1. Introduction
Recent studies have shown activated immune cells to undergo distinct metabolic changes, which not only fulfill the energy needs of these cells but also contribute to their effector function (reviewed in [
1]). These metabolic changes are part of trained immune responses, which summarize the long-term functional reprogramming of innate immune cells evoked by exogenous or endogenous insults, leading to an altered response towards a second challenge after the return to a nonactivated state [
2]. Innate immune cells undergoing trained immune responses can either mount stronger (trained immunity) or weaker (trained tolerance) immune responses mediated by a combination of epigenetic modifications and metabolic reprogramming of innate effector cells, as well as the long-term reprogramming of their bone marrow progenitor cells [
2].
Accordingly, better understanding the mechanisms underlying trained immune responses will allow for a new generation of therapeutics and vaccines that combine trained immune responses in innate immune cells with the induction of classical antigen-specific adaptive immune responses.
As a model for such novel therapeutics in the field of allergies, we generated a recombinant fusion protein, rFlaA:Betv1, that combines the TLR5-ligand flagellin A from
Listeria monocytogenes with the major birch pollen allergen Bet v 1 into a single molecule [
3]. In previous studies, we were able to show that rFlaA:Betv1 efficiently suppressed allergic sensitization in vivo [
3] while also activating myeloid DCs (mDCs) [
3,
4], macrophages [
5], epithelial cells [
6], and B cells [
7], leading to pro-inflammatory cytokine production and the subsequent formation of allergen-specific TH1 responses with TH2-suppressing capacity [
3,
5]. Furthermore, rFlaA:Betv1 induced a prominent production of the anti-inflammatory cytokine IL-10 in all investigated types of antigen presenting cells (APCs) (mDCs, macrophages, and B cells), which was shown to significantly suppress TH2 responses [
3,
5,
7].
Interestingly, previous research also showed both rFlaA:Betv1-stimulated mDCs and macrophages to undergo a metabolic switch towards a strongly glycolytic phenotype, resulting in lactate-induced acidification of the culture medium and a prominent color change in the contained pH indicator [
3,
5], referred to as the “Warburg Effect” [
