
INO80 Is Required for the Cell Cycle Control, Survival, and Differentiation of Mouse ESCs by Transcriptional Regulation

 ,
,  , ,
, ,  and
and 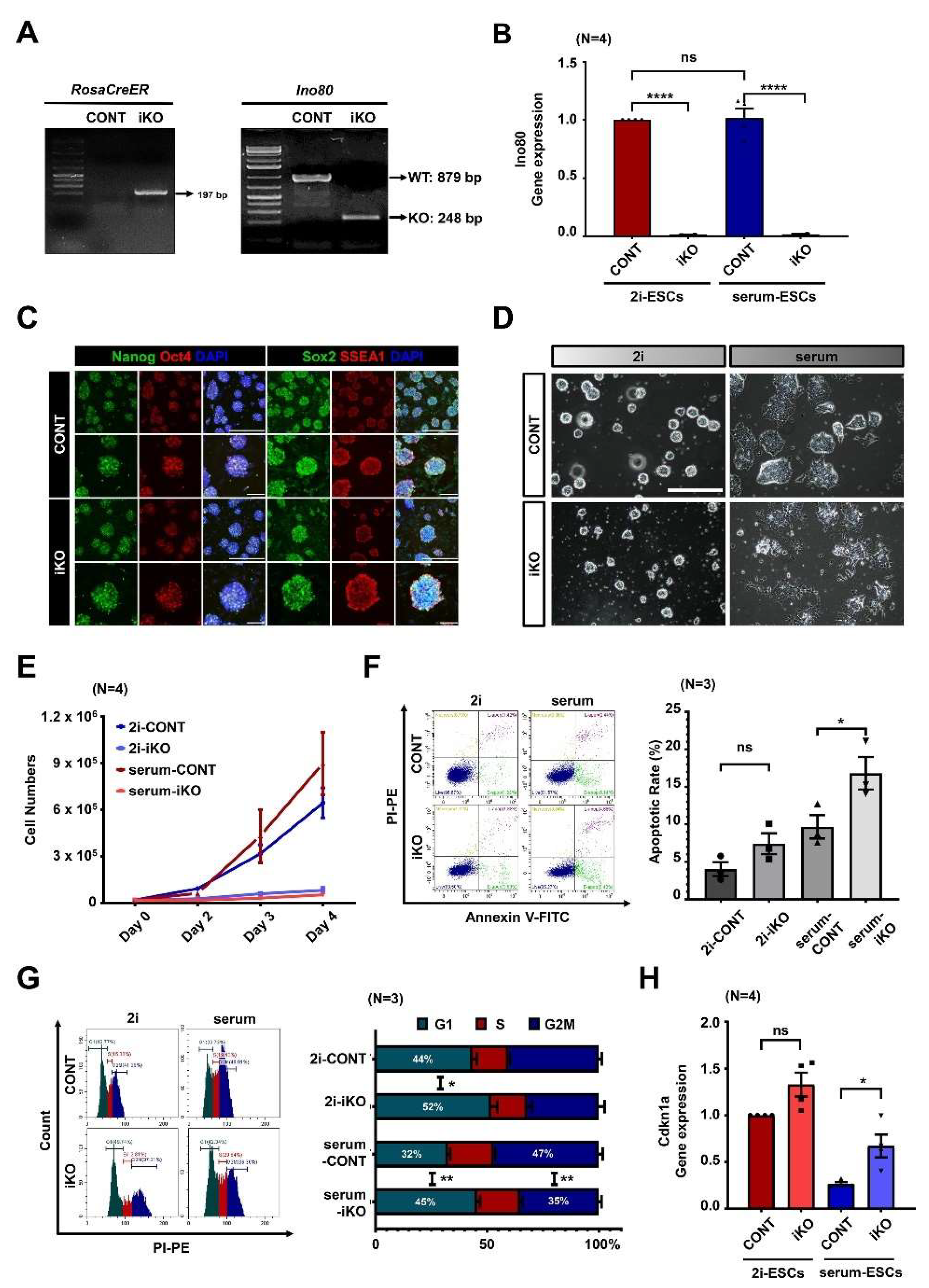{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ino80 Loss in ESCs Causes Aberrant Cell Cycle and Cell Death
2.2. Ino80 Loss Alters Expression of Genes Related to Cell Cycle
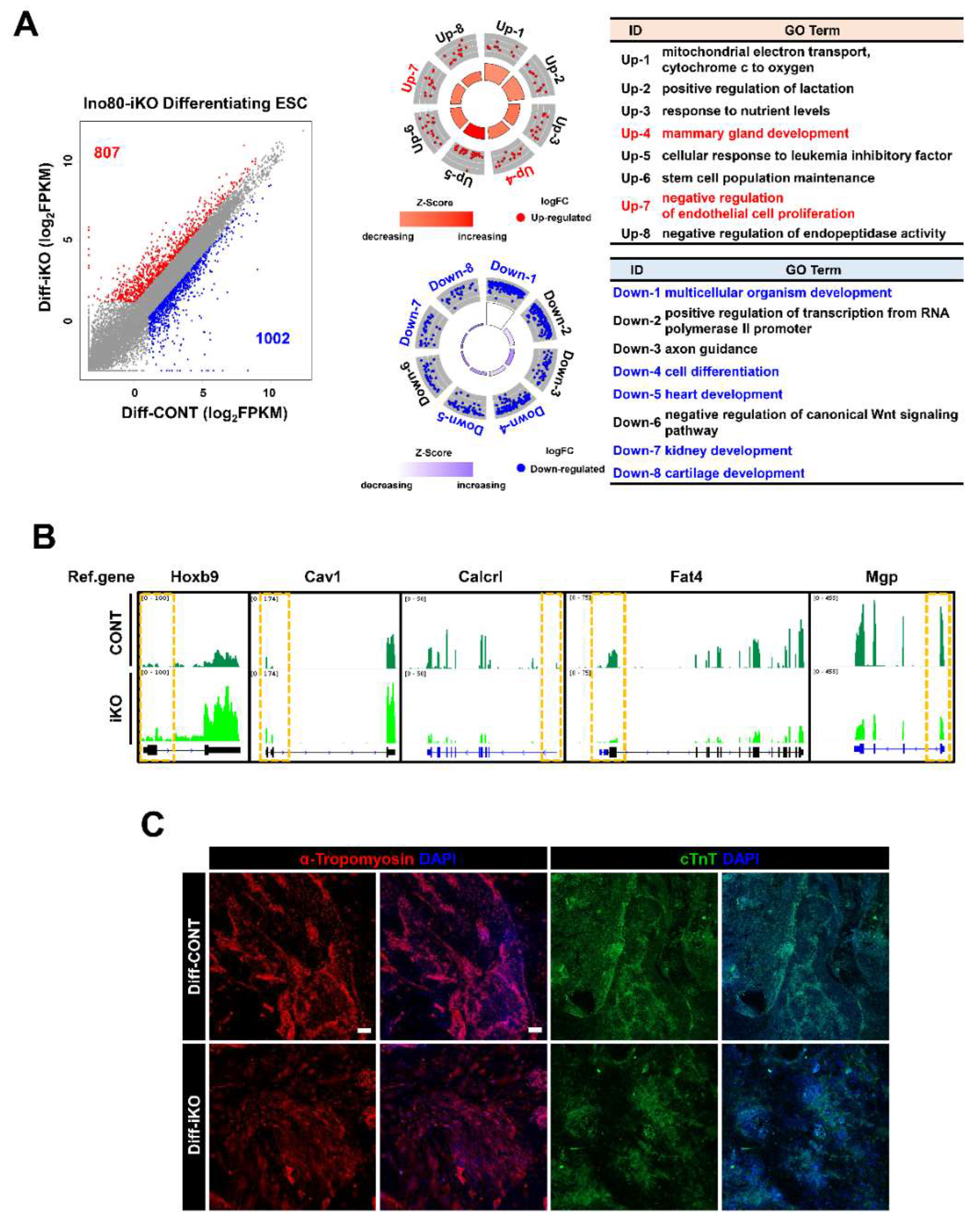
2.3. Abnormal Cell Cycle Profile Is Persistent in Differentiating ESCs
2.4. Ino80 Loss Impairs Expression of Genes Involved in Differentiation of Mesodermal Lineages
2.5. Conserved and Unique Alteration of Gene Expression between Undifferentiated and Differentiating iKO ESCs
3. Discussion
4. Materials and Methods
4.1. Animals and Generation of Ino80 iKO ESCs
4.2. ESC Differentiation
4.3. PCR Genotyping
4.4. Analyses of Cell Proliferation, Cell Cycle, and Apoptosis
4.5. Immunofluorescence
4.6. Quantitative RT-PCR Analysis
4.7. RNA-seq and ChIP-seq Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- White, J.; Dalton, S. Cell cycle control of embryonic stem cells. Stem Cell Rev. Rep. 2005, 1, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Soufi, A.; Dalton, S. Cycling through developmental decisions: How cell cycle dynamics control pluripotency, differentiation and reprogramming. Development 2016, 143, 4301–4311. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, S.; Dalton, S. Molecular and biological properties of pluripotent embryonic stem cells. Gene Ther. 2008, 15, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Ter Huurne, M.; Chappell, J.; Dalton, S.; Stunnenberg, H.G. Distinct cell-cycle control in two different states of mouse pluripotency. Cell Stem Cell 2017, 21, 449–455.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ter Huurne, M.; Peng, T.; Yi, G.; van Mierlo, G.; Marks, H.; Stunnenberg, H.G. Critical role for P53 in regulating the cell cycle of ground state embryonic stem cells. Stem Cell Rep. 2020, 14, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Faast, R.; White, J.; Cartwright, P.; Crocker, L.; Sarcevic, B.; Dalton, S. Cdk6–cyclin D3 activity in murine ES cells is resistant to inhibition by p16INK4a. Oncogene 2004, 23, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Stead, E.; White, J.; Faast, R.; Conn, S.; Goldstone, S.; Rathjen, J.; Dhingra, U.; Rathjen, P.; Walker, D.; Dalton, S. Pluripotent cell division cycles are driven by ectopic Cdk2, cyclin A/E and E2F activities. Oncogene 2002, 21, 8320–8333. [Google Scholar] [CrossRef] [Green Version]
