Low Temperature Inhibits the Defoliation Efficiency of Thidiazuron in Cotton by Regulating Plant Hormone Synthesis and the Signaling Pathway

 ,
, 
 and
and 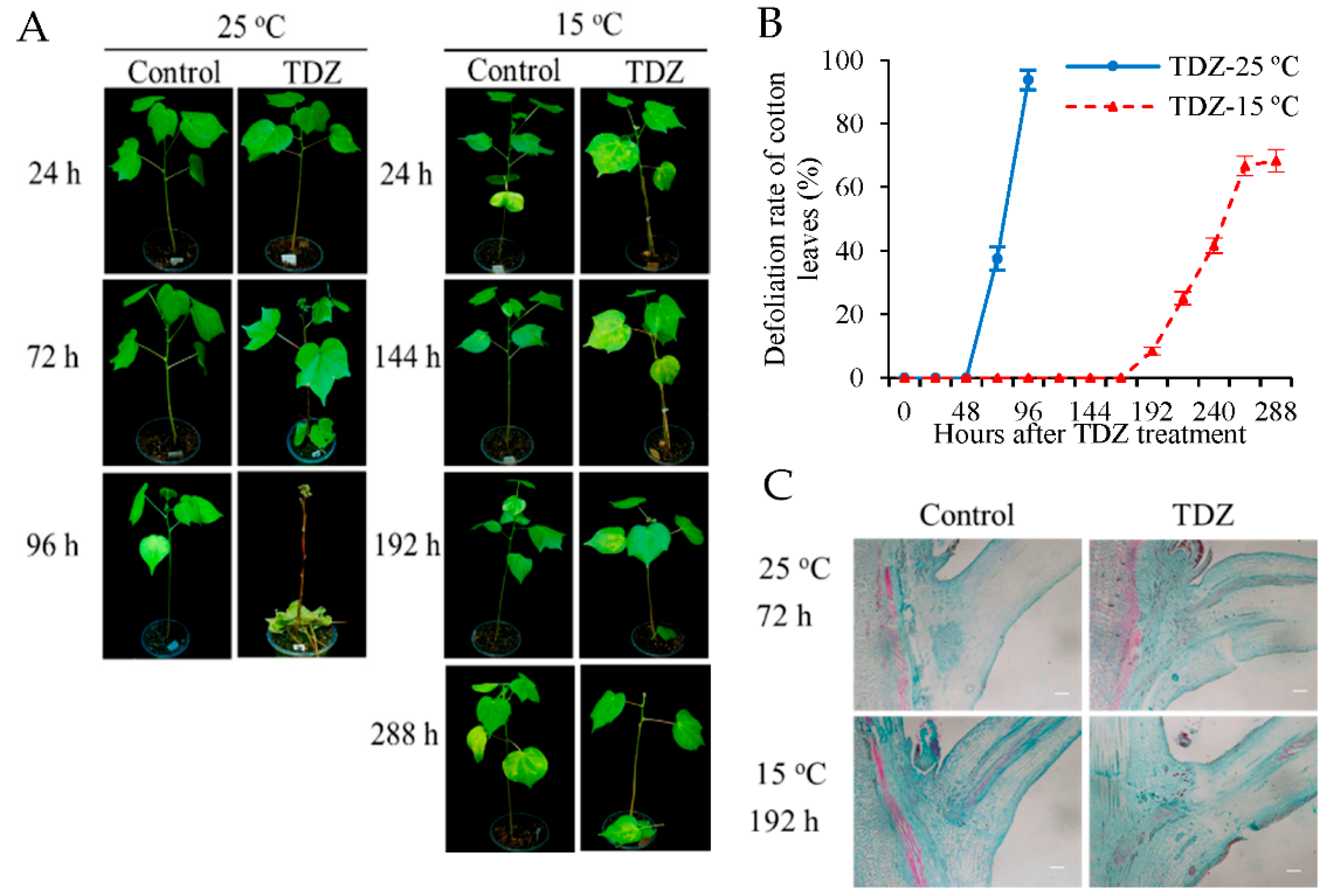{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. The Defoliation Rate of Cotton Leaves Treated with TDZ Is Reduced by Low Temperature Conditions
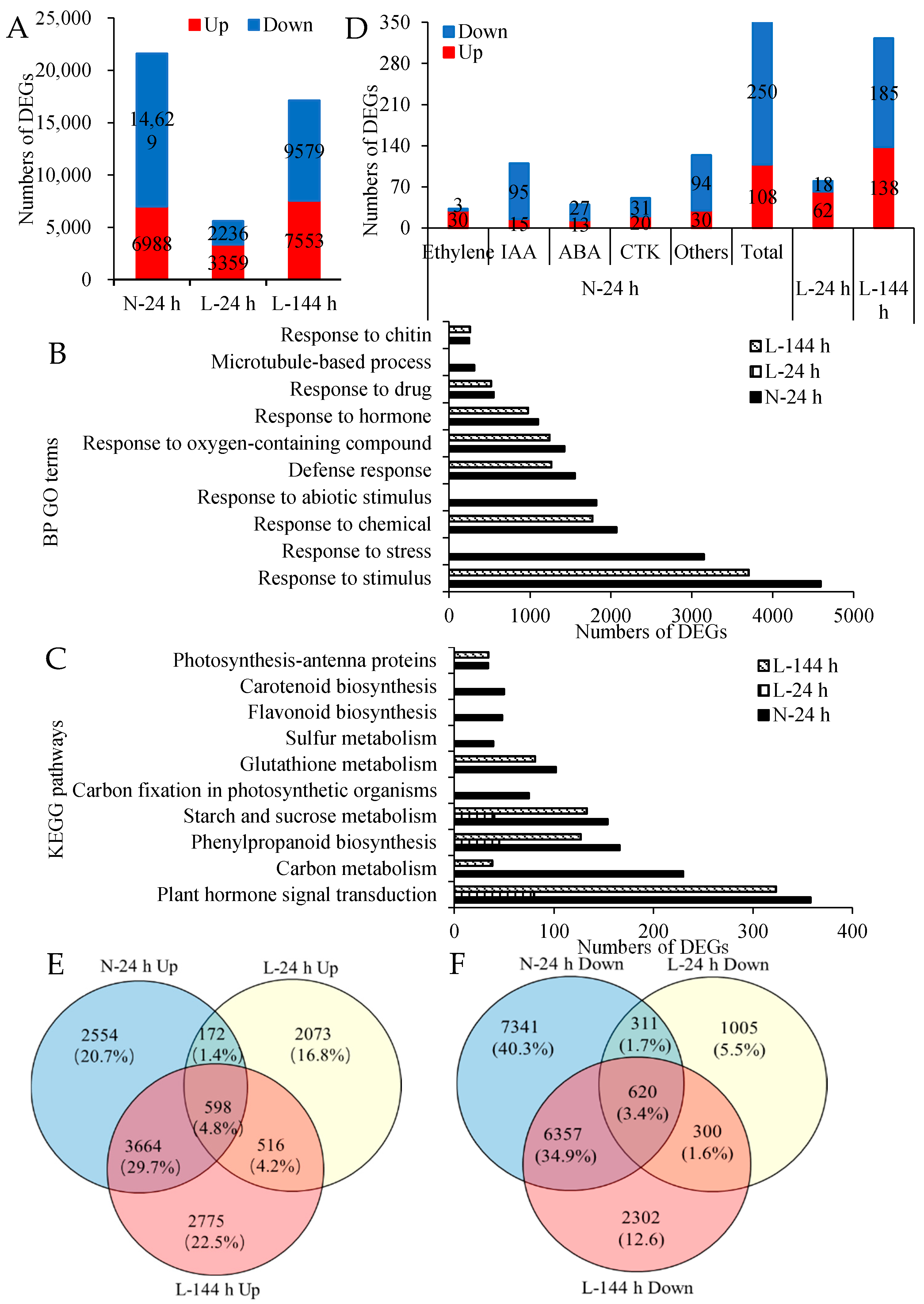
2.2. Transcriptome Change in Cotton AZs Treated with TDZ under Low Temperatures
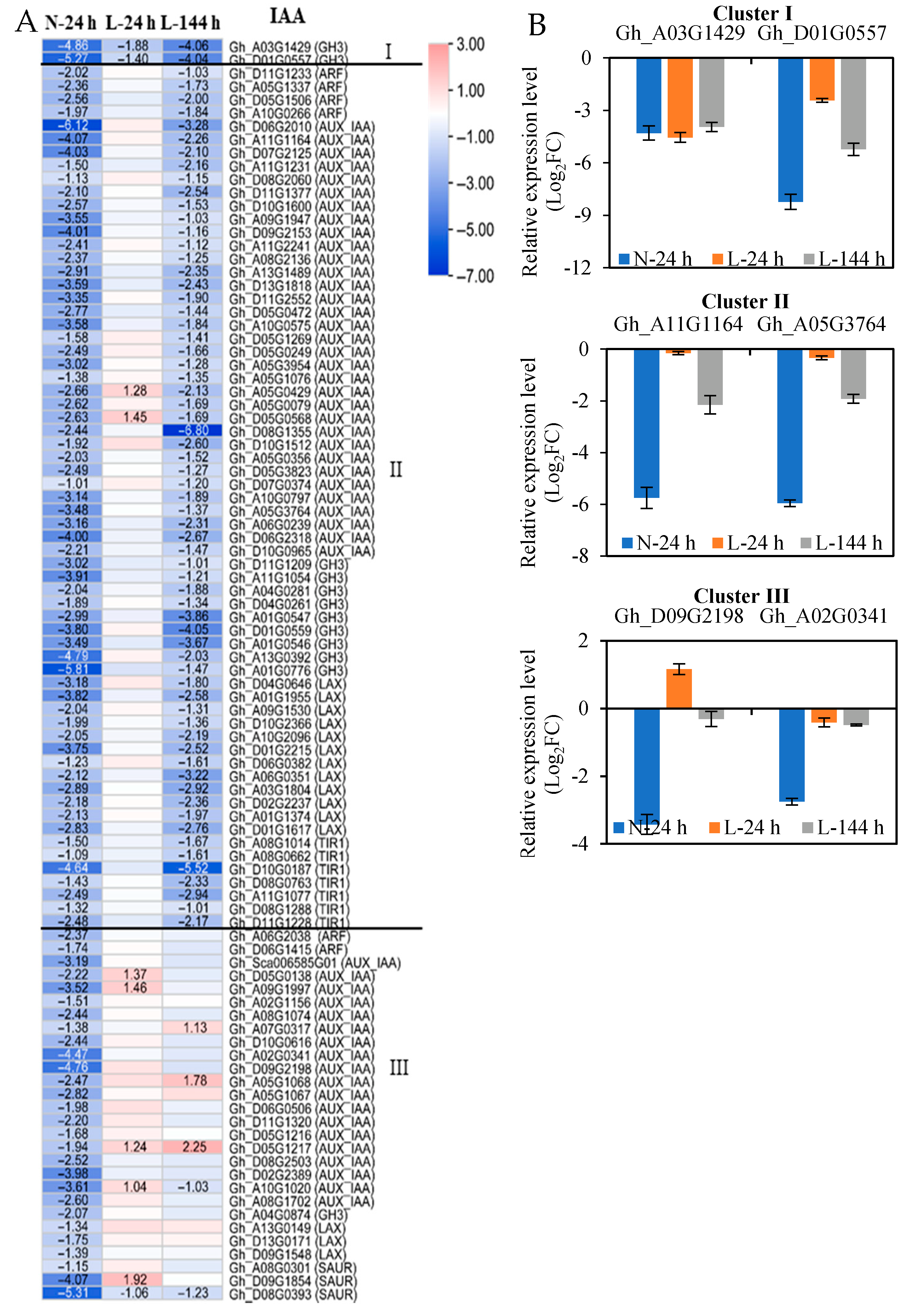
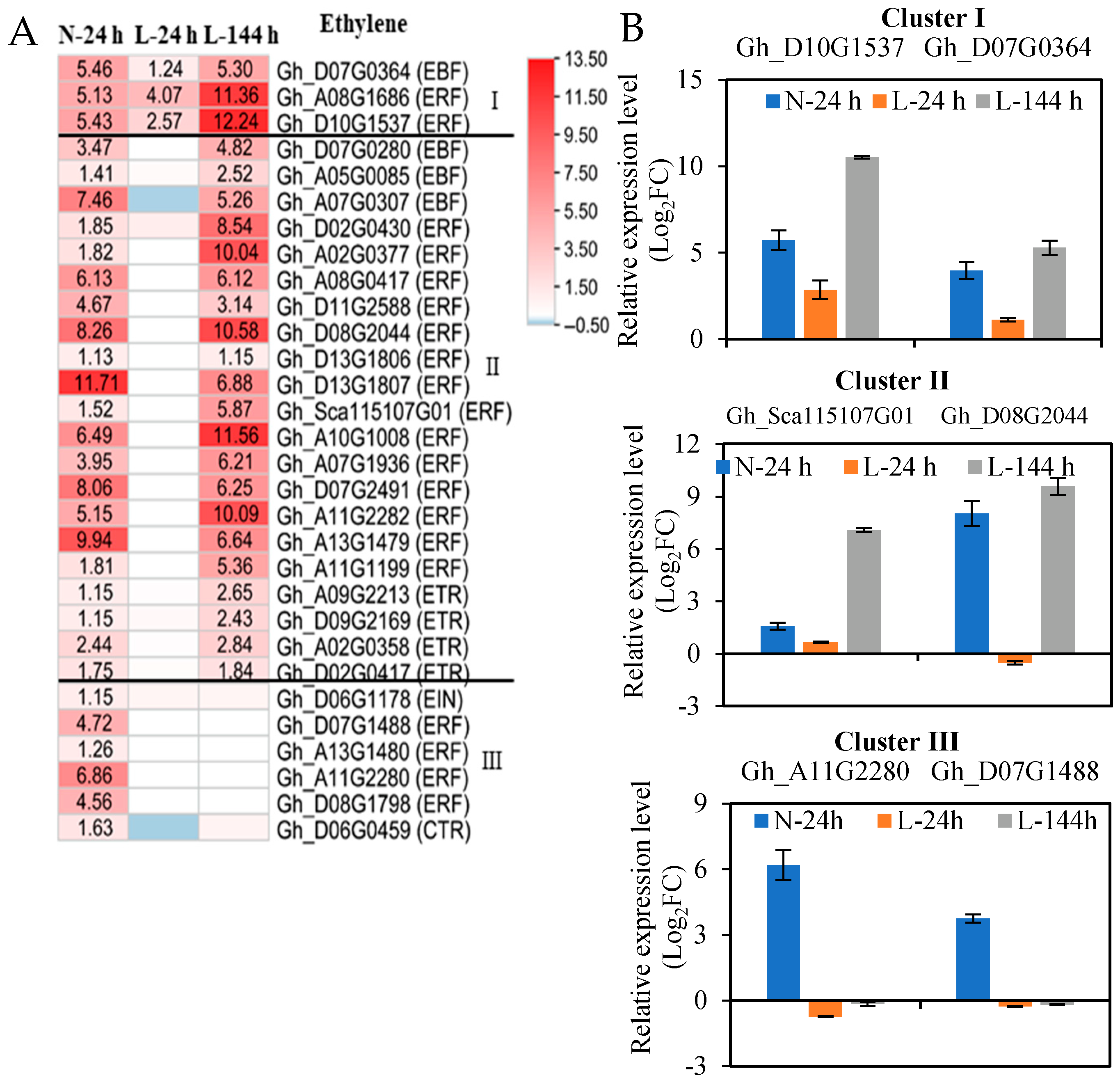
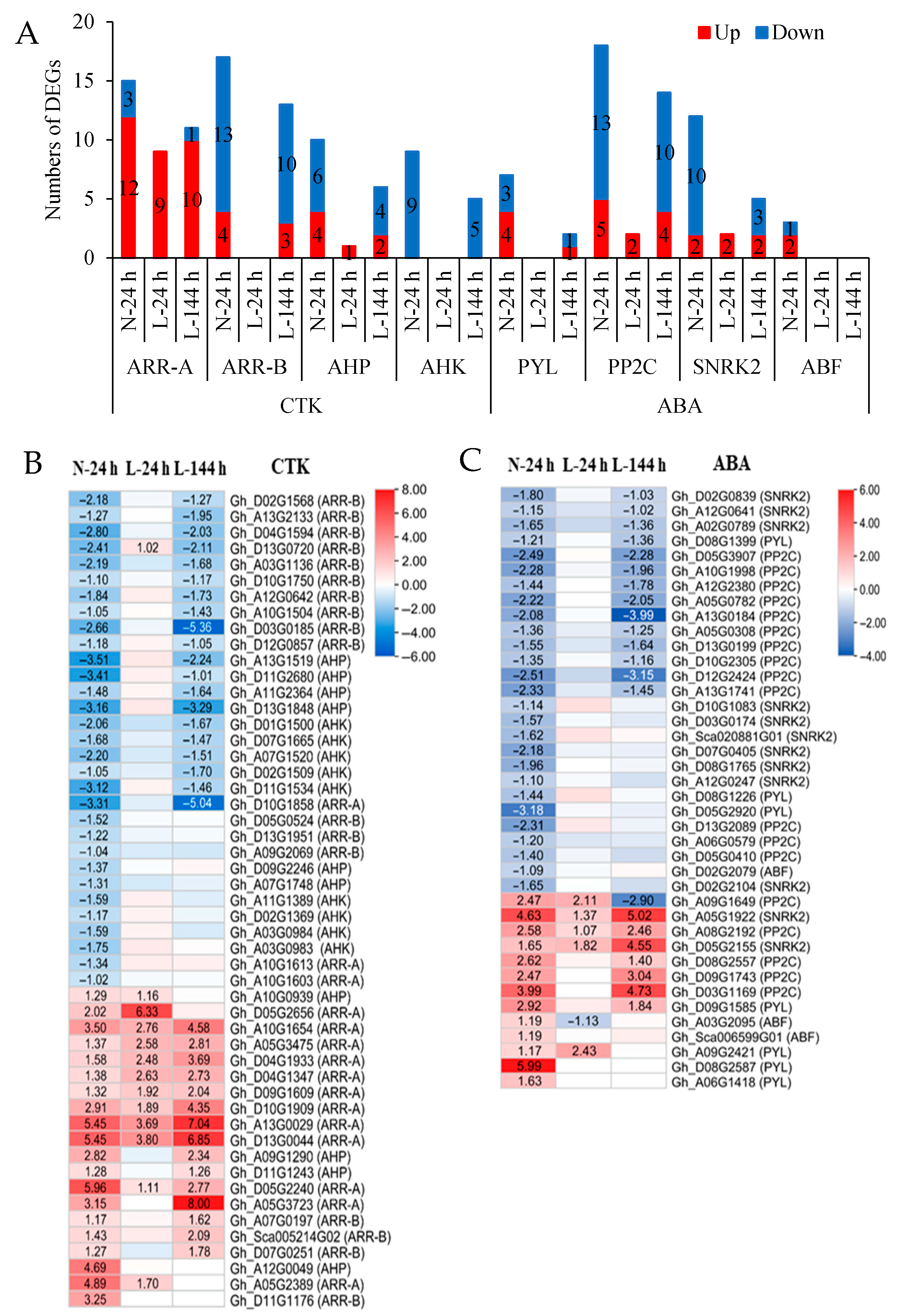
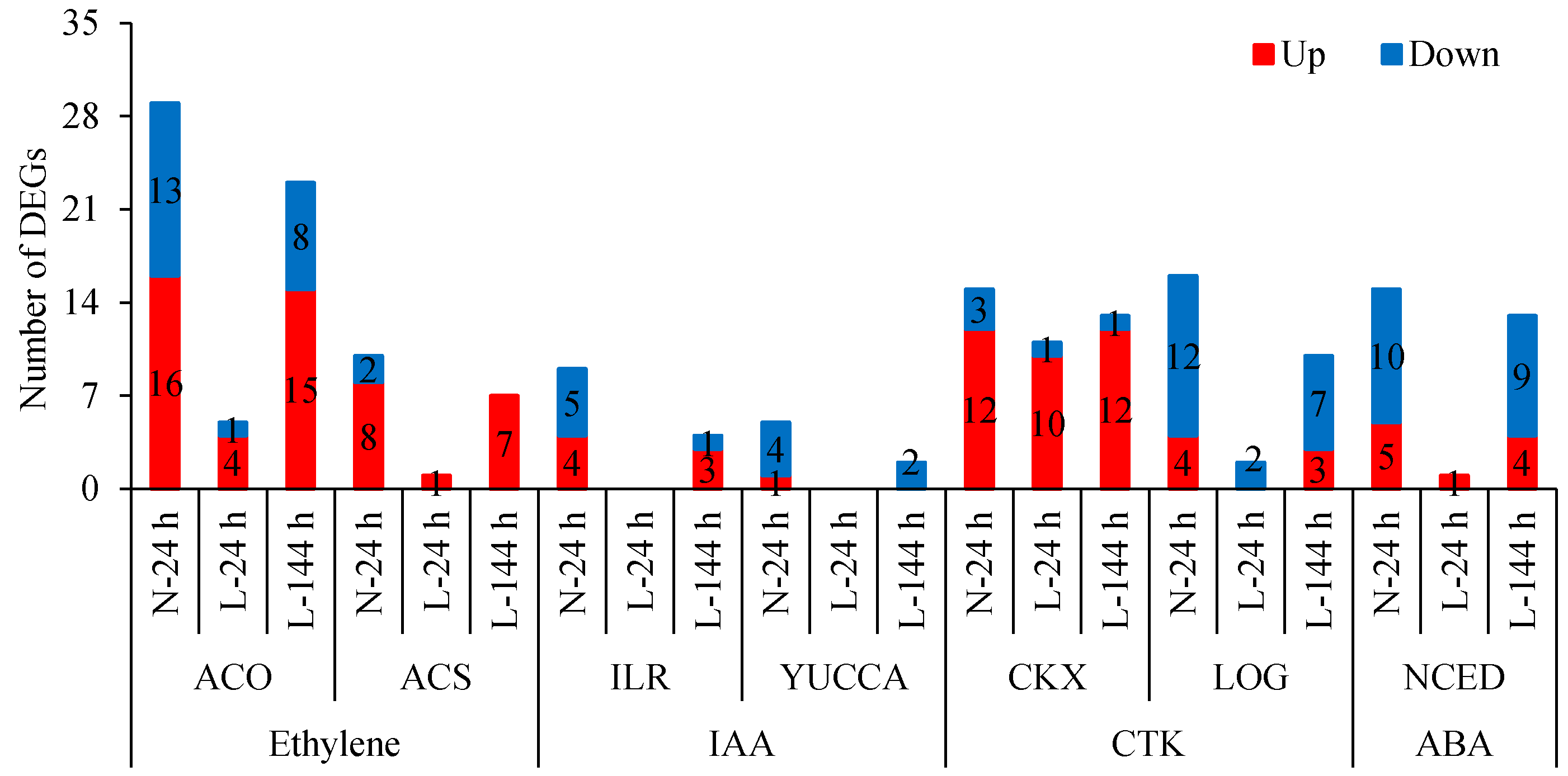
2.3. The Response of Plant Hormone Signaling Genes in Cotton AZs to TDZ Are Regulated by Low Temperatures
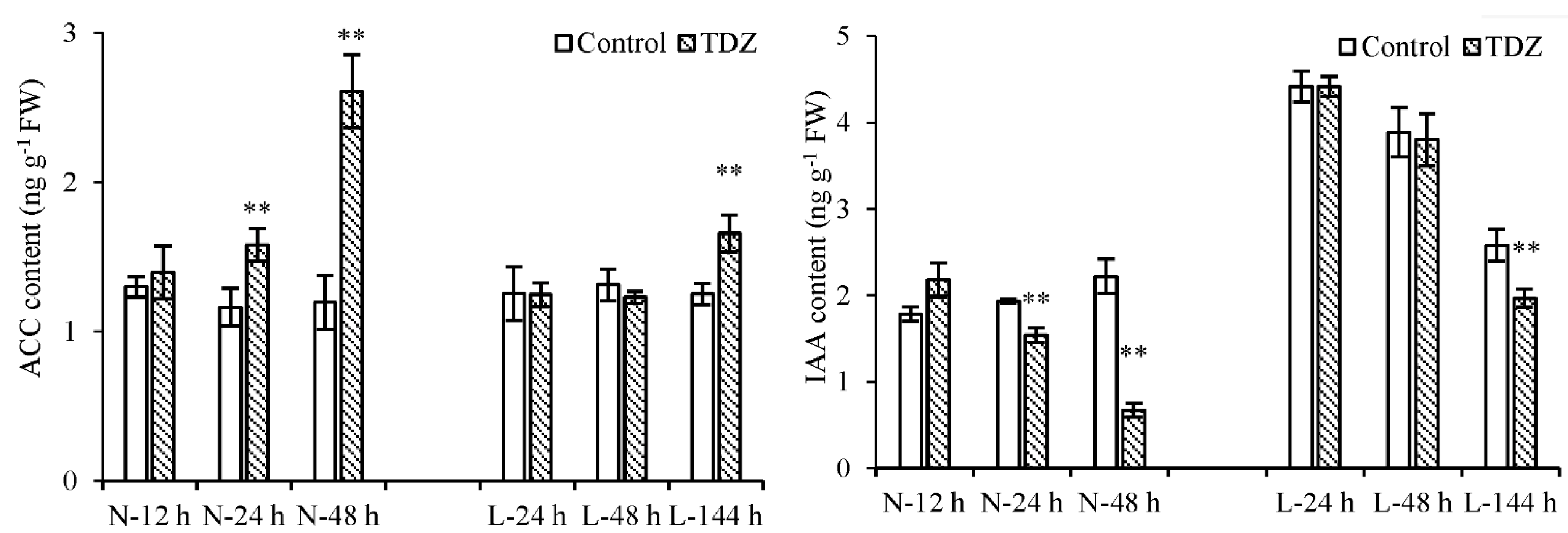
2.4. The Effect of Low Temperature on the Synthesis and Metabolism of Endogenous Plant Hormones in Cotton AZs Treated with TDZ
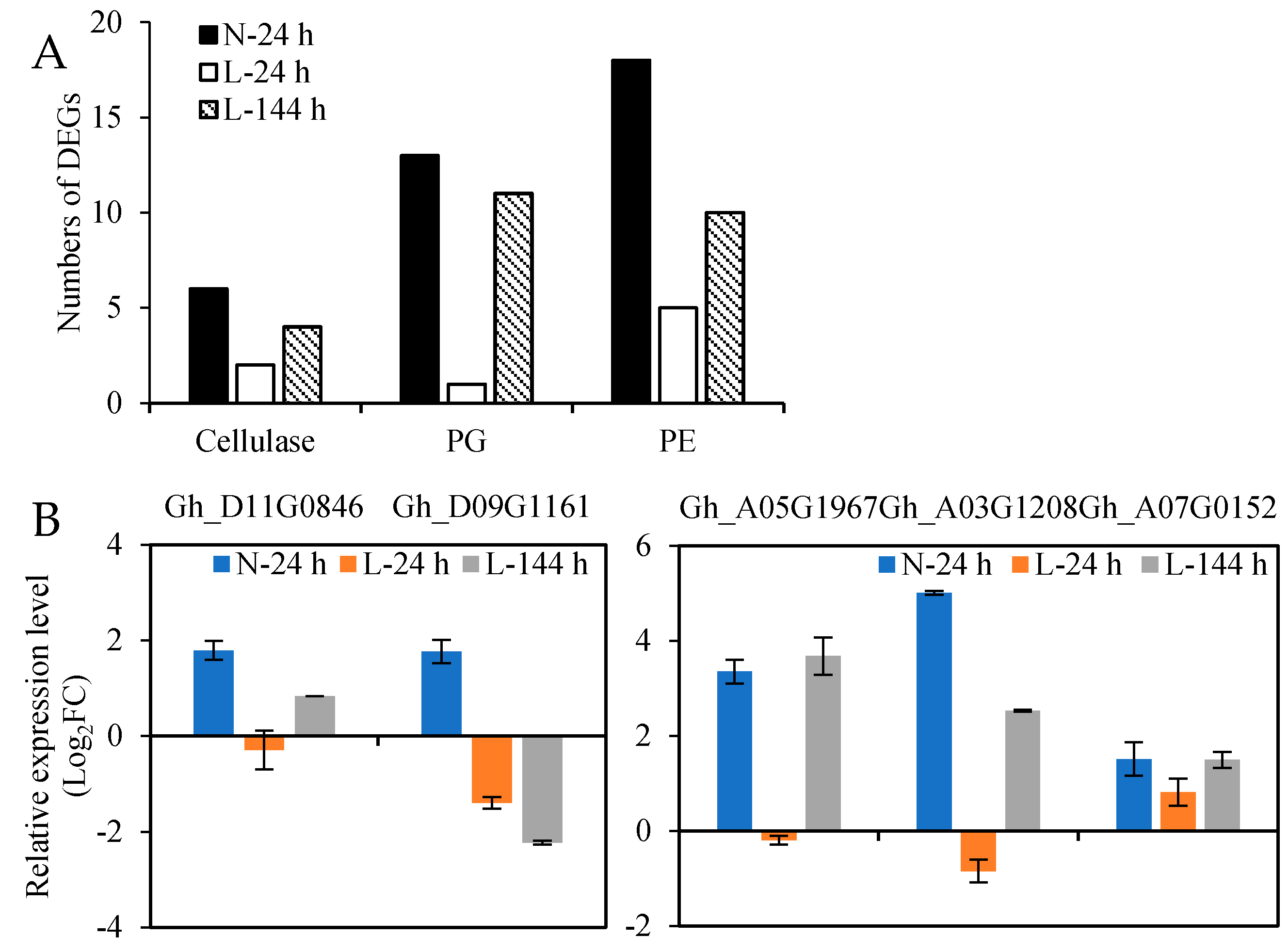
2.5. The Effect of Low Temperature on the Expression of Cell Wall Hydrolase Genes in Cotton AZs Treated with TDZ
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Histological Analysis
4.3. RNA Extraction and Sequencing
4.4. Analysis of DEGs
4.5. Plant Hormone Content
4.6. RT-qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Xu, J.; Chen, L.; Sun, H.; Wusiman, N.; Sun, W.; Li, B.; Gao, Y.; Kong, J.; Zhang, D.; Zhang, X.; et al. Crosstalk between cytokinin and ethylene signaling pathways regulates leaf abscission in cotton in response to chemical defoliants. J. Exp. Bot. 2019, 70, 1525–1538. