

New TMA (4,6,4′-Trimethyl angelicin) Analogues as Anti-Inflammatory Agents in the Treatment of Cystic Fibrosis Lung Disease

 ,
,  ,
,  , , ,
, , , 

Abstract
1. Introduction
2. Results and Discussion
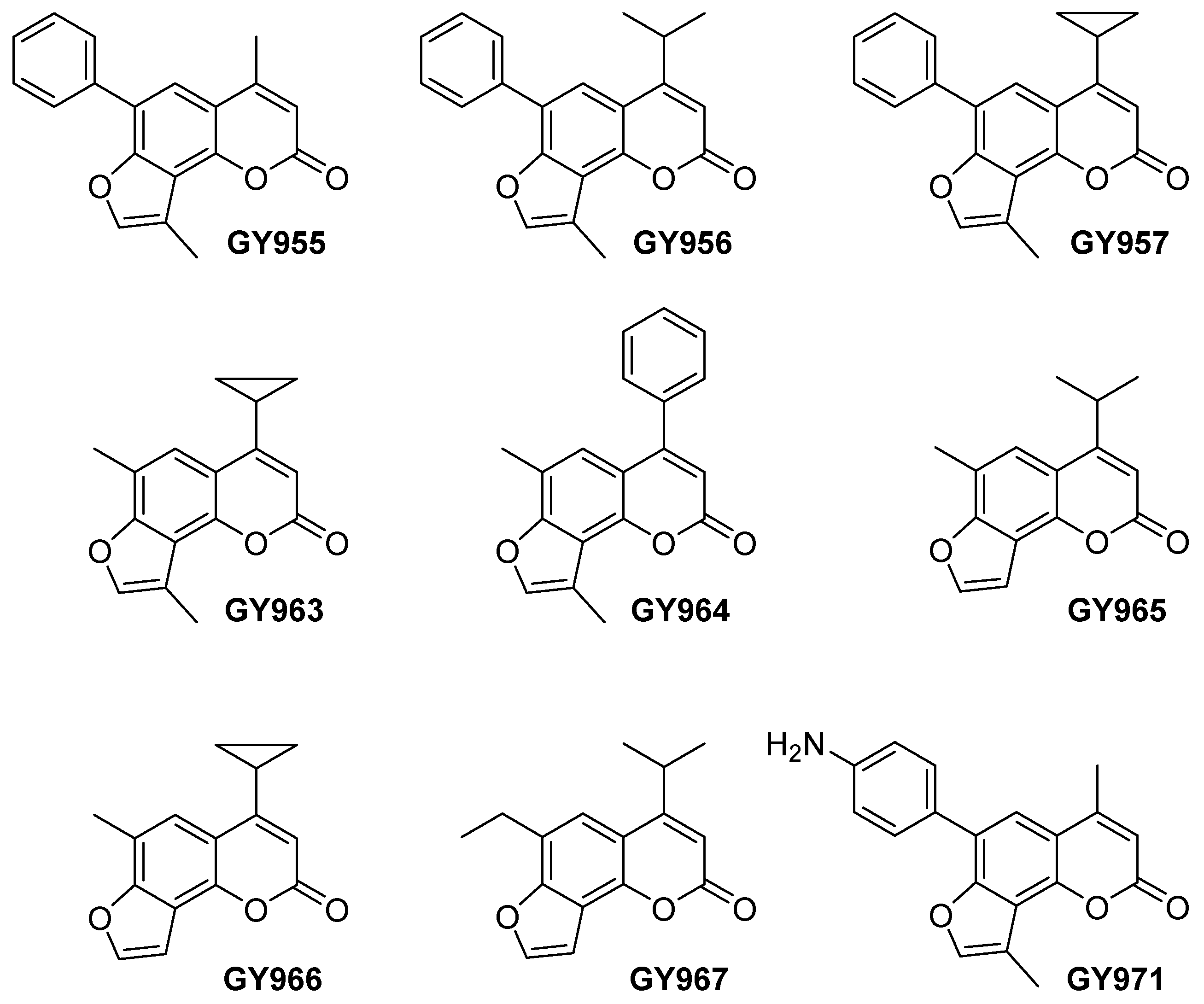
2.1. Synthesis of TMA Analogues
2.2. In Vitro Experiments
2.2.1. NF-κB Inhibition Activity of TMA Analogues (EMSA)
2.2.2. NF-κB-Mediated IL-8 Expression and Release after Treatment with TMA Analogues
Effect of TMA Analogues on IL-8 Expression in IB3-1 F508del/W1282X Cells Infected with P. aeruginosa
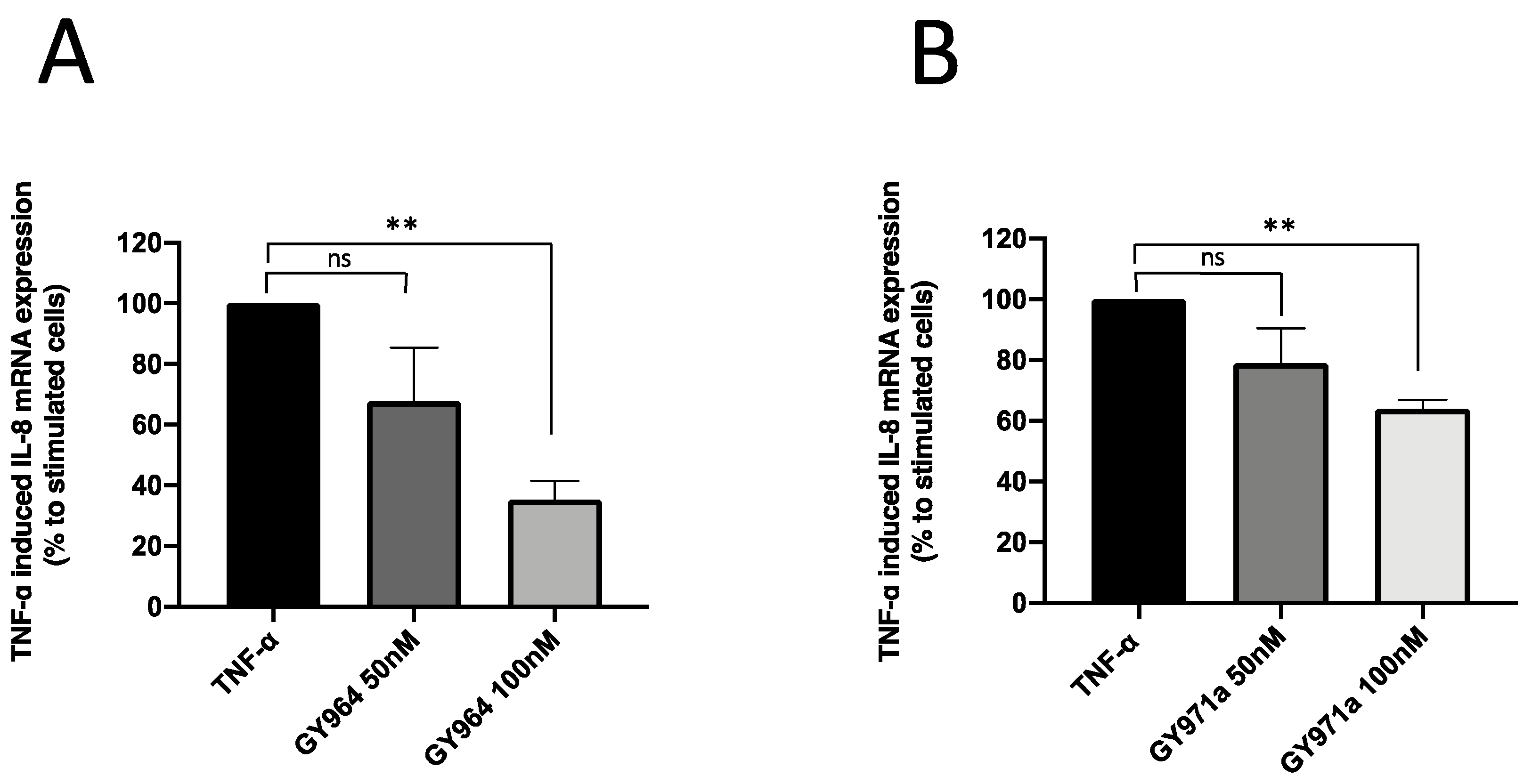
Effect of TMA Analogues on IL-8 Expression in TNF-α-Stimulated IB3-1 508Fdel/W1282X Cells
IL-8 Release in IB3-1 Cells Stimulated with TNF-α or Infected with P. aeruginosa
2.2.3. Toxicity Studies (Antiproliferative and Apoptotic Activity Evaluation)
2.3. In Vivo Experiments
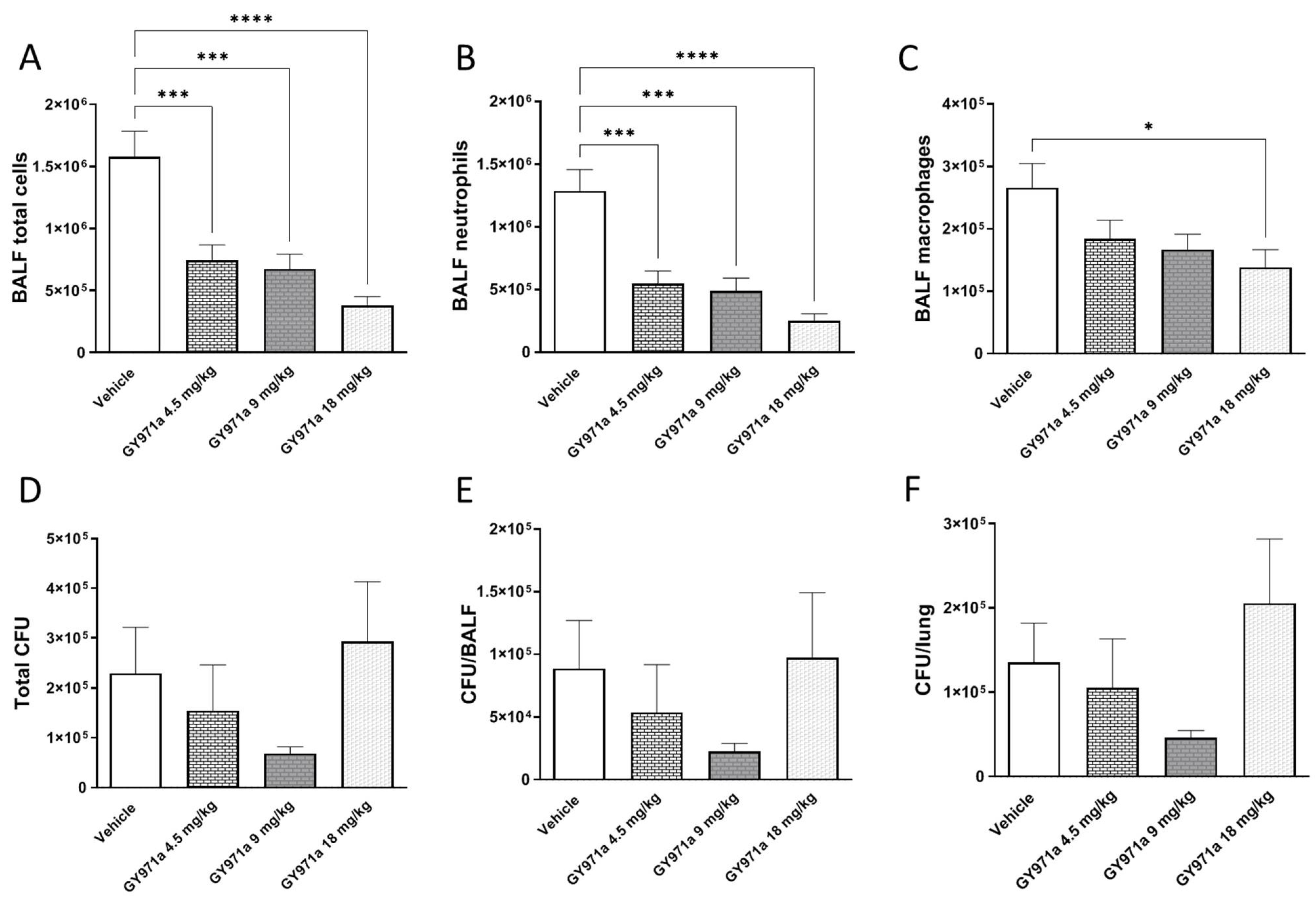
In Vivo Toxicity and Efficacy of TMA Analogue GY971a (pANDMA) in a Murine Model of P. aeruginosa Acute Infection
3. Materials and Methods
3.1. Synthesis of TMA Derivatives
3.2. In Vitro Assays
3.2.1. EMSA Experiments
3.2.2. Gene Expression Analysis Induced by TMA Analogues on PAO1-Infected or TNF-alpha-Stimulated IB3-1 Cells
- Cell culture conditions, TMA analogue treatment, infection with Pseudomonas aeruginosa or addition of TNF-alpha stimulus:
- Gene expression analysis:
- Statistical analysis
3.2.3. Protein Expression Analysis Induced by TMA Analogues (ELISA)
3.2.4. Toxicity
- Cell Proliferation Assay
- Proapoptotic activity (Annexin V method)
- Antimicrobial activity (Broth Dilution Method)
- Statistical analysis
3.3. In Vivo Assays
- Ethics Statement
3.3.1. Toxicity
3.3.2. Mouse Models of Acute P. aeruginosa Infection
- Histopathological evaluation
