
PCSK9 Confers Inflammatory Properties to Extracellular Vesicles Released by Vascular Smooth Muscle Cells
 , , , , , ,
, , , , , ,  ,
,  ,
, 
 , ,
, ,  , and
, and 
Abstract
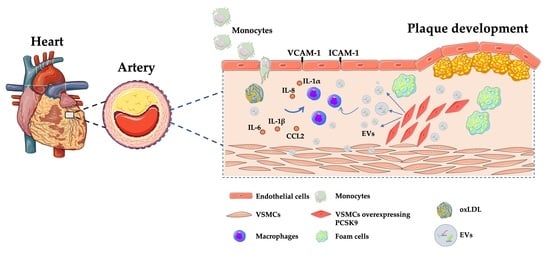
1. Introduction
2. Results
2.1. Characterization of VSMCs Overexpressing PCSK9
2.2. Characterization of VSMCPCSK9-Derived EVs
2.3. Impact of EVs on Pro-Inflammatory Markers in Endothelial Cells, Monocytes and Monocyte-Derived Macrophages
2.4. Effect of EVs on Cell Migration, ox-LDL Uptake and Mitochondrial Functionality in Recipient Cells
2.5. Evaluation of Macrophage Recruitment upon EVs Injection in Zebrafish
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Extracellular Vesicles Isolation
4.3. Evaluation of the Uptake of EVs
4.4. Nanosight Tracking Analysis (NTA)
4.5. Transmission Electron Microscopy (TEM)
4.6. qPCR
4.7. Western Blot
4.8. Enzyme-Linked Immunosorbent Assay
4.9. Flow Cytometry
4.10. Migration Assay
4.11. Preparation of Oxidized LDL (oxLDL) and Lipoprotein-Induced Cholesterol Accumulation
4.12. Mitochondrial Bioenergetic Evaluation
4.13. Mass Spectrometry Analysis
4.14. Gene Ontology Analysis
4.15. EV-miRNAs Isolation and Analysis
4.16. Zebrafish Husbandry
4.17. Microinjection of Zebrafish Embryos and Immunostaining Analysis
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgozoglu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Prim. 2019, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Bjorkegren, J.L.M.; Lusis, A.J. Atherosclerosis: Recent developments. Cell 2022, 185, 1630–1645. [Google Scholar] [CrossRef] [PubMed]
- Basatemur, G.L.; Jorgensen, H.F.; Clarke, M.C.H.; Bennett, M.R.; Mallat, Z. Vascular smooth muscle cells in atherosclerosis. Nat. Rev. Cardiol. 2019, 16, 727–744. [Google Scholar] [CrossRef]
- Grootaert, M.O.J.; Bennett, M.R. Vascular smooth muscle cells in atherosclerosis: Time for a re-assessment. Cardiovasc. Res. 2021, 117, 2326–2339. [Google Scholar] [CrossRef] [PubMed]
- Lupo, M.G.; Bressan, A.; Donato, M.; Canzano, P.; Camera, M.; Poggio, P.; Greco, M.F.; Garofalo, M.; De Martin, S.; Panighel, G.; et al. PCSK9 promotes arterial medial calcification. Atherosclerosis 2022, 346, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 2019, 43, 273–303. [Google Scholar] [CrossRef]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating Extracellular Vesicles in Human Disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Guay, C.; Regazzi, R. Exosomes as new players in metabolic organ cross-talk. Diabetes Obes. Metab. 2017, 19 (Suppl. 1), 137–146. [Google Scholar] [CrossRef] [PubMed]
- Konkoth, A.; Saraswat, R.; Dubrou, C.; Sabatier, F.; Leroyer, A.S.; Lacroix, R.; Duchez, A.C.; Dignat-George, F. Multifaceted role of extracellular vesicles in atherosclerosis. Atherosclerosis 2021, 319, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Giro, O.; Jimenez, A.; Pane, A.; Badimon, L.; Ortega, E.; Chiva-Blanch, G. Extracellular vesicles in atherothrombosis and cardiovascular disease: Friends and foes. Atherosclerosis 2021, 330, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Macchi, C.; Greco, M.F.; Favero, C.; Dioni, L.; Cantone, L.; Hoxha, M.; Vigna, L.; Solazzo, G.; Corsini, A.; Banach, M.; et al. Associations Among PCSK9 Levels, Atherosclerosis-Derived Extracellular Vesicles, and Their miRNA Content in Adults With Obesity. Front. Cardiovasc. Med. 2021, 8, 785250. [Google Scholar] [CrossRef] [PubMed]