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, R.; Wang, X.; Zhao, L.; Chen, J.; Yang, C.; Zhang, G.; Ni, W.; Zhang, L. Involvement of hydrogen peroxide in cotton leaf abscission induced by thidiazuron. J. Plant Growth. Regul. 2021, 40, 1667–1673. [Google Scholar] [CrossRef]
- Jin, D.; Wang, X.; Xu, Y.; Gui, H.; Zhang, H.; Dong, Q.; Sikder, R.K.; Yang, G.; Song, M. Chemical defoliant promotes leaf abscission by altering ROS metabolism and photosynthetic efficiency in Gossypium hirsutum. Int. J. Mol. Sci. 2020, 21, 2738. [Google Scholar] [CrossRef] [PubMed]
- Siebert, J.D.; Stewart, A.M. Correlation of defoliation timing methods to optimize cotton yield, quality and revenue. J. Cotton Sci. 2006, 10, 146–154. [Google Scholar]
- Du, M.; Li, Y.; Tian, X.; Duan, L.; Zhang, M.; Tan, W.; Xu, D.; Li, Z. The phytotoxin coronatine induces abscission-related gene expression and boll ripening during defoliation of cotton. PLoS ONE 2014, 9, e97652. [Google Scholar] [CrossRef]
- Kim, J. Four shades of detachment: Regulation of floral organ abscission. Plant Signal. Behav. 2014, 9, e976154. [Google Scholar] [CrossRef]
- Patharkar, O.R.; Walker, J.C. Advances in abscission signaling. J. Exp. Bot. 2018, 69, 733–740. [Google Scholar] [CrossRef]
- Li, F.; Wu, Q.; Liao, B.; Yu, K.; Huo, Y.; Meng, L.; Wang, S.; Wang, B.; Du, M.; Tian, X.; et al. Thidiazuron promotes leaf abscission by regulating the crosstalk complexities between ethylene, auxin, and cytokinin in cotton. Int. J. Mol. Sci. 2022, 23, 2696. [Google Scholar] [CrossRef]
