Foxo1/Prg4/HIF3α pathway activation and autophagy |
| [147] |
| Quercetin | MMX + ACLT + PCLT-induced OA in rabbits | 25 mg/kg from the 5th week on, once daily for 4 weeks Intragastric delivery | Reduction in OA progression |
| [148] |
| Quercetin encapsulated in the mPEG-polypeptide thermogel | ACLT-induced OA model in rats | 50 or 500 μg IA injection | Relief of pain symptoms and delay of OA progression |
| [149] |
| Palmitoylethanolamide co-ultra micronized with quercetin (PEA-Q) | carrageenan paw oedema and MIA-induced OA models in rats | 10 mg/kg three times per week for 4 weeks Oral administration | Reduction in inflammatory and pain responses | carrageenan paw oedema
| [150] |
| Curcumin | MMTL-induced OA in mice | curcumin nanoparticles (0.07 mg of 10 μg curcumin/1 mg nanoparticles) once daily for 8 weeks Topic administration | Delay of OA progression and pain relief no significant effect on OA pain relief. |
| [151] |
| Next Generation Ultrasol Curcumin (NGUC) | MIA-induced OA in rats | 100 mg/kg of NGUC (20 mg/kg of curcuminoids) 200 mg/kg of NGUC (40 mg/kg of curcuminoids) Oral administration for 4 weeks | Enhanced bioavailability of curcumin and amelioration of OA pathophysiology |
| [152] |
| Malvidin | MIA-induced OA in rats | 10, 20 mg/kg/day, dissolved in saline (vehicle)] was administered for 14 days starting on the same day of MIA injection IA injection | Pain-relieving effects |
| [153] |
| PCA loaded into a delivery system (MOF@HA@PCA) pH-responsive nanoparticle system | ACLT-induced OA in rats | 6 μg/mL(once/week) for 4 and 8 weeks from ACLT surgery IA injection | Attenuation of OA progression |
| [157] |
| Genistein | DMM + ACLT + MCLT-induced OA in rats | 20 mg/kg/each day for 6 weeks Intragastric delivery | Suppression of inflammation, cartilage degradation and apoptosis |
| [166] |
| Apigenin | The IL1-β-treated knee joint of rats | 50–100 μM pretreatment IA injection | Chondroprotection |
| [170] |
| Luteolin | IL-1β-treated rabbit articular chondrocytes | 50–100 μM pretreatment Articular injection | Chondroprotection |
| [172] |
| Hesperidin | Medial collateral ligament transection + medial meniscus removal- induced OA model in rats | 200 mg/kg body weight once 118 days for 28 days Oral administration | Delay of cartilage degeneration |
| [178] |
| Sesamin | Papain-induced OA model in rats | 1,10 μM sesamin injection every 5 days for 25 days | Inhibit the pathological progression |
| [196] |
| Apigenin and synovial membrane-derived mesenchymal stem cells (SMMSCs) | ACLT-induced OA in rats | Apigenin (0.1–0.3 μM) ± SMMSC (3 × 106) once weekly for 3 weeks IA injection | Suppression of inflammation and oxidative stress supplementary benefits when combined with cell-based therapy |
| [198] |
| Apigenin and kaempferol ± SMMSCs | ACLT-induced OA in rats | kaempferol (10 or 20 μM) + apigenin (0.1 μM) weekly, for three weeks IA injection | Coupling the two molecules has synergistic benefits in reducing inflammation and cartilage degradation |
| [199] |
| Capsaicin | MIA-induced OA in rats | 0.5% two weeks before OA induction single IA injection | Suppression of mechanical pain and bone erosion |
| [200] |
| Epigallocatechin 3-gallate | Spontaneous OA in guinea pig | 10 µM once a week for 12 consecutive weeks from 6 months of age IA Injection | Inhibition of age-related-OA changes |
| [201] |
| Epigallocatechin 3-gallate | ACLT-induced OA model in rats | 10 μM once every three days for 5 weeks, 2 weeks after the operation IA Injection | Prevention of inflammation and the promotion of autophagy |
| [202] |
| Genistein | MIA-induced OA in rats | 30 mg/kg/day for 8 weeks Intragastric delivery | Prevention of cartilage damage |
| [203] |