- Macchi, C.; Ferri, N.; Sirtori, C.R.; Corsini, A.; Banach, M.; Ruscica, M. Proprotein Convertase Subtilisin/Kexin Type 9: A View beyond the Canonical Cholesterol-Lowering Impact. Am. J. Pathol. 2021, 191, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Seidah, N.G.; Garcon, D. Expanding Biology of PCSK9: Roles in Atherosclerosis and Beyond. Curr. Atheroscler. Rep. 2022, 24, 821–830. [Google Scholar] [CrossRef]
- Ferri, N.; Tibolla, G.; Pirillo, A.; Cipollone, F.; Mezzetti, A.; Pacia, S.; Corsini, A.; Catapano, A.L. Proprotein convertase subtilisin kexin type 9 (PCSK9) secreted by cultured smooth muscle cells reduces macrophages LDLR levels. Atherosclerosis 2012, 220, 381–386. [Google Scholar] [CrossRef]
- Ding, Z.; Liu, S.; Wang, X.; Deng, X.; Fan, Y.; Sun, C.; Wang, Y.; Mehta, J.L. Hemodynamic shear stress via ROS modulates PCSK9 expression in human vascular endothelial and smooth muscle cells and along the mouse aorta. Antioxid. Redox Signal 2015, 22, 760–771. [Google Scholar] [CrossRef]
- Ferri, N.; Marchiano, S.; Tibolla, G.; Baetta, R.; Dhyani, A.; Ruscica, M.; Uboldi, P.; Catapano, A.L.; Corsini, A. PCSK9 knock-out mice are protected from neointimal formation in response to perivascular carotid collar placement. Atherosclerosis 2016, 253, 214–224. [Google Scholar] [CrossRef]
- Guo, Y.; Tang, Z.; Yan, B.; Yin, H.; Tai, S.; Peng, J.; Cui, Y.; Gui, Y.; Belke, D.; Zhou, S.; et al. PCSK9 (Proprotein Convertase Subtilisin/Kexin Type 9) Triggers Vascular Smooth Muscle Cell Senescence and Apoptosis: Implication of Its Direct Role in Degenerative Vascular Disease. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 67–86. [Google Scholar] [CrossRef]
- Comfort, N.; Cai, K.; Bloomquist, T.R.; Strait, M.D.; Ferrante, A.W., Jr.; Baccarelli, A.A. Nanoparticle Tracking Analysis for the Quantification and Size Determination of Extracellular Vesicles. J. Vis. Exp. 2021, 169, e62447. [Google Scholar] [CrossRef]
- Boyer, M.J.; Kimura, Y.; Akiyama, T.; Baggett, A.Y.; Preston, K.J.; Scalia, R.; Eguchi, S.; Rizzo, V. Endothelial cell-derived extracellular vesicles alter vascular smooth muscle cell phenotype through high-mobility group box proteins. J. Extracell. Vesicles 2020, 9, 1781427. [Google Scholar] [CrossRef]
- Chen, Q.; Lv, J.; Yang, W.; Xu, B.; Wang, Z.; Yu, Z.; Wu, J.; Yang, Y.; Han, Y. Targeted inhibition of STAT3 as a potential treatment strategy for atherosclerosis. Theranostics 2019, 9, 6424–6442. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Mazza, E.M.; Lucchesi, D.; Cifola, I.; Gemelli, C.; Grande, A.; Battaglia, C.; Bicciato, S.; Boraschi, D. Transcriptomic profiling of the development of the inflammatory response in human monocytes in vitro. PLoS ONE 2014, 9, e87680. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Wang, X.; Zhao, M.; Cai, T.; Liu, P.; Li, J.; Willard, B.; Zu, L.; Zhou, E.; Li, Y.; et al. Macrophage Foam Cell-Derived Extracellular Vesicles Promote Vascular Smooth Muscle Cell Migration and Adhesion. J. Am. Heart Assoc. 2016, 5, e004099. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Ren, K.D.; Luan, Y.; Chen, X.; Yang, Y. Mitochondrial Dynamics: Pathogenesis and Therapeutic Targets of Vascular Diseases. Front. Cardiovasc. Med. 2021, 8, 770574. [Google Scholar] [CrossRef]