- Statistical analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Riordan, J.R.; Rommens, J.M.; Kerem, B.S.; Alon, N.O.A.; Rozmahel, R.; Grzelczak, Z.; Zielenski, J.; Lok, S.I.; Plavsic, N.; Chou, J.L.; et al. Identification of the Cystic Fibrosis Gene: Cloning and Characterization of Complementary DNA. Science 1989, 245, 1066–1073. [Google Scholar] [CrossRef]
- Rommens, J.M.; Ianuzzi, M.C.; Kerem, B.-S.; Drumm, M.L.; Melmer, G.; Dean, M.; Rozmahel, R.; Cole, J.L.; Kennedy, D.; Ilidaka, N.; et al. Identification of the cystic fibrosis gene: Chromosome walking and jumping. Science 1989, 245, 1058–1065. [Google Scholar] [CrossRef]
- Kerem, B.S.; Rommens, J.M.; Buchanan, J.A.; Markiewicz, D.; Cox, T.K.; Chakravarti, A.; Buchwald, M.; Tsui, L.C. Identification of the Cystic Fibrosis Gene: Genetic Analysis. Science 1989, 245, 1073–1080. [Google Scholar] [CrossRef]
- Khan, T.Z.; Wagener, J.S.; Bost, T.; Martinez, J.; Accurso, F.J.; Riches, D.W. Early pulmonary inflammation in infants with cystic fibrosis. Am. J. Respir. Crit. Care Med. 1995, 151, 1075–1082. [Google Scholar]
- Noah, T.L.; Black, H.R.; Cheng, P.W.; Wood, R.E.; Leigh, M.W. Nasal and Bronchoalveolar Lavage Fluid Cytokines in Early Cystic Fibrosis. J. Infect. Dis. 1997, 175, 638–647. [Google Scholar] [CrossRef]
- Armstrong, D.S.; Grimwood, K.; Carlin, J.B.; Carzino, R.; Gutièrrez, J.P.; Hull, J.; Olinsky, A.; Phelan, E.M.; Robertson, C.F.; Phelan, P.D. Lower Airway Inflammation in Infants and Young Children with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 1997, 156, 1197–1204. [Google Scholar] [CrossRef]
- Stoltz, D.A.; Meyerholz, D.K.; Welsh, M.J. Origins of Cystic Fibrosis Lung Disease David. N. Engl. J. Med. 2015, 256, 351–362. [Google Scholar] [CrossRef]
- Boucher, R.C. Evidence for Airway Surface Dehydration as the Initiating Event in CF Airway Disease. J. Intern. Med. 2007, 261, 5–16. [Google Scholar] [CrossRef]
