
Expression of DNA Methyltransferase 3B Isoforms Is Associated with DNA Satellite 2 Hypomethylation and Clinical Prognosis in Advanced High-Grade Serous Ovarian Carcinoma

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
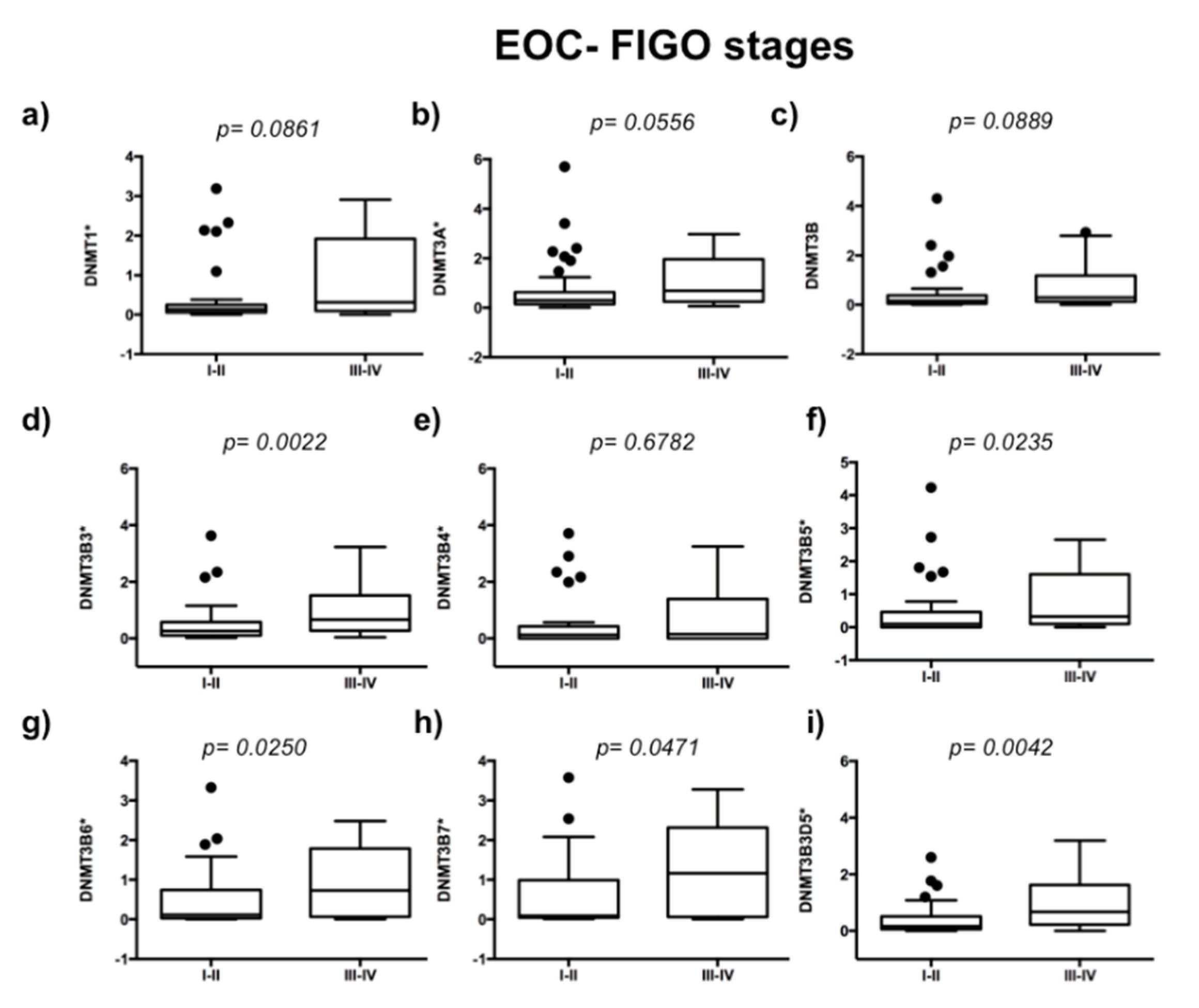
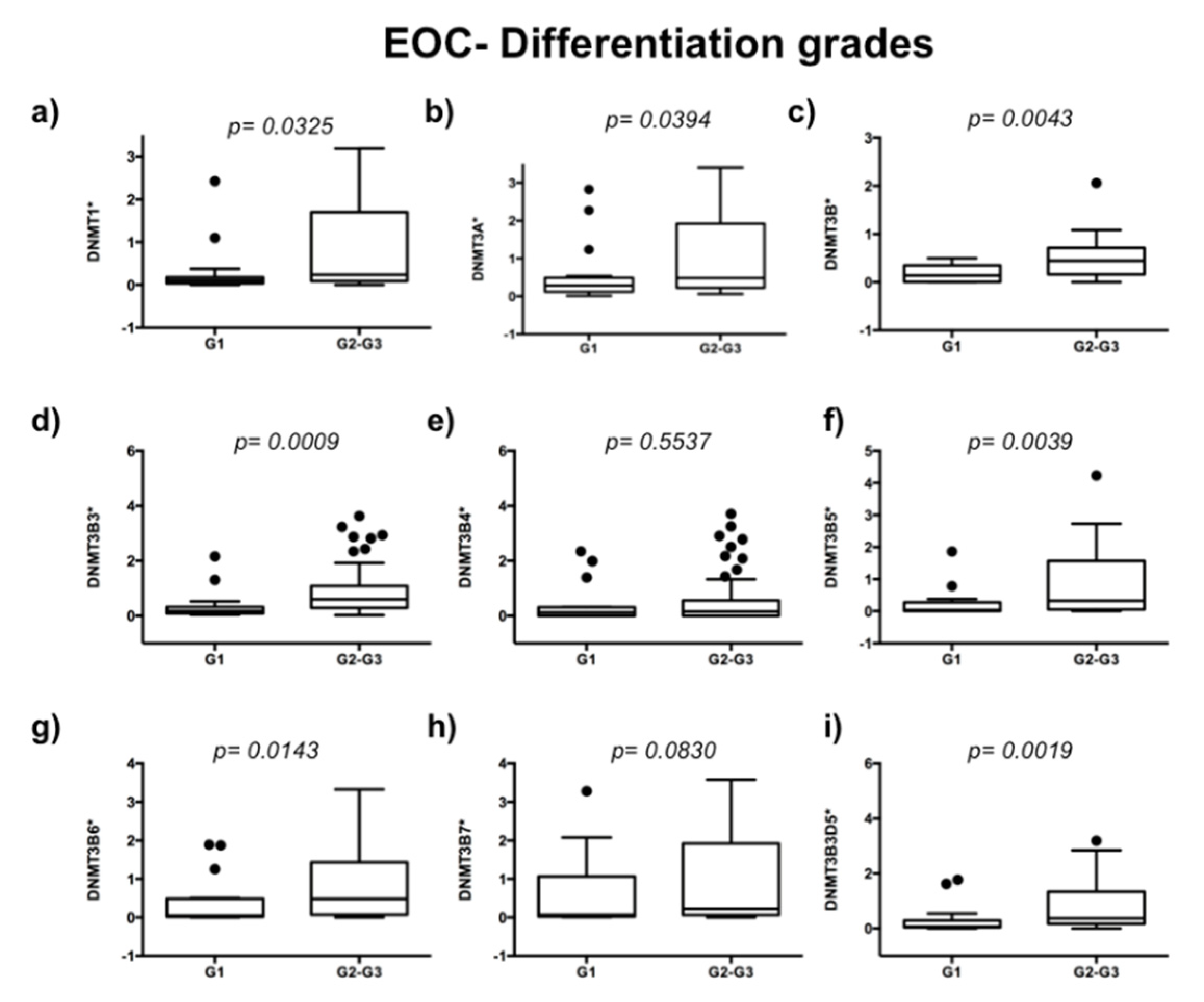
2.1. DNA Methyltransferases Expression in Ovarian Carcinoma
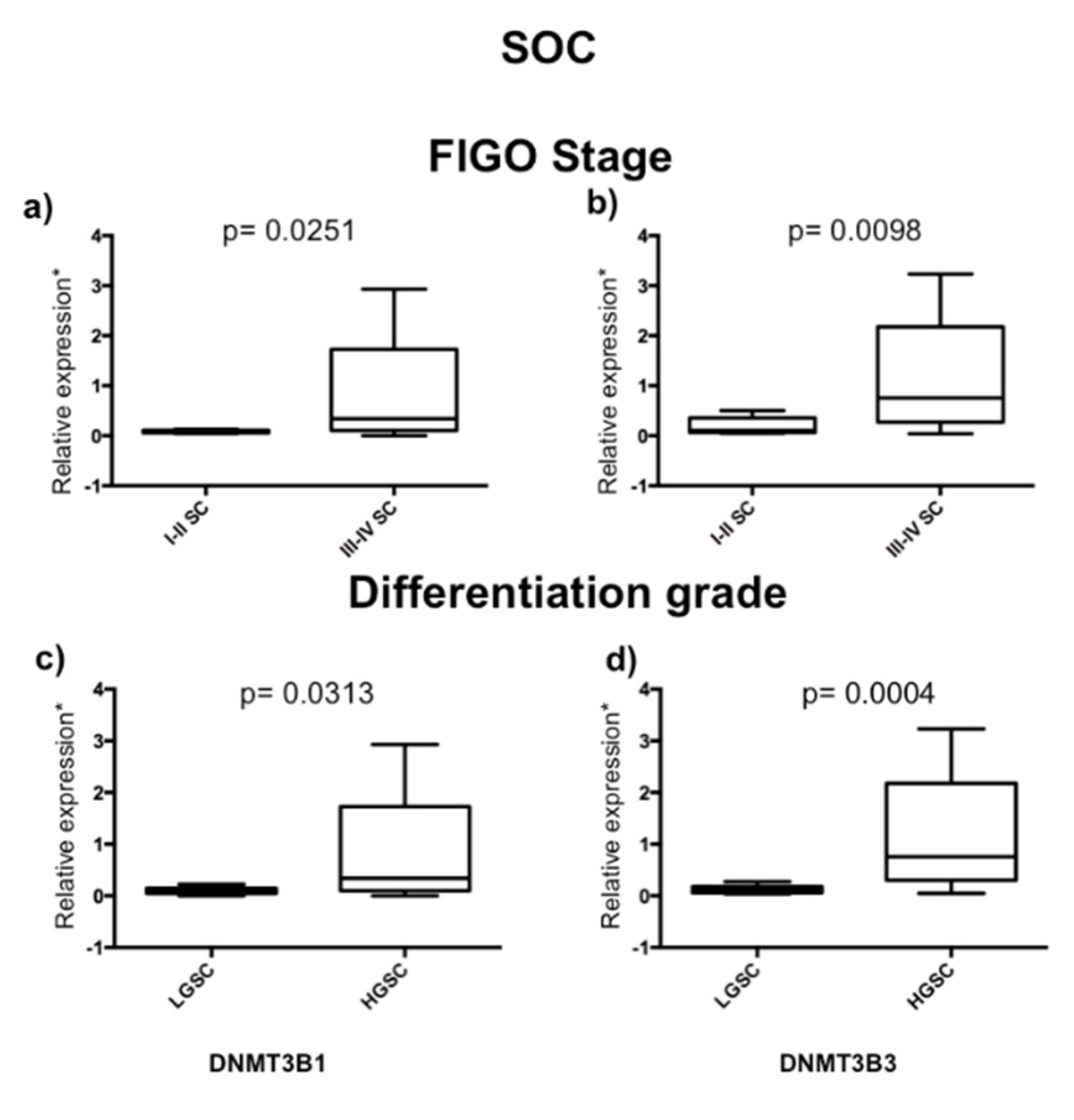
2.2. DNMT3B1 and DNMT3B3 Are Overexpressed in High-Grade and Advanced Stages of Serous Ovarian Carcinoma
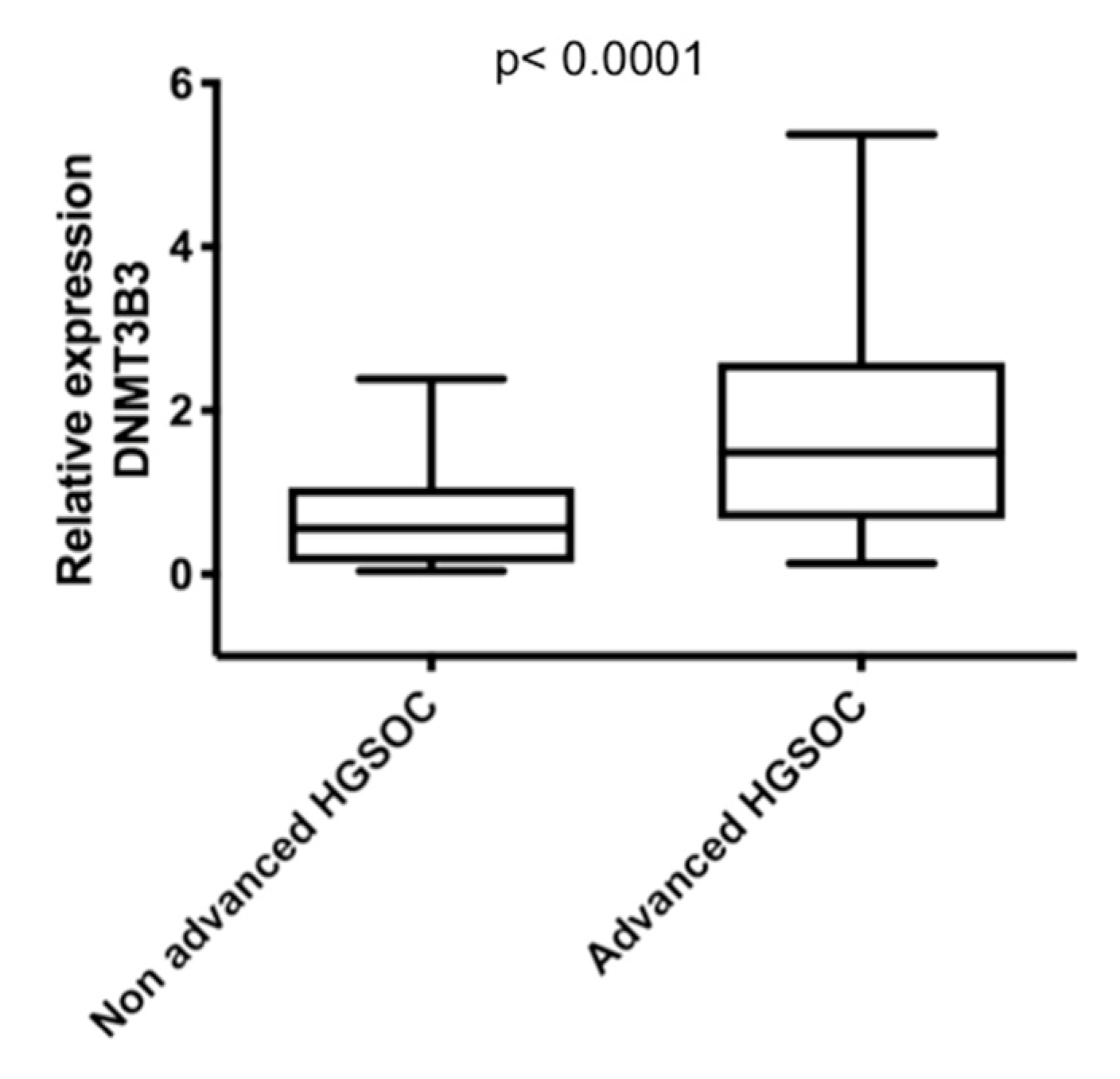
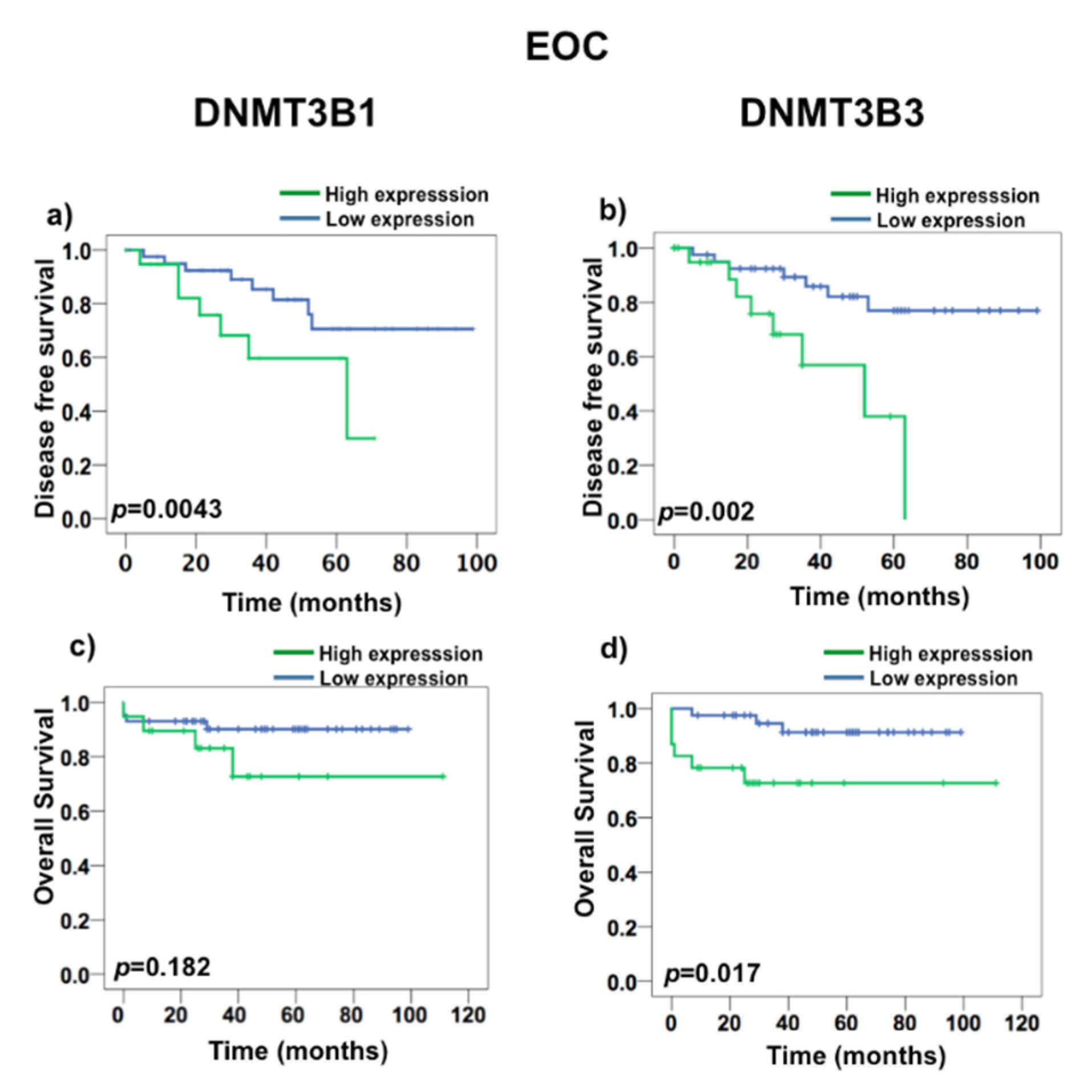
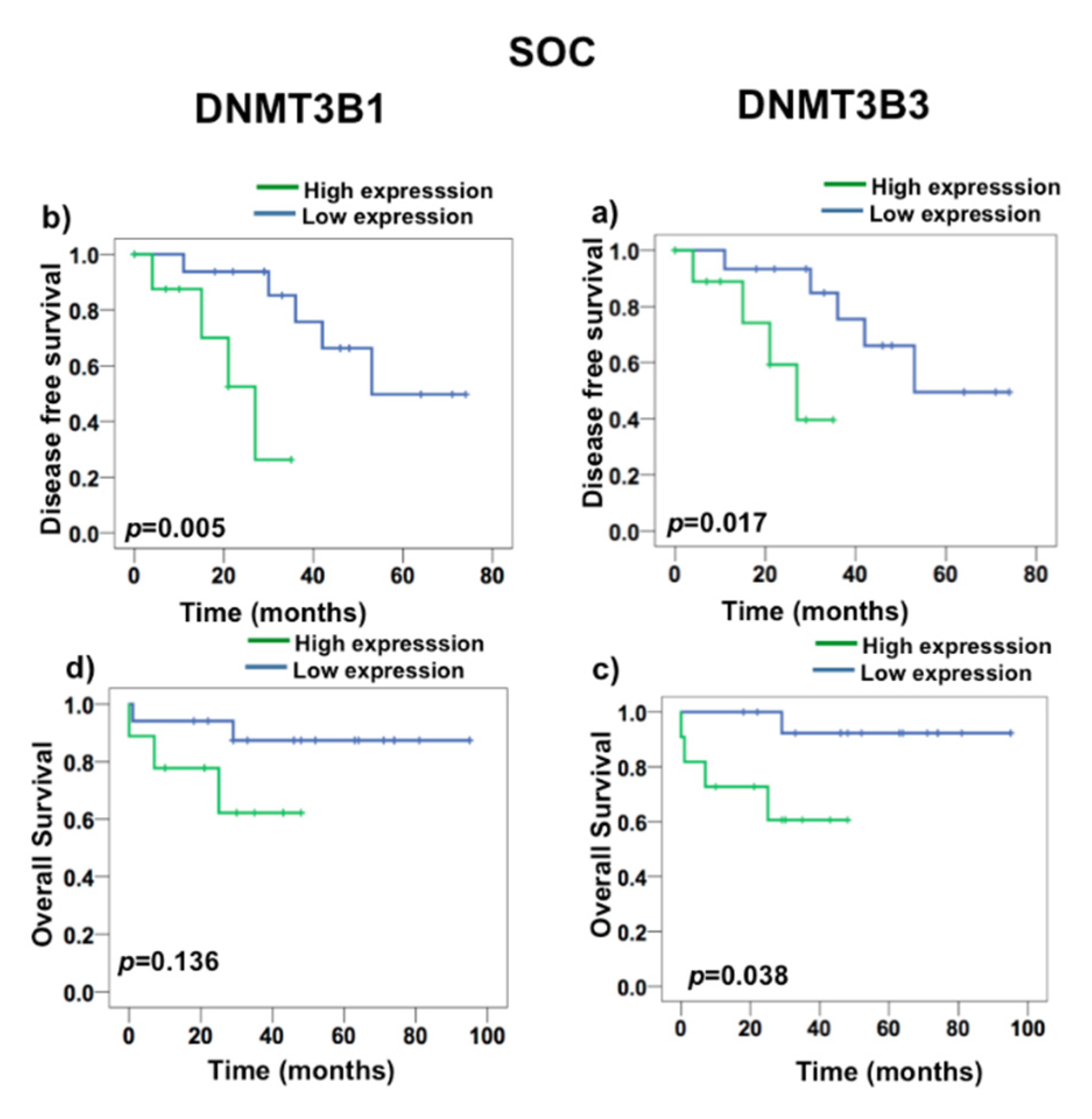
2.3. DNMT3B3 Overexpression Is Associated with Poor Prognosis in Serous Ovarian Carcinoma
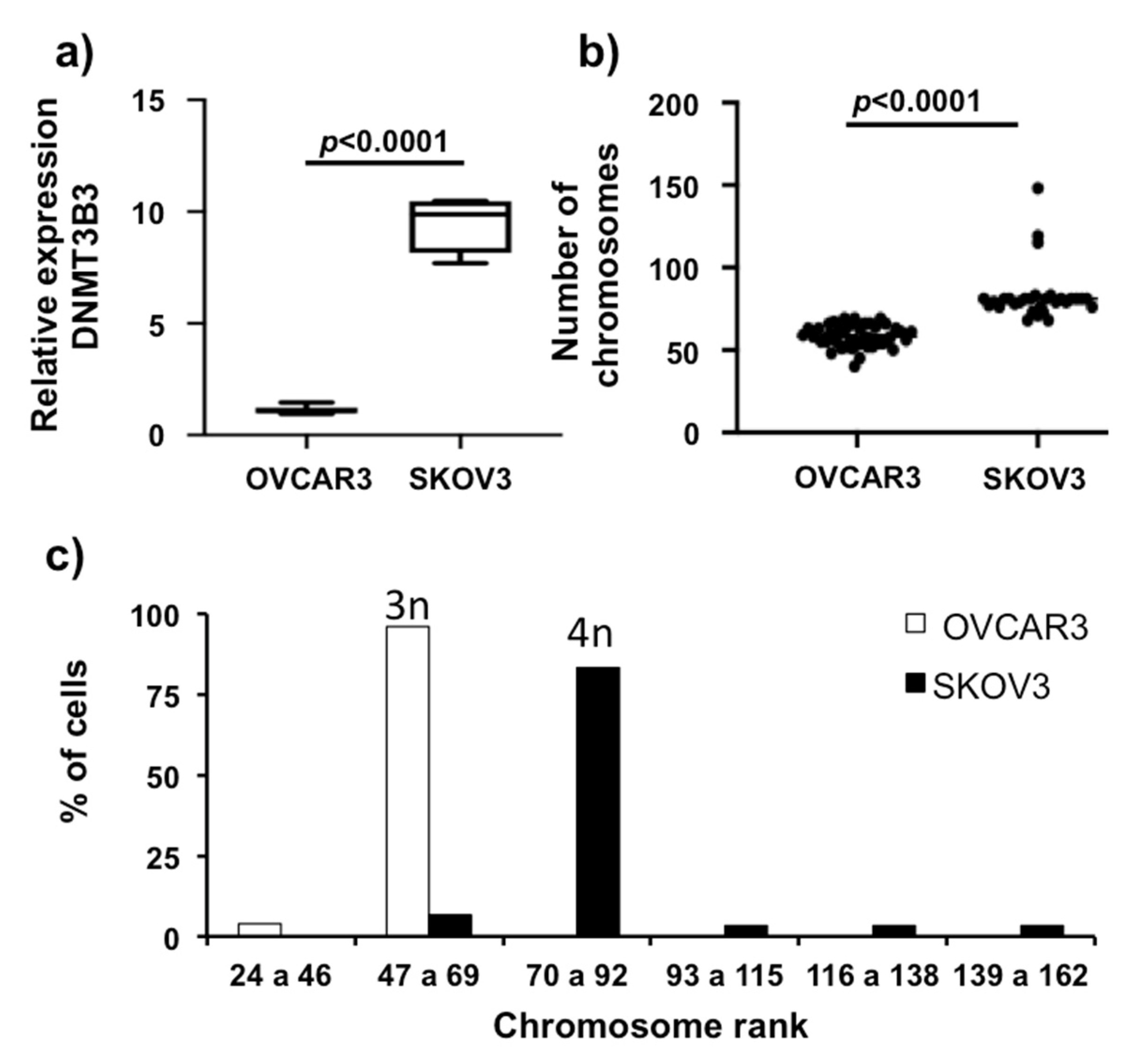
2.4. DNA Methyltransferase 3B3 Expression in High-Grade, Advanced Ovarian Carcinoma Cell Lines
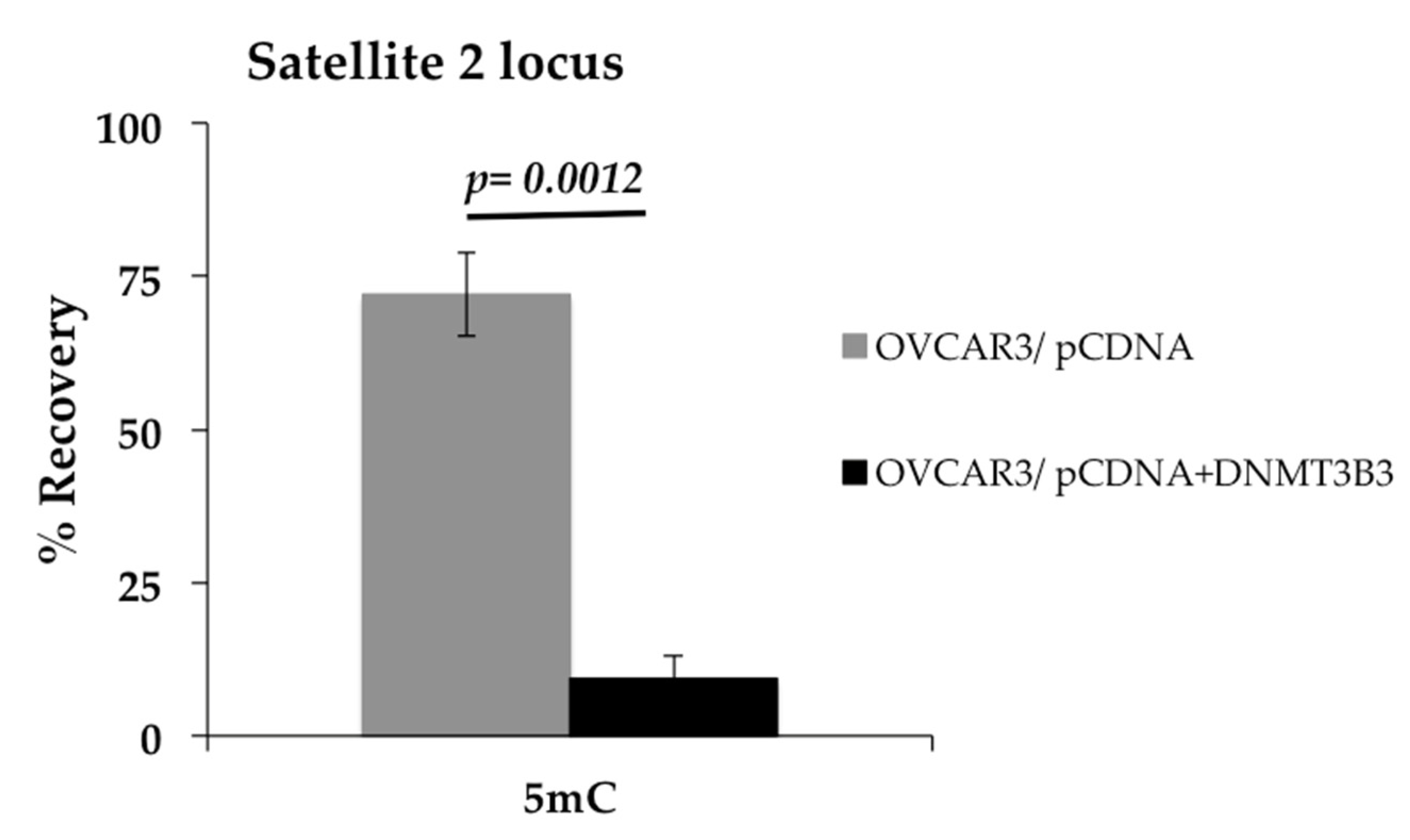
2.5. Overexpression of DNA Methyltransferase 3B3 Is a Cause of DNA Hypomethylation in the OVCAR3 Cell Line
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Shih, I.-M. The Origin and Pathogenesis of Epithelial Ovarian Cancer: A Proposed Unifying Theory. Am. J. Surg. Pathol. 2010, 34, 433–443. [Google Scholar] [CrossRef] [PubMed]
- McCluggage, W.G. Morphological Subtypes of Ovarian Carcinoma: A Review with Emphasis on New Developments and Pathogenesis. Pathology 2011, 43, 420–432. [Google Scholar] [CrossRef]
- Li, E.; Zhang, Y. DNA Methylation in Mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a019133. [Google Scholar] [CrossRef] [PubMed]
- Jurkowska, R.Z.; Jurkowski, T.P.; Jeltsch, A. Structure and Function of Mammalian DNA Methyltransferases. ChemBioChem 2011, 12, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Vavouri, T.; Lehner, B. Human Genes with CpG Island Promoters Have a Distinct Transcription-Associated Chromatin Organization. Genome Biol. 2012, 13, R110. [Google Scholar] [CrossRef]