8]. Further analyses showed the anti-inflammatory IL-10 secretion by rFlaA:Betv1 to be dependent on the mechanistic target of rapamycin (mTOR), a master regulator of both cellular metabolism and immune function [
3]. In contrast with this, rFlaA:Betv1-induced pro-inflammatory cytokine secretion in mDCs was shown to be largely mTOR independent, but depended on MAP kinase signaling [
4]. In line with this, a differential role of metabolic pathways in the pro- and anti-inflammatory cytokine secretion induced by rFlaA:Betv1 has been proposed [
3].
Although the metabolic changes that occur in immune cells activated by different stimuli are well described [
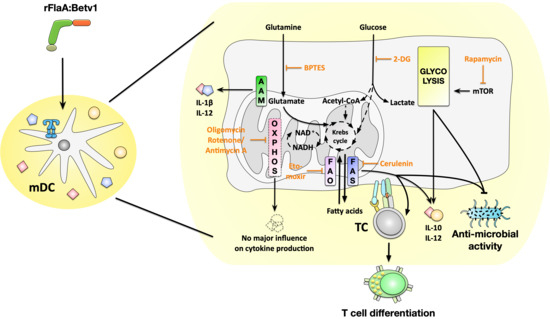
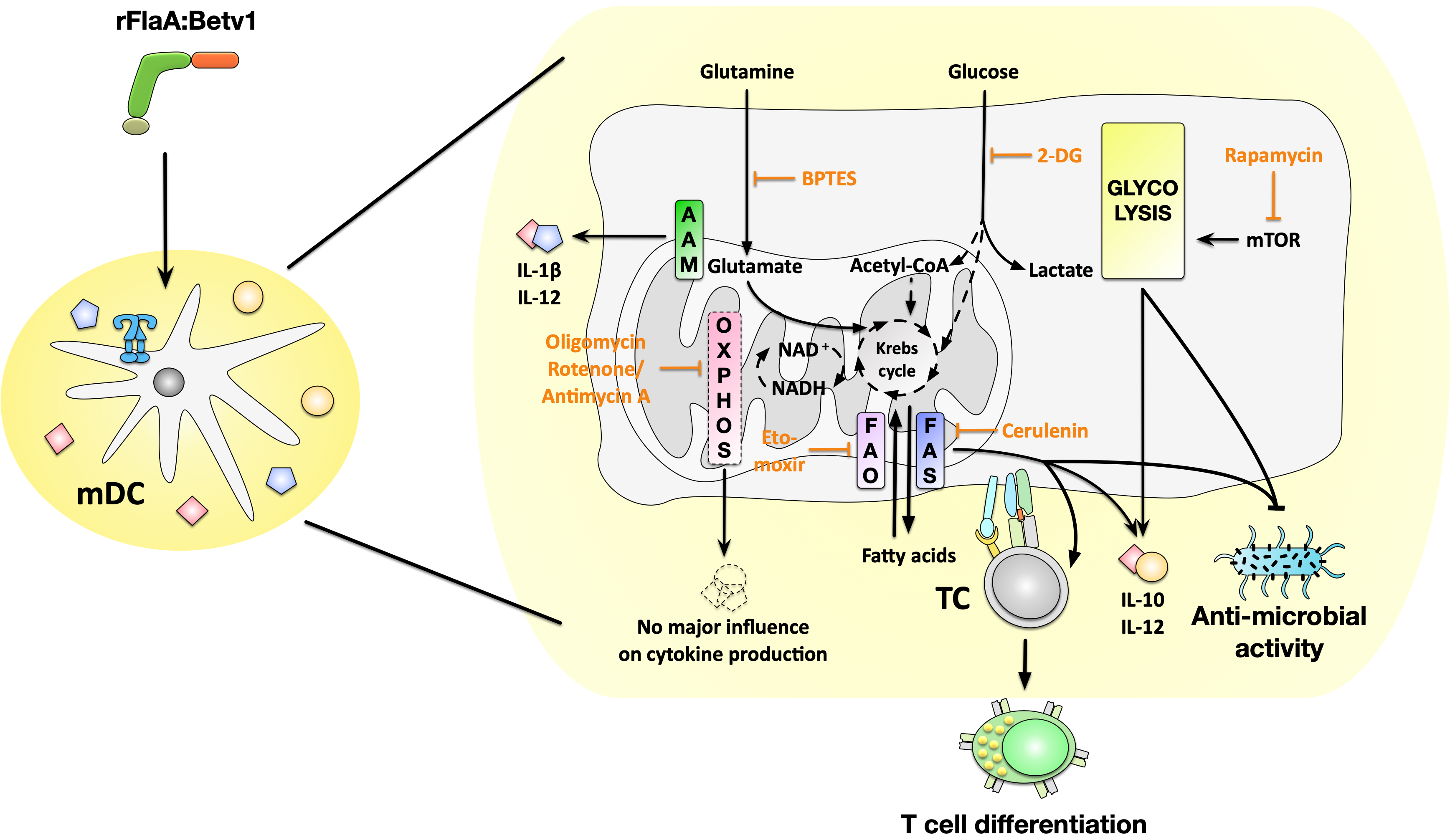
9], both the activation of immune metabolism by potential future therapeutics, such as the fusion protein rFlaA:Betv1, and its contribution to the immune-modulating properties (such as cytokine secretion, influence on activation status, and antimicrobial properties) are not fully understood. Therefore, the aim of this study was to investigate the contribution of distinct metabolic pathways, including glycolysis, mitochondrial respiration, amino acid-, or fatty acid metabolism, to the rFlaA:Betv1-mediated activation of mDCs.
3. Discussion
Over the last few years, the induction and modulation of trained immune responses in innate immune cells has become a topic of great interest in the vaccination field. Trained immunity describes the heightened reactivity of innate immune cells after prior activation either to the same or an unrelated stimulus [
2]. It is based on metabolic and epigenetic changes in both terminally differentiated immune cells, as well as their bone marrow progenitors [
2]. In addition to whole pathogens, TLR-ligands and C-type lectins were among the first described stimuli to trigger trained immune responses, making them interesting adjuvant candidates [
16,
17].
We recently generated and characterized a fusion protein consisting of the TLR5-ligand flagellin and the major birch pollen allergen Bet v 1 (rFlaA:Betv1) [
3], which was meanwhile shown to suppress TH2 responses in vitro and allergic sensitization in vivo [
3]. These immune modulating properties of rFlaA:Betv1 were paralleled by a pronounced activation of glycolytic metabolism in both mDCs [
3] and macrophages [
5], as well as a strong activation of epithelial cells [
6] and B cells [
7].