- Ribeiro, C.M.P.; McElvaney, N.G.; Cabrini, G. Editorial: Novel Anti-Inflammatory Approaches for Cystic Fibrosis Lung Disease: Identification of Molecular Targets and Design of Innovative Therapies. Front. Pharmacol. 2021, 12, 10–13. [Google Scholar] [CrossRef]
- Cabrini, G.; Rimessi, A.; Borgatti, M.; Lampronti, I.; Finotti, A.; Pinton, P.; Gambari, R. Role of Cystic Fibrosis Bronchial Epithelium in Neutrophil Chemotaxis. Front. Immunol. 2020, 11, 1438. [Google Scholar] [CrossRef]
- Hartl, D.; Latzin, P.; Hordijk, P.; Marcos, V.; Rudolph, C.; Woischnik, M.; Krauss-Etschmann, S.; Koller, B.; Reinhardt, D.; Roscher, A.A.; et al. Cleavage of CXCR1 on Neutrophils Disables Bacterial Killing in Cystic Fibrosis Lung Disease. Nat. Med. 2007, 13, 1423–1430. [Google Scholar] [CrossRef]
- Galli, F.; Battistoni, A.; Gambari, R.; Pompella, A.; Bragonzi, A.; Pilolli, F.; Iuliano, L.; Piroddi, M.; Dechecchi, M.C.; Cabrini, G. Oxidative Stress and Antioxidant Therapy in Cystic Fibrosis. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 690–713. [Google Scholar] [CrossRef]
- Marcos, V.; Zhou-Suckow, Z.; Önder Yildirim, A.; Bohla, A.; Hector, A.; Vitkov, L.; Krautgartner, W.D.; Stoiber, W.; Griese, M.; Eickelberg, O.; et al. Free DNA in Cystic Fibrosis Airway Fluids Correlates with Airflow Obstruction. Mediat. Inflamm. 2015, 2015, 408935. [Google Scholar] [CrossRef]
- McElvaney, O.J.; McElvaney, N.G. Targeting IL-8 in cystic fibrosis: Enough but not too much. Am. J. Respir. Cell. Mol. Biol. 2018, 59, 401–402. [Google Scholar] [CrossRef]
- Laselva, O.; Criscione, M.L.; Allegretta, C.; Di Gioia, S.; Liso, A.; Conese, M. Like Growth Factor Binding Protein (IGFBP-6) as a Novel Regulator of Inflammatory Response in Cystic Fibrosis Airway Cells. Front. Mol. Biosci. 2022, 12, 905468. [Google Scholar] [CrossRef]
- Tamanini, A.; Borgatti, M.; Finotti, A.; Piccagli, L.; Bezzerri, V.; Favia, M.; Guerra, L.; Lampronti, I.; Bianchi, N.; Dall’Acqua, F.; et al. Trimethylangelicin Reduces IL-8 Transcription and Potentiates CFTR Function. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, 380–390. [Google Scholar] [CrossRef]
