

In Vitro and In Silico Analysis of New n-Butyl and Isobutyl Quinoxaline-7-carboxylate 1,4-di-N-oxide Derivatives against Trypanosoma cruzi as Trypanothione Reductase Inhibitors

 , , , , ,
, , , , , 
Abstract
1. Introduction
2. Results
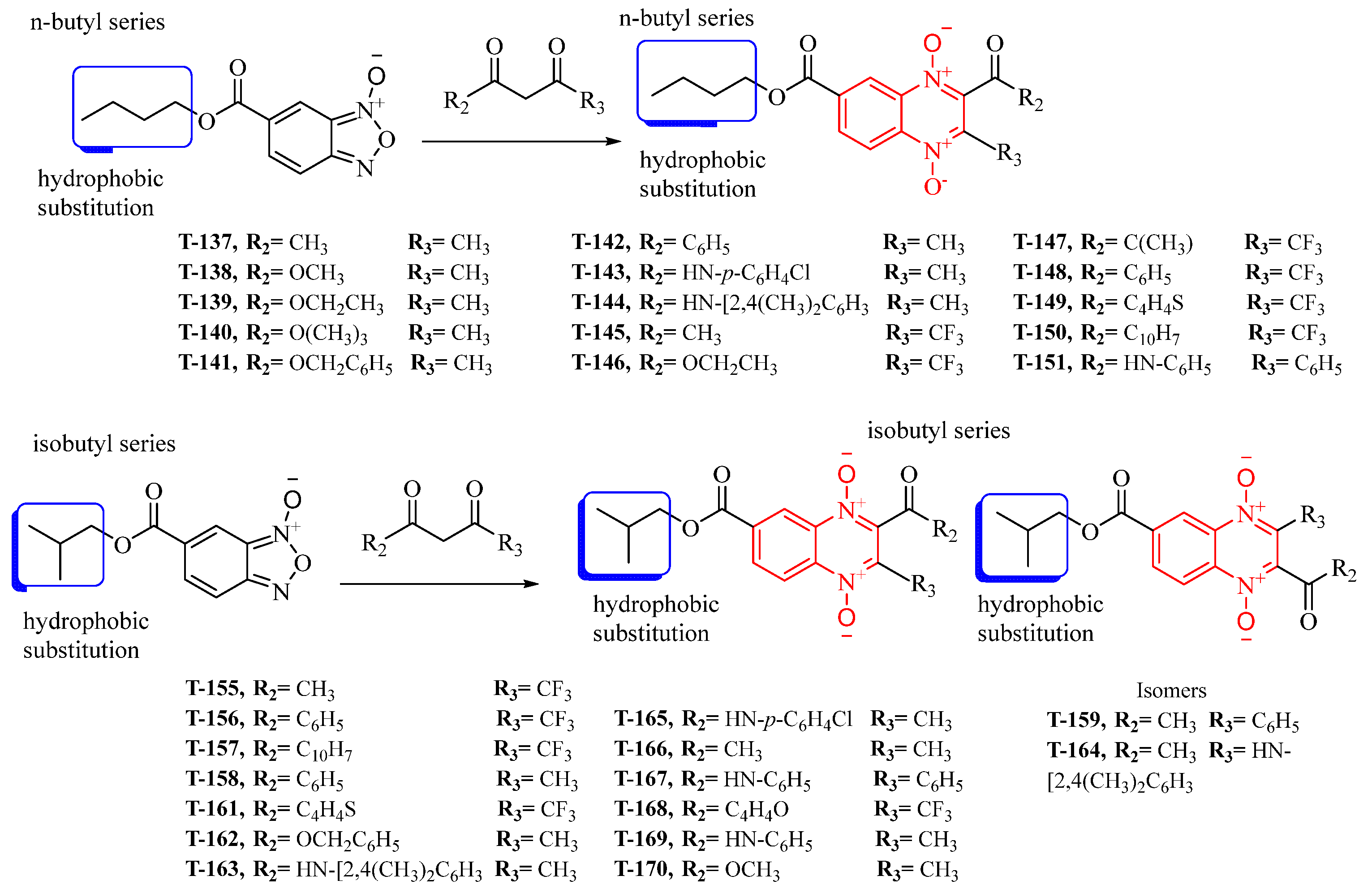
2.1. Synthesis
2.2. Biological Evaluation
Trypanocidal Activity against Trypomastigotes
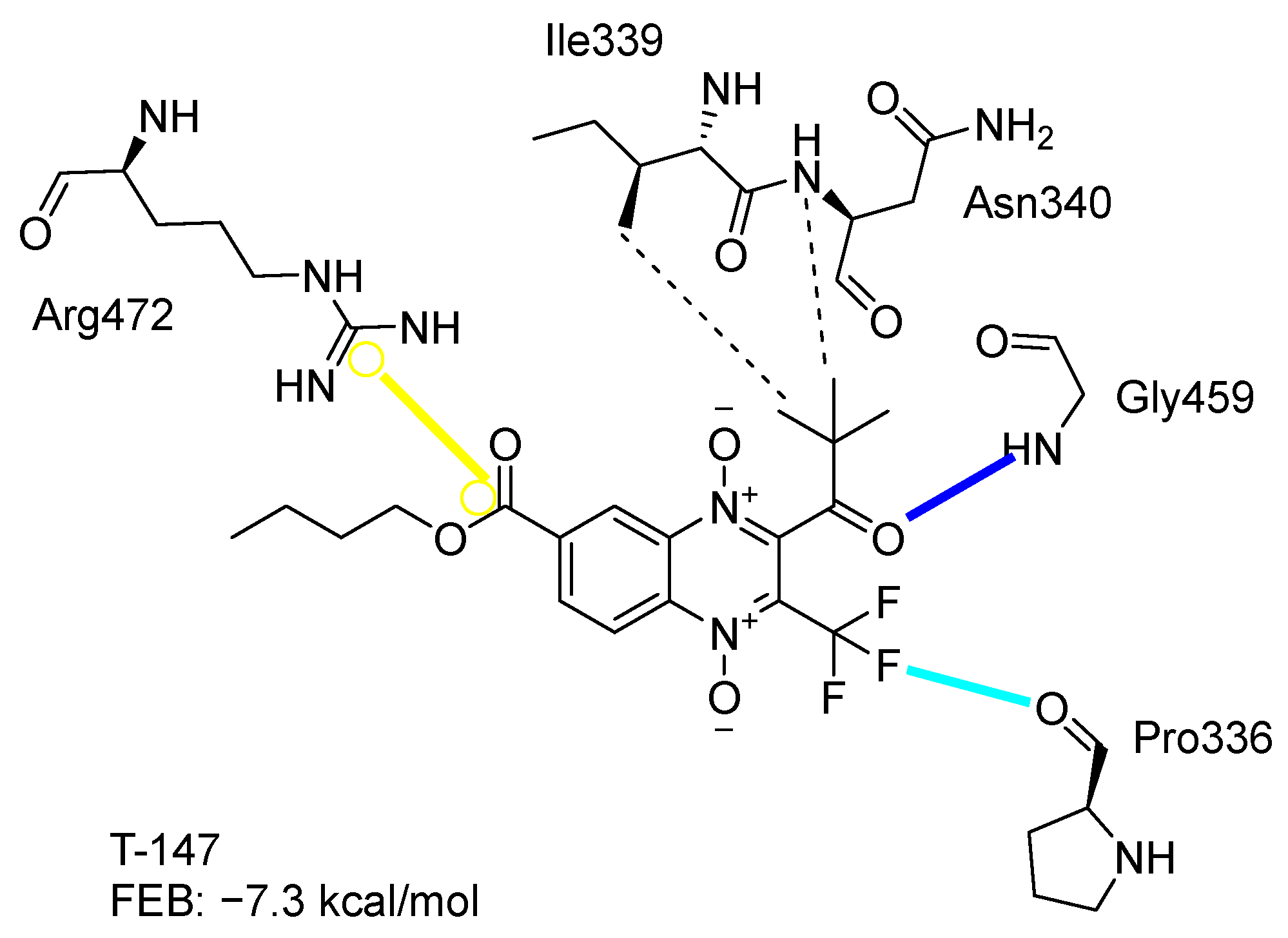
2.3. Molecular Docking on TcTR
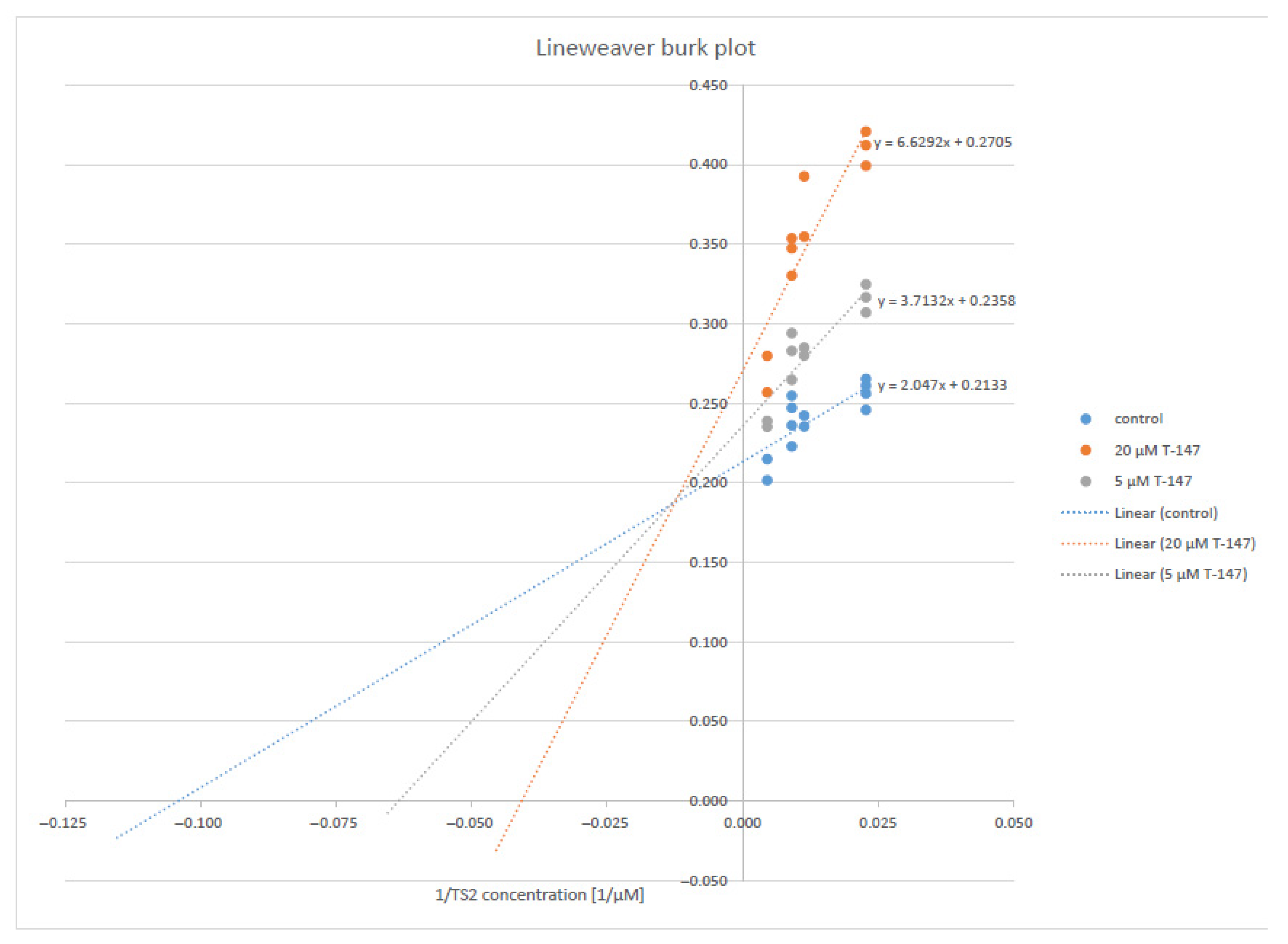