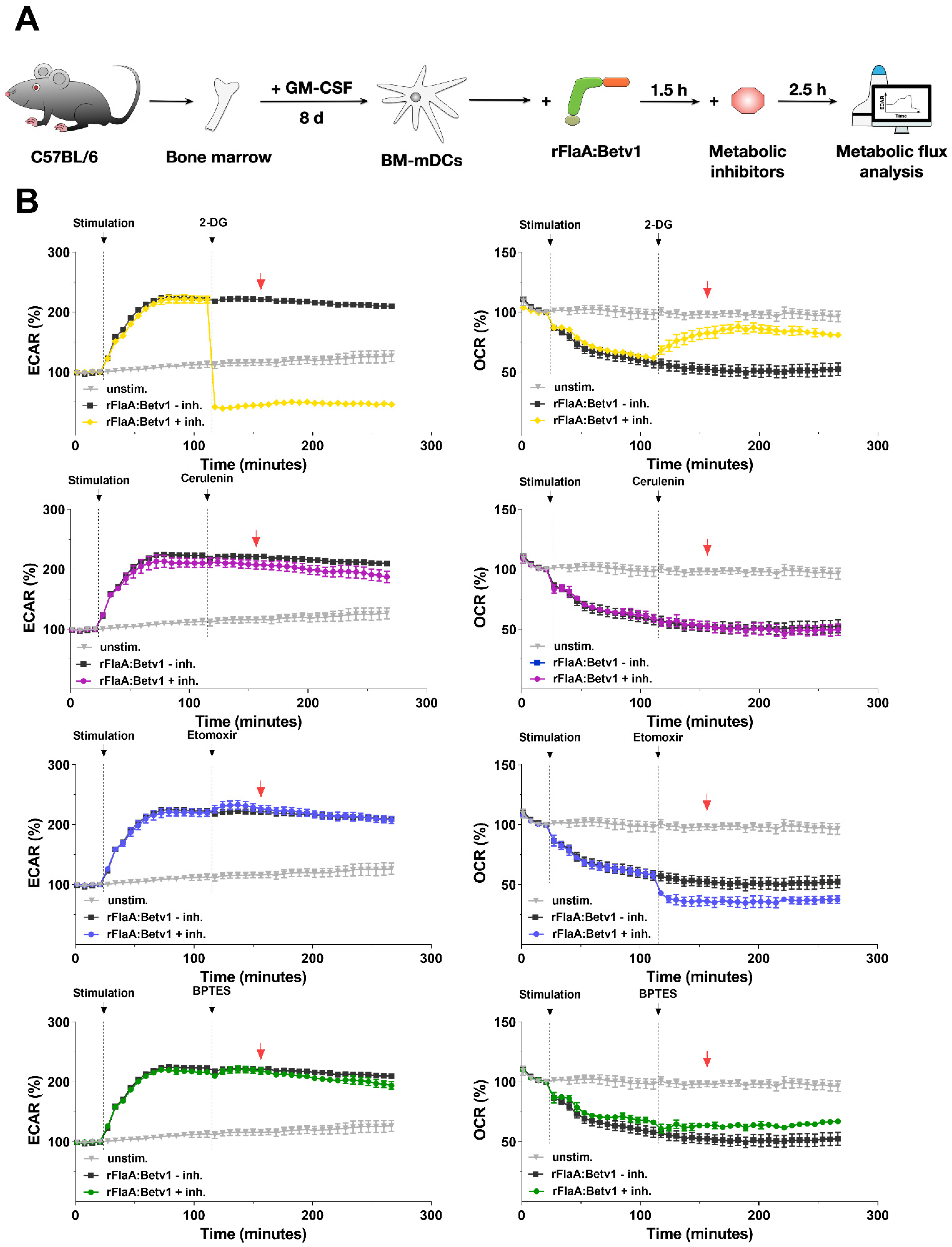
In the present study, we compared the immune–metabolic changes induced by rFlaA:Betv1 in mDCs to other well-established TLR-ligands. In addition to nonreactivity of the investigated mDCs to polyI:C, likely caused by these cells expressing only low levels of the corresponding pattern recognition receptor TLR3 [
18], all other TLR-ligands induced a shift towards glycolytic metabolism (indicated by enhanced ECAR values) while also reducing oxygen consumption (indicated by decreases in OCR) to varying degrees. The same metabolic response was observed for rFlaA:Betv1-stimulated mDCs, making rFlaA:Betv1 a suitable model to study the immune metabolic effects of novel vaccine candidates that incorporate antigens.
