
Regulation of Apoptosis and Oxidative Stress by Oral Boswellia Serrata Gum Resin Extract in a Rat Model of Endometriosis
 ,
,  ,
,  , ,
, ,  ,
,  ,
,  , ,
, ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
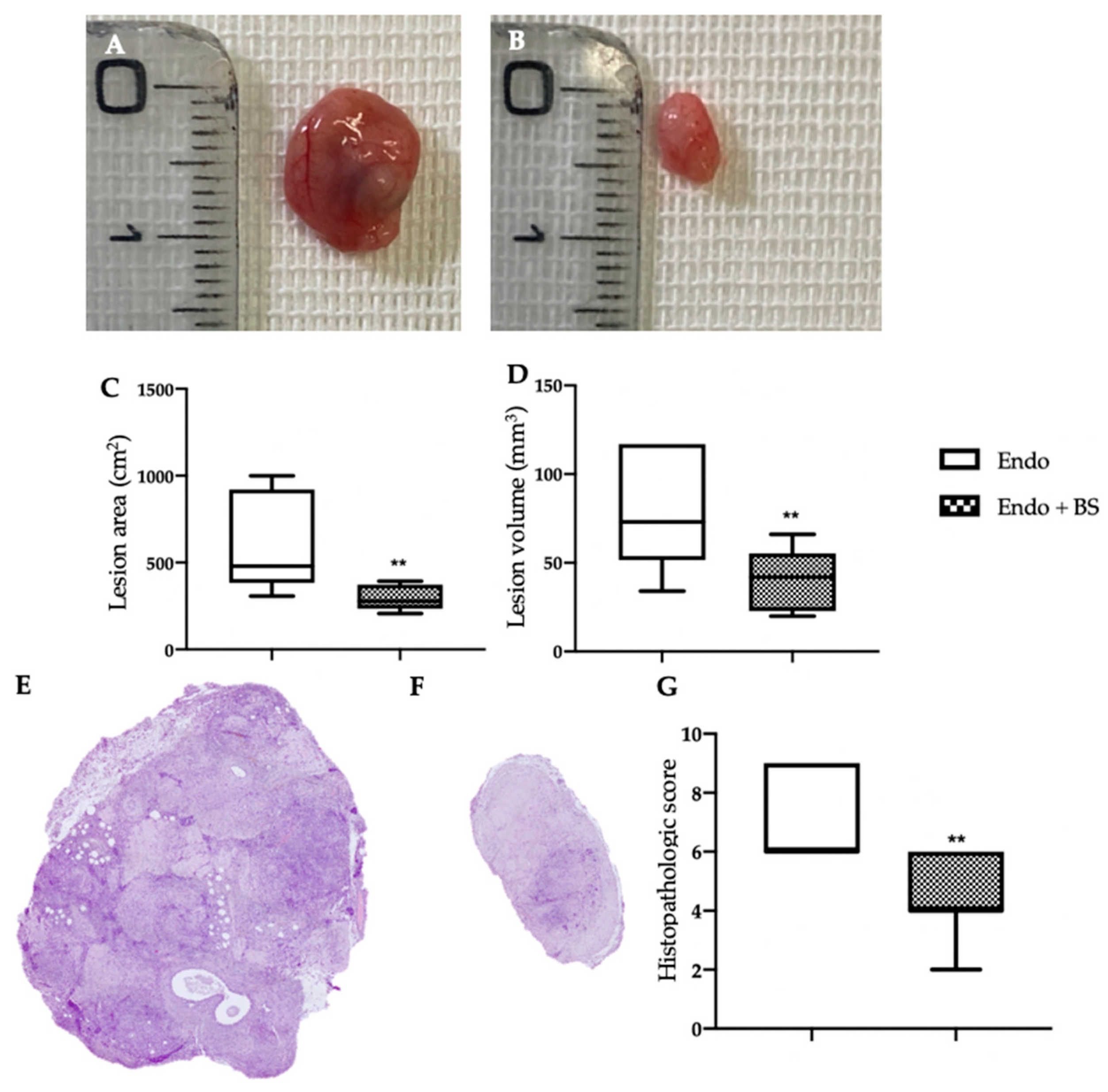
2.1. Monitoring of Endometriotic Lesions Development
2.2. Effect of Oral BS Administration on Macroscopic and Histological Analysis
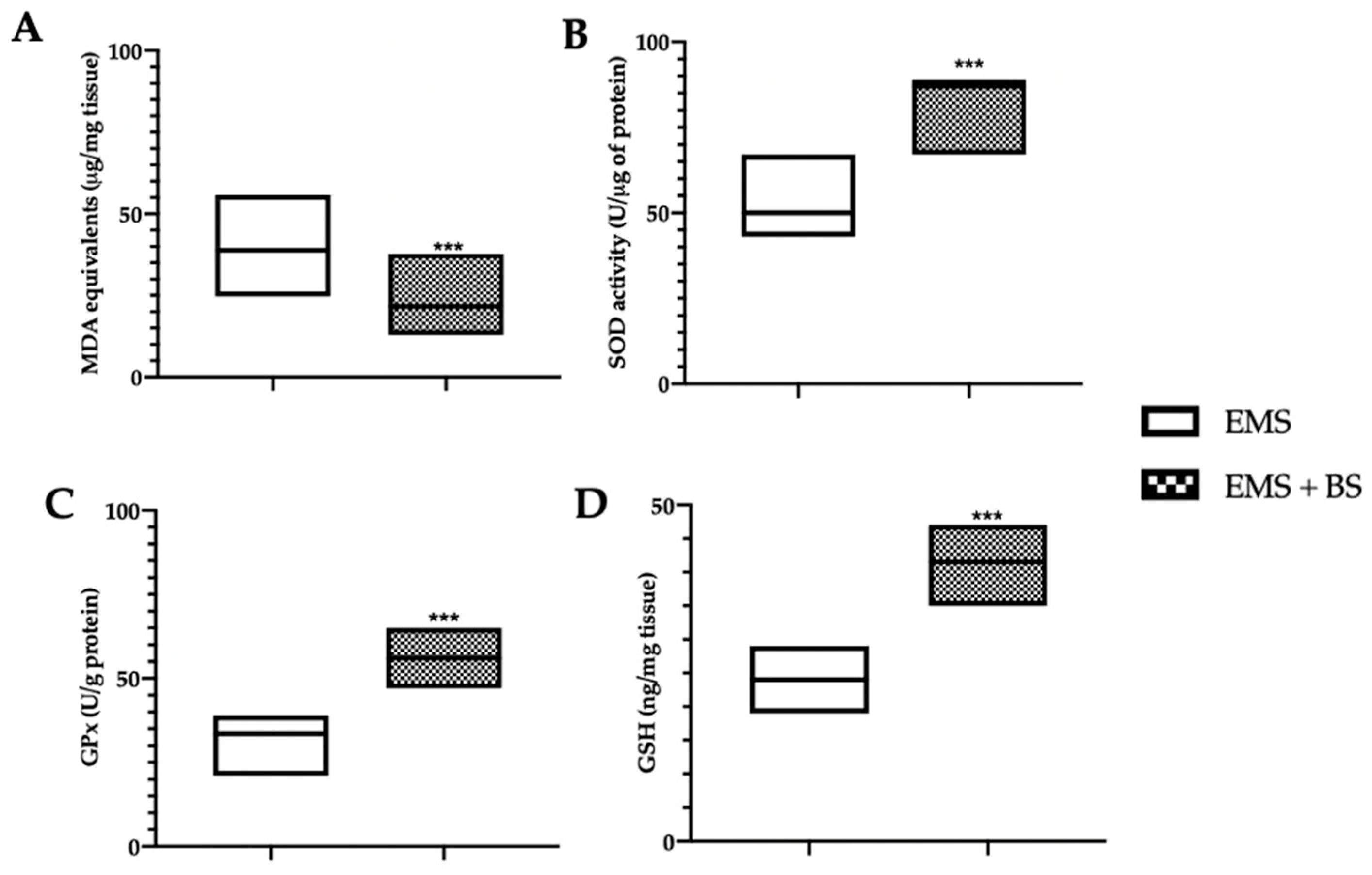
2.3. Effect of Oral BS Administration on the Oxidative Stress
2.4. Effect of Oral BS Administration on the Apoptosis Pathway
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Model Induction
4.3. Experimental Groups
- -
- EMS group: animals were subjected to the above described induction and vehicle (2% gum acacia was orally administered on the seventh day until the fourteenth day);
- -
- EMS + BS group: animals were subjected to the above described induction and BS (100 mg/Kg) was orally administered on the seventh day until the fourteenth day;
- -
- Sham group: animals were subjected to the above described induction but they were intraperitoneally injected with 500 μL of PBS along the midventral line instead of endometriotic tissue.
