
Knockdown of CLAUDIN-6 Inhibited Apoptosis and Induced Proliferation of Bovine Cumulus Cells

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Expression of CLDN6 Gene and Protein in Bovine CCs
2.2. Screening of Effective siRNA Sequences Targeting Bovine CLDN6 Gene
2.3. Effects of CLDN6 Knockdown on the Apoptosis of Bovine CCs
2.4. Effects of CLDN6 Knockdown on the Proliferation of Bovine CCs
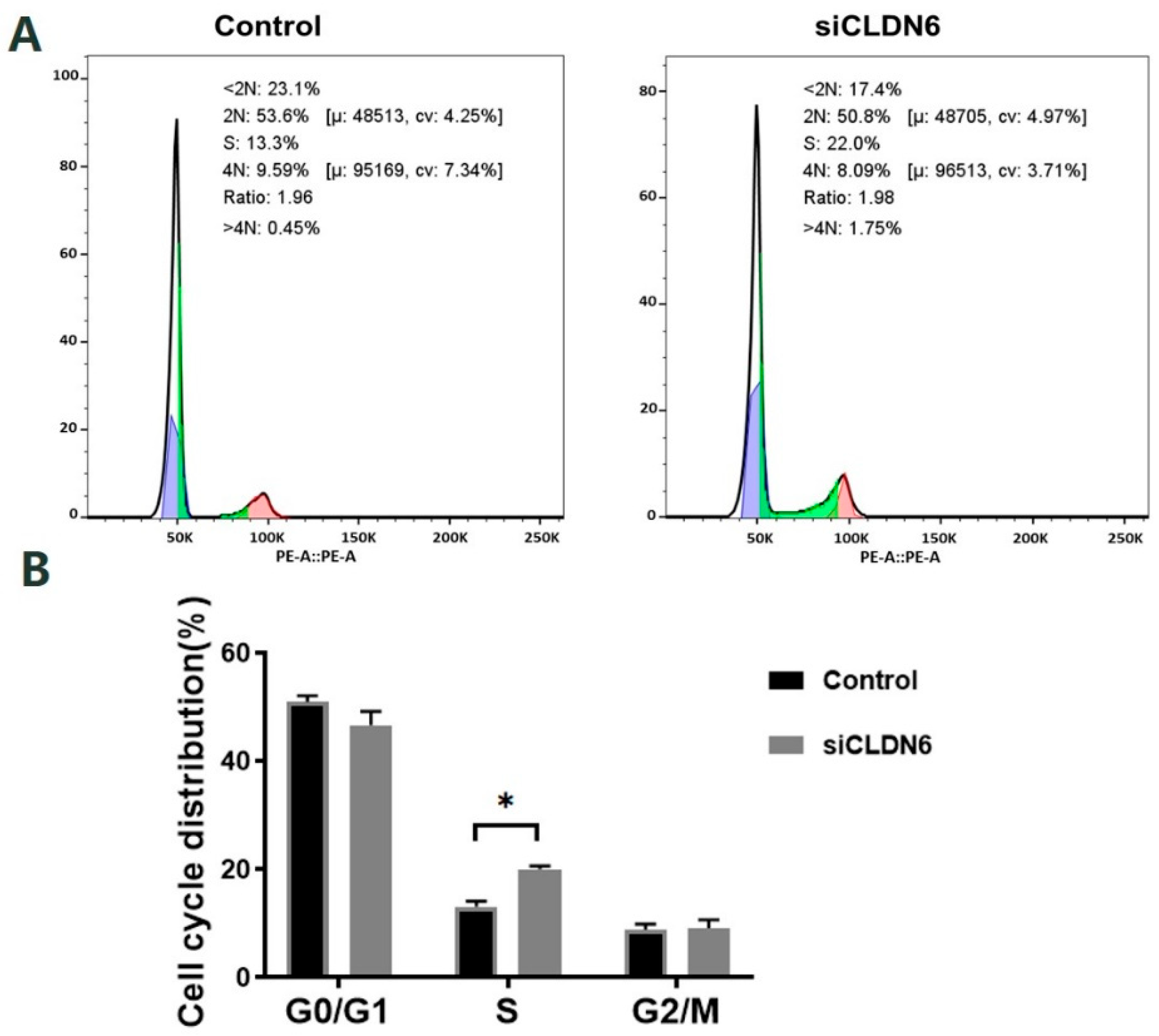
2.5. Effects of CLDN6 Knockdown on the Cell Cycle of Bovine CCs
3. Discussion
4. Materials and Methods
4.1. Culture of Bovine CCs
4.2. Synthesis and Transfection of Small Interfering RNA (siRNA)
4.3. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.4. Immunofluorescence Staining
4.5. Western Blotting (WB)
4.6. Apoptosis Analysis
4.7. Cell Proliferation Assay
4.8. Cell Cycle Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diaz, F.J.; O’Brien, M.J.; Wigglesworth, K.; Eppig, J.J. The preantral granulosa cell to cumulus cell transition in the mouse ovary: Development of competence to undergo expansion. Dev. Biol. 2006, 299, 91–104. [Google Scholar] [CrossRef]
- Auclair, S.; Uzbekov, R.; Elis, S.; Sanchez, L.; Kireev, I.; Lardic, L.; Dalbies-Tran, R.; Uzbekova, S. Absence of cumulus cells during in vitro maturation affects lipid metabolism in bovine oocytes. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E599–E613. [Google Scholar] [CrossRef] [PubMed]
- Hughes, F.M., Jr.; Gorospe, W.C. Biochemical identification of apoptosis (programmed cell death) in granulosa cells: Evidence for a potential mechanism underlying follicular atresia. Endocrinology 1991, 129, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Elgebaly, M.M.; Hazaa, A.B.M.; Amer, H.A.; Mesalam, A. L-Cysteine improves bovine oocyte developmental competence in vitro via activation of oocyte-derived growth factors BMP-15 and GDF-9. Reprod. Domest. Anim. 2022, 57, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Schutz, L.F.; Robinson, C.L.; Totty, M.L.; Spicer, L.J. Evidence that gene expression of ovarian follicular tight junction proteins is regulated in vivo and in vitro in cattle. J. Anim. Sci. 2017, 95, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.L.; Singh, K.; Mangray, S.; Tavares, R.; Noble, L.; Resnick, M.B.; Yakirevich, E. Claudin expression in high-grade invasive ductal carcinoma of the breast: Correlation with the molecular subtype. Mod. Pathol. 2013, 26, 485–495. [Google Scholar] [CrossRef]
- Gunzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef]