- Fujii-Yamamoto, H.; Kim, J.M.; Arai, K.-I.; Masai, H. Cell cycle and developmental regulations of replication factors in mouse embryonic stem cells. J. Biol. Chem. 2005, 280, 12976–12987. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.; Crabtree, G.R. Chromatin remodelling during development. Nature 2010, 463, 474–484. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Ding, J.; Dong, L.; Zhao, J.; Su, J.; Wang, L.; Sui, Y.; Zhao, T.; Wang, F.; Jin, J.; et al. Negative regulation of p21Waf1/Cip1 by human INO80 chromatin remodeling complex is implicated in cell cycle phase G2/M arrest and abnormal chromosome stability. PLoS ONE 2015, 10, e0137411. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.; Chung, J.I.; King, D.A.; D’Amato, G.; Paik, D.T.; Duan, A.; Chang, A.; Nagelberg, D.; Sharma, B.; Jeong, Y.; et al. Endothelial deletion of Ino80 disrupts coronary angiogenesis and causes congenital heart disease. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Zou, J.; Zou, S.; Li, X. INO80 is required for osteogenic differentiation of human mesenchymal stem cells. Sci. Rep. 2016, 6, 35924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutt, C.; Hallmann, A.; Hachim, S.; Klockner, I.; Valussi, M.; Atzberger, A.; Graumann, J.; Braun, T.; Boettger, T. Linc- MYH configures INO 80 to regulate muscle stem cell numbers and skeletal muscle hypertrophy. EMBO J. 2020, 39, e105098. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Elsayed, Z.; Peterkin, V.; Alkatib, S.; Bennett, D.; Landry, J.W. Ino80 is essential for proximal-distal axis asymmetry in part by regulating Bmp4 expression. BMC Biol. 2016, 14, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, J.-N.; Tian, Y.; Xiao, Y.; Wu, L.; Li, L.; Chang, S. The mINO80 chromatin remodeling complex is required for efficient telomere replication and maintenance of genome stability. Cell Res. 2013, 23, 1396–1413. [Google Scholar] [CrossRef] [Green Version]
- Alatwi, H.E.; Downs, J.A. Removal of H2A.Z by INO 80 promotes homologous recombination. EMBO Rep. 2015, 16, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Magnuson, T. INO80 requires a polycomb subunit to regulate the establishment of poised chromatin in murine spermatocytes. Development 2022, 149, dev200089. [Google Scholar] [CrossRef]
- Xian, S.; Zhang, Y.; Wang, L.; Yao, F.; Ding, J.; Wang, Y.; Yang, X.; Dai, F.; Yin, T.; Cheng, Y. INO80 participates in the pathogenesis of recurrent miscarriage by epigenetically regulating trophoblast migration and invasion. J. Cell. Mol. Med. 2021, 25, 3885–3897. [Google Scholar] [CrossRef] [PubMed]