- Zeimet, A.G.; Fiegl, H.; Goebel, G.; Kopp, F.; Allasia, C.; Reimer, D.; Steppan, I.; Mueller-Holzner, E.; Ehrlich, M.; Marth, C. DNA Ploidy, Nuclear Size, Proliferation Index and DNA-Hypomethylation in Ovarian Cancer. Gynecol. Oncol. 2011, 121, 24–31. [Google Scholar] [CrossRef]
- Widschwendter, M.; Jiang, G.; Woods, C.; Müller, H.M.; Fiegl, H.; Goebel, G.; Marth, C.; Müller-Holzner, E.; Zeimet, A.G.; Laird, P.W.; et al. DNA Hypomethylation and Ovarian Cancer Biology. Cancer Res. 2004, 64, 4472–4480. [Google Scholar] [CrossRef]
- Wiley, A.; Katsaros, D.; Chen, H.; Rigault de la Longrais, I.A.; Beeghly, A.; Puopolo, M.; Singal, R.; Zhang, Y.; Amoako, A.; Zelterman, D.; et al. Aberrant Promoter Methylation of Multiple Genes in Malignant Ovarian Tumors and in Ovarian Tumors with Low Malignant Potential. Cancer 2006, 107, 299–308. [Google Scholar] [CrossRef]
- Shao, C.; Lacey, M.; Dubeau, L.; Ehrlich, M. Hemimethylation Footprints of DNA Demethylation in Cancer. Epigenetics 2009, 4, 165–175. [Google Scholar] [CrossRef]
- Press, J.Z.; De Luca, A.; Boyd, N.; Young, S.; Troussard, A.; Ridge, Y.; Kaurah, P.; Kalloger, S.E.; Blood, K.A.; Smith, M.; et al. Ovarian Carcinomas with Genetic and Epigenetic BRCA1 Loss Have Distinct Molecular Abnormalities. BMC Cancer 2008, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- De Caceres, I.I.; Battagli, C.; Esteller, M.; Herman, J.G.; Dulaimi, E.; Edelson, M.I.; Bergman, C.; Ehya, H.; Eisenberg, B.L.; Cairns, P. Tumor Cell-Specific BRCA1 and RASSF1A Hypermethylation in Serum, Plasma, and Peritoneal Fluid from Ovarian Cancer Patients. Cancer Res. 2004, 64, 6476–6481. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, F.; Altinisik, J.; Karateke, A.; Coksuer, H.; Buyru, N. Methylation of Tumor Suppressor Genes in Ovarian Cancer. Exp. Ther. Med. 2012, 4, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Moselhy, S.S.; Kumosani, T.A.; Kamal, I.; Jalal, J.; Jabaar, H.S.A.; Dalol, A. Hypermethylation of P 15, P 16, and E-Cadherin Genes in Ovarian Cancer. Toxicol. Ind. Health 2015, 31, 924–930. [Google Scholar] [CrossRef]
- Chédin, F. The DNMT3 Family of Mammalian De Novo DNA Methyltransferases. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2011; Volume 101, pp. 255–285. ISBN 978-0-12-387685-0. [Google Scholar]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for De Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Ratnam, S.; Mertineit, C.; Ding, F.; Howell, C.Y.; Clarke, H.J.; Bestor, T.H.; Chaillet, J.R.; Trasler, J.M. Dynamics of Dnmt1 Methyltransferase Expression and Intracellular Localization during Oogenesis and Preimplantation Development. Dev. Biol. 2002, 245, 304–314. [Google Scholar] [CrossRef][Green Version]
- Manzo, M.; Wirz, J.; Ambrosi, C.; Villaseñor, R.; Roschitzki, B.; Baubec, T. Isoform-specific Localization of DNMT3A Regulates DNA Methylation Fidelity at Bivalent CpG Islands. EMBO J. 2017, 36, 3421–3434. [Google Scholar] [CrossRef]
- Ostler, K.R.; Davis, E.M.; Payne, S.L.; Gosalia, B.B.; Expósito-Céspedes, J.; Le Beau, M.M.; Godley, L.A. Cancer Cells Express Aberrant DNMT3B Transcripts Encoding Truncated Proteins. Oncogene 2007, 26, 5553–5563. [Google Scholar] [CrossRef]
- Ma, M.Z.; Lin, R.; Carrillo, J.; Bhutani, M.; Pathak, A.; Ren, H.; Li, Y.; Song, J.; Mao, L. ∆ DNMT3B4-Del Contributes to Aberrant DNA Methylation Patterns in Lung Tumorigenesis. EBioMedicine 2015, 2, 1340–1350. [Google Scholar] [CrossRef]