- Wang, H.F.; Zheng, S.F.; Chen, Y.; Zhou, Z.Y.; Xu, J. Correlations between claudin-1 and PIGF expressions in retinoblastoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4196–4203. [Google Scholar] [PubMed]
- Schaefer, I.M.; Agaimy, A.; Fletcher, C.D.; Hornick, J.L. Claudin-4 expression distinguishes SWI/SNF complex-deficient undifferentiated carcinomas from sarcomas. Mod. Pathol. 2017, 30, 539–548. [Google Scholar] [CrossRef]
- Abuazza, G.; Becker, A.; Williams, S.S.; Chakravarty, S.; Truong, H.T.; Lin, F.; Baum, M. Claudins 6, 9, and 13 are developmentally expressed renal tight junction proteins. Am. J. Physiol. Renal Physiol. 2006, 291, F1132–F1141. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, T.; Sherman-Baust, C.A.; Poosala, S.; Mullin, J.M.; Morin, P.J. Age-related changes of claudin expression in mouse liver, kidney, and pancreas. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 1146–1153. [Google Scholar] [CrossRef]
- Ben-David, U.; Nudel, N.; Benvenisty, N. Immunologic and chemical targeting of the tight-junction protein Claudin-6 eliminates tumorigenic human pluripotent stem cells. Nat. Commun. 2013, 4, 1992. [Google Scholar] [CrossRef]
- Wang, L.L.; Xue, Y.; Shen, Y.H.; Li, W.; Cheng, Y.; Yan, X.S.; Shi, W.S.; Wang, J.; Gong, Z.J.; Yang, G.H.; et al. Claudin 6: A novel surface marker for characterizing mouse pluripotent stem cells. Cell Res. 2012, 22, 1082–1085. [Google Scholar] [CrossRef] [PubMed]
- Troy, T.C.; Turksen, K. The targeted overexpression of a Claudin mutant in the epidermis of transgenic mice elicits striking epidermal and hair follicle abnormalities. Mol. Biotechnol. 2007, 36, 166–174. [Google Scholar] [CrossRef]
- Moriwaki, K.; Tsukita, S.; Furuse, M. Tight junctions containing claudin 4 and 6 are essential for blastocyst formation in preimplantation mouse embryos. Dev. Biol. 2007, 312, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, X.L.; Li, C.C.; Mao, B.Y. Xenopus Claudin-6 is required for embryonic pronephros morphogenesis and terminal differentiation. Biochem. Biophys. Res. Commun. 2015, 462, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Yang, X.Y.; Fan, J.J.; Du, X.B. Claudin 6: Therapeutic prospects for tumours, and mechanisms of expression and regulation (Review). Mol. Med. Rep. 2021, 24, 677. [Google Scholar] [CrossRef]