- Lampronti, I.; Manzione, M.G.; Sacchetti, G.; Ferrari, D.; Spisani, S.; Bezzerri, V.; Finotti, A.; Borgatti, M.; Dechecchi, M.C.; Miolo, G.; et al. Differential Effects of Angelicin Analogues on NF-ΚB Activity and IL-8 Gene Expression in Cystic Fibrosis IB3-1 Cells. Mediat. Inflamm. 2017, 2017, 2389487. [Google Scholar] [CrossRef]
- Marzaro, G.; Lampronti, I.; D’Aversa, E.; Sacchetti, G.; Miolo, G.; Vaccarin, C.; Cabrini, G.; Dechecchi, M.C.; Gambari, R.; Chilin, A. Design, Synthesis and Biological Evaluation of Novel Trimethylangelicin Analogues Targeting Nuclear Factor kB (NF-kB). Eur. J. Med. Chem. 2018, 151, 285–293. [Google Scholar] [CrossRef]
- Vaccarin, C.; Gabbia, D.; De Martin, S.; Franceschinis, E.; Roverso, M.; Bogialli, S.; Sacchetti, G.; Tupini, C.; Lampronti, I.; Gambari, G.; et al. Improved Trimethylangelicin Analogues for Cystic Fibrosis: Design, synthesis and preliminary screening. Int. J. Mol. Sci. 2022, 23, 11528. [Google Scholar] [CrossRef]
- Bragonzi, A. Murine Models of Acute and Chronic Lung Infection with Cystic Fibrosis Pathogens. Int. J. Med. Microbiol. 2010, 300, 584–593. [Google Scholar] [CrossRef]
- Facchini, M.; De Fino, I.; Riva, C.; Bragonzi, A. Long term chronic Pseudomonas aeruginosa airway infection in mice. J. Vis. Exp. 2014, 85, 51019. [Google Scholar] [CrossRef] [PubMed]
- Montagner, G.; Bezzerri, V.; Cabrini, G.; Fabbri, E.; Borgatti, M.; Lampronti, I.; Finotti, A.; Nielsen, P.E.; Gambari, R. An antisense peptide nucleic acid against Pseudomonas aeruginosa inhibiting bacterial-induced inflammatory responses in the cystic fibrosis IB3-1 cellular model system. Int. J. Biol. Macromol. 