- Wang, J.; Bhutani, M.; Pathak, A.K.; Lang, W.; Ren, H.; Jelinek, J.; He, R.; Shen, L.; Issa, J.-P.; Mao, L. ΔDNMT3B Variants Regulate DNA Methylation in a Promoter-Specific Manner. Cancer Res. 2007, 67, 10647–10652. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Sun, S.; Rodriguez, M.; Yue, P.; Jang, S.; Mao, L. A Novel DNMT3B Subfamily, ΔDNMT3B, Is the Predominant Form of DNMT3B in Non-Small Cell Lung Cancer. Int. J. Oncol. 2006, 29, 201–207. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Van Emburgh, B.O.; Shan, J.; Su, Z.; Fields, C.R.; Vieweg, J.; Hamazaki, T.; Schwartz, P.H.; Terada, N.; Robertson, K.D. A Novel DNMT3B Splice Variant Expressed in Tumor and Pluripotent Cells Modulates Genomic DNA Methylation Patterns and Displays Altered DNA Binding. Mol. Cancer Res. 2009, 7, 1622–1634. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Xie, S.; Li, E. Cloning and Characterization of a Family of Novel Mammalian DNA (Cytosine-5) Methyltransferases. Nat. Genet. 1998, 19, 219–220. [Google Scholar] [CrossRef] [PubMed]
- Weisenberger, D.J.; Velicescu, M.; Cheng, J.C.; Gonzales, F.A.; Liang, G.; Jones, P.A. Role of the DNA Methyltransferase Variant DNMT3b3 in DNA Methylation. Mol. Cancer Res. 2004, 1, 62–72. [Google Scholar] [CrossRef]
- Aoki, A. Enzymatic Properties of de Novo-Type Mouse DNA (Cytosine-5) Methyltransferases. Nucleic Acids Res. 2001, 29, 3506–3512. [Google Scholar] [CrossRef]