- Ito, Y.; Takasawa, A.; Takasawa, K.; Murakami, T.; Akimoto, T.; Kyuno, D.; Kawata, Y.; Shano, K.; Kirisawa, K.; Ota, M.; et al. Aberrant expression of claudin-6 contributes to malignant potentials and drug resistance of cervical adenocarcinoma. Cancer Sci. 2022, 113, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.T.; Zhang, Y.Q.; Li, Q.L.; Zhang, Z.Z.; Zhao, G.; Xu, J. CLDN6 promotes tumor progression through the YAP1-snail1 axis in gastric cancer. Cell Death Dis. 2019, 10, 949. [Google Scholar] [CrossRef]
- Cao, X.; He, G.Z. Knockdown of CLDN6 inhibits cell proliferation and migration via PI3K/AKT/mTOR signaling pathway in endometrial carcinoma cell line HEC-1-B. Onco Targets Ther. 2018, 11, 6351–6360. [Google Scholar] [CrossRef]
- Kwon, M.J. Emerging roles of claudins in human cancer. Int. J. Mol. Sci. 2013, 14, 18148–18180. [Google Scholar] [CrossRef]
- Lewis, J.B.; Milner, D.C.; Lewis, A.L.; Dunaway, T.M.; Egbert, K.M.; Albright, S.C.; Merrell, B.J.; Monson, T.D.; Broberg, D.S.; Gassman, J.R.; et al. Up-regulation of Claudin-6 in the distal lung impacts secondhand smoke-induced inflammation. Int. J. Environ. Res. Public Health 2016, 13, 1018. [Google Scholar] [CrossRef]
- Sugimoto, K.; Ichikawa-Tomikawa, N.; Satohisa, S.; Akashi, Y.; Kanai, R.; Saito, T.; Sawada, N.; Chiba, H. The tight-junction protein claudin-6 induces epithelial differentiation from mouse F9 and embryonic stem cells. PLoS ONE 2013, 8, e75106. [Google Scholar] [CrossRef]
- Kohmoto, T.; Masuda, K.; Shoda, K.; Takahashi, R.; Ujiro, S.; Tange, S.; Ichikawa, D.; Otsuji, E.; Imoto, I. Claudin-6 is a single prognostic marker and functions as a tumor-promoting gene in a subgroup of intestinal type gastric cancer. Gastric Cancer 2020, 23, 403–417. [Google Scholar] [CrossRef]
- Song, P.Y.; Li, Y.R.; Dong, Y.; Liang, Y.Y.; Qu, H.N.; Qi, D.; Lu, Y.; Jin, X.S.; Guo, Y.T.; Jia, Y.Y.; et al. Estrogen receptor beta inhibits breast cancer cells migration and invasion through CLDN6-mediated autophagy. J. Exp. Clin. Cancer Res. 2019, 38, 354. [Google Scholar] [CrossRef]
- Birks, D.K.; Kleinschmidt-DeMasters, B.K.; Donson, A.M.; Barton, V.N.; McNatt, S.A.; Foreman, N.K.; Handler, M.H. Claudin 6 is a positive marker for atypical teratoid/rhabdoid tumors. Brain Pathol. 2010, 20, 140–150. [Google Scholar] [CrossRef]
- Antonelli, M.; Hasselblatt, M.; Haberler, C.; Di Giannatale, A.; Garre, M.L.; Donofrio, V.; Lauriola, L.; Ridola, V.; Arcella, A.; Fruhwald, M.; et al. Claudin-6 is of limited sensitivity and specificity for the diagnosis of atypical teratoid/rhabdoid tumors. Brain Pathol. 2011, 21, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Sharma, A.; Dhawan, P. Claudin family of proteins and cancer: An overview. J. Oncol. 2010, 2010, 541957. [Google Scholar] [CrossRef]