2017, 99, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Milani, R.; Marcellini, A.; Montagner, G.; Baldisserotto, A.; Manfredini, S.; Gambari, R.; Lampronti, I. Phloridzin derivatives inhibiting pro-inflammatory cytokine expression in human cystic fibrosis IB3-1 cells. Eur. J. Pharm. Sci. 2015, 78, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, E.; Borgatti, M.; Montagner, G.; Bianchi, N.; Finotti, A.; Lampronti, I.; Bezzerri, V.; Dechecchi, M.C.; Cabrini, G.; Gambari, R. Expression of MicroRNA-93 and Interleukin-8 during Pseudomonas Aeruginosa-Mediated Induction of Proinflammatory Responses. Am. J. Respir. Cell Mol. Biol. 2014, 50, 1144–1155. [Google Scholar] [CrossRef]
- Scattolin, T.; Caligiuri, I.; Canovese, L.; Demitri, N.; Gambari, R.; Lampronti, I.; Rizzolio, F.; Santo, C.; Visentin, F. Synthesis of new allyl palladium complexes bearing purine-based NHC ligands with antiproliferative and proapoptotic activities on human ovarian cancer cell lines. Dalton Trans. 2018, 47, 13616–13630. [Google Scholar] [CrossRef]
- Lampronti, I.; Dechecchi, M.C.; Rimessi, A.; Bezzerri, V.; Nicolis, E.; Guerrini, A.; Tacchini, M.; Tamanini, A.; Munari, S.; D’Aversa, E.; et al. β-Sitosterol Reduces the Expression of Chemotactic Cytokine Genes in Cystic Fibrosis Bronchial Epithelial Cells. Front. Pharmacol. 2017, 8, 236. [Google Scholar] [CrossRef]
- Cigana, C.; Ranucci, S.; Rossi, A.; De Fino, I.; Melessike, M.; Bragonzi, A. Antibiotic Efficacy Varies Based on the Infection Model and Treatment Regimen for Pseudomonas Aeruginosa. Eur. Respir. J. 2020, 55, 1802456. [Google Scholar] [CrossRef]
- Kukavica-Ibrulj, I.; Facchini, M.; Cigana, C.; Levesque, R.C.; Bragonzi, A. Assessing Pseudomonas aeruginosa virulence and the host response using murine models of acute and chronic lung infection. Methods Mol. Biol. 2014, 1149, 757–771. [Google Scholar]