- Mishra, A.; Khare, S.; Trivedi, P.K.; Nath, P. Ethylene induced cotton leaf abscission is associated with higher expression of cellulose (GhCell) and increased activities of ethylene biosynthesis enzymes in abscission zone. Plant Physiol. Bioch. 2008, 46, 54–63. [Google Scholar] [CrossRef]
- Mishra, A.; Khare, S.; Trivedi, P.K.; Nath, P. Effect of ethylene, 1-MCP, ABA and IAA on break strength, cellulose and polygalacturonase activities during cotton leaf abscission. S. Afr. J. Bot. 2008, 74, 282–287. [Google Scholar] [CrossRef]
- Brown, K.M. Ethylene and abscission. Plant Physiol. 1997, 100, 567–576. [Google Scholar] [CrossRef]
- Meir, S.; Philosoph-Hadas, S.; Sundaresan, S.; Selvaraj, K.S.V.; Burd, S.; Ophir, R.; Kochanek, B.; Reid, M.S.; Jiang, C.Z.; Lers, A. Microarray analysis of the abscission-related transcriptome in the tomato flower abscission zone in response to auxin depletion. Plant Physiol. 2010, 154, 1929–1956. [Google Scholar] [CrossRef]
- Liu, J.; Moore, S.; Chen, C.; Lindsey, K. Crosstalk complexities between auxin, cytokinin, and ethylene in Arabidopsis root development: From experiments to systems modeling, and back again. Mol. Plant 2017, 10, 1480–1496. [Google Scholar] [CrossRef]
- Brummel, D.A.; Hall, B.D.; Bennett, A.B. Antisense suppression of tomato endo-1,4-beta-glucanase Cel2 mRNA accumulation increases the force required to break fruit abscission zones but does not affect fruit softening. Plant Mol. Biol. 1999, 40, 615–622. [Google Scholar] [CrossRef]