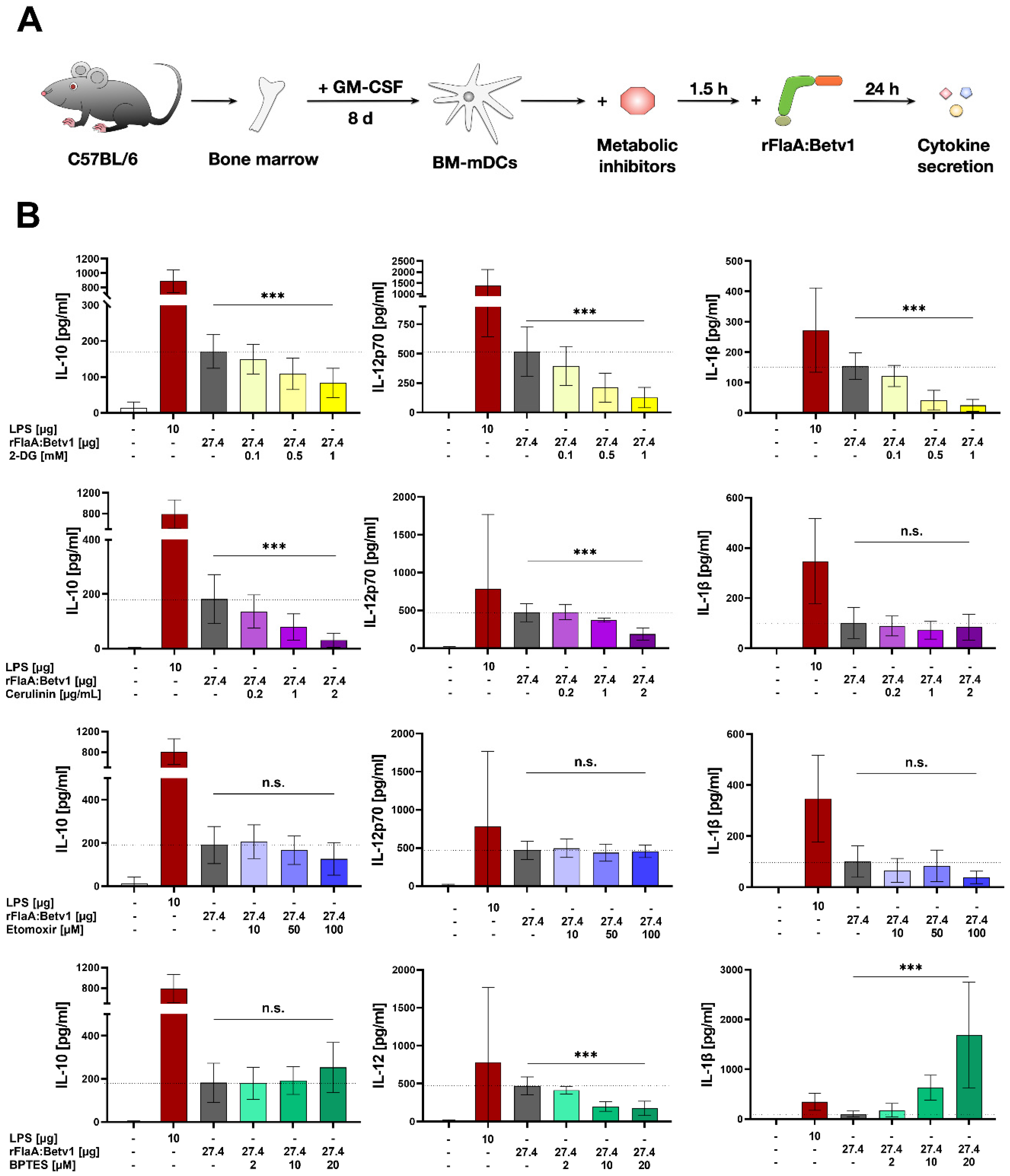
We then further characterized the metabolic phenotype of rFlaA:Betv1-stimulated mDCs in more detail. The increased glycolysis in rFlaA:Betv1-stimulated mDCs was paralleled by a marked decrease in mitochondrial respiration. Furthermore, while inhibition of glycolysis abrogated the observed metabolic switch and dose-dependently suppressed secretion of all investigated cytokines, inhibition of the mitochondrion by antimycin A further enhanced the rFlaA:Betv1-induced metabolic switch towards glycolysis. Here, neither the inhibition of ATP synthase by oligomycin nor inhibition of complex III with antimycin A had an effect on the secretion of the anti-inflammatory cytokine IL-10.