4.4. Abdominal High-Frequency Ultrasound
4.5. Biochemical Analysis
4.6. Histological Examination
4.7. Western Blot Analysis
4.8. Terminal Deoxynucleotidyl Nick-End Labeling (TUNEL) Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jouhari, S.; Mohammadzadeh, A.; Soltanghoraee, H.; Mohammadi, Z.; Khazali, S.; Mirzadegan, E.; Lakpour, N.; Fatemi, F.; Zafardoust, S.; Mohazzab, A.; et al. Effects of silymarin, cabergoline and letrozole on rat model of endometriosis. Taiwan J. Obstet. Gynecol. 2018, 57, 830–835. [Google Scholar] [CrossRef]
- Fritzer, N.; Tammaa, A.; Haas, D.; Oppelt, P.; Renner, S.; Hornung, D.; Wölfler, M.; Ulrich, U.; Hudelist, G. When sex is not on fire: A prospective multicentre study evaluating the short-term effects of radical resection of endometriosis on quality of sex life and dyspareunia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 197, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Van Langendonckt, A.; Casanas-Roux, F.; Donnez, J. Oxidative stress and peritoneal endometriosis. Fertil. Steril. 2002, 77, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Foyouzi, N.; Berkkanoglu, M.; Arici, A.; Kwintkiewicz, J.; Izquierdo, D.; Duleba, A.J. Effects of oxidants and antioxidants on proliferation of endometrial stromal cells. Fertil. Steril. 2004, 82, 1019–1022. [Google Scholar] [CrossRef]
- Chen, N.; Du, B.; Zhou, H.; Shen, F.; Li, J.; Xie, Z. Abnormal expression of Nrf2 may play an important role in the pathogenesis and development of adenomyosis. PLoS ONE 2017, 12, e0182773. [Google Scholar] [CrossRef] [Green Version]
- Ekici, E.I.; Guney, M.; Naziroglu, M. Protective effect of cabergoline on mitochondrial oxidative stress-induced apoptosis is mediated by modulations of TRPM2 in neutrophils of patients with endometriosis. J. Bioenerg. Biomembr. 2020, 52, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Tischner, D.; Woess, C.; Ottina, E.; Villunger, A. Bcl-2-regulated cell death signalling in the prevention of autoimmunity. Cell Death Dis. 2010, 1, e48. [Google Scholar] [CrossRef] [Green Version]
- Kulsoom, B.; Shamsi, T.S.; Afsar, N.A.; Memon, Z.; Ahmed, N.; Hasnain, S.N. Bax, Bcl-2, and Bax/Bcl-2 as prognostic markers in acute myeloid leukemia: Are we ready for Bcl-2-directed therapy? Cancer Manag. Res. 2018, 10, 403. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, D.M.; Davis, R.B.; Ettner, S.L.; Appel, S.; Wilkey, S.; Van Rompay, M.; Kessler, R.C. Trends in alternative medicine use in the United States, 1990-1997: Results of a follow-up national survey. JAMA 1998, 280, 1569–1575. [Google Scholar] [CrossRef] [Green Version]
- Bardaweel, S.K.; Shehadeh, M.; Suaifan, G.A.; Kilani, M.-V.Z. Complementary and alternative medicine utilization by a sample of infertile couples in Jordan for infertility treatment: Clinics-based survey. BMC Complement. Altern. Med. 2013, 13, 35. [Google Scholar] [CrossRef]
- Comar, K.M.; Kirby, D.F. Herbal remedies in gastroenterology. J. Clin. Gastroenterol. 2005, 39, 457–468. [Google Scholar] [CrossRef]
- Tindle, H.A.; Davis, R.B.; Phillips, R.S.; Eisenberg, D.M. Trends in use of complementary and alternative medicine by US adults: 1997-2002. Altern. Ther. Health Med. 2005, 11, 42. [Google Scholar]
- Cox, H.; Henderson, L.; Wood, R.; Cagliarini, G. Learning to take charge: Women’s experiences of living with endometriosis. Complement. Ther. Nurs. Midwifery 2003, 9, 62–68. [Google Scholar] [CrossRef]
- Fugh-Berman, A.; Kronenberg, F. Complementary and alternative medicine (CAM) in reproductive-age women: A review of randomized controlled trials. Reprod. Toxicol. 2003, 17, 137–152. [Google Scholar] [CrossRef]