- Gordon, C.A.; Hartono, S.R.; Chédin, F. Inactive DNMT3B Splice Variants Modulate De Novo DNA Methylation. PLoS ONE 2013, 8, e69486. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Zhang, R.; Zhang, S.; Jiang, S.; Liu, Y.; Zhang, W.; Zhang, Y.; Li, J.; Gong, K.; Hu, X.-R.; et al. Splice Variants DNMT3B4 and DNMT3B7 Overexpression Inhibit Cell Proliferation in 293A Cell Line. Vitro Cell. Dev. Biol. Anim. 2013, 49, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Ostler, K.R.; Yang, Q.; Looney, T.J.; Zhang, L.; Vasanthakumar, A.; Tian, Y.; Kocherginsky, M.; Raimondi, S.L.; DeMaio, J.G.; Salwen, H.R.; et al. Truncated DNMT3B Isoform DNMT3B7 Suppresses Growth, Induces Differentiation, and Alters DNA Methylation in Human Neuroblastoma. Cancer Res. 2012, 72, 4714–4723. [Google Scholar] [CrossRef]
- Gopalakrishna-Pillai, S.; Iverson, L.E. A DNMT3B Alternatively Spliced Exon and Encoded Peptide Are Novel Biomarkers of Human Pluripotent Stem Cells. PLoS ONE 2011, 6, e20663. [Google Scholar] [CrossRef] [PubMed]
- Plourde, K.V.; Labrie, Y.; Ouellette, G.; Pouliot, M.-C.; Durocher, F. Genome-Wide Methylation Analysis of DNMT3B Gene Isoforms Revealed Specific Methylation Profiles in Breast Cell Lines. Epigenomics 2016, 8, 1209–1226. [Google Scholar] [CrossRef]
- Duymich, C.E.; Charlet, J.; Yang, X.; Jones, P.A.; Liang, G. DNMT3B Isoforms without Catalytic Activity Stimulate Gene Body Methylation as Accessory Proteins in Somatic Cells. Nat. Commun. 2016, 7, 11453. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Lv, C.; Qiao, F.; Qiu, X.; Huang, W.; Wu, Q.; Zhao, Z.; Fan, H. Expression Pattern and Clinical Significance of DNA Methyltransferase 3B Variants in Gastric Carcinoma. Oncol. Rep. 2010, 8, 819–826. [Google Scholar]