- Arabzadeh, A.; Troy, T.C.; Turksen, K. Role of the Cldn6 cytoplasmic tail domain in membrane targeting and epidermal differentiation in vivo. Mol. Cell Biol. 2006, 26, 5876–5887. [Google Scholar] [CrossRef]
- Sullivan, L.M.; Yankovich, T.; Le, P.; Martinez, D.; Santi, M.; Biegel, J.A.; Pawel, B.R.; Judkins, A.R. Claudin-6 is a nonspecific marker for malignant rhabdoid and other pediatric tumors. Am. J. Surg. Pathol. 2012, 36, 73–80. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, Q.; Liu, Y.F.; Xu, X.M.; Quan, C.S. Gene silencing of claudin-6 enhances cell proliferation and migration accompanied with increased MMP-2 activity via p38 MAPK signaling pathway in human breast epithelium cell line HBL-100. Mol. Med. Rep. 2013, 8, 1505–1510. [Google Scholar] [CrossRef]
- Zhang, X.W.; Ruan, Y.; Li, Y.R.; Lin, D.J.; Liu, Z.J.; Quan, C.S. Expression of apoptosis signal-regulating kinase 1 is associated with tight junction protein claudin-6 in cervical carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 5535–5541. [Google Scholar]
- Lu, Y.; Dang, Q.H.; Bo, Y.; Su, X.J.; Wang, L.P.; Sun, J.Q.; Wei, J.Y.; Quan, C.S.; Li, Y.R. The expression of CLDN6 in hepatocellular carcinoma tissue and the effects of CLDN6 on biological phenotypes of hepatocellular carcinoma cells. J. Cancer 2021, 12, 5454–5463. [Google Scholar] [CrossRef]
- Guo, Y.X.; Xu, X.M.; Liu, Z.J.; Zhang, T.; Zhang, X.W.; Wang, L.P.; Wang, M.; Liu, Y.Y.; Lu, Y.; Liu, Y.P.; et al. Apoptosis signal-regulating kinase 1 is associated with the effect of claudin-6 in breast cancer. Diagn. Pathol. 2012, 7, 111. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.R.; Wu, S.; Tang, J.H.; Chen, G.G.; Li, F.G. miR-135a suppresses granulosa cell growth by targeting Tgfbr1 and Ccnd2 during folliculogenesis in mice. Cells 2021, 10, 2104. [Google Scholar] [CrossRef]
- Robker, R.L.; Richards, J.S. Hormone-induced proliferation and differentiation of granulosa cells: A coordinated balance of the cell cycle regulators cyclin D2 and p27Kip1. Mol. Endocrinol. 1998, 12, 924–940. [Google Scholar] [CrossRef] [PubMed]
- Mei, Q.J.; Li, H.Y.; Liu, Y.; Wang, X.F.; Xiang, W.P. Advances in the study of CDC42 in the female reproductive system. J. Cell Mol. Med. 2022, 26, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Yang, Y.F.; Wang, Y.L.; Li, Y.; He, Y.M.; Duan, J.X.; Xu, D.J.; Pei, Y.F.; Cheng, J.Y.; Yang, L.; et al. Phospholipase C inhibits apoptosis of porcine primary granulosa cells cultured in vitro. J. Ovarian Res. 2019, 12, 90. [Google Scholar] [CrossRef]