- Robertson, T.F.; Huttenlocher, A. Real-time imaging of inflammation and its resolution: It’s apparent because it’s transparent. Immunol. Rev. 2022, 306, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Chi, M.; Phan, Q.T.; Gonzalez, C.; Dubremetz, J.F.; Levraud, J.P.; Lutfalla, G. Transient infection of the zebrafish notochord with E. coli induces chronic inflammation. Dis. Model Mech. 2014, 7, 871–882. [Google Scholar] [CrossRef]
- Ashraf, J.V.; Al Haj Zen, A. Role of Vascular Smooth Muscle Cell Phenotype Switching in Arteriogenesis. Int. J. Mol. Sci. 2021, 22, 10585. [Google Scholar] [CrossRef]
- Ding, Z.; Liu, S.; Wang, X.; Deng, X.; Fan, Y.; Shahanawaz, J.; Shmookler Reis, R.J.; Varughese, K.I.; Sawamura, T.; Mehta, J.L. Cross-talk between LOX-1 and PCSK9 in vascular tissues. Cardiovasc. Res. 2015, 107, 556–567. [Google Scholar] [CrossRef]
- Grune, J.; Meyborg, H.; Bezhaeva, T.; Kappert, K.; Hillmeister, P.; Kintscher, U.; Pieske, B.; Stawowy, P. PCSK9 regulates the chemokine receptor CCR2 on monocytes. Biochem. Biophys. Res. Commun. 2017, 485, 312–318. [Google Scholar] [CrossRef]
- Zou, Y.; Chen, Z.; Zhang, X.; Yu, J.; Xu, H.; Cui, J.; Li, Y.; Niu, Y.; Zhou, C.; Xia, J.; et al. Targeting PCSK9 Ameliorates Graft Vascular Disease in Mice by Inhibiting NLRP3 Inflammasome Activation in Vascular Smooth Muscle Cells. Front. Immunol. 2022, 13, 894789. [Google Scholar] [CrossRef]
- Iida, Y.; Tanaka, H.; Sano, H.; Suzuki, Y.; Shimizu, H.; Urano, T. Ectopic Expression of PCSK9 by Smooth Muscle Cells Contributes to Aortic Dissection. Ann. Vasc. Surg. 2018, 48, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Akbulut, A.C.; Kaczor, D.M.; Halder, M.; Koenen, R.R.; Kramann, R. Initiation and Propagation of Vascular Calcification Is Regulated by a Concert of Platelet- and Smooth Muscle Cell-Derived Extracellular Vesicles. Front. Cardiovasc. Med. 2018, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Suades, R.; Greco, M.F.; Padro, T.; Badimon, L. Extracellular Vesicles as Drivers of Immunoinflammation in Atherothrombosis. Cells 2022, 11, 1845. [Google Scholar] [CrossRef] [PubMed]
- Coly, P.M.; Boulanger, C.M. Role of extracellular vesicles in atherosclerosis: An update. J. Leukoc. Biol. 2022, 111, 51–62. [Google Scholar] [CrossRef]
- Lupo, M.G.; Marchiano, S.; Adorni, M.P.; Zimetti, F.; Ruscica, M.; Greco, M.F.; Corsini, A.; Ferri, N. PCSK9 Induces Rat Smooth Muscle Cell Proliferation and Counteracts the Pleiotropic Effects of Simvastatin. Int. J. Mol. Sci. 2021, 22, 4114. [Google Scholar] [CrossRef]
- Gialeli, C.; Shami, A.; Goncalves, I. Extracellular matrix: Paving the way to the newest trends in atherosclerosis. Curr. Opin. Lipidol. 2021, 32, 277–285. [Google Scholar] [CrossRef]
- Punch, E.; Klein, J.; Diaba-Nuhoho, P.; Morawietz, H.; Garelnabi, M. Effects of PCSK9 Targeting: Alleviating Oxidation, Inflammation, and Atherosclerosis. J. Am. Heart Assoc. 2022, 11, e023328. [Google Scholar] [CrossRef]
- Lu, Y.; Thavarajah, T.; Gu, W.; Cai, J.; Xu, Q. Impact of miRNA in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e159–e170. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; Garcia-Cardena, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef]