- Gonzalez-Carranza, Z.H.; Whitelaw, C.A.; Swarup, R.; Roberts, J.A. Temporal and spatial expression of a polygalacturonase during leaf and flower abscission in oilseed rape and arabidopsis. Plant Physiol. 2002, 128, 534–543. [Google Scholar] [CrossRef]
- Snipes, C.E.; Wills, G.D. Influence of temperature and adjuvants on thidiazuron activity in cotton leaves. Weed Sci. 1994, 42, 13–17. [Google Scholar] [CrossRef]
- Eder, Z.P.; Sgingh, S.; Fromme, D.D.; Mott, D.A.; Ibrahim, A.M.H.; Morgan, G.D. Cotton harvest aid regimes and their interaction with cotton cultivar characteristics impacting leaf grade. Agron. J. 2017, 109, 2714–2722. [Google Scholar] [CrossRef]
- Gwathney, C.O.; Hayes, R.M. Harvest-aid interactions under different temperature regimes in field-grown cotton. J. Cotton Sci. 1997, 1, 1–28. [Google Scholar]
- Stewart, A.M.; Edmisten, K.L.; Wells, R. Boll openers in cotton: Effectiveness and environmental influences. Field Crops Res. 2000, 67, 83–90. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Baren, M.J.V.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Taylor, J.E.; Whitelaw, C.A. Signals in abscission. New Phytol. 2001, 151, 323–340. [Google Scholar] [CrossRef]
- Olsson, V.; Butenko, M.A. Abscission in plants. Curr. Biol. 2018, 28, 338–339. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.B.; Wang, G.; Li, Y.Y.; Wang, B.; Zhang, P.; Peng, M. Reactive oxygen species regulate leaf pulvinus abscission zone cell separation in response to water-deficit stress in cassava. Sci. Rep. 2016, 6, 21542. [Google Scholar] [CrossRef] [PubMed]