2.4. Enzymatic Activity Evaluation
Trypanothione Reductase Inhibition
2.5. Molecular Dynamics on TcTR
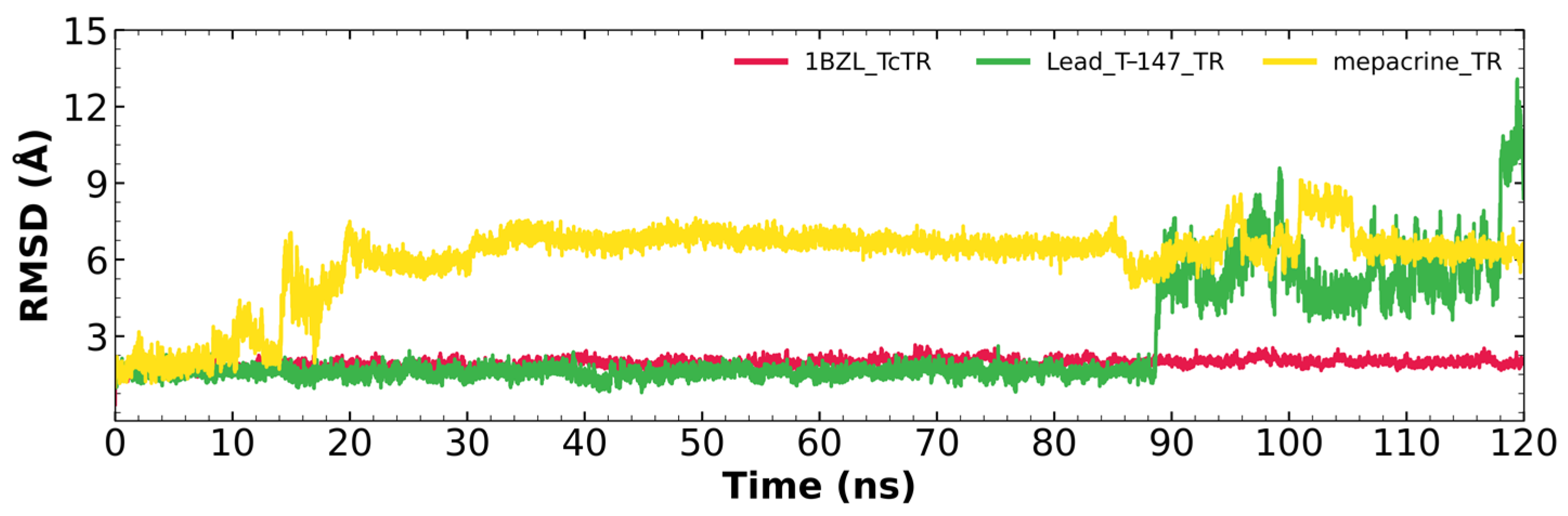
2.5.1. RMSD Analysis
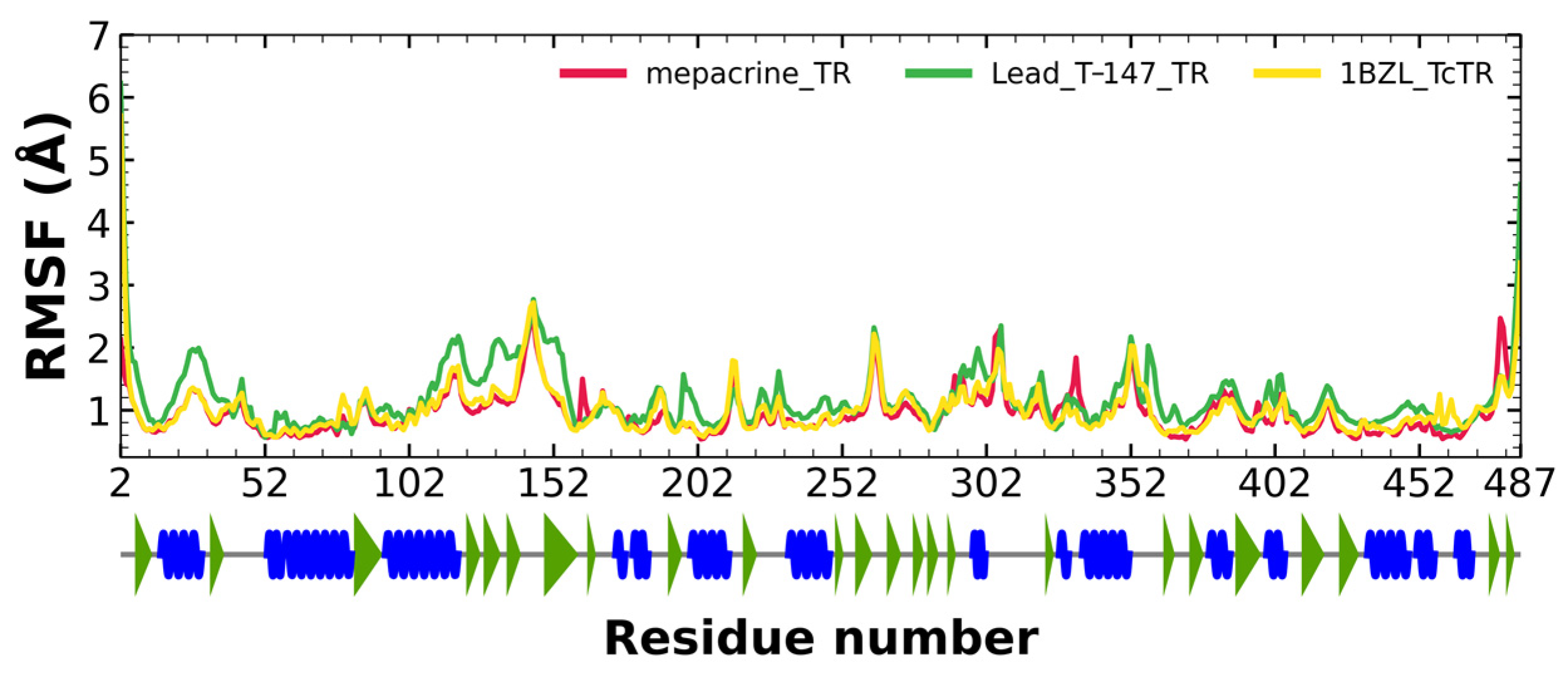
2.5.2. RMSF Analysis
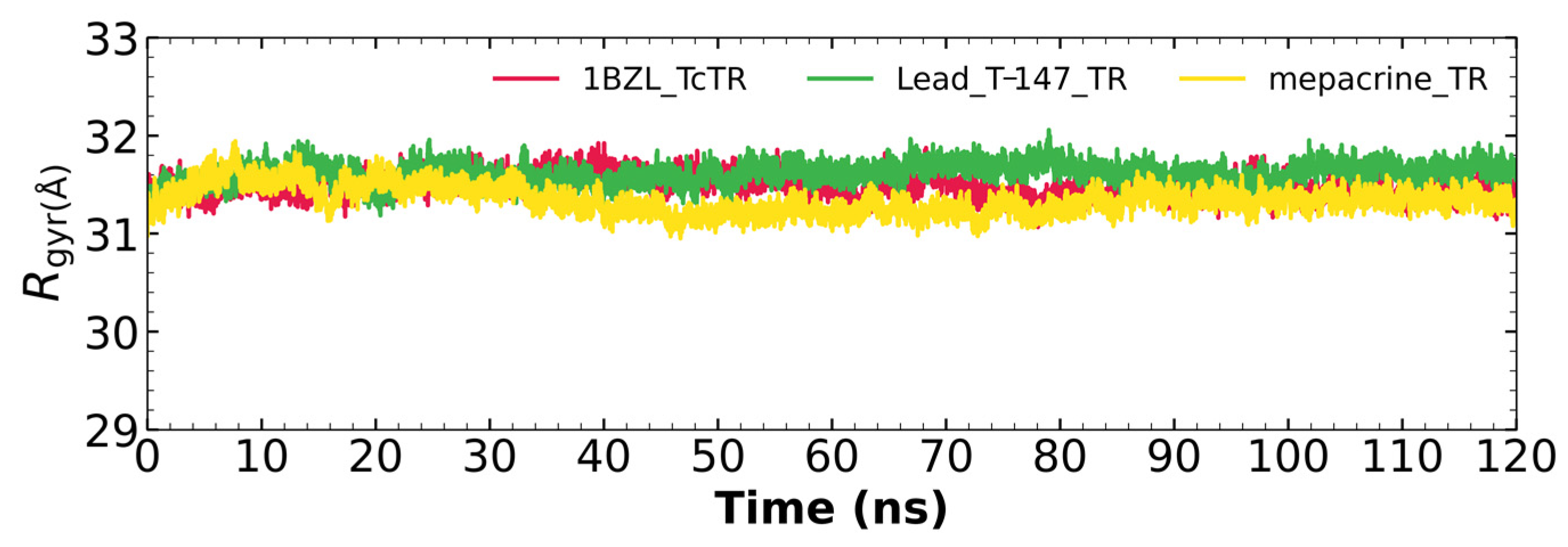
2.5.3. Radius of Gyration
2.6. In Vitro Selectivity Assessment of Enzymatic Activity
2.7. Molecular Dynamics on hGR
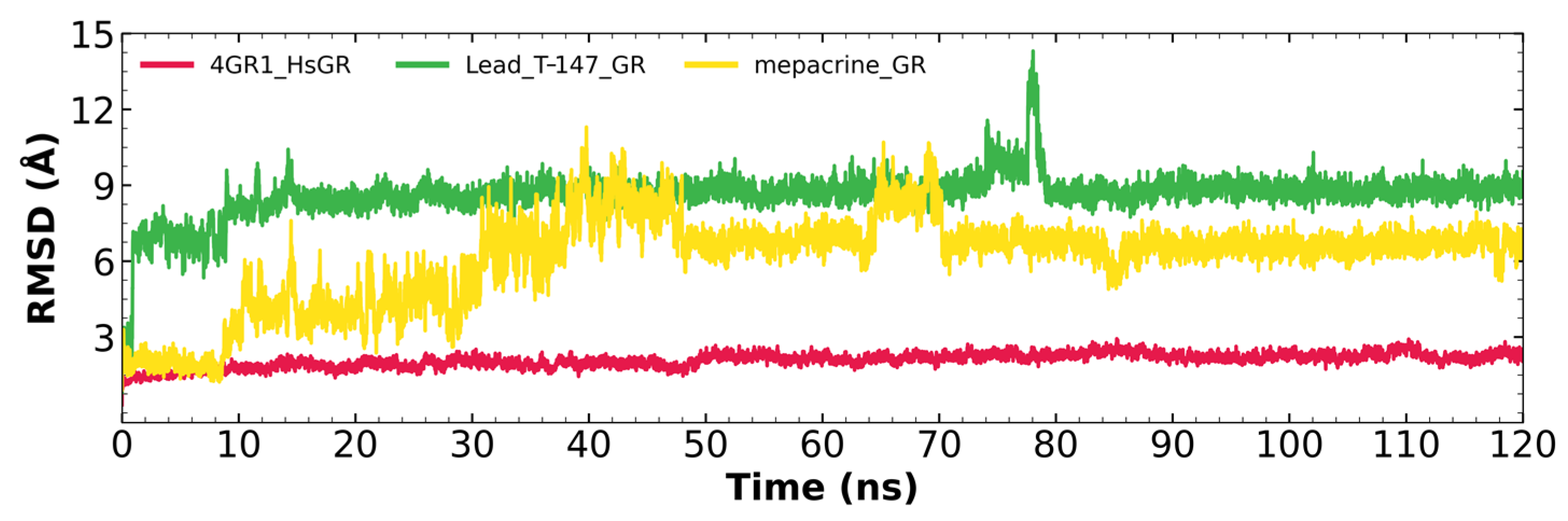
2.7.1. RMSD Analysis
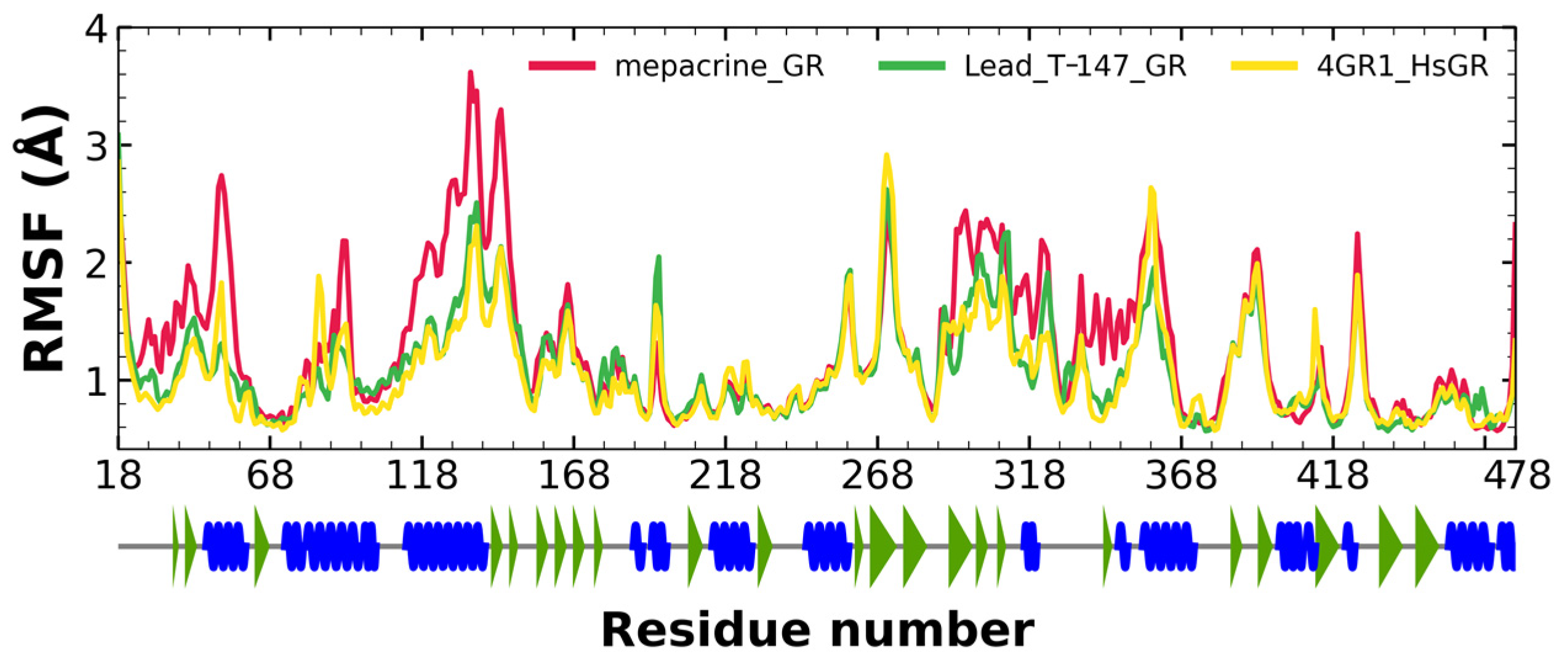