Biochemically, the shift towards preferred lactate production from pyruvate is mediated via a HIF-1α-dependent upregulation of different pyruvate dehydrogenase kinases (PDHK) [
19]. In addition, the upregulation of glycolytic enzymes via HIF-1α activation further increases the speed of glycolysis [
20]. Indeed, Western Blot analyses of rFlaA:Betv1-stimulated mDCs showed a distinct upregulation of HIF-1α both 12 and 24 h post-stimulation.
Under hypoxic conditions, HIF-1α promotes the expression of many genes, but is rapidly degraded by the ubiquitin proteasome pathway under ambient conditions due to its oxygen sensitivity [
21]. In addition, immune cell activation under normoxic conditions can also result in HIF-1α stabilization. For example, Tannahill et al. first reported that LPS-stimulation of macrophages led to the accumulation of the Krebs cycle intermediate succinate via glutamine-dependent anaplerosis and the “GABA-shunt” pathway [
22]. In this context, succinate acted as a cell-intrinsic danger signal, driving both glycolysis and pro-inflammatory IL-1β production via stabilization of HIF-1α (termed “pseudohypoxia”) [
22,
23]. Interestingly, the inhibition of glycolysis using 2-DG decreased the amount of rFlaA:Betv1-induced IL-10 and IL-1β. In contrast, inhibition of HIF-1α activity by either blocking HIF-1α expression using YC-I or enhancing the degradation of HIF-1α by DBA had no impact on the production of IL-1β. At the same time, both inhibitors led to increased secretion of IL-10 (Repos.
Figure S8B). In this experimental setting, only YC-I decreased rFlaA:Betv1-induced IL-12p70 secretion (Repos.
Figure S8B). None of the used HIF-1α-inhibitors significantly affected rFlaA:Betv1-induced antimicrobial activity (Repos.
Figure S8C). These results suggest that, while levels of IL-10 and IL-12p70 are connected to HIF-1α signaling in rFlaA:Betv1-stimulated mDCs, both IL-1β production and antimicrobial activity are independent of HIF-1α and may be triggered by different mechanisms such as mitochondrial stress and the assembly of the inflammasome (see below).
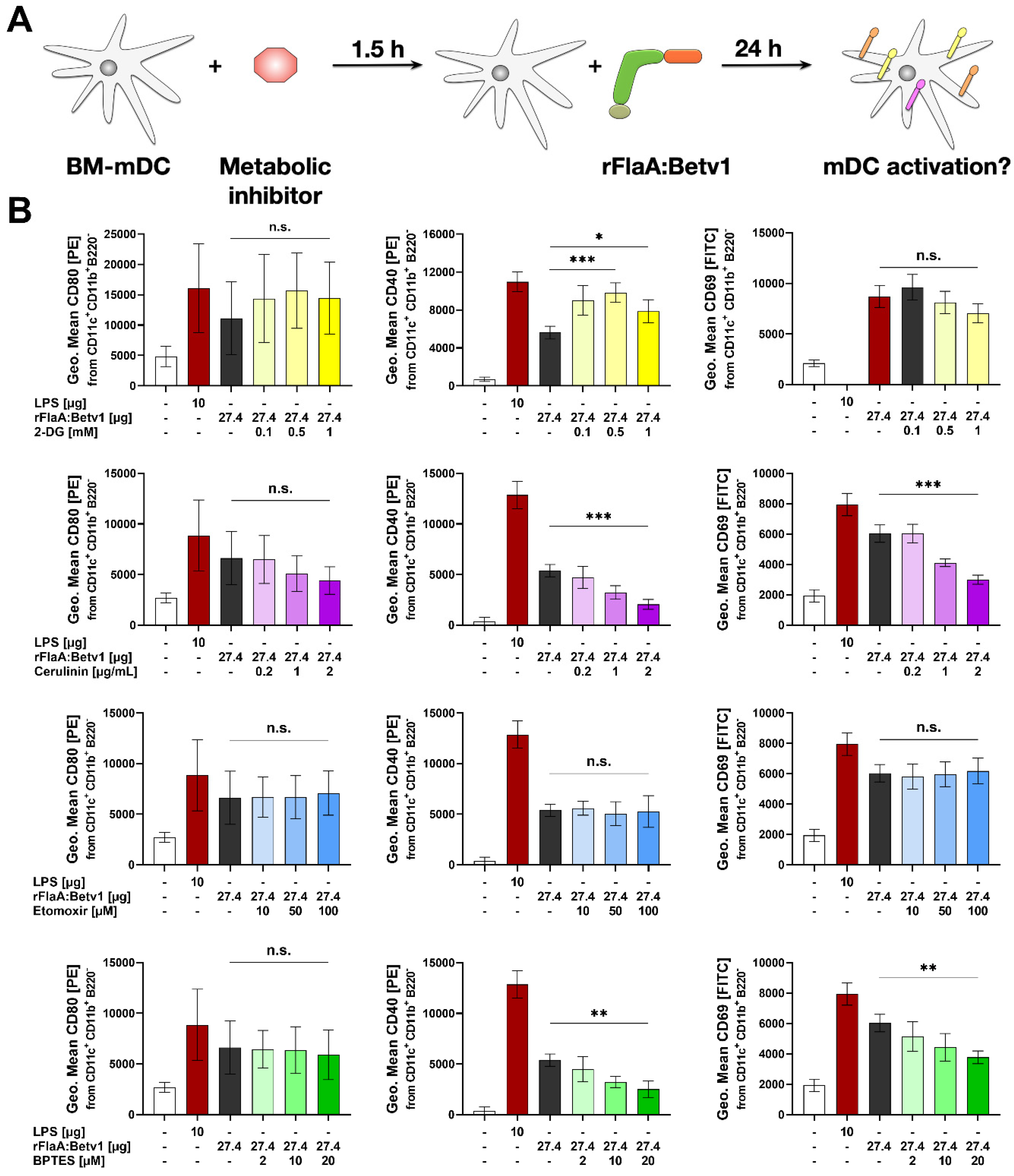
Moreover, inhibition of glutaminase as part of amino acid metabolism by BPTES slightly reduced the rFlaA:Betv1-induced increase in ECAR and slightly increased OCR. rFlaA:Betv1-induced IL-10 secretion was independent of BPTES pretreatment while IL-12p70 was dose-dependently reduced, and IL-1β secretion strongly increased by BPTES. Besides, BPTES also reduced the expression levels of CD40 and CD69.