- Li, M.; Qian, M.; Kyler, K.; Xu, J. Endothelial-Vascular Smooth Muscle Cells Interactions in Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 151. [Google Scholar] [CrossRef]
- Cai, Q.; Lanting, L.; Natarajan, R. Interaction of monocytes with vascular smooth muscle cells regulates monocyte survival and differentiation through distinct pathways. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Alexander, Y.; Osto, E.; Schmidt-Trucksass, A.; Shechter, M.; Trifunovic, D.; Duncker, D.J.; Aboyans, V.; Back, M.; Badimon, L.; Cosentino, F.; et al. Endothelial function in cardiovascular medicine: A consensus paper of the European Society of Cardiology Working Groups on Atherosclerosis and Vascular Biology, Aorta and Peripheral Vascular Diseases, Coronary Pathophysiology and Microcirculation, and Thrombosis. Cardiovasc. Res. 2021, 117, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Krauss, R.M.; Chang, J.T.; Teng, B.B. PCSK9 deficiency reduces atherosclerosis, apolipoprotein B secretion, and endothelial dysfunction. J. Lipid Res. 2018, 59, 207–223. [Google Scholar] [CrossRef]
- Bernelot Moens, S.J.; Neele, A.E.; Kroon, J.; van der Valk, F.M.; Van den Bossche, J.; Hoeksema, M.A.; Hoogeveen, R.M.; Schnitzler, J.G.; Baccara-Dinet, M.T.; Manvelian, G.; et al. PCSK9 monoclonal antibodies reverse the pro-inflammatory profile of monocytes in familial hypercholesterolaemia. Eur. Heart J. 2017, 38, 1584–1593. [Google Scholar] [CrossRef]
- Ding, Z.; Pothineni, N.V.K.; Goel, A.; Luscher, T.F.; Mehta, J.L. PCSK9 and inflammation: Role of shear stress, pro-inflammatory cytokines, and LOX-1. Cardiovasc. Res. 2020, 116, 908–915. [Google Scholar] [CrossRef]
- Ding, Z.; Wang, X.; Liu, S.; Zhou, S.; Kore, R.A.; Mu, S.; Deng, X.; Fan, Y.; Mehta, J.L. NLRP3 inflammasome via IL-1beta regulates PCSK9 secretion. Theranostics 2020, 10, 7100–7110. [Google Scholar] [CrossRef]
- Dutzmann, J.; Daniel, J.M.; Bauersachs, J.; Hilfiker-Kleiner, D.; Sedding, D.G. Emerging translational approaches to target STAT3 signalling and its impact on vascular disease. Cardiovasc. Res. 2015, 106, 365–374. [Google Scholar] [CrossRef]
- Babon, J.J.; Kershaw, N.J.; Murphy, J.M.; Varghese, L.N.; Laktyushin, A.; Young, S.N.; Lucet, I.S.; Norton, R.S.; Nicola, N.A. Suppression of cytokine signaling by SOCS3: Characterization of the mode of inhibition and the basis of its specificity. Immunity 2012, 36, 239–250. [Google Scholar] [CrossRef]
- Ruscica, M.; Ricci, C.; Macchi, C.; Magni, P.; Cristofani, R.; Liu, J.; Corsini, A.; Ferri, N. Suppressor of Cytokine Signaling-3 (SOCS-3) Induces Proprotein Convertase Subtilisin Kexin Type 9 (PCSK9) Expression in Hepatic HepG2 Cell Line. J. Biol. Chem. 2016, 291, 3508–3519. [Google Scholar] [CrossRef]
- Macchi, C.; Greco, M.F.; Botta, M.; Sperandeo, P.; Dongiovanni, P.; Valenti, L.; Cicero, A.F.G.; Borghi, C.; Lupo, M.G.; Romeo, S.; et al. Leptin, Resistin, and Proprotein Convertase Subtilisin/Kexin Type 9: The Role of STAT3. Am. J. Pathol. 2020, 190, 2226–2236. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in atherosclerosis: A dynamic balance. Nat. Rev. Immunol. 2013, 13, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Luquero, A.; Crespo, J.; Pena, E.; Borrell-Pages, M. PCSK9 and LRP5 in macrophage lipid internalization and inflammation. Cardiovasc. Res. 2021, 117, 2054–2068. [Google Scholar] [CrossRef] [PubMed]