2.7.2. RMSF Analysis
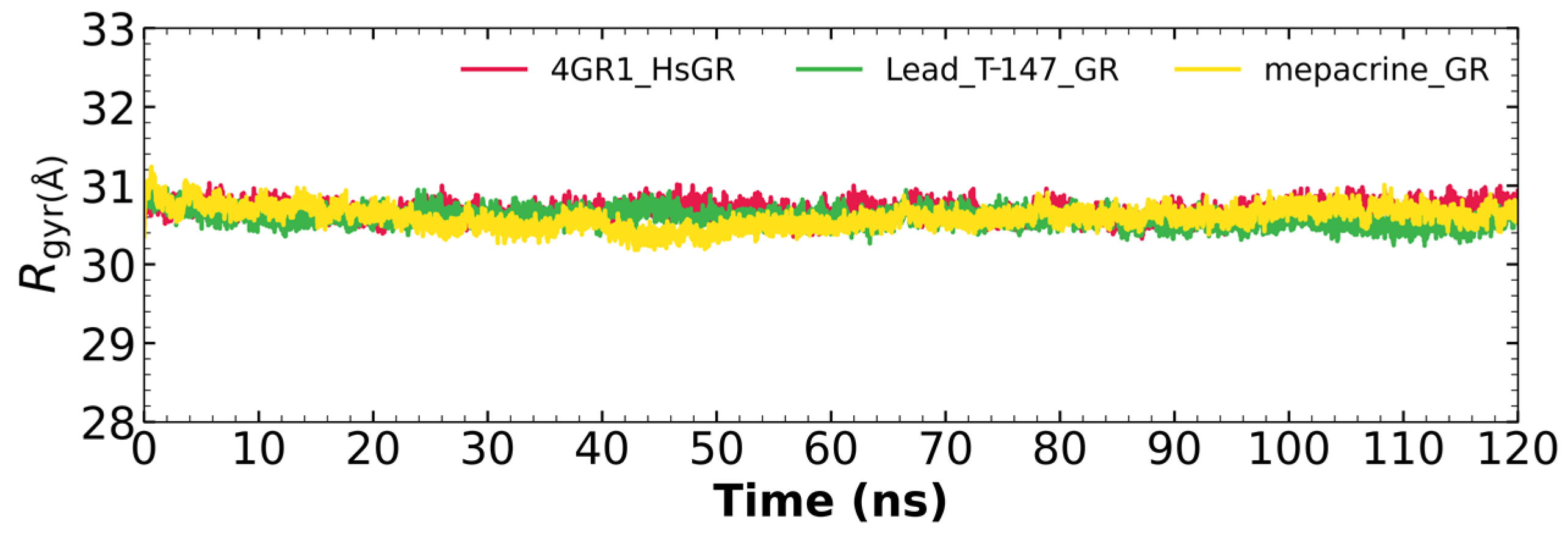
2.7.3. Radius of Gyration
3. Discussion
3.1. Biological Activity
3.1.1. Trypanocidal Activity
3.1.2. Structure-Activity Relationship
3.2. Molecular Docking on Trypansoma cruzi Trypanothione Reductase (TcTR)
3.3. Trypanothione Reductase Inhibition
3.4. In Silico Analysis of Inhibitor Binding TcTR
3.5. Glutathione Reductase Inhibition
3.6. In Silico Analysis of Inhibitor Binding hGR
4. Materials and Methods
4.1. Chemical Synthesis
4.2. Biological Evaluation
4.2.1. Parasite Culture
4.2.2. Trypanocidal Activity against Trypomastigotes
4.3. Molecular Docking Analysis
4.4. Molecular Dynamics
4.5. Enzymatic Activity Evaluation
4.5.1. Trypanothione Reductase
4.5.2. Glutathione Reductase
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO/Chagas Disease (American Trypanosomiasis). Available online: http://www.who.int/mediacentre/factsheets/fs340/en/ (accessed on 18 March 2018).