- Wieser, F.; Cohen, M.; Gaeddert, A.; Yu, J.; Burks-Wicks, C.; Berga, S.L.; Taylor, R.N. Evolution of medical treatment for endometriosis: Back to the roots? Hum. Reprod. Update 2007, 13, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-J.; Nilsson, Å.; Oredsson, S.; Badmaev, V.; Zhao, W.-Z.; Duan, R.-D. Boswellic acids trigger apoptosis via a pathway dependent on caspase-8 activation but independent on Fas/Fas ligand interaction in colon cancer HT-29 cells. Carcinogenesis 2002, 23, 2087–2093. [Google Scholar] [CrossRef] [Green Version]
- Han, R. Highlight on the studies of anticancer drugs derived from plants in China. Stem Cells 1994, 12, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Thawani, V.; Hingorani, L.; Shrivastava, M.; Bhate, V.; Khiyani, R. Pharmacokinetic study of 11-keto β-Boswellic acid. Phytomedicine 2004, 11, 255–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, G.K.; Chandrakasan, G.; Dhar, S. Studies on the metabolism of glycosaminoglycans under the influence of new herbal anti-inflammatory agents. Biochem. Pharmacol. 1989, 38, 3527–3534. [Google Scholar] [CrossRef]
- Safayhi, H.; Mack, T.; Sabieraj, J.; Anazodo, M.I.; Subramanian, L.R.; Ammon, H. Boswellic acids: Novel, specific, nonredox inhibitors of 5-lipoxygenase. J. Pharmacol. Exp. Ther. 1992, 261, 1143–1146. [Google Scholar]
- Elshazly, S.M.; Abd El Motteleb, D.M.; Nassar, N.N. The selective 5-LOX inhibitor 11-keto-β-boswellic acid protects against myocardial ischemia reperfusion injury in rats: Involvement of redox and inflammatory cascades. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2013, 386, 823–833. [Google Scholar] [CrossRef]
- Hartmann, R.M.; Morgan Martins, M.I.; Tieppo, J.; Fillmann, H.S.; Marroni, N.P. Effect of Boswellia serrata on antioxidant status in an experimental model of colitis rats induced by acetic acid. Dig. Dis. Sci. 2012, 57, 2038–2044. [Google Scholar] [CrossRef] [PubMed]
- Ali, E.N.; Mansour, S.Z. Boswellic acids extract attenuates pulmonary fibrosis induced by bleomycin and oxidative stress from gamma irradiation in rats. Chin. Med. 2011, 6, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, Z.-T.; Gong, X.-J.; Sun, H.-B.; Li, Y.-M.; Ji, H. Protective effects of oleanolic acid on cerebral ischemic damage in vivo and H2O2-induced injury in vitro. Pharm. Biol. 2011, 49, 78–85. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, M.; Wang, M.; Wang, M.; Zhang, T.; Park, J.; Zhu, Y.; Guo, C.; Jia, Y.; Li, Y. Neuroprotection by acetyl-11-keto-β-boswellic acid, in ischemic brain injury involves the Nrf2/HO-1 defense pathway. Sci. Rep. 2014, 4, 7002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-J.; Nilsson, A.; Oredsson, S.; Badmaev, V.; Duan, R.-D. Keto-and acetyl-keto-boswellic acids inhibit proliferation and induce apoptosis in Hep G2 cells via a caspase-8 dependent pathway. Int. J. Mol. Med. 2002, 10, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Lee, J.H.; Harwalkar, J.A.; Bondar, J.; Safayhi, H.; Golubic, M. Acetyl-11-Keto-ß-Boswellic acid (Akba) is cytotoxic for meningioma cells and inhibits phosphorylation of the extracellular-signal regulated kinase 1 and 2. In Eicosanoids and Other Bioactive Lipids in Cancer, Inflammation, and Radiation Injury 5; Springer: Berlin/Heidelberg, Germany, 2002; pp. 387–393. [Google Scholar]