| Protocatechuic acid (PCA) | ACLT-induced OA in rats | 50 mg/kg/day for 4 weeks from ACLT surgery Intragastric delivery | Suppression of osteoclastogenesis |
| [204] |
| Poly(ethylene glycol) (PEG)-formononetin (FMN) nanodrug | ACLT-induced OA in rats | 1.25 μg/mL (once per week) for 4 or 8 weeks IA injection | Anti-inflammatory and chondroprotective effects |
| [205] |
| Quercetin | Papain-induced OA in rats | 1, 4 and 7 days post-quercetin treatment 5–10 mg/kg Intragastric delivery | Prevention of OA progression |
| [206] |
| Quercetin | MIA-induced OA model in mice | 20 mg/kg for 28 days Intragastric delivery | Reduction in OA symptoms and cartilage degradation |
| [207] |
| Quercetin | MNX-induced OA model in rats | 100 mg/kg on day 0 for consecutive 7 days once daily Oral administration | Relief of OA conditions |
| [208] |
| Quercetin-loaded lecithin-chitosan nanoparticles | DMM and MIA-induced OA models in rats | Dose 1 (0.84 mg/g gel), dose 2 (1.68 mg/g gel), dose 3 (3.36 mg/g gel) on the 29th day Topic administration | Prevention of cartilage degradation |
| [209] |
| Resveratrol | DMM-induced OA model in rats | 0.8 mL of 5, 10, and 15 μmol/L, 6 weeks after OA induction IA injection | Cartilage protection |
| [210] |
| Resveratrol | MIA-induced OA model in rats | 5 or 10 mg/kg body/daily after OA induction for up to 14 days Oral administration | Inhibition of inflammatory mediators |
| [211] |
| Resveratrol | MIA-induced OA model in rats | 50 μg/10 μL, 250 μg/10 μL, and 500 μg/10 μL for 8 consecutive days from day 14 today 21 after OA inductions Intrathecal administration | Reduction in mechanical allodynia |
| [212] |
| Resveratrol | Type II collagenase-induced OA model in rats | 40 and 80 mg/kg, twice daily for 4 weeks Intragastric administration | Reduction in joint destruction via anti-inflammatory and antiapoptotic effects |
| [213] |
| Resveratrol | Obesity-related OA model: high-fat diet (HFD)-induced OA in mice | 22.5 and 45 mg/kg for 12 weeks, after OA induction. Intragastric administration | Alleviation of OA pathology via the reduction in systemic inflammation |
| [214] |
| OA induced by treadmill-running exercise in rats | 25 mg Resveratrol or RLTA (containing 25 mg Res), daily for 3 weeks Intra-gastric administration | Reduction in inflammation |
| [215] |
| Vanillic acid | MIA-induced OA in rats | 30 mg/kg/day for 14 days from MIA Intragastric delivery | Reduction in synovitis and pain-related behaviour |
| [216] |
| Molecule Tested | Study Types | Patients Data | Treatments (Route-Doses) | Follow-Up | Main Effects | Ref. |
|---|---|---|---|---|---|---|
| Curcumin (Cur) versus diclofenac (Df) | Randomized open-label parallel-arm study | knee OA VAS score ≥ 4 N = 140 N = 71 Cur + Df (50 ♂ + 21♀) N = 69 Df (48 ♂ + 21♀) aged 38–65 year | Cur complex 500 mg + Df 50 mg (Cur + Df)/ 2 times daily for 28 days Df 50mg alone 2 times daily for 28 days Oral administration | Baseline, Day 14 Day 28 |
| [217] |
| Curcumin-phosphatidylcholine phytosome complex (Meriva®) | clinical trial | knee OA mild to moderate pain N = 100 N = 50 NSAIDs N = 50 Meriva® | NSAIDs vs. Meriva® Two 500 mg tablets daily (200 mg curcumin/day) Oral administration | Baseline, 8 months |
| [218] |
| Resveratrol | Open-labelednoncontrolled clinical trial | knee OA mild-to-moderate OA N = 28 (8 ♂ + 20♀) KOOS = 46 ± 13 VAS = 74 ± 8 | 500 mg/day in a single oral dose for 90 days (monotherapy) Oral administration | Baseline, day 30, 3 months |
| [219] |
| Resveratrol + meloxicam | Randomized Placebo-Controlled Study | knee OA mild-to-moderate OA N = 110 VAS = 100
| 15 mg/day meloxicam and either 500 mg/day Resveratrol or placebo for 90 days Oral administration | 12 weeks 90-days |
| [220] |