- Liu, Y.; Sun, L.; Fong, P.; Yang, J.; Zhang, Z.; Yin, S.; Jiang, S.; Liu, X.; Ju, H.; Huang, L.; et al. An Association between Overexpression of DNA Methyltransferase 3B4 and Clear Cell Renal Cell Carcinoma. Oncotarget 2017, 8, 19712–19722. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Kanai, Y.; Sakamoto, M.; Saito, H.; Ishii, H.; Hirohashi, S. Overexpression of a Splice Variant of DNA Methyltransferase 3b, DNMT3b4, Associated with DNA Hypomethylation on Pericentromeric Satellite Regions during Human Hepatocarcinogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 10060–10065. [Google Scholar] [CrossRef] [PubMed]
- Brambert, P.R.; Kelpsch, D.J.; Hameed, R.; Desai, C.V.; Calafiore, G.; Godley, L.A.; Raimondi, S.L. DNMT3B7 Expression Promotes Tumor Progression to a More Aggressive Phenotype in Breast Cancer Cells. PLoS ONE 2015, 10, e0117310. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; White, M.W.; Schroeder, A.M.; DeLuca, N.V.; Leszczynski, A.L.; Raimondi, S.L. Aberrant DNMT3B7 Expression Correlates to Tissue Type, Stage, and Survival across Cancers. PLoS ONE 2018, 13, e0201522. [Google Scholar] [CrossRef] [PubMed]
- Ahluwalia, A.; Hurteau, J.A.; Bigsby, R.M.; Nephew, K.P. DNA Methylation in Ovarian Cancer: II. Expression of DNA Methyltransferases in Ovarian Cancer Cell Lines and Normal Ovarian Epithelial Cells. Gynecol. Oncol. 2001, 82, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Van Emburgh, B.O.; Robertson, K.D. Modulation of Dnmt3b Function in Vitro by Interactions with Dnmt3L, Dnmt3a and Dnmt3b Splice Variants. Nucleic Acids Res. 2011, 39, 4984–5002. [Google Scholar] [CrossRef] [PubMed]
- Robertson, K.D.; Uzvolgyi, E.; Liang, G.; Talmadge, C.; Sumegi, J.; Gonzales, F.A.; Jones, P.A. The Human DNA Methyltransferases (DNMTs) 1, 3a and 3b: Coordinate MRNA Expression in Normal Tissues and Overexpression in Tumors. Nucleic Acids Res. 1999, 27, 2291–2298. [Google Scholar] [CrossRef]