- Syrovets, T.; Büchele, B.; Krauss, C.; Laumonnier, Y.; Simmet, T. Acetyl-boswellic acids inhibit lipopolysaccharide-mediated TNF-α induction in monocytes by direct interaction with IκB kinases. J. Immunol. 2005, 174, 498–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winking, M.; Sarikaya, S.; Rahmanian, A.; Jödicke, A.; Böker, D.-K. Boswellic acids inhibit glioma growth: A new treatment option? J. Neuro-Oncol. 2000, 46, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Entschladen, F.; Liu, H.; Niggemann, B.; Fang, Q.; Zaenker, K.S.; Han, R. Boswellic acid acetate induces differentiation and apoptosis in highly metastatic melanoma and fibrosarcoma cells. Cancer Detect. Prev. 2003, 27, 67–75. [Google Scholar] [CrossRef]
- Harlev, A.; Gupta, S.; Agarwal, A. Targeting oxidative stress to treat endometriosis. Expert Opin. Ther. Targets 2015, 19, 1447–1464. [Google Scholar] [CrossRef]
- Rosa e Silva, J.C.; do Amara, V.F.; Mendonca, J.L.; Rosa e Silva, A.C.; Nakao, L.S.; Poli Neto, O.B.; Ferriani, R.A. Serum markers of oxidative stress and endometriosis. Clin. Exp. Obstet. Gynecol. 2014, 41, 371–374. [Google Scholar] [CrossRef]
- Ngo, C.; Chereau, C.; Nicco, C.; Weill, B.; Chapron, C.; Batteux, F. Reactive oxygen species controls endometriosis progression. Am. J. Pathol. 2009, 175, 225–234. [Google Scholar] [CrossRef]
- Di Paola, D.; Natale, S.; Iaria, C.; Crupi, R.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; Peritore, A.F. Environmental Co-Exposure to Potassium Perchlorate and Cd Caused Toxicity and Thyroid Endocrine Disruption in Zebrafish Embryos and Larvae (Danio rerio). Toxics 2022, 10, 198. [Google Scholar] [CrossRef]
- Alfieri, A.; Srivastava, S.; Siow, R.C.; Modo, M.; Fraser, P.A.; Mann, G.E. Targeting the Nrf2–Keap1 antioxidant defence pathway for neurovascular protection in stroke. J. Physiol. 2011, 589, 4125–4136. [Google Scholar] [CrossRef]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1-Nrf2-ARE Pathway As a Potential Preventive and Therapeutic Target: An Update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V.; Canning, P. Keap1, the cysteine-based mammalian intracellular sensor for electrophiles and oxidants. Arch. Biochem. Biophys. 2017, 617, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niture, S.K.; Jaiswal, A.K. Nrf2 protein up-regulates antiapoptotic protein Bcl-2 and prevents cellular apoptosis. J. Biol. Chem. 2012, 287, 9873–9886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Lim, W.; Bazer, F.W.; Song, G. Apigenin induces ROS-dependent apoptosis and ER stress in human endometriosis cells. J. Cell. Physiol. 2018, 233, 3055–3065. [Google Scholar] [CrossRef]
- McLaren, J.; Prentice, A.; Charnock-Jones, D.S.; Sharkey, A.M.; Smith, S.K. Immunolocalization of the apoptosis regulating proteins Bcl-2 and Bax in human endometrium and isolated peritoneal fluid macrophages in endometriosis. Hum. Reprod. 1997, 12, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Talebi, H.; Farahpour, M.R.; Hamishehkar, H. The effectiveness of Rutin for prevention of surgical induced endometriosis development in a rat model. Sci. Rep. 2021, 11, 7180. [Google Scholar] [CrossRef]
- Cho, Y.J.; Lee, S.H.; Park, J.W.; Han, M.; Park, M.J.; Han, S.J. Dysfunctional signaling underlying endometriosis: Current state of knowledge. J. Mol. Endocrinol. 2018, 60, R97–R113. [Google Scholar] [CrossRef] [Green Version]
- Cordaro, M.; Trovato Salinaro, A.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Interdonato, L.; Crea, R.; Fusco, R.; et al. Hidrox((R)) and Endometriosis: Biochemical Evaluation of Oxidative Stress and Pain. Antioxidants 2021, 10, 720. [Google Scholar] [CrossRef]
- Umar, S.; Umar, K.; Sarwar, A.H.M.G.; Khan, A.; Ahmad, N.; Ahmad, S.; Katiyar, C.K.; Husain, S.A.; Khan, H.A. Boswellia serrata extract attenuates inflammatory mediators and oxidative stress in collagen induced arthritis. Phytomedicine 2014, 21, 847–856. [Google Scholar] [CrossRef]