Mechanistically, BPTES blocks glutaminase (GLS1), which converts glutamine to glutamate and ammonia, leading to an increase in cellular glutamine levels [
24]. As well as being imported into the mitochondrion, glutamine can also be used for the generation of uridine diphosphate-N-acetyl-glucosamine in the cytoplasm, which is an essential substrate for subsequent glycosylation reactions, making the availability of glutamine crucial to maintain proper protein function as well as signal transduction through glycosylation reactions [
25,
26].
In combination with mitochondrial ROS generated by mitochondrial damage, glutamine can further promote the assembly of the NLRP3 inflammasome complex [
27], enhancing IL-1β production. This theory is supported by the fact that, in addition to BPTES, oligomycin pretreatment in rFlaA:Betv1-stimulated mDCs also increased IL-1β production, presumably caused by increased rates of mitochondrial oxidative stress.
In line with this complex network, BPTES was described to have pleiotropic effects affecting numerous metabolic pathways, including glycolysis, the Krebs cycle, as well as nucleotide and amino acid metabolism in two breast cancer cell lines [
28], suggesting the effects of BPTES on overall cellular metabolism to be more complex than the simple inhibition of glutamate generation in the mitochondrion.
We further analyzed the contribution of fatty acid metabolism to the observed activation of mDCs by rFlaA:Betv1. Metabolically, only inhibition of fatty acid synthesis by cerulenin slightly reduced glycolysis (while having no effect on OCR). Moreover, cerulenin pretreatment dose-dependently suppressed the rFlaA:Betv1-induced IL-10- and IL-12p70 secretion, while IL-1β secretion remained unchanged. In contrast, etomoxir pretreatment slightly reduced mitochondrial oxygen consumption and interfered with antimicrobial activity otherwise observed in supernatants of rFlaA:Betv1-stimulated mDCs, but had no impact on the investigated ECAR, cytokine secretion, or expression of co-stimulatory molecules. Etomoxir concentrations greater than 5 µM were recently shown to induce mitochondrial stress associated with ROS production in human T cells independently of its effect on carnitine palmitoyltransferase 1 (CPT1a) [
29]. On the basis of these results, similar off-target effects of etomoxir in mDCs cannot be excluded. Therefore, our results obtained in mDCs pre-treated with etomoxir have to be interpreted cautiously.