- Patharkar, O.R.; Walker, J.C. Core mechanisms regulating developmentally timed and environmentally triggered abscission. Plant Physiol. 2016, 172, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.E.; Bleecker, A.B. Ethylene-dependent and-independent processes associated with floral organ abscission in Arabidopsis. Plant Physiol. 2004, 134, 194–203. [Google Scholar] [CrossRef]
- Basu, M.M.; Gonzalez-Carranza, Z.H.; Azam-Ali, S.; Tang, S.Y.; Shahid, A.A.; Roberts, J.A. The manipulation of auxin in the abscission zone cells of Arabidopsis flowers reveals that indoleacetic acid signaling is a prerequisite for organ shedding. Plant Physiol. 2013, 162, 96–106. [Google Scholar] [CrossRef]
- Kucko, A.; Wilmowicz, E.; Pokora, W.; Alche, J.D.D. Disruption of the auxin gradient in the abscission zone area evokes asymmetrical changes leading to flower separation in yellow lupine. Int. J. Mol. Sci. 2020, 21, 3815. [Google Scholar] [CrossRef]
- Suttle, J.C. Disruption of the polar auxin transport system in cotton seedlings following treatment with the defoliant thidiazuron. Plant Physiol. 1988, 86, 241–245. [Google Scholar] [CrossRef]
- Jin, X.; Zimmermann, J.; Polle, A.; Fischer, U. Auxin is a long-range signal that acts independently of ethylene signaling on leaf abscission in Populus. Front. Plant Sci. 2015, 6, 634. [Google Scholar] [CrossRef]