- Wang, J.; Walsh, G.; Liu, D.D.; Lee, J.J.; Mao, L. Expression of ΔDNMT3B Variants and Its Association with Promoter Methylation of P16 and RASSF1A in Primary Non–Small Cell Lung Cancer. Cancer Res. 2006, 66, 8361–8366. [Google Scholar] [CrossRef]
- Ehrlich, M.; Woods, C.B.; Yu, M.C.; Dubeau, L.; Yang, F.; Campan, M.; Weisenberger, D.J.; Long, T.; Youn, B.; Fiala, E.S.; et al. Quantitative Analysis of Associations between DNA Hypermethylation, Hypomethylation, and DNMT RNA Levels in Ovarian Tumors. Oncogene 2006, 25, 2636–2645. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Category | n | % |
|---|---|---|---|
| Stage | I | 29 | 46 |
| II | 7 | 11 | |
| III | 18 | 29 | |
| IV | 9 | 14 | |
| Histologic type | Serous | 27 | 43 |
| Endometrioid | 15 | 24 | |
| Clear cells | 11 | 17 | |
| Mucinous | 8 | 13 | |
| Other | 2 | 3 | |
| Tumor grade | G1 | 18 | 29 |
| G2 | 9 | 14 | |
| G3 | 36 | 57 | |
| Recurrence | Yes | 12 | 16 |
| No | 51 | 84 | |
| Current final status | Dead | 9 | 13 |
| Alive | 54 | 87 |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| DNMT1 | AACCTTCACCTAGCCCCAG | CTCATCCGATTTGGCTCTTTCA |
| DNMT3A | GACAAGAATGCCACCAAAGC | CCATCTCCGAACCACATGAC |
| DNMT3B1 | CCATCAAAGTTTCTGCTGCT | GAAGAGGTGTCGGATGACAG |
| DNMT3B3 | CCGGGATGAACAGGATCTTT | AGTAGTCCTTCAGAGGGGCG |
| DNMT3B4 | CGGTTCCTGGAGTGTAATCC | GGTTATTGTCTGTACTTTCTTTAACTGTT |
| DNMT3B5 | AATACAATAGGATAGCCAAGGATCT | TTCAGAGGGGCGAAGAGG |
| DNMT3B6 | CCAAGCTTGGAAAGCATGAA | CCGTTGACGAGGATCGAGT |
| DNMT3B7 | CAGTCTAATTACCTTTCACAGAGAACA | GCTTTGAGGCGCTTGGGT |
| DNMT3B3Δ5 | GAAAGCCCAGCTTCCCTGAGA | AGTTGTGTCCTCTGTGTCGTCTGT |