- Gospodinov, A.; Vaissiere, T.; Krastev, D.B.; Legube, G.; Anachkova, B.; Herceg, Z. Mammalian Ino80 mediates double-strand break repair through its role in DNA end strand resection. Mol. Cell. Biol. 2011, 31, 4735–4745. [Google Scholar] [CrossRef] [Green Version]
- Niculescu, A.B.; Chen, X.; Smeets, M.; Hengst, L.; Prives, C.; Reed, S.I. Effects of p21 Cip1/Waf1 at both the G 1 /S and the G 2 /M cell cycle transitions: pRb is a critical determinant in blocking DNA replication and in preventing endoreduplication. Mol. Cell Biol. 1998, 18, 629–643. [Google Scholar] [CrossRef]
- Liu, C.; Wang, R.; He, Z.; Osteil, P.; Wilkie, E.; Yang, X.; Chen, J.; Cui, G.; Guo, W.; Chen, Y.; et al. Suppressing nodal signaling activity predisposes ectodermal differentiation of epiblast stem cells. Stem Cell Rep. 2018, 11, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Wang, L.; Zhang, S.; Bennett, B.D.; He, F.; Zhang, Y.; Xiong, C.; Han, L.; Diao, L.; Li, P.; et al. INO80 governs superenhancer-mediated oncogenic transcription and tumor growth in melanoma. Genes Dev. 2016, 30, 1440–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-S.; Lee, S.-A.; Hur, S.-K.; Seo, J.-W.; Kwon, J. Stabilization and targeting of INO80 to replication forks by BAP1 during normal DNA synthesis. Nat. Commun. 2014, 5, 5128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Du, Y.; Ward, J.M.; Shimbo, T.; Lackford, B.; Zheng, X.; Miao, Y.-L.; Zhou, B.; Han, L.; Fargo, D.C.; et al. INO80 facilitates pluripotency gene activation in embryonic stem cell self-renewal, reprogramming, and blastocyst development. Cell Stem Cell 2014, 14, 575–591. [Google Scholar] [CrossRef] [Green Version]
- Cheon, Y.; Han, S.; Kim, T.; Hwang, D.; Lee, D. The chromatin remodeler Ino80 mediates RNAPII pausing site determination. Genome Biol. 2021, 22, 1–29. [Google Scholar] [CrossRef]
- Prendergast, L.; McClurg, U.L.; Hristova, R.; Berlinguer-Palmini, R.; Greener, S.; Veitch, K.; Hernandez, I.; Pasero, P.; Rico, D.; Higgins, J.M.G.; et al. Resolution of R-loops by INO80 promotes DNA replication and maintains cancer cell proliferation and viability. Nat. Commun. 2020, 11, 4534. [Google Scholar] [CrossRef]
- Keil, J.M.; Doyle, D.Z.; Qalieh, A.; Lam, M.M.; Funk, O.H.; Qalieh, Y.; Shi, L.; Mohan, N.; Sorel, A.; Kwan, K.Y. Symmetric neural progenitor divisions require chromatin-mediated homologous recombination DNA repair by Ino80. Nat. Commun. 2020, 11, 3839. [Google Scholar] [CrossRef]
- Yu, H.; Wang, J.; Lackford, B.; Bennett, B.; Li, J.-L.; Hu, G. INO80 promotes H2A.Z occupancy to regulate cell fate transition in pluripotent stem cells. Nucleic Acids Res. 2021, 49, 6739–6755. [Google Scholar] [CrossRef]