- Patterson, S.E.; Bolivar-Medina, J.L.; Falbel, T.G.; Hedtcke, J.L.; Nevarez-McBride, D.; Maule, A.F.; Zalapa, J.E. Are we on the right track: Can our understanding of abscission in model systems promote or derail making improvements in less studied crops? Front. Plant Sci. 2016, 6, 1268. [Google Scholar] [CrossRef]
- Marciniak, K.; Kucko, A.; Wilmowicz, E.; Swidzinski, M.; Przedniczek, K.; Kopcewicz, J. Gibberellic acid affects the functioning of the flower abscission zone in Lupinus Luteus via cooperation with the ethylene precursor independently of abscisic acid. J. Plant Physiol. 2018, 229, 170–174. [Google Scholar] [CrossRef]
- Thompson, D.S.; Osborne, D.J. A role for the stele in intertissue signaling in the initiation of abscission in bean leaves (Phaseolus vulgaris L.). Plant Physiol. 1994, 105, 341–347. [Google Scholar] [CrossRef][Green Version]
- Wilmowicz, E.; Frankowski, K.; Kucko, A.; Swidzinski, M.; Alche, J.D.; Nowakowska, A.; Kopcewicza, J. The influence of abscisic acid on the ethylene biosynthesis pathway in the functioning of the flower abscission zone in Lupinus luteus. J. Plant Physiol. 2016, 206, 49–58. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, W.; Zeng, Q.; Song, S.; Zhang, M.; Li, X.; Hou, L.; Xiao, Y.; Luo, M.; Li, D.; et al. Moderately enhancing cytokinin level by down-regulation of GhCKX expression in cotton concurrently increases fiber and seed yield. Mol. Breeding 2015, 35, 60. [Google Scholar] [CrossRef]