- Xu, R.F.; Qin, N.; Xu, X.X.; Sun, X.; Chen, X.X.; Zhao, J.H. Inhibitory effect of SLIT2 on granulosa cell proliferation mediated by the CDC42-PAKs-ERK1/2 MAPK pathway in the prehierarchical follicles of the chicken ovary. Sci. Rep. 2018, 8, 9168. [Google Scholar] [CrossRef]
- Scott, R.P.; Hawley, S.P.; Ruston, J.; Du, J.M.; Brakebusch, C.; Jones, N.; Pawson, T. Podocyte-specific loss of Cdc42 leads to congenital nephropathy. J. Am. Soc. Nephrol. 2012, 23, 1149–1154. [Google Scholar] [CrossRef]
- Wallace, S.W.; Durgan, J.; Jin, D.; Hall, A. Cdc42 regulates apical junction formation in human bronchial epithelial cells through PAK4 and Par6B. Mol. Biol. Cell 2010, 21, 2996–3006. [Google Scholar] [CrossRef]
- Cardano, M.; Tribioli, C.; Prosperi, E. Targeting proliferating cell nuclear antigen (PCNA) as an effective strategy to inhibit tumor cell proliferation. Curr. Cancer Drug Targets 2020, 20, 240–252. [Google Scholar] [CrossRef]
- Celis, J.E.; Celis, A. Cell cycle-dependent variations in the distribution of the nuclear protein cyclin proliferating cell nuclear antigen in cultured cells: Subdivision of S phase. Proc. Natl. Acad. Sci. USA 1985, 82, 3262–3266. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Hua, J.; Zhang, Y.W.; Jiang, X.H.; Zhang, H.; Ma, T.L.; Zheng, W.; Sun, R.; Shen, W.; Sha, J.H.; et al. Proliferating cell nuclear antigen (PCNA) regulates primordial follicle assembly by promoting apoptosis of oocytes in fetal and neonatal mouse ovaries. PLoS ONE 2011, 6, e16046. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, Y.F.; Ren, Y.; Xu, X.M.; Yu, L.N.; Li, Y.L.; Quan, C.S. Tight junction protein, claudin-6, downregulates the malignant phenotype of breast carcinoma. Eur. J. Cancer Prev. 2010, 19, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Y.; Zhu, F.F.; Chen, P.X. miR-7 and miR-218 epigenetically control tumor suppressor genes RASSF1A and Claudin-6 by targeting HoxB3 in breast cancer. Biochem. Biophys. Res. Commun. 2012, 424, 28–33. [Google Scholar] [CrossRef]
- Sun, W.J.; Pang, Y.W.; Liu, Y.; Hao, H.S.; Zhao, X.M.; Qin, T.; Zhu, H.B.; Du, W.H. Exogenous glutathione supplementation in culture medium improves the bovine embryo development after in vitro fertilization. Theriogenology 2015, 84, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.Q.; Li, X.L.; Wang, W.J.; Hao, H.S.; Zou, H.Y.; Pang, Y.W.; Zhao, X.M.; Zhu, H.B.; Du, W.H. Knockdown of bone morphogenetic protein 4 gene induces apoptosis and inhibits proliferation of bovine cumulus cells. Theriogenology 2022, 188, 28–36. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA | Sequences (5′–3′) | |