| GAPDH | TCGTTCCCAAAGTCCTCCTGTTTC | TCCGCAGCCGCCTGGTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Castillo Falconi, V.M.; Díaz-Chávez, J.; Torres-Arciga, K.; Luna-Maldonado, F.; Gudiño-Gomez, A.A.; Pedroza-Torres, A.; Castro-Hernández, C.; Cantú de León, D.; Herrera, L.A. Expression of DNA Methyltransferase 3B Isoforms Is Associated with DNA Satellite 2 Hypomethylation and Clinical Prognosis in Advanced High-Grade Serous Ovarian Carcinoma. Int. J. Mol. Sci. 2022, 23, 12759. https://doi.org/10.3390/ijms232112759
Del Castillo Falconi VM, Díaz-Chávez J, Torres-Arciga K, Luna-Maldonado F, Gudiño-Gomez AA, Pedroza-Torres A, Castro-Hernández C, Cantú de León D, Herrera LA. Expression of DNA Methyltransferase 3B Isoforms Is Associated with DNA Satellite 2 Hypomethylation and Clinical Prognosis in Advanced High-Grade Serous Ovarian Carcinoma. International Journal of Molecular Sciences. 2022; 23(21):12759. https://doi.org/10.3390/ijms232112759
Chicago/Turabian StyleDel Castillo Falconi, Victor M., José Díaz-Chávez, Karla Torres-Arciga, Fernando Luna-Maldonado, Adriana A. Gudiño-Gomez, Abraham Pedroza-Torres, Clementina Castro-Hernández, David Cantú de León, and Luis A. Herrera. 2022. "Expression of DNA Methyltransferase 3B Isoforms Is Associated with DNA Satellite 2 Hypomethylation and Clinical Prognosis in Advanced High-Grade Serous Ovarian Carcinoma" International Journal of Molecular Sciences 23, no. 21: 12759. https://doi.org/10.3390/ijms232112759
APA StyleDel Castillo Falconi, V. M., Díaz-Chávez, J., Torres-Arciga, K., Luna-Maldonado, F., Gudiño-Gomez, A. A., Pedroza-Torres, A., Castro-Hernández, C., Cantú de León, D., & Herrera, L. A. (2022). Expression of DNA Methyltransferase 3B Isoforms Is Associated with DNA Satellite 2 Hypomethylation and Clinical Prognosis in Advanced High-Grade Serous Ovarian Carcinoma. International Journal of Molecular Sciences, 23(21), 12759. https://doi.org/10.3390/ijms232112759