- Wang, Y.; Li, T.; Meng, H.; Sun, X. Optimal and spatial analysis of hormones, degrading enzymes and isozyme profiles in tomato pedicel explants during ethylene-induced abscission. Plant Growth Regul. 2005, 46, 97–107. [Google Scholar] [CrossRef]
- Merelo, P.; Agusti, J.; Arbona, V.; Costa, M.L.; Estornell, L.H.; Gomez-Cadenas, A.; Coimbra, S.; Gomes, M.D.; Perez-Amador, M.A.; Domingo, C.; et al. Cell wall remodeling in abscission zone cells during ethylene-promoted fruit abscission in citrus. Front. Plant Sci. 2017, 8, 126. [Google Scholar]
- Yang, Y.; Anderson, C.T.; Cao, J. Polygalacturonase45 cleaves pectin and links cell proliferation and morphogenesis to leaf curvature in Arabidopsis thaliana. Plant J. 2021, 106, 1493–1508. [Google Scholar] [CrossRef]
- Zhai, Z.; Feng, C.; Wang, Y.; Sun, Y.; Peng, X.; Xiao, Y.; Zhang, X.; Zhou, X.; Jiao, J.; Wang, W.; et al. Genome-wide identification of the xyloglucan endotransglucosylase /hydrolase (XTH) and polygalacturonase (PG) genes and characterization of their role in fruit softening of sweet cherry. Int. J. Mol. Sci. 2021, 22, 12331. [Google Scholar] [CrossRef]
- McManus, M.T. Further examination of abscission zone cells as ethylene target cells in higher plants. Ann. Bot. 2008, 101, 285–292. [Google Scholar] [CrossRef]
- Jin, D.; Xu, Y.; Gui, H.; Zhang, H.; Dong, Q.; Sikder, R.K.; Wang, X.; Yang, G.; Song, M. Evaluation of cotton (Gossypium hirsutum L.) leaf abscission sensitivity triggered by thidiazuron through membership function Value. Plants 2021, 10, 49. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc.TM-1) provides a resource for fiber improvement. Nat. Biotech. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Ruth, W.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, H.; Sun, S.; Wang, X.; Yang, C.; Zhang, G.; Meng, Y.; Wang, Y.; Hu, W.; Liu, R. Low Temperature Inhibits the Defoliation Efficiency of Thidiazuron in Cotton by Regulating Plant Hormone Synthesis and the Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 14208. https://doi.org/10.3390/ijms232214208
Shu H, Sun S, Wang X, Yang C, Zhang G, Meng Y, Wang Y, Hu W, Liu R. Low Temperature Inhibits the Defoliation Efficiency of Thidiazuron in Cotton by Regulating Plant Hormone Synthesis and the Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(22):14208. https://doi.org/10.3390/ijms232214208
Chicago/Turabian StyleShu, Hongmei, Shangwen Sun, Xiaojing Wang, Changqin Yang, Guowei Zhang, Yali Meng, Youhua Wang, Wei Hu, and Ruixian Liu. 2022. "Low Temperature Inhibits the Defoliation Efficiency of Thidiazuron in Cotton by Regulating Plant Hormone Synthesis and the Signaling Pathway" International Journal of Molecular Sciences 23, no. 22: 14208. https://doi.org/10.3390/ijms232214208
APA StyleShu, H., Sun, S., Wang, X., Yang, C., Zhang, G., Meng, Y., Wang, Y., Hu, W., & Liu, R. (2022). Low Temperature Inhibits the Defoliation Efficiency of Thidiazuron in Cotton by Regulating Plant Hormone Synthesis and the Signaling Pathway. International Journal of Molecular Sciences, 23(22), 14208. https://doi.org/10.3390/ijms232214208