- Pereira, K.S.; Schmidt, F.L.; Barbosa, R.L.; Guaraldo, A.M.; Franco, R.M.; Dias, V.L.; Passos, L.A. Transmission of Chagas Disease (American Trypanosomiasis) by Food. Adv. Food Nutr. Res. 2010, 59, 63–85. [Google Scholar] [CrossRef]
- Briceno, L.; Mosca, W. Quello Che Non Si Cerca Difficilmente Si Trova: La Malattia Di Chagas. G Ital. Cardiol. 2016, 17, 343–347. [Google Scholar] [CrossRef]
- Rajão, M.A.; Furtado, C.; Alves, C.L.; Passos-Silva, D.G.; de Moura, M.B.; Schamber-Reis, B.L.; Kunrath-Lima, M.; Zuma, A.A.; Vieira-da-Rocha, J.P.; Garcia, J.B.F.; et al. Unveiling Benznidazole’s mechanism of action through overexpression of DNA repair proteins in Trypanosoma cruzi. Environ. Mol. Mutagen. 2014, 55, 309–321. [Google Scholar] [CrossRef]
- Vazquez-Jimenez, L.K.; Hernandez-Posada, M.I.; Paz-Gonzalez, A.D.; Nogueda-Torres, B.; Martinez-Vazquez, A.V.; Herrera-Mayorga, V.; Bocanegra-García, V.; Rivera, G. Analysis of the Effect of Methyl 2-Acetamide-3-Methylquinoxaline-7-Carboxylate 1,4-Di-N-Oxide on the Relative Expression of the Trypanothione Reductase Gene in Trypanosoma Cruzi Epimastigotes. Pak. J. Pharm. Sci. 2019, 32, 1447–1452. [Google Scholar]
- Moreno-Herrera, A.; Cortez-Maya, S.; Bocanegra-Garcia, V.; Banik, B.K.; Rivera, G. Recent Advances in the Development of Broad-Spectrum Antiprotozoal Agents. Curr. Med. Chem. 2021, 28, 583–606. [Google Scholar] [CrossRef]
- Palos, I.; Moo-Puc, R.; Vique-Sánchez, J.L.; Benítez-Cardoza, C.G.; Monge, A.; Villalobos-Rocha, J.C.; Paz-Gonzalez, A.D.; Rivera, G. Esters of Quinoxaline-7-Carboxylate-1,4-Di-N-Oxide as Trichomonas Vaginalis Triosephosphate Isomerase Inhibitors. Acta Pharm. 2021, 71, 485–495. [Google Scholar] [CrossRef]
- Sanchez-Sanchez, M.; Rivera, G.; A. Garcia, E.; Bocanegra-Garcia, V. Therapeutic Targets for the Development of Anti-Trypanosoma Cruzi Drugs: A Brief Review. Mini-Rev. Org. Chem. 2016, 13, 227–243. [Google Scholar] [CrossRef]
- Arioka, S.; Sakagami, M.; Uematsu, R.; Yamaguchi, H.; Togame, H.; Takemoto, H.; Hinou, H.; Nishimura, S.I. Potent Inhibitor Scaffold against Trypanosoma Cruzi Trans-Sialidase. Bioorg. Med. Chem. 2010, 18, 1633–1640. [Google Scholar] [CrossRef]
- Kashif, M.; Chacón-Vargas, K.F.; López-Cedillo, J.C.; Nogueda-Torres, B.; Paz-González, A.D.; Ramírez-Moreno, E.; Agusti, R.; Uhrig, M.L.; Reyes-Arellano, A.; Peralta-Cruz, J.; et al. Synthesis, Molecular Docking and Biological Evaluation of Novel Phthaloyl Derivatives of 3-Amino-3-Aryl Propionic Acids as Inhibitors of Trypanosoma Cruzi Trans-Sialidase. Eur. J. Med. Chem. 2018, 156, 252–268. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Jiménez, L.K.; Paz-González, A.D.; Juárez-Saldivar, A.; Uhrig, M.L.; Agusti, R.; Reyes-Arellano, A.; Nogueda-Torres, B.; Rivera, G. Structure-Based Virtual Screening of New Benzoic Acid Derivatives as Trypanosoma Cruzi Trans-Sialidase Inhibitors. Med. Chem. 2021, 17, 724–731. [Google Scholar] [CrossRef]
- Kashif, M.; Moreno-Herrera, A.; Lara-Ramirez, E.E.; Ramírez-Moreno, E.; Bocanegra-García, V.; Ashfaq, M.; Rivera, G. Recent Developments in Trans-Sialidase Inhibitors of Trypanosoma Cruzi. J. Drug Target. 2017, 25, 485–498. [Google Scholar] [CrossRef]
- Vázquez-Jiménez, L.K.; Paz-González, A.D.; Kashif, M.; Juárez-Rendón, K.J.; Nogueda-Torres, B.; Bocanegra-García, V.; Rivera, G. Effect of 4-Amino-3-Nitrobenzoic Acid on the Expression Level of the Trans-Sialidase Gene in Trypanosoma Cruzi Epimastigotes. Pak. J. Pharm. Sci. 2019, 32, 825–829. [Google Scholar]
- Kashif, M.; Moreno-Herrera, A.; Villalobos-Rocha, J.C.; Nogueda-Torres, B.; Pérez-Villanueva, J.; Rodríguez-Villar, K.; Medina-Franco, J.L.; de Andrade, P.; Carvalho, I.; Rivera, G. Benzoic Acid Derivatives with Trypanocidal Activity: Enzymatic Analysis and Molecular Docking Studies toward Trans-Sialidase. Molecules 2017, 22, 1863. [Google Scholar] [CrossRef]
- Herrera-Mayorga, V.; Lara-Ramírez, E.E.; Chacón-Vargas, K.F.; Aguirre-Alvarado, C.; Rodríguez-Páez, L.; Alcántara-Farfán, V.; Cordero-Martínez, J.; Nogueda-Torres, B.; Reyes-Espinosa, F.; Bocanegra-García, V.; et al. Structure-Based Virtual Screening and In Vitro Evaluation of New Trypanosoma Cruzi Cruzain Inhibitors. Int. J. Mol. Sci. 2019, 20, 1742. [Google Scholar] [CrossRef]
- Rocha, D.A.; Silva, E.B.; Fortes, I.S.; Lopes, M.S.; Ferreira, R.S.; Andrade, S.F. Synthesis and Structure-Activity Relationship Studies of Cruzain and Rhodesain Inhibitors. Eur. J. Med. Chem. 2018, 157, 1426–1459. [Google Scholar] [CrossRef]
- Delgado-Maldonado, T.; Nogueda-Torres, B.; Espinoza-Hicks, J.C.; Vázquez-Jiménez, L.K.; Paz-González, A.D.; Juárez-Saldívar, A.; Rivera, G. Synthesis and Biological Evaluation in Vitro and in Silico of N-Propionyl-N′-Benzeneacylhydrazone Derivatives as Cruzain Inhibitors of Trypanosoma Cruzi. Mol. Divers. 2020, 1, 39–50. [Google Scholar] [CrossRef]
- Palos, I.; Lara-Ramirez, E.E.; Lopez-Cedillo, J.C.; Garcia-Perez, C.; Kashif, M.; Bocanegra-Garcia, V.; Nogueda-Torres, B.; Rivera, G. Repositioning FDA Drugs as Potential Cruzain Inhibitors from Trypanosoma Cruzi: Virtual Screening, In Vitro and In Vivo Studies. Molecules 2017, 22, 1015. [Google Scholar] [CrossRef]
- Peloso, E.F.; Gonalves, C.C.; Silva, T.M.; Ribeiro, L.H.G.; Piñeyro, M.D.; Robello, C.; Gadelha, F.R. Tryparedoxin Peroxidases and Superoxide Dismutases Expression as Well as ROS Release Are Related to Trypanosoma Cruzi Epimastigotes Growth Phases. Arch. Biochem. Biophys. 2012, 520, 117–122. [Google Scholar] [CrossRef]