- Döring, G.; Bragonzi, A.; Paroni, M.; Aktürk, F.-F.; Cigana, C.; Schmidt, A.; Gilpin, D.; Heyder, S.; Born, T.; Smaczny, C.; et al. BIIL 284 reduces neutrophil numbers but increases P. aeruginosa bacteremia and inflammation in mouse lungs. J. Cyst. Fibros. 2014, 13, 156–163. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Derivative | IUPAC Name (ID Name) | MIC 1 |
|---|---|---|
| GY955 | 6-phenyl-4,4′-dimethyl-angelicin (6-PhDMA) | 30 μM |
| GY956 | 4-isopropyl-6-phenyl-4′-methyl-angelicin (IPPhMA) | 60 μM |
| GY957 | 4-cyclopropyl-6 phenyl-4′-methyl-angelicin (CPPhMA) | >100 μM |
| GY963 | 4-cyclopropyl-6,4′-dimethyl-angelicin (CPDMA) | 100 μM |
| GY964 | 4-phenyl-6,4′-dimethyl-angelicin (4-PhDMA) | 30 μM |
| GY965 | 4-isopropyl-6-methyl-angelicin (IPMA) | 80 μM |
| GY966 | 4-cyclopropyl-6-methyl-angelicin (CPMA) | 18 μM |
| GY967 | 4-isopropyl-6-ethyil-angelicin (IPEA) | >100 μM |
| GY971 | 6-p-aminophenyl-4,4′-dimethyl-angelicin (pANDMA) | <12.5 μM |
| Sample | Live | Early Apoptosis | Late Apoptosis | Dead | Total Apoptosis |
|---|---|---|---|---|---|
| Untreated cells (C-) | 97.85 | 0.72 | 1.02 | 0.41 | 1.74 |
| Vehicle | 97.01 | 1.06 | 1.42 | 0.51 | 2.48 |
| TMA 50 nM | 96.20 | 0.65 | 1.30 | 1.85 | 1.95 |
| TMA 100 nM | 95.95 | 0.50 | 1.50 | 2.05 | 2.00 |
| TMA 200 nM | 97.57 | 0.56 | 1.01 | 0.86 | 1.57 |
| GY964 50 nM | 98.05 | 0.75 | 0.95 | 0.25 | 1.70 |
| GY964 100 nM | 97.55 | 0.50 | 1.20 | 0.75 | 1.70 |
| GY964 200 nM | 97.02 | 1.21 | 1.26 | 0.51 | 2.48 |
| GY971a 50 nM | 96.80 | 0.75 | 1.55 | 0.90 | 2.30 |
| GY971a 100 nM | 97.70 | 0.65 | 1.25 | 0.40 | 1.90 |
| GY971a 200 nM | 96.99 | 0.97 | 1.63 | 0.41 | 2.60 |
| Sample | Live | Early Apoptosis | Late Apoptosis | Dead | Total Apoptosis |
|---|---|---|---|---|---|
| Untreated cells (C-) | 93.25 | 0.65 | 4.00 | 2.10 | 4.65 |
| Vehicle | 95.64 | 0.53 | 2.86 | 0.98 | 3.38 |
| TMA 50 nM | 92.69 | 0.59 | 5.07 | 1.65 | 5.67 |
| TMA 100 nM | 93.40 | 0.70 | 5.00 | 0.90 | 5.70 |
| TMA 200 nM | 93.50 | 1.16 | 3.75 | 1.60 | 4.91 |