- Cai, Y.; Jin, J.; Yao, T.; Gottschalk, A.J.; Swanson, S.K.; Wu, S.; Shi, Y.; Washburn, M.; Florens, L.; Conaway, R.C.; et al. YY1 functions with INO80 to activate transcription. Nat. Struct. Mol. Biol. 2007, 14, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Gorski, J.W.; Ueland, F.R.; Kolesar, J.M. CCNE1 amplification as a predictive biomarker of chemotherapy resistance in epithelial ovarian cancer. Diagnostics 2020, 10, 279. [Google Scholar] [CrossRef]
- Xu, H.; George, E.; Kinose, Y.; Kim, H.; Shah, J.B.; Peake, J.D.; Ferman, B.; Medvedev, S.; Murtha, T.; Barger, C.J.; et al. CCNE1 copy number is a biomarker for response to combination WEE1-ATR inhibition in ovarian and endometrial cancer models. Cell Rep. Med. 2021, 2, 100394. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Li, Y.; Zhao, Q.; Fan, L.; Tan, B.; Gu, J. Downregulation of Annexin A11 (ANXA11) Inhibits Cell Proliferation, Invasion, and Migration via the AKT/GSK-3β Pathway in Gastric Cancer. J. Pharmacol. Exp. Ther. 2018, 24, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, I.; González-Medina, A.; Gaspa, L.; Hidalgo, E.; Ayté, J. The INO80 complex activates the transcription of S-phase genes in a cell cycle-regulated manner. FEBS J. 2018, 285, 3870–3881. [Google Scholar] [CrossRef]
- Gowans, G.J.; Schep, A.N.; Wong, K.M.; King, D.A.; Greenleaf, W.J.; Morrison, A.J. INO80 chromatin remodeling coordinates metabolic homeostasis with cell division. Cell Rep. 2018, 22, 611–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirt, S.E.; Adler, A.S.; Gebala, V.; Weimann, J.M.; Schaffer, B.E.; Saddic, L.A.; Viatour, P.; Vogel, H.; Chang, H.Y.; Meissner, A.; et al. G1 arrest and differentiation can occur independently of Rb family function. J. Cell Biol. 2010, 191, 809–825. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Utter, M.B.; Frias, M.A.; Foster, D.A. Cancer cells with defective RB and CDKN2A are resistant to the apoptotic effects of rapamycin. Cancer Lett. 2021, 522, 164–170. [Google Scholar] [CrossRef]
- Satoh, T.; Kaida, D. Upregulation of p27 cyclin-dependent kinase inhibitor and a C-terminus truncated form of p27 contributes to G1 phase arrest. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Fu, X.-D.; Zhou, Y.; Zhang, Y. Down-regulation of the cyclin E1 oncogene expression by microRNA-16-1 induces cell cycle arrest in human cancer cells. BMB Rep. 2009, 42, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, K.; DeGregori, J.; Nevins, J.R. Regulation of the cyclin E gene by transcription factor E2F1. Proc. Natl. Acad. Sci. USA 1995, 92, 12146–12150. [Google Scholar] [CrossRef] [Green Version]