- Pérez-Fuentes, R.; Torres-Rasgado, E.; Salgado-Rosas, H.; Zamora-Ginez, I.; Sánchez-Guillén, M.C. The Anti-Oxidant Defence Response in Individuals with the Indeterminate Form of Chagas Disease (American Trypanosomiasis). Ann. Trop. Med. Parasitol. 2008, 102, 189–197. [Google Scholar] [CrossRef]
- Vázquez-Jiménez, L.K.; Moreno-Herrera, A.; Juárez-Saldivar, A.; González-González, A.; Ortiz-Pérez, E.; Paz-González, A.D.; Palos-Pizarro, I.; Ramírez-Moreno, E.; Rivera, G. Recent Advances in the Development of Triose Phosphate Isomerase Inhibitors as Antiprotozoal Agents. Curr. Med. Chem. 2021, 28, 2504–2529. [Google Scholar] [CrossRef]
- Machado-Silva, A.; Cerqueira, P.G.; Grazielle-Silva, V.; Gadelha, F.R.; de Figueiredo Peloso, E.; Teixeira, S.M.R.; Machado, C.R. How Trypanosoma Cruzi Deals with Oxidative Stress: Antioxidant Defence and DNA Repair Pathways. Mutat. Res. Rev. Mutat. Res. 2016, 767, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, K.; Paulino, M.; O. Salas, C.; J. Zarate-Ramos, J.; Vera, B.; Rivera, G. Trypanothione Reductase: A Target for the Development of Anti- Trypanosoma Cruzi Drugs. Mini-Rev. Med. Chem. 2017, 17, 939–946. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Becerra, N.; Kashif, M.; González, M.; Cerecetto, H.; Aguilera, E.; Nogueda-Torres, B.; Chacón-Vargas, K.F.; José Zarate-Ramos, J.; Castillo-Velázquez, U.; et al. In Vitro and in Silico Evaluations of New Aryloxy-1,4-Naphthoquinones as Anti-Trypanosoma Cruzi Agents. Med. Chem. Res. 2020, 29, 665–674. [Google Scholar] [CrossRef]
- Cardoso, M.S.; Reis-Cunha, J.L.; Bartholomeu, D.C. Evasion of the Immune Response by Trypanosoma Cruzi during Acute Infection. Front. Immunol. 2016, 6, 659. [Google Scholar] [CrossRef] [PubMed]
- Battista, T.; Colotti, G.; Ilari, A.; Fiorillo, A. Targeting Trypanothione Reductase, a Key Enzyme in the Redox Trypanosomatid Metabolism, to Develop New Drugs against Leishmaniasis and Trypanosomiases. Molecules 2020, 25, 1924. [Google Scholar] [CrossRef]
- Beig, M.; Oellien, F.; Garoff, L.; Noack, S.; Krauth-Siegel, R.L.; Selzer, P.M. Trypanothione Reductase: A Target Protein for a Combined in Vitro and in Silico Screening Approach. PLoS Negl. Trop. Dis. 2015, 9, e0003773. [Google Scholar] [CrossRef]
- Rivera, G. Quinoxaline 1,4-Di-N-Oxide Derivatives: Are They Unselective or Selective Inhibitors? Mini-Rev. Med. Chem. 2022, 22, 15–25. [Google Scholar] [CrossRef]
- El Newahie, A.M.S.; Ismail, N.S.M.; Abou El Ella, D.A.; Abouzid, K.A.M. Quinoxaline-Based Scaffolds Targeting Tyrosine Kinases and Their Potential Anticancer Activity. Arch. Pharm. 2016, 349, 309–326. [Google Scholar] [CrossRef]
- Rivera, G.; Andrade-Ochoa, S.; Ortega Romero, M.S.; Palos, I.; Monge, A.; Sanchez-Torres, L.E. Ester of Quinoxaline-7-Carboxylate 1,4-Di-N-Oxide as Apoptosis Inductors in K-562 Cell Line: An in Vitro, QSAR and DFT Study. Anti-Cancer Agents Med. Chem. 2017, 17, 682–691. [Google Scholar] [CrossRef]
- Rivera, G.; Ahmad Shah, S.S.; Arrieta-Baez, D.; Palos, I.; Mongue, A.; Sánchez-Torres, L.E. Esters of Quinoxaline 1’4-Di-N-Oxide with Cytotoxic Activity on Tumor Cell Lines Based on NCI-60 Panel. Iran. J. Pharm. Res. 2017, 16, 953–965. [Google Scholar]
- Keri, R.S.; Pandule, S.S.; Budagumpi, S.; Nagaraja, B.M. Quinoxaline and Quinoxaline-1,4-Di-N-Oxides: An Emerging Class of Antimycobacterials. Arch. Pharm. 2018, 351, 1700325. [Google Scholar] [CrossRef] [PubMed]
- Palos, I.; Luna-Herrera, J.; Lara-Ramírez, E.E.; Loera-Piedra, A.; Fernández-Ramírez, E.; Guadalupe Aguilera-Arreola, M.; Paz-González, A.D.; Monge, A.; Wan, B.; Franzblau, S.; et al. Anti-Mycobacterium Tuberculosis Activity of Esters of Quinoxaline 1,4-Di-N-Oxide. Molecules 2018, 23, 1453. [Google Scholar] [CrossRef] [PubMed]
- El-Attar, M.A.Z.; Elbayaa, R.Y.; Shaaban, O.G.; Habib, N.S.; Abdel Wahab, A.E.; Abdelwahab, I.A.; El-Hawash, S.A.M. Design, Synthesis, Antibacterial Evaluation and Molecular Docking Studies of Some New Quinoxaline Derivatives Targeting Dihyropteroate Synthase Enzyme. Bioorg. Chem. 2018, 76, 437–448. [Google Scholar] [CrossRef]
- Patel, N.B.; Patel, J.N.; Purohit, A.C.; Patel, V.M.; Rajani, D.P.; Moo-Puc, R.; Lopez-Cedillo, J.C.; Nogueda-Torres, B.; Rivera, G. In Vitro and in Vivo Assessment of Newer Quinoxaline–Oxadiazole Hybrids as Antimicrobial and Antiprotozoal Agents. Int. J. Antimicrob. Agents 2017, 50, 413–418. [Google Scholar] [CrossRef]
- Estevez, Y.; Quiliano, M.; Burguete, A.; Cabanillas, B.; Zimic, M.; Málaga, E.; Verástegui, M.; Pérez-Silanes, S.; Aldana, I.; Monge, A.; et al. Trypanocidal Properties, Structure–Activity Relationship and Computational Studies of Quinoxaline 1,4-Di-N-Oxide Derivatives. Exp. Parasitol. 2011, 127, 745–751. [Google Scholar] [CrossRef]
- Duque-Montaño, B.E.; Gómez-Caro, L.C.; Sanchez-Sanchez, M.; Monge, A.; Hernández-Baltazar, E.; Rivera, G.; Torres-Angeles, O. Synthesis and in Vitro Evaluation of New Ethyl and Methyl Quinoxaline-7-Carboxylate 1,4-Di-N-Oxide against Entamoeba Histolytica. Bioorg. Med. Chem. 2013, 21, 4550–4558. [Google Scholar] [CrossRef]
- Soto-Sánchez, J.; Caro-Gómez, L.A.; Paz-González, A.D.; Marchat, L.A.; Rivera, G.; Moo-Puc, R.; Arias, D.G.; Ramírez-Moreno, E. Biological Activity of Esters of Quinoxaline-7-Carboxylate 1,4-Di-N-Oxide against E. Histolytica and Their Analysis as Potential Thioredoxin Reductase Inhibitors. Parasitol. Res. 2020, 119, 695–711. [Google Scholar] [CrossRef]
- Chacón-Vargas, K.F.; Andrade-Ochoa, S.; Nogueda-Torres, B.; Juárez-Ramírez, D.C.; Lara-Ramírez, E.E.; Mondragón-Flores, R.; Monge, A.; Rivera, G.; Sánchez-Torres, L.E. Isopropyl Quinoxaline-7-Carboxylate 1,4-Di-N-Oxide Derivatives Induce Regulated Necrosis-like Cell Death on Leishmania (Leishmania) Mexicana. Parasitol. Res. 2018, 117, 45–58. [Google Scholar] [CrossRef]