| GY964 50 nM | 94.80 | 0.50 | 3.25 | 1.45 | 3.75 |
| GY964 100 nM | 91.82 | 3.45 | 4.09 | 0.63 | 7.54 |
| GY964 200 nM | 93.85 | 0.80 | 3.65 | 1.70 | 4.45 |
| GY971a 50 nM | 94.20 | 0.95 | 3.20 | 1.65 | 4.15 |
| GY971a 100 nM | 94.75 | 0.70 | 3.25 | 1.30 | 3.95 |
| GY971a 200 nM | 92.95 | 0.45 | 5.00 | 1.60 | 5.45 |
| Cytokine/Chemokine | Concentration pg × 700 µg−1 | |||
|---|---|---|---|---|
| Vehicle (H2O/DMSO 4%) | 4.5 mg/Kg GY971a | 9 mg/kg GY971a | 18 mg/kg GY971a | |
| IL-1α | 88.45 ± 10.30 | 65.95 ± 7.91 | 54.31 ± 7.29 * | 43.51 ± 3.03 ** |
| IL-1β | 9.21 ± 1.49 | 6.95 ± 1.05 | 4.91 ± 0.59 * | 4.89 ± 0.75 * |
| IL-2 | 6.27 ± 0.28 | 5.58 ± 0.30 | 5.76 ± 0.24 | 5.28 ± 0.20 * |
| IL-3 | 5.11 ± 0.40 | 4.58 ± 0.24 | 4.36 ± 0.41 | 3.67 ± 0.25 * |
| IL-4 | 1.29 ± 0.09 | 1.23 ± 0.12 | 0.93 ± 0.14 | 0.87 ± 0.11 * |
| IL-5 | 8.46 ± 0.68 | 8.13 ± 0.37 | 7.89 ± 0.63 | 7.26 ± 0.44 |
| IL-6 | 110.0 ± 27.68 | 64.30 ± 10.86 | 49.64 ± 9.34 * | 69.79 ± 8.27 |
| IL-9 | 38.58 ± 1.01 | 35.43 ± 1.74 | 32.17 ± 2.04 * | 29.64 ± 0.69 *** |
| IL-10 | 20.76 ± 0.95 | 17.55 ± 1.23 | 16.41 ± 1.56 * | 14.79 ± 1.05 ** |
| IL-12p40 | 71.29 ± 4.32 | 61.72 ± 5.31 | 52.88 ± 4.67 * | 45.67 ± 2.62 *** |
| IL-12p70 | 167.6 ± 16.08 | 123.7 ± 12.91 | 110.1 ± 13.58 ** | 94.45 ± 6.61 ** |
| IL-13 | 88.02 ± 3.81 | 82.53 ± 2.63 | 80.58 ± 4.30 | 78.14 ± 3.36 |
| IL-17A | 5.49 ± 0.63 | 4.27 ± 0.26 | 3.98 ± 0.30 * | 3.84 ± 0.17 * |
| Eotaxin | 1627 ± 160.8 | 1638 ± 124.4 | 1618 ± 87.33 | 1414 ± 65.28 |
| G-CSF | 444.1 ± 86.41 | 259.1 ± 36.89 | 297.8 ± 58.72 | 181.7 ± 19.88 * |
| GM-CSF | 48.66 ± 5.94 | 45.70 ± 1.52 | 33.60 ± 4.60 * | 42.55 ± 3.18 |
| IFN-γ | 30.10 ± 1.43 | 27.02 ± 1.51 | 23.76 ± 1.65 ** | 22.51 ± 0.96 ** |
| KC | 2413 ± 456.1 | 2085 ± 263.6 | 1486 ± 150.8 | 2048 ± 248.3 |
| MCP-1 | 1813 ± 322.6 | 1171 ± 136.4 | 979.0 ± 144.3 * | 692.2 ± 78.41 ** |
| MIP-1α | 186.9 ± 36.52 | 131.7 ± 23.94 | 107.0 ± 12.10 | 92.11 ± 10.33 * |