- Murad, H.; Hawat, M.; Ekhtiar, A.; Aljapawe, A.; Abbas, A.; Darwish, H.; Sbenati, O.; Ghannam, A. Induction of G1-phase cell cycle arrest and apoptosis pathway in MDA-MB-231 human breast cancer cells by sulfated polysaccharide extracted from Laurencia papillosa. Cancer Cell Int. 2016, 16, 39. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, F.; Bai, C.; Yao, C.; Zhong, H.; Zou, C.; Chen, X. Sophoridine induces apoptosis and S phase arrest via ROS-dependent JNK and ERK activation in human pancreatic cancer cells. J. Exp. Clin. Cancer Res. 2017, 36, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and abundance estimation from RNA-Seq reveals thousands of new transcripts and switching among isoforms. Nat Biotechnol. 2011, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, A.; Schwartz, M.; et al. gplots: Various R Programming Tools for Plotting Data. The Comprehensive R Archive Network. 2016. Available online: http://CRAN.R-project.org/package=gplots (accessed on 15 September 2019).
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 genome project data processing subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [Green Version]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, S.; Lee, E.J.; Thang, N.X.; La, H.; Lee, H.; Park, C.; Han, D.W.; Uhm, S.J.; Song, H.; Do, J.T.; et al. INO80 Is Required for the Cell Cycle Control, Survival, and Differentiation of Mouse ESCs by Transcriptional Regulation. Int. J. Mol. Sci. 2022, 23, 15402. https://doi.org/10.3390/ijms232315402
Yoo S, Lee EJ, Thang NX, La H, Lee H, Park C, Han DW, Uhm SJ, Song H, Do JT, et al. INO80 Is Required for the Cell Cycle Control, Survival, and Differentiation of Mouse ESCs by Transcriptional Regulation. International Journal of Molecular Sciences. 2022; 23(23):15402. https://doi.org/10.3390/ijms232315402
Chicago/Turabian StyleYoo, Seonho, Eun Joo Lee, Nguyen Xuan Thang, Hyeonwoo La, Hyeonji Lee, Chanhyeok Park, Dong Wook Han, Sang Jun Uhm, Hyuk Song, Jeong Tae Do, and et al. 2022. "INO80 Is Required for the Cell Cycle Control, Survival, and Differentiation of Mouse ESCs by Transcriptional Regulation" International Journal of Molecular Sciences 23, no. 23: 15402. https://doi.org/10.3390/ijms232315402
APA StyleYoo, S., Lee, E. J., Thang, N. X., La, H., Lee, H., Park, C., Han, D. W., Uhm, S. J., Song, H., Do, J. T., Choi, Y., & Hong, K. (2022). INO80 Is Required for the Cell Cycle Control, Survival, and Differentiation of Mouse ESCs by Transcriptional Regulation. International Journal of Molecular Sciences, 23(23), 15402. https://doi.org/10.3390/ijms232315402