| Combination of a glucosamine-chondroitin-quercetin glucoside (GCQG) | Clinical study | Knee OA mild-to-moderate OA 62.5 ± 9.4 years N = 46 (7 ♂ + 39 ♀) | glucosamine hydrochloride (1200 mg/d), shark cartilage powder (300 mg/d) containing 75–111 mg of chondroitin, and 45 mg of quercetin Oral administration | 3 months |
| [221] |
| Glucosamine hydrochloride, chondroitin sulfate and quercetin glycosides(GCQ supplement) | A randomized double-blind, placebo-controlled study | Knee OA N = 40 N = 20 GCQ treatment (4 ♂ + 16♀) N = 20 placebo (3 ♂ + 17♀) KL grade I-II | 1200 mg glucosamine hydrochloride, 60 mg chondroitin sulfate and 45 mg quercetin glycosides per day Dietary supplementation | 16 weeks |
| [222] |
| Capsaicin cream | Randomized, open-label series of n-of-1 trials | Radiographic knee OA N = 22 17 participants completed the trial. | Participants received 5% w/w ibuprofen gel and 0025% w/w capsaicin cream four times/day three treatment cycles (six paired periods) Topic administration | 3 cycles, each comprising one treatment for 4 weeks, an individualized washout period (maximum 4 weeks), then the other treatment for 4 weeks | Of 22 participants, 4 (18%) had a greater response to ibuprofen, 9 (41%) to capsaicin, 4 (18%) had similar responses, and 5 (23%) were undetermined | [223] |
| IA Trans-Capsaicin (CNTX) | Randomized, Double-Blind, Placebo-Controlled Trial | Knee OA moderate-to-severe OA 45–80 years N = 172 N = 69 placebo group N = 33 CNTX-4975 0.5 mg group N = 70 CNTX-4975 1.0 mg group | Treatments: Placebo, CNTX-4975 0.5 mg, or CNTX-4975 1.0 mg Single IA injection | 12 weeks 24 weeks |
| [224] |
| Pycnogenol® | A randomized, double-blind, placebo-controlled trial with a parallel group design | Knee OA N = 37 N = 19 Pycnogenol (♂ + 17♀) N = 18 Placebo (1 ♂ + 17♀) | Patients received either a placebo or Pycnogenol pills (50 mg, three times daily) for 3 months | 60 days 90 days | ↓ pain, stiffness, physical function, and composite WOMAC score | [225] |
| Pycnogenol® | A double-blind, placebo-controlled study | Knee OA mild to moderate OA N = 156 I or II OA grade (X-ray analysis). | Pycnogenol group versus the placebo group 100 mg/daily Oral administration | Baseline 3 months |
| [226] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambari, L.; Cellamare, A.; Grassi, F.; Grigolo, B.; Panciera, A.; Ruffilli, A.; Faldini, C.; Desando, G. Overview of Anti-Inflammatory and Anti-Nociceptive Effects of Polyphenols to Halt Osteoarthritis: From Preclinical Studies to New Clinical Insights. Int. J. Mol. Sci. 2022, 23, 15861. https://doi.org/10.3390/ijms232415861
Gambari L, Cellamare A, Grassi F, Grigolo B, Panciera A, Ruffilli A, Faldini C, Desando G. Overview of Anti-Inflammatory and Anti-Nociceptive Effects of Polyphenols to Halt Osteoarthritis: From Preclinical Studies to New Clinical Insights. International Journal of Molecular Sciences. 2022; 23(24):15861. https://doi.org/10.3390/ijms232415861
Chicago/Turabian StyleGambari, Laura, Antonella Cellamare, Francesco Grassi, Brunella Grigolo, Alessandro Panciera, Alberto Ruffilli, Cesare Faldini, and Giovanna Desando. 2022. "Overview of Anti-Inflammatory and Anti-Nociceptive Effects of Polyphenols to Halt Osteoarthritis: From Preclinical Studies to New Clinical Insights" International Journal of Molecular Sciences 23, no. 24: 15861. https://doi.org/10.3390/ijms232415861
APA StyleGambari, L., Cellamare, A., Grassi, F., Grigolo, B., Panciera, A., Ruffilli, A., Faldini, C., & Desando, G. (2022). Overview of Anti-Inflammatory and Anti-Nociceptive Effects of Polyphenols to Halt Osteoarthritis: From Preclinical Studies to New Clinical Insights. International Journal of Molecular Sciences, 23(24), 15861. https://doi.org/10.3390/ijms232415861