- Barbosa-Cabrera, E.; Moo-Puc, R.; Monge, A.; Paz-González, A.D.; Bocanegra-García, V.; Rivera, G. In Vitro and in Vivo Evaluation of Quinoxaline 1,4-Di-N-Oxide Against Giardia Lamblia. Lett. Drug Des. Discov. 2020, 17, 428–433. [Google Scholar] [CrossRef]
- Torres, E.; Moreno-Viguri, E.; Galiano, S.; Devarapally, G.; Crawford, P.W.; Azqueta, A.; Arbillaga, L.; Varela, J.; Birriel, E.; di Maio, R.; et al. Novel Quinoxaline 1,4-Di-N-Oxide Derivatives as New Potential Antichagasic Agents. Eur. J. Med. Chem. 2013, 66, 324–334. [Google Scholar] [CrossRef]
- Ancizu, S.; Moreno, E.; Torres, E.; Burguete, A.; Pérez-Silanes, S.; Benítez, D.; Villar, R.; Solano, B.; Marín, A.; Aldana, I.; et al. Heterocyclic-2-Carboxylic Acid (3-Cyano-1,4-Di-N-Oxidequinoxalin-2-Yl)Amide Derivatives as Hits for the Development of Neglected Disease Drugs. Molecules 2009, 14, 2256–2272. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Rocha, J.C.; Sánchez-Torres, L.; Nogueda-Torres, B.; Segura-Cabrera, A.; García-Pérez, C.A.; Bocanegra-García, V.; Palos, I.; Monge, A.; Rivera, G. Anti-Trypanosoma Cruzi and Anti-Leishmanial Activity by Quinoxaline-7-Carboxylate 1,4-Di-N-Oxide Derivatives. Parasitol. Res. 2014, 113, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Chacón-Vargas, K.F.; Nogueda-Torres, B.; Sánchez-Torres, L.E.; Suarez-Contreras, E.; Villalobos-Rocha, J.C.; Torres-Martinez, Y.; Lara-Ramirez, E.E.; Fiorani, G.; Krauth-Siegel, R.L.; Bolognesi, M.L.; et al. Trypanocidal Activity of Quinoxaline 1,4 Di-N-Oxide Derivatives as Trypanothione Reductase Inhibitors. Molecules 2017, 22, 220. [Google Scholar] [CrossRef]
- Pérez-Silanes, S.; Torres, E.; Arbillaga, L.; Varela, J.; Cerecetto, H.; González, M.; Azqueta, A.; Moreno-Viguri, E. Synthesis and Biological Evaluation of Quinoxaline Di-N-Oxide Derivatives with in Vitro Trypanocidal Activity. Bioorg. Med. Chem. Lett. 2016, 26, 903–906. [Google Scholar] [CrossRef]
- Hossain, M.U.; Oany, A.R.; Ahmad, S.A.I.; Hasan, M.A.; Khan, M.A.; Siddikey, M.A.A. Identification of Potential Inhibitor and Enzyme-Inhibitor Complex on Trypanothione Reductase to Control Chagas Disease. Comput. Biol. Chem. 2016, 65, 29–36. [Google Scholar] [CrossRef]
- Khan, M.O.F.; Austin, S.E.; Chan, C.; Yin, H.; Marks, D.; Vaghjiani, S.N.; Kendrick, H.; Yardley, V.; Croft, S.L.; Douglas, K.T. Use of an Additional Hydrophobic Binding Site, the Z Site, in the Rational Drug Design of a New Class of Stronger Trypanothione Reductase Inhibitor, Quaternary Alkylammonium Phenothiazines. J. Med. Chem. 2000, 43, 3148–3156. [Google Scholar] [CrossRef] [PubMed]
- González-Chávez, Z.; Olin-Sandoval, V.; Rodíguez-Zavala, J.S.; Moreno-Sánchez, R.; Saavedra, E. Metabolic Control Analysis of the Trypanosoma Cruzi Peroxide Detoxification Pathway Identifies Tryparedoxin as a Suitable Drug Target. Biochim. Biophys. Acta 2015, 1850, 263–273. [Google Scholar] [CrossRef]
- Gómez-Caro, L.C.; Sánchez-Sánchez, M.; Bocanegra-García, V.; Rivera, G.; Monge, A. Synthesis of Quinoxaline 1,4-Di-n-Oxide Derivatives on Solid Support Using Room Temperature and Microwave-Assisted Solvent-Free Procedures. Quim. Nova 2011, 34, 1147–1151. [Google Scholar] [CrossRef]
- Bosseno, M.F.; Barnabé, C.; Gastélum, E.M.; Lozano Kasten, F.; Ramsey, J.; Espinoza, B.; Brenière, S.F. Predominance of Trypanosoma Cruzi Lineage I in Mexico. J. Clin. Microbiol. 2002, 40, 627–632. [Google Scholar] [CrossRef]
- Brener, Z. Therapeutic activity and criterion of cure on mice experimentally infected with Trypanosoma cruzi. Rev. Inst. Med. Trop. Sao Paulo 1962, 4, 389–396. [Google Scholar]
- Kemmer, G.; Keller, S. Nonlinear Least-Squares Data Fitting in Excel Spreadsheets. Nat. Protoc. 2010, 5, 267–281. [Google Scholar] [CrossRef]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the Scope of the Protein–Ligand Interaction Profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindah, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Polishchuk, P.; Kutlushina, A.; Bashirova, D.; Mokshyna, O.; Madzhidov, T. Virtual Screening Using Pharmacophore Models Retrieved from Molecular Dynamic Simulations. Int. J. Mol. Sci. 2019, 20, 5834. [Google Scholar] [CrossRef] [PubMed]
- Lemkul, J.A. From Proteins to Perturbed Hamiltonians: A Suite of Tutorials for the GROMACS-2018 Molecular Simulation Package [Article v1.0]. Living J. Comput. Mol. Sci. 2019, 1, 5068. [Google Scholar] [CrossRef]
- Comini, M.A.; Dirdjaja, N.; Kaschel, M.; Krauth-Siegel, R.L. Preparative Enzymatic Synthesis of Trypanothione and Trypanothione Analogues. Int. J. Parasitol. 2009, 39, 1059–1062. [Google Scholar] [CrossRef]
- Jockers-Scherübl, M.C.; Schirmer, R.H.; Krauth-Siegel, R.L. Trypanothione Reductase from Trypanosoma Cruzi. Eur. J. Biochem. 1989, 180, 267–272. [Google Scholar] [CrossRef]
- Brown, A.M. A Step-by-Step Guide to Non-Linear Regression Analysis of Experimental Data Using a Microsoft Excel Spreadsheet. Comput. Methods Programs Biomed. 2001, 65, 191–200. [Google Scholar] [CrossRef]
- Juárez-Saldivar, A.; Barbosa-Cabrera, E.; Lara-Ramírez, E.E.; Paz-González, A.D.; Martínez-Vázquez, A.V.; Bocanegra-García, V.; Palos, I.; Campillo, N.E.; Rivera, G. Virtual Screening of Fda-approved Drugs against Triose Phosphate Isomerase from Entamoeba Histolytica and Giardia Lamblia Identifies Inhibitors of Their Trophozoite Growth Phase. Int. J. Mol. Sci. 2021, 22, 5943. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||||