|---|---|---|
| siRNA-1 | Sense Antisense | GCUUCCGCUGGUCUGCAAATT UUUGCAGACCAGCGGAAGCTT |
| siRNA-2 | Sense Antisense | GGAAGGUGACCGCCUUCAUTT AUGAAGGCGGUCACCUUCCTT |
| siRNA-3 | Sense Antisense | GGACCGCCCACACCAUCAUTT AUGAUGGUGUGGGCGGUCCTT |
| Negative control (NC) | Sense Antisense | UUCUCCGAACGUGUCACGUTT ACGUGACACGUUCGGAGAATT |
| Genes | Accession No. | Primer Sequences (5′–3′) | Product Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| CLDN6 | NM_001205697.1 | F: GCUUCCGCUGGUCUGCAAATT | 192 | 59 |
| R: UUUGCAGACCAGCGGAAGCTT | ||||
| BAX | NM_173894 | F:GGCTGGACATTGGACTTCCTTC | 112 | 61 |
| R:TGGTCACTGTCTGCCATGTGG | ||||
| BCL-2 | NM_001077486 | F:GATGACTTCTCTCGGCGCTA | 165 | 60 |
| R:GACCCCTCCGAACTCAAAGA | ||||
| CASPASE-3 | NM_001077840 | F:TACTTGGGAAGGTGTGAGAAAACTAA | 71 | 59 |
| R:AACCCGTCTCCCTTTATATTGCT | ||||
| PCNA | NM_001034494 | F:GTCCAGGGCTCCATCTTGAA | 123 | 58 |
| R:CAAGGAGACATGAGACGAGT | ||||
| CDC42 | NM_001046332 | F:GTTGTTGTGGGTGATGGTGC | 128 | 58 |
| R:TCCCCACCAATCATAACTGT | ||||
| CCND2 | NM_001076372 | F:TGACCGCTGAGAAGTTATGC | 104 | 59 |
| R:CGCCAGGTTCCATTTCAACT | ||||
| GAPDH | NM-001034034.2 | F:GGGTCATCATCTCTGCACCT | 177 | 59 |
| R:GGTCATAAGTCCCTCCACGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Zou, H.; Chen, N.; Tian, Y.; Hao, H.; Pang, Y.; Zhao, X.; Zhu, H.; Yu, D.; Du, W. Knockdown of CLAUDIN-6 Inhibited Apoptosis and Induced Proliferation of Bovine Cumulus Cells. Int. J. Mol. Sci. 2022, 23, 13222. https://doi.org/10.3390/ijms232113222
Wang W, Zou H, Chen N, Tian Y, Hao H, Pang Y, Zhao X, Zhu H, Yu D, Du W. Knockdown of CLAUDIN-6 Inhibited Apoptosis and Induced Proliferation of Bovine Cumulus Cells. International Journal of Molecular Sciences. 2022; 23(21):13222. https://doi.org/10.3390/ijms232113222
Chicago/Turabian StyleWang, Wanjie, Huiying Zou, Nanzhu Chen, Yaqing Tian, Haisheng Hao, Yunwei Pang, Xueming Zhao, Huabin Zhu, Dawei Yu, and Weihua Du. 2022. "Knockdown of CLAUDIN-6 Inhibited Apoptosis and Induced Proliferation of Bovine Cumulus Cells" International Journal of Molecular Sciences 23, no. 21: 13222. https://doi.org/10.3390/ijms232113222
APA StyleWang, W., Zou, H., Chen, N., Tian, Y., Hao, H., Pang, Y., Zhao, X., Zhu, H., Yu, D., & Du, W. (2022). Knockdown of CLAUDIN-6 Inhibited Apoptosis and Induced Proliferation of Bovine Cumulus Cells. International Journal of Molecular Sciences, 23(21), 13222. https://doi.org/10.3390/ijms232113222