It is described that in activated immune cells, a metabolic switch towards glycolysis is usually paralleled by a “disrupted” mitochondrial Krebs cycle resulting from an insufficient supply of the mitochondrion with pyruvate (which is preferentially used to regenerate NAD via the generation of lactate). This “disrupted” Krebs cycle was shown to facilitate the generation of both directly and indirectly antimicrobial molecules such as prostaglandins, nitric oxide (NO), ROS, or itaconate, all of which are important immune effector molecules (reviewed in [
15]).
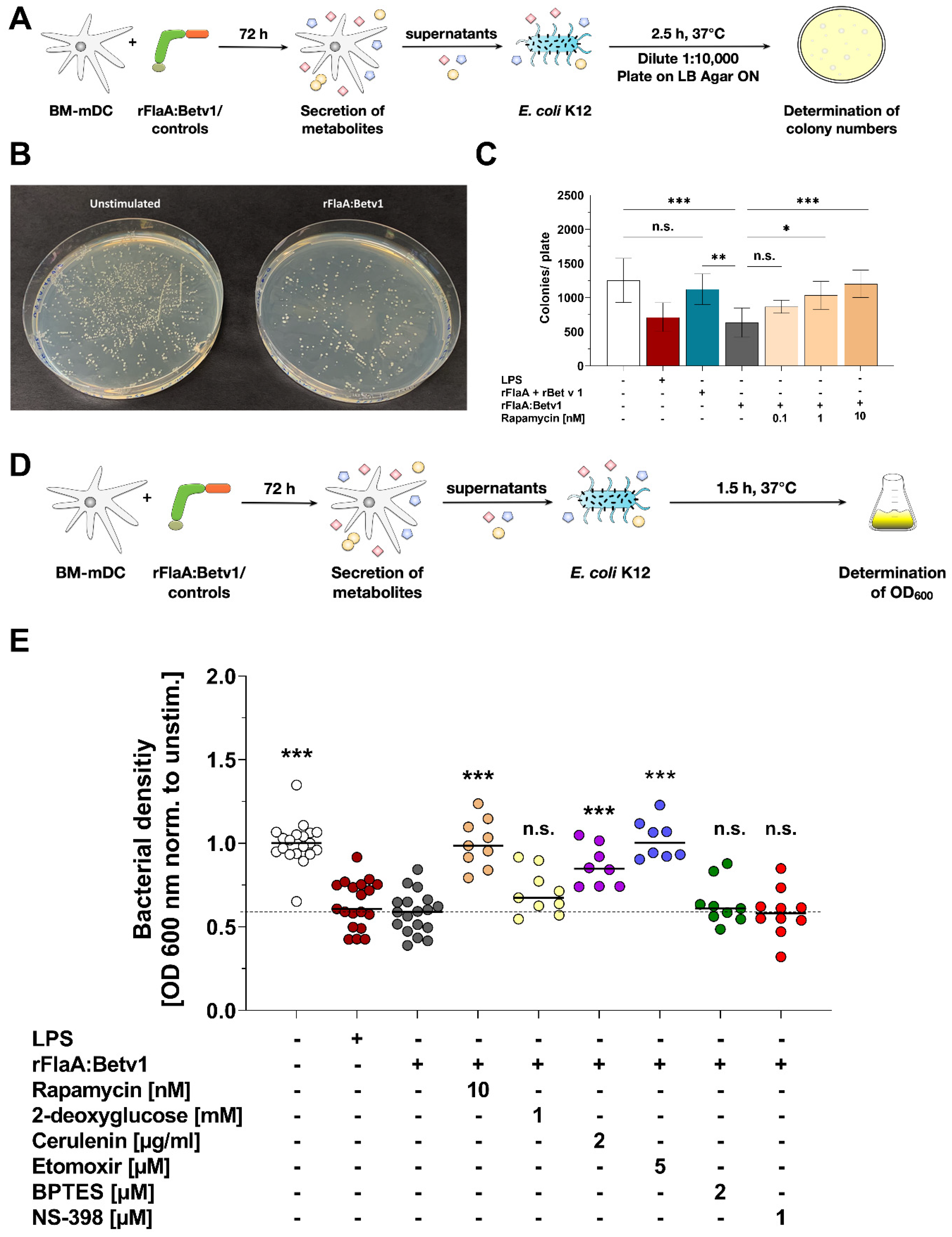
Indeed, we observed a direct antimicrobial activity of supernatants derived from rFlaA:Betv1-stimulated mDCs on E. coli K12 cultures. This antimicrobial activity was shown to depend on both mTOR signaling and fatty acid metabolism as inhibition of either fatty acid oxidation, fatty acid synthesis, or mTOR was shown to prevent mDCs from producing these antimicrobial molecules.