| Code | R2 | R3 | R7 | Trypomastigotes | |||
| % mortality at 50 µg/mL | LC50 (µM) | ||||||
| NINOA | INC-5 | NINOA | INC-5 | ||||
| T-137 | -CH3 | -CH3 | CH3(CH2)3- | 49.5 ± 2.1 | 27.2 ± 4.1 | 118.2 ± 2.2 | 192.6 ± 7.5 |
| T-138 | -OCH3 | -CH3 | CH3(CH2)3- | 58.1 ± 1.3 | 40 ± 3.4 | 87.96 ± 3.6 | 269.5 ± 10 |
| T-139 | -OCH2CH3 | -CH3 | CH3(CH2)3- | 58.2 ± 0.9 | 33.1 ± 1.9 | 96 ± 1.4 | 163 ± 5.2 |
| T-140 | -OC(CH3)3 | -CH3 | CH3(CH2)3- | 45.3 ± 1.7 | 50.8 ± 4.6 | 122 ± 2.1 | 104 ± 5.6 |
| T-141 | -OCH2 C6H5 | -CH3 | CH3(CH2)3- | 71 ± 1.9 | 40.2 ± 3.1 | 38.9 ± 1.4 | 118 ± 6.3 |
| T-142 | -C6H5 | -CH3 | CH3(CH2)3- | 67.6 ± 3.5 | 36.2 ± 4.3 | 44.0 ± 2.6 | 160 ± 6.2 |
| T-143 | -HN-p-C6H4Cl | -CH3 | CH3(CH2)3- | 63.8 ± 3.2 | 37.8 ± 2.3 | 51.1 ± 1.9 | 121 ± 6.3 |
| T-144 | -NH-[2,4(CH3)2C6H3] | -CH3 | CH3(CH2)3- | 57.1 ± 1.8 | 37.4 ± 1.1 | 66.3 ± 4.5 | 122 ± 5.7 |
| T-145 | -CH3 | -CF3 | CH3(CH2)3- | 28.6 ± 5.6 | 45 ± 3.1 | 167 ± 7.8 | 242 ± 7 |
| T-146 | -OCH2CH3 | -CF3 | CH3(CH2)3- | 38.1 ± 1.9 | 48 ± 1.7 | 123 ± 5.2 | 222 ± 8 |
| T-147 | -C(CH3)3 | -CF3 | CH3(CH2)3- | 45.2 ± 2.2 | 48.8 ± 2.4 | 116 ± 5.1 | 124 ± 3.6 |
| T-148 | -C6H5 | -CF3 | CH3(CH2)3- | 56.7 ± 3.7 | 35 ± 1.6 | 65.3 ± 2.1 | 128 ± 4.4 |
| T-149 | -C4H3S | -CF3 | CH3(CH2)3- | 48.1 ± 1.6 | 34.3 ± 2.7 | 96.1 ± 4.1 | 135 ± 3.8 |
| T-150 | -C10H7 | -CF3 | CH3(CH2)3- | 53.3 ± 4.1 | 50.4 ± 1.9 | 64.3 ± 1.7 | 81.5 ± 1.4 |
| T-151 | -NH-C6H5 | -C6H5 | CH3(CH2)3- | 41 ± 0.8 | 30.5 ± 1.8 | 130 ± 5.0 | 103 ± 4.2 |
| T-155 | -CH3 | -CF3 | (CH3)2CHCH2- | 25.2 ± 1.4 | 53.7 ± 4.1 | 186 ± 7.5 | 97.6 ± 4.3 |
| T-156 | -C6H5 | -CF3 | (CH3)2CHCH2- | 26.2 ± 1.1 | 31.2 ± 1.3 | 194 ± 7.1 | 105 ± 3.7 |
| T-157 | -C10H7 | -CF3 | (CH3)2CHCH2- | 25.9 ± 3.1 | 15.3 ± 1.1 | 136 ± 4.3 | 119 ± 3.9 |
| T-158 | -C6H5 | -CH3 | (CH3)2CHCH2- | 24.9 ± 1.4 | 27.2 ± 2.3 | 171 ± 7.6 | 148 ± 3.5 |
| T-159 | -CH3 | -C6H5 | (CH3)2CHCH2- | 9.7 ± 1.5 | 14.9 ± 1.9 | 257 ± 8.5 | 189 ± 5.3 |
| T-161 | -C4H3S | -CF3 | (CH3)2CHCH2- | 46.3 ± 3.4 | 21.4 ± 2.1 | 116 ± 5.7 | 130 ± 5.5 |
| T-162 | -OCH2 C6H5 | -CH3 | (CH3)2CHCH2- | 25.6 ± 1.9 | 30.7 ± 2.8 | 160 ± 7.8 | 117 ± 4.4 |
| T-163 | -NH-[2,4(CH3)2C6H3] | -CH3 | (CH3)2CHCH2- | 27.4 ± 4.2 | 53.5 ± 3.3 | 154 ± 6.4 | 84.5 ± 4.3 |
| T-164 | -CH3 | -NH-[2,4(CH3)2C6H3] | (CH3)2CHCH2- | 8.7 ± 1.2 | 32.7 ± 1.9 | 556 ± 13.2 | 134 ± 4.3 |
| T-165 | -HN-p-C6H4Cl | -CH3 | (CH3)2CHCH2- | 12.5 ± 2.3 | 24.3 ± 0.7 | 157 ± 6.1 | 165 ± 7.2 |
| T-166 | -CH3 | -CH3 | (CH3)2CHCH2- | 45.7 ± 1.1 | 11.8 ± 1.6 | 159 ± 6.0 | 233 ± 8.8 |
| T-167 | -NHC6H5 | -C6H5 | (CH3)2CHCH2- | 61.2 ± 3.9 | 74.8 ± 0.8 | 78.1 ± 2.8 | 136 ± 4.4 |
| T-168 | -C4H3O | -CF3 | (CH3)2CHCH2- | 69.4 ± 5.1 | 55.2 ± 1.3 | 70.0 ± 4.0 | 198 ± 5.7 |
| T-169 | -NHC6H5 | -CH3 | (CH3)2CHCH2- | 78.3 ± 1.7 | 68.1 ± 3.1 | 56.9 ± 0.2 | 62.2 ± 3.3 |
| T-170 | -OCH3 | -CH3 | (CH3)2CHCH2- | 76.9 ± 0.6 | 60.4 ± 0.9 | 33.3 ± 1.2 | 199 ± 10.2 |
| Bnz | 68.8 ± 1.7 | 51.4 ± 1.4 | 130.7 ± 8.8 | 191.3 ± 1.8 | |||
| Nfx | 76.6 ± 1.5 | 53.8 ± 1.3 | 70.4 ± 8.0 | 139.4 ± 3.0 | |||
| 2D-Interaction Profiles | |
|---|---|
 |  |
 |  |
 |  |
 |  |
| Compound | Inhibitor [µM] | % Inhibition of TR | % Inhibition of TR |
|---|---|---|---|
| 44 µM [TS2] | 100 µM [TS2] | ||
| T-147 | 20 | 35 | 28 |
| 5 | 18 | 13 | |
| T-148 | 20 | 36 | 33 |
| 5 | 18 | 16 | |
| T-150 | 20 | insoluble | insoluble |
| 5 | 13 | 0 |
| Substrate GSSG [µM] | % Inhibition of hGR | % Inhibition of hGR |
|---|---|---|
| 5 µM [T-147] | 20 µM [T-147] | |
| 37 | 14 | 45 |
| 92 | 24 | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-González, A.; Sánchez-Sánchez, O.; Krauth-Siegel, R.L.; Bolognesi, M.L.; Gớmez-Escobedo, R.; Nogueda-Torres, B.; Vázquez-Jiménez, L.K.; Saavedra, E.; Encalada, R.; Espinoza-Hicks, J.C.; et al. In Vitro and In Silico Analysis of New n-Butyl and Isobutyl Quinoxaline-7-carboxylate 1,4-di-N-oxide Derivatives against Trypanosoma cruzi as Trypanothione Reductase Inhibitors. Int. J. Mol. Sci. 2022, 23, 13315. https://doi.org/10.3390/ijms232113315
González-González A, Sánchez-Sánchez O, Krauth-Siegel RL, Bolognesi ML, Gớmez-Escobedo R, Nogueda-Torres B, Vázquez-Jiménez LK, Saavedra E, Encalada R, Espinoza-Hicks JC, et al. In Vitro and In Silico Analysis of New n-Butyl and Isobutyl Quinoxaline-7-carboxylate 1,4-di-N-oxide Derivatives against Trypanosoma cruzi as Trypanothione Reductase Inhibitors. International Journal of Molecular Sciences. 2022; 23(21):13315. https://doi.org/10.3390/ijms232113315
Chicago/Turabian StyleGonzález-González, Alonzo, Oscar Sánchez-Sánchez, R. Luise Krauth-Siegel, Maria Laura Bolognesi, Rogelio Gớmez-Escobedo, Benjamín Nogueda-Torres, Lenci K. Vázquez-Jiménez, Emma Saavedra, Rusely Encalada, José Carlos Espinoza-Hicks, and et al. 2022. "In Vitro and In Silico Analysis of New n-Butyl and Isobutyl Quinoxaline-7-carboxylate 1,4-di-N-oxide Derivatives against Trypanosoma cruzi as Trypanothione Reductase Inhibitors" International Journal of Molecular Sciences 23, no. 21: 13315. https://doi.org/10.3390/ijms232113315
APA StyleGonzález-González, A., Sánchez-Sánchez, O., Krauth-Siegel, R. L., Bolognesi, M. L., Gớmez-Escobedo, R., Nogueda-Torres, B., Vázquez-Jiménez, L. K., Saavedra, E., Encalada, R., Espinoza-Hicks, J. C., Paz-González, A. D., & Rivera, G. (2022). In Vitro and In Silico Analysis of New n-Butyl and Isobutyl Quinoxaline-7-carboxylate 1,4-di-N-oxide Derivatives against Trypanosoma cruzi as Trypanothione Reductase Inhibitors. International Journal of Molecular Sciences, 23(21), 13315. https://doi.org/10.3390/ijms232113315