- Ricci, C.; Ruscica, M.; Camera, M.; Rossetti, L.; Macchi, C.; Colciago, A.; Zanotti, I.; Lupo, M.G.; Adorni, M.P.; Cicero, A.F.G.; et al. PCSK9 induces a pro-inflammatory response in macrophages. Sci. Rep. 2018, 8, 2267. [Google Scholar] [CrossRef] [PubMed]
- Katsuki, S.; Kumar Jha, P.; Lupieri, A.; Nakano, T.; Passos, L.S.A.; Rogers, M.A.; Becker-Greene, D.; Le, T.D.; Decano, J.L.; Ho Lee, L.; et al. Proprotein Convertase Subtilisin/Kexin 9 Promotes Macrophage Activation via LDL Receptor-Independent Mechanisms. Circ. Res. 2022, e320056. [Google Scholar] [CrossRef]
- Jaen, R.I.; Povo-Retana, A.; Rosales-Mendoza, C.; Capillas-Herrero, P.; Sanchez-Garcia, S.; Martin-Sanz, P.; Mojena, M.; Prieto, P.; Bosca, L. Functional Crosstalk between PCSK9 Internalization and Pro-Inflammatory Activation in Human Macrophages: Role of Reactive Oxygen Species Release. Int. J. Mol. Sci. 2022, 23, 9114. [Google Scholar] [CrossRef]
- Moore, K.J.; Koplev, S.; Fisher, E.A.; Tabas, I.; Bjorkegren, J.L.M.; Doran, A.C.; Kovacic, J.C. Macrophage Trafficking, Inflammatory Resolution, and Genomics in Atherosclerosis: JACC Macrophage in CVD Series (Part 2). J. Am. Coll. Cardiol. 2018, 72, 2181–2197. [Google Scholar] [CrossRef]
- Gerrity, R.G.; Naito, H.K. Lipid clearance from fatty streak lesions by foam cell migration. Artery 1980, 8, 215–219. [Google Scholar]
- Kzhyshkowska, J.; Neyen, C.; Gordon, S. Role of macrophage scavenger receptors in atherosclerosis. Immunobiology 2012, 217, 492–502. [Google Scholar] [CrossRef]
- Shu, H.; Peng, Y.; Hang, W.; Nie, J.; Zhou, N.; Wang, D.W. The role of CD36 in cardiovascular disease. Cardiovasc. Res. 2022, 118, 115–129. [Google Scholar] [CrossRef]
- Tang, Z.; Jiang, L.; Peng, J.; Ren, Z.; Wei, D.; Wu, C.; Pan, L.; Jiang, Z.; Liu, L. PCSK9 siRNA suppresses the inflammatory response induced by oxLDL through inhibition of NF-kappaB activation in THP-1-derived macrophages. Int. J. Mol. Med. 2012, 30, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, X.; Liu, J.; Wang, Y.; Zhang, S. Inclisiran inhibits oxidized low-density lipoprotein-induced foam cell formation in Raw264.7 macrophages via activating the PPARgamma pathway. Autoimmunity 2022, 55, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Novoa, B.; Figueras, A. Zebrafish: Model for the study of inflammation and the innate immune response to infectious diseases. Adv. Exp. Med. Biol. 2012, 946, 253–275. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Interleukin-1 Beta as a Target for Atherosclerosis Therapy: Biological Basis of CANTOS and Beyond. J. Am. Coll. Cardiol. 2017, 70, 2278–2289. [Google Scholar] [CrossRef]
- Murphy, E.; Ardehali, H.; Balaban, R.S.; DiLisa, F.; Dorn, G.W., 2nd; Kitsis, R.N.; Otsu, K.; Ping, P.; Rizzuto, R.; Sack, M.N.; et al. Mitochondrial Function, Biology, and Role in Disease: A Scientific Statement From the American Heart Association. Circ. Res. 2016, 118, 1960–1991. [Google Scholar] [CrossRef] [PubMed]
- Dumont, A.; Lee, M.; Barouillet, T.; Murphy, A.; Yvan-Charvet, L. Mitochondria orchestrate macrophage effector functions in atherosclerosis. Mol. Aspects Med. 2021, 77, 100922. [Google Scholar] [CrossRef]