- Genovese, T.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Caudullo, S.; Raffone, E.; Macri, F.; Interdonato, L.; Gugliandolo, E.; Interlandi, C.; et al. Molecular and Biochemical Mechanism of Cannabidiol in the Management of the Inflammatory and Oxidative Processes Associated with Endometriosis. Int. J. Mol. Sci. 2022, 23, 5427. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G.; et al. Cashew (Anacardium occidentale L.) Nuts Counteract Oxidative Stress and Inflammation in an Acute Experimental Model of Carrageenan-Induced Paw Edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef]
- D’Amico, R.; Trovato Salinaro, A.; Fusco, R.; Cordaro, M.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Lo Dico, G.; Cuzzocrea, S.; Di Paola, R. Hericium erinaceus and coriolus versicolor modulate molecular and biochemical changes after traumatic brain injury. Antioxidants 2021, 10, 898. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; D’Amico, R.; Licata, P.; Crupi, R.; Gugliandolo, E. Effects of hydroxytyrosol against lipopolysaccharide-induced inflammation and oxidative stress in bovine mammary epithelial cells: A natural therapeutic tool for bovine mastitis. Antioxidants 2020, 9, 693. [Google Scholar] [CrossRef]
- Rashid, K.; Das, J.; Sil, P.C. Taurine ameliorate alloxan induced oxidative stress and intrinsic apoptotic pathway in the hepatic tissue of diabetic rats. Food Chem. Toxicol. 2013, 51, 317–329. [Google Scholar] [CrossRef]
- Cordaro, M.; D’Amico, R.; Morabito, R.; Fusco, R.; Siracusa, R.; Peritore, A.F.; Impellizzeri, D.; Genovese, T.; Crupi, R.; Gugliandolo, E. Physiological and Biochemical Changes in NRF2 Pathway in Aged Animals Subjected to Brain Injury. Cell. Physiol. Biochem. 2021, 55, 160–179. [Google Scholar] [PubMed]
- D’Amico, R.; Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Scuto, M.; Cuzzocrea, S.; Di Paola, R.; et al. Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. Int. J. Mol. Sci. 2020, 21, 2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, R.; Gugliandolo, E.; Campolo, M.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. Effect of a new formulation of micronized and ultramicronized N-palmitoylethanolamine in a tibia fracture mouse model of complex regional pain syndrome. PLoS ONE 2017, 12, e0178553. [Google Scholar] [CrossRef] [PubMed]
- Peritore, A.F.; Crupi, R.; Scuto, M.; Gugliandolo, E.; Siracusa, R.; Impellizzeri, D.; Cordaro, M.; D’Amico, R.; Fusco, R.; Di Paola, R.; et al. The Role of Annexin A1 and Formyl Peptide Receptor 2/3 Signaling in Chronic Corticosterone-Induced Depression-Like behaviors and Impairment in Hippocampal-Dependent Memory. CNS Neurol. Disord. Drug Targets 2020, 19, 27–43. [Google Scholar] [CrossRef]
- Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Cordaro, M.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Salinaro, A.T.; Raffone, E.; Genovese, T. Autophagy and mitophagy promotion in a rat model of endometriosis. Int. J. Mol. Sci. 2021, 22, 5074. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Salinaro, A.T.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Crea, R.; Cordaro, M.; Cuzzocrea, S. Hidrox® counteracts cyclophosphamide-induced male infertility through NRF2 pathways in a mouse model. Antioxidants 2021, 10, 778. [Google Scholar] [CrossRef]
- Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; D’Amico, R.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. Consumption of Anacardium occidentale L. (Cashew Nuts) Inhibits Oxidative Stress through Modulation of the Nrf2/HO-1 and NF-kB Pathways. Molecules 2020, 25, 4426. [Google Scholar] [CrossRef]