| MIP-1β | 291.2 ± 30.26 | 254.0 ± 28.16 | 219.8 ± 20.41 | 212.2 ± 15.66 |
| RANTES | 110.3 ± 10.35 | 123.0 ± 9.93 | 109.9 ± 9.72 | 101.9 ± 9.40 |
| TNF-α | 104.6 ± 9.05 | 93.45 ± 6.93 | 95.61 ± 11.04 | 84.06 ± 5.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tupini, C.; Chilin, A.; Rossi, A.; De Fino, I.; Bragonzi, A.; D’Aversa, E.; Cosenza, L.C.; Vaccarin, C.; Sacchetti, G.; Borgatti, M.; et al. New TMA (4,6,4′-Trimethyl angelicin) Analogues as Anti-Inflammatory Agents in the Treatment of Cystic Fibrosis Lung Disease. Int. J. Mol. Sci. 2022, 23, 14483. https://doi.org/10.3390/ijms232214483
Tupini C, Chilin A, Rossi A, De Fino I, Bragonzi A, D’Aversa E, Cosenza LC, Vaccarin C, Sacchetti G, Borgatti M, et al. New TMA (4,6,4′-Trimethyl angelicin) Analogues as Anti-Inflammatory Agents in the Treatment of Cystic Fibrosis Lung Disease. International Journal of Molecular Sciences. 2022; 23(22):14483. https://doi.org/10.3390/ijms232214483
Chicago/Turabian StyleTupini, Chiara, Adriana Chilin, Alice Rossi, Ida De Fino, Alessandra Bragonzi, Elisabetta D’Aversa, Lucia Carmela Cosenza, Christian Vaccarin, Gianni Sacchetti, Monica Borgatti, and et al. 2022. "New TMA (4,6,4′-Trimethyl angelicin) Analogues as Anti-Inflammatory Agents in the Treatment of Cystic Fibrosis Lung Disease" International Journal of Molecular Sciences 23, no. 22: 14483. https://doi.org/10.3390/ijms232214483
APA StyleTupini, C., Chilin, A., Rossi, A., De Fino, I., Bragonzi, A., D’Aversa, E., Cosenza, L. C., Vaccarin, C., Sacchetti, G., Borgatti, M., Tamanini, A., Dechecchi, M. C., Sanvito, F., Gambari, R., Cabrini, G., & Lampronti, I. (2022). New TMA (4,6,4′-Trimethyl angelicin) Analogues as Anti-Inflammatory Agents in the Treatment of Cystic Fibrosis Lung Disease. International Journal of Molecular Sciences, 23(22), 14483. https://doi.org/10.3390/ijms232214483