- Soto-Heredero, G.; Gomez de Las Heras, M.M.; Gabande-Rodriguez, E.; Oller, J.; Mittelbrunn, M. Glycolysis—A key player in the inflammatory response. FEBS J. 2020, 287, 3350–3369. [Google Scholar] [CrossRef]
- Macchi, C.; Moregola, A.; Greco, M.F.; Svecla, M.; Bonacina, F.; Dhup, S.; Dadhich, R.K.; Audano, M.; Sonveaux, P.; Mauro, C.; et al. Monocarboxylate transporter 1 deficiency impacts CD8(+) T lymphocytes proliferation and recruitment to adipose tissue during obesity. iScience 2022, 25, 104435. [Google Scholar] [CrossRef]
- Ferri, N.; Colombo, G.; Ferrandi, C.; Raines, E.W.; Levkau, B.; Corsini, A. Simvastatin reduces MMP1 expression in human smooth muscle cells cultured on polymerized collagen by inhibiting Rac1 activation. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1043–1049. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Favari, E.; Zimetti, F.; Bortnick, A.E.; Adorni, M.P.; Zanotti, I.; Canavesi, M.; Bernini, F. Impaired ATP-binding cassette transporter A1-mediated sterol efflux from oxidized LDL-loaded macrophages. FEBS Lett. 2005, 579, 6537–6542. [Google Scholar] [CrossRef] [PubMed]
- Kritharides, L.; Jessup, W.; Mander, E.L.; Dean, R.T. Apolipoprotein A-I-mediated efflux of sterols from oxidized LDL-loaded macrophages. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 276–289. [Google Scholar] [CrossRef]
- Macchi, C.; Bonalume, V.; Greco, M.F.; Mozzo, M.; Melfi, V.; Sirtori, C.R.; Magnaghi, V.; Corsini, A.; Ruscica, M. Impact of Atorvastatin on Skeletal Muscle Mitochondrial Activity, Locomotion and Axonal Excitability-Evidence from ApoE(-/-) Mice. Int. J. Mol. Sci. 2022, 23, 5415. [Google Scholar] [CrossRef]
- Brioschi, M.; Lento, S.; Tremoli, E.; Banfi, C. Proteomic analysis of endothelial cell secretome: A means of studying the pleiotropic effects of Hmg-CoA reductase inhibitors. J. Proteom. 2013, 78, 346–361. [Google Scholar] [CrossRef]
- Brioschi, M.; Eligini, S.; Crisci, M.; Fiorelli, S.; Tremoli, E.; Colli, S.; Banfi, C. A mass spectrometry-based workflow for the proteomic analysis of in vitro cultured cell subsets isolated by means of laser capture microdissection. Anal. Bioanal. Chem. 2014, 406, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Distler, U.; Kuharev, J.; Navarro, P.; Tenzer, S. Label-free quantification in ion mobility-enhanced data-independent acquisition proteomics. Nat. Protoc. 2016, 11, 795–812. [Google Scholar] [CrossRef]
- Roverso, M.; Brioschi, M.; Banfi, C.; Visentin, S.; Burlina, S.; Seraglia, R.; Traldi, P.; Lapolla, A. A preliminary study on human placental tissue impaired by gestational diabetes: A comparison of gel-based versus gel-free proteomics approaches. Eur. J. Mass. Spectrom. (Chichester) 2016, 22, 71–82. [Google Scholar] [CrossRef]
- Amadio, P.; Macchi, C.; Favero, C.; Zara, M.; Solazzo, G.; Dioni, L.; Sandrini, L.; Vigna, L.; Greco, M.F.; Buoli, M.; et al. Brain-Derived Neurotrophic Factor and Extracellular Vesicle-Derived miRNAs in an Italian Cohort of Individuals With Obesity: A Key to Explain the Link Between Depression and Atherothrombosis. Front. Cardiovasc. Med. 2022, 9, 906483. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Cafora, M.; Hoxha, M.; Cantone, L.; Bollati, V.; Pistocchi, A.; Ferrari, L. Assessment of innate immune response activation following the injection of extracellular vesicles isolated from human cell cultures in zebrafish embryos. Methods Enzymol. 2020, 645, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Spreafico, M.; Gruszka, A.M.; Valli, D.; Mazzola, M.; Deflorian, G.; Quinte, A.; Totaro, M.G.; Battaglia, C.; Alcalay, M.; Marozzi, A.; et al. HDAC8: A Promising Therapeutic Target for Acute Myeloid Leukemia. Front. Cell Dev. Biol. 2020, 8, 844. [Google Scholar] [CrossRef] [PubMed]
- Pfrieger, F.W.; Vitale, N. Cholesterol and the journey of extracellular vesicles. J. Lipid Res. 2018, 59, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Khatua, S.; Simal-Gandara, J.; Acharya, K. Understanding immune-modulatory efficacy in vitro. Chem. Biol. Interact. 2022, 352, 109776. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Peptide Count | Unique Peptides | Confidence Score | p Value | Max Fold Change |
|---|---|---|---|---|---|---|
| P20592; P20591 | Interferon-induced GTP-binding protein Mx2 | 2 | 2 | 10.92 | 2.88 × 10−5 | 3.57 |
| P13807 | Glycogen (starch) synthase_ muscle | 4 | 4 | 23.71 | 1.83 × 10−13 | 2.95 |
| P68036; A0A1B0GUS4 | Ubiquitin-conjugating enzyme E2 L3 | 5 | 5 | 53.67 | 1.97 × 10−13 | 2.72 |
| Q02750; P36507 | Dual specificity mitogen-activated protein kinase kinase 1 | 6 | 2 | 36.97 | 4.70 × 10−7 | 2.64 |
| P17302 | Gap junction alpha-1 protein | 2 | 2 | 13.03 | 9.33 × 10−8 | 2.36 |
| P04075; P05062 | Fructose-bisphosphate aldolase A | 15 | 7 | 154.95 | 1.12 × 10−13 | 2.19 |
| Q13228 | Methanethiol oxidase | 3 | 3 | 17.18 | 3.04 × 10−8 | 2.07 |
| Q96HE7 | ERO1-like protein alpha | 6 | 3 | 37.10 | 1.39 × 10−10 | 2.03 |
| P00338 | L-lactate dehydrogenase A chain | 17 | 10 | 149.76 | 2.92 × 10−12 | 2.01 |
| P00558 | Phosphoglycerate kinase 1 | 17 | 11 | 169.71 | 3.76 × 10−11 | 1.87 |
| P30044 | Peroxiredoxin-5_ mitochondrial | 4 | 4 | 39.60 | 2.53 × 10−8 | 1.84 |
| Q9Y6B6 | GTP-binding protein SAR1b | 4 | 2 | 34.89 | 0.000135 | 1.81 |
| O00499 | Myc box-dependent-interacting protein 1 | 4 | 3 | 20.62 | 1.42 × 10−9 | 1.75 |
| Q9BVA1 | Tubulin beta-2B chain | 20 | 3 | 179.13 | 0.001302 | 1.74 |
| Q12797 | Aspartyl/asparaginyl beta-hydroxylase | 4 | 3 | 19.84 | 2.91 × 10−11 | 1.71 |
| Q10471 | Polypeptide N-acetylgalactosaminyltransferase 2 | 5 | 2 | 27.74 | 6.82 × 10−8 | 1.69 |
| Q12840 | Kinesin heavy chain isoform 5A | 14 | 4 | 86.31 | 4.71 × 10−7 | 1.66 |
| Q8NBP7 | Proprotein convertase subtilisin/kexin type 9 | 6 | 5 | 39.04 | 2.34 × 10−6 | 1.65 |
| P43490 | Nicotinamide phosphoribosyltransferase | 7 | 4 | 40.76 | 1.29 × 10−8 | 1.64 |
| P23381 | Tryptophan-tRNA ligase_ cytoplasmic | 5 | 4 | 33.11 | 3.90 × 10−10 | 1.64 |
| P48059 | LIM and senescent cell antigen-like-containing domain protein 1 | 2 | 2 | 11.07 | 4.80 × 10−5 | 1.63 |
| Q15050 | Ribosome biogenesis regulatory protein homolog | 2 | 2 | 11.53 | 2.63 × 10−6 | 1.61 |
| O95379 | Tumour necrosis factor alpha-induced protein 8 | 2 | 2 | 11.95 | 0.000177 | 1.60 |
| P07948 | Tyrosine-protein kinase Lyn | 3 | 2 | 19.58 | 7.27 × 10−9 | 1.58 |
| P31150 | Rab GDP dissociation inhibitor alpha | 12 | 4 | 99.46 | 1.58 × 10−7 | 1.57 |
| P14854 | Cytochrome c oxidase subunit 6B1 | 3 | 3 | 17.72 | 1.91 × 10−13 | 1.56 |