- Peritore, A.F.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Fusco, R.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Di Paola, R.; Cuzzocrea, S.; et al. Management of Acute Lung Injury: Palmitoylethanolamide as a New Approach. Int. J. Mol. Sci. 2021, 22, 5533. [Google Scholar] [CrossRef]
- Cordaro, M.; Fusco, R.; D’Amico, R.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Mandalari, G.; Cuzzocrea, S.; et al. Cashew (Anacardium occidentale L.) Nuts Modulate the Nrf2 and NLRP3 Pathways in Pancreas and Lung after Induction of Acute Pancreatitis by Cerulein. Antioxidants 2020, 9, 992. [Google Scholar] [CrossRef]
- Cordaro, M.; Scuto, M.; Siracusa, R.; D’Amico, R.; Filippo Peritore, A.; Gugliandolo, E.; Fusco, R.; Crupi, R.; Impellizzeri, D.; Pozzebon, M.; et al. Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J. 2020, 34, 4085–4106. [Google Scholar] [CrossRef]
- Crupi, R.; Palma, E.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; Cordaro, M.; Impellizzeri, D.; De Caro, C.; Calzetta, L.; Cuzzocrea, S.; et al. Protective Effect of Hydroxytyrosol Against Oxidative Stress Induced by the Ochratoxin in Kidney Cells: In vitro and in vivo Study. Front. Vet. Sci. 2020, 7, 136. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, R.; Cordaro, M.; Fusco, R.; Peritore, A.F.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Mandalari, G.; Caccamo, D.; Cuzzocrea, S.; et al. Consumption of Cashew (Anacardium occidentale L.) Nuts Counteracts Oxidative Stress and Tissue Inflammation in Mild Hyperhomocysteinemia in Rats. Nutrients 2022, 14, 1474. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Xiang, W.; Zhang, T.; Zeng, L.; Yang, K.; Li, J. Effectiveness of Boswellia and Boswellia extract for osteoarthritis patients: A systematic review and meta-analysis. BMC Complement. Med. Ther. 2020, 20, 225. [Google Scholar] [CrossRef] [PubMed]
- Suchita, W.; Raman, D.M.R.; Kaur, C.D. A Review on phytochemistry and pharmacological activities of Boswellia serrata: A natural remedy. Int. J. Pharmacogn. 2021, 8, 454–461. [Google Scholar]
- Aronson, J.K. Meyler’s Side Effects of Herbal Medicines; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Interdonato, L.; Crupi, R.; Gugliandolo, E.; Macrì, F.; Di Paola, D.; Peritore, A.F.; et al. Regulation of Apoptosis and Oxidative Stress by Oral Boswellia Serrata Gum Resin Extract in a Rat Model of Endometriosis. Int. J. Mol. Sci. 2022, 23, 15348. https://doi.org/10.3390/ijms232315348
D’Amico R, Impellizzeri D, Cordaro M, Siracusa R, Interdonato L, Crupi R, Gugliandolo E, Macrì F, Di Paola D, Peritore AF, et al. Regulation of Apoptosis and Oxidative Stress by Oral Boswellia Serrata Gum Resin Extract in a Rat Model of Endometriosis. International Journal of Molecular Sciences. 2022; 23(23):15348. https://doi.org/10.3390/ijms232315348
Chicago/Turabian StyleD’Amico, Ramona, Daniela Impellizzeri, Marika Cordaro, Rosalba Siracusa, Livia Interdonato, Rosalia Crupi, Enrico Gugliandolo, Francesco Macrì, Davide Di Paola, Alessio Filippo Peritore, and et al. 2022. "Regulation of Apoptosis and Oxidative Stress by Oral Boswellia Serrata Gum Resin Extract in a Rat Model of Endometriosis" International Journal of Molecular Sciences 23, no. 23: 15348. https://doi.org/10.3390/ijms232315348
APA StyleD’Amico, R., Impellizzeri, D., Cordaro, M., Siracusa, R., Interdonato, L., Crupi, R., Gugliandolo, E., Macrì, F., Di Paola, D., Peritore, A. F., Fusco, R., Cuzzocrea, S., & Di Paola, R. (2022). Regulation of Apoptosis and Oxidative Stress by Oral Boswellia Serrata Gum Resin Extract in a Rat Model of Endometriosis. International Journal of Molecular Sciences, 23(23), 15348. https://doi.org/10.3390/ijms232315348