| Q06330 | Recombining binding protein suppressor of hairless | 5 | 4 | 27.02 | 1.45 × 10−8 | 1.56 |
| P60174 | Triosephosphate isomerase | 12 | 7 | 99.95 | 2.06 × 10−8 | 1.55 |
| Q9HC10 | Otoferlin | 4 | 2 | 20.14 | 3.29 × 10−8 | 1.54 |
| P68366 | Tubulin alpha-4A chain | 18 | 2 | 187.63 | 3.90 × 10−8 | 1.53 |
| P46108 | Adapter molecule crk | 4 | 4 | 25.52 | 3.67 × 10−6 | 1.53 |
| P58546 | Myotrophin | 3 | 2 | 18.91 | 4.77 × 10−5 | 1.52 |
| Q9BQE3 | Tubulin alpha-1C chain | 22 | 2 | 232.79 | 5.32 × 10−7 | 1.50 |
| Accession | Description | Peptide Count | Unique Peptides | Confidence Score | p Value | Max Fold Change |
|---|---|---|---|---|---|---|
| Q8NBP7 | Proprotein convertase subtilisin/kexin type 9 | 12 | 12 | 105.04 | 8.05 × 10−5 | 7.10 |
| O95967 | EGF-containing fibulin-like extracellular matrix protein 2 | 7 | 7 | 64.03 | 2.27 × 10−4 | 4.30 |
| Q9Y6C2 | EMILIN-1 | 12 | 12 | 92.98 | 8.26 × 10−5 | 3.42 |
| P04908 | Histone H2A type 1-B/E | 3 | 3 | 23.21 | 1.21 × 10−3 | 2.90 |
| P62805 | Histone H4 | 5 | 5 | 38.37 | 3.81 × 10−4 | 2.76 |
| P35442 | Thrombospondin-2 | 10 | 9 | 61.02 | 4.07 × 10−4 | 2.61 |
| P01024 | Complement C3 | 7 | 7 | 44.15 | 3.35 × 10−4 | 2.40 |
| Q14112 | Nidogen-2 | 10 | 9 | 64.07 | 1.09 × 10−5 | 2.29 |
| P17987 | T-complex protein 1 subunit alpha | 3 | 3 | 16.69 | 4.26 × 10−3 | 2.25 |
| Q9UQP3 | Tenascin-N | 4 | 4 | 22.53 | 2.15 × 10−3 | 2.08 |
| P98160 | Basement membrane-specific heparan sulfate proteoglycan core protein | 40 | 40 | 264.35 | 2.40 × 10−4 | 2.01 |
| O60814 | Histone H2B type 1-K | 5 | 5 | 36.96 | 5.82 × 10−3 | 1.83 |
| Q8WUM4 | Programmed cell death 6-interacting protein | 15 | 15 | 97.41 | 2.69 × 10−3 | 1.55 |
| P78509 | Reelin | 7 | 7 | 37.44 | 8.13 × 10−3 | 1.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greco, M.F.; Rizzuto, A.S.; Zarà, M.; Cafora, M.; Favero, C.; Solazzo, G.; Giusti, I.; Adorni, M.P.; Zimetti, F.; Dolo, V.; et al. PCSK9 Confers Inflammatory Properties to Extracellular Vesicles Released by Vascular Smooth Muscle Cells. Int. J. Mol. Sci. 2022, 23, 13065. https://doi.org/10.3390/ijms232113065
Greco MF, Rizzuto AS, Zarà M, Cafora M, Favero C, Solazzo G, Giusti I, Adorni MP, Zimetti F, Dolo V, et al. PCSK9 Confers Inflammatory Properties to Extracellular Vesicles Released by Vascular Smooth Muscle Cells. International Journal of Molecular Sciences. 2022; 23(21):13065. https://doi.org/10.3390/ijms232113065
Chicago/Turabian StyleGreco, Maria Francesca, Alessandra Stefania Rizzuto, Marta Zarà, Marco Cafora, Chiara Favero, Giulia Solazzo, Ilaria Giusti, Maria Pia Adorni, Francesca Zimetti, Vincenza Dolo, and et al. 2022. "PCSK9 Confers Inflammatory Properties to Extracellular Vesicles Released by Vascular Smooth Muscle Cells" International Journal of Molecular Sciences 23, no. 21: 13065. https://doi.org/10.3390/ijms232113065
APA StyleGreco, M. F., Rizzuto, A. S., Zarà, M., Cafora, M., Favero, C., Solazzo, G., Giusti, I., Adorni, M. P., Zimetti, F., Dolo, V., Banfi, C., Ferri, N., Sirtori, C. R., Corsini, A., Barbieri, S. S., Pistocchi, A., Bollati, V., Macchi, C., & Ruscica, M. (2022). PCSK9 Confers Inflammatory Properties to Extracellular Vesicles Released by Vascular Smooth Muscle Cells. International Journal of Molecular Sciences, 23(21), 13065. https://doi.org/10.3390/ijms232113065