Currently, the exact mechanisms of the observed antimicrobial activity are unclear. Recently, mTOR-dependent autophagy has emerged as an innate immune response pathway that contributes to antimicrobial activity by targeting cytosolic bacteria to autophagosomes in order to restrict bacterial growth (reviewed in [
30]). Therefore, mTOR-dependent autophagy of the bacteria inside the rFlaA:Betv1-stimulated mDCs could contribute to antibacterial responses.
Additionally, as we still observed antimicrobial activity in cell-free, previously frozen supernatants of rFlaA:Betv1-stimulated mDCs and antibacterial activity was shown to be dependent on both mTOR and fatty acid metabolism, our results suggest that the production of antimicrobial molecules requires the mTOR-dependent switch in cellular metabolism which results in the above-described “disrupted” Krebs cycle. The intermediates accumulating from the “disrupted” Krebs cycle are the building blocks of important immune active and antimicrobial compounds [
31].
One example is the accumulation of citrate, cis-aconitate, and D-isocitrate (caused by decreased expression of the enzyme isocitrate dehydrogenase within the “disrupted” Krebs cycle, which normally converts D-isocitrate to α-ketoglutarate), resulting in the generation of prostaglandins, NO, ROS, or itaconate (reviewed in [
15]). In particular, itaconate, which is produced from citrate by activated DCs (and macrophages) has been reported not only to have antibacterial activity, but also to reduce inflammation by inhibiting succinate dehydrogenase [
32]. Furthermore, a recent study by Jaiswal and colleagues was able to identify itaconate as a key Krebs cycle intermediate with the ability to reduce TH2 cytokine production [
33]. Therefore, itaconate could play a role in the immune-modulating properties of rFlaA:Betv1-stimulated mDCs.
Moreover, the formation of certain antimicrobial metabolites such as prostaglandins depends on fatty acid metabolism. As a precursor, fatty acids are metabolized into either arachidonic acid or eicopentaenoic acid, which are subsequently turned into prostaglandins and thromboxanes via cyclooxygenases 1 and 2 (COX1/2) [
34]. In an initial experiment, we analyzed if the antimicrobial effect of rFlaA:Betv1-stimulated mDC-derived supernatants was caused by the production of prostaglandins, by inhibiting COX2 using NS-398. Here, inhibition of COX2 by NS-398 did not modify the rFlaA:Betv1-induced antimicrobial effect (
Figure 5E) and only slightly reduced IL-12p70 secretion (Repos.
Figure S8B). As the bacterial density of cultures incubated with NS-398 pretreated rFlaA:Betv1-stimulated mDC supernatants was equal to those incubated with simply rFlaA:Betv1-stimulated mDC supernatants, we suggest that molecules other than prostaglandins generated from Krebs cycle intermediates, such as, e.g., itaconate, ROS, or NOS, could be responsible for the antimicrobial activity observed in rFlaA:Betv1-stimulated mDC supernatants. In addition, arachidonic acid and other unsaturated fatty acids can also directly function as endogenous antimicrobial molecules [
35]. Therefore, the observed dependency of antimicrobial factor secretion on both mTOR activation and fatty acid metabolism is in line with the reported biochemical alterations occurring in predominantly glycolytic, activated innate immune cells. Nevertheless, we could show the stimulation of mDCs with rFlaA:Betv1 to result in the secretion of antimicrobial molecules, but did not identify the respective substances. Elucidating which molecules mediated the observed antimicrobial activity will be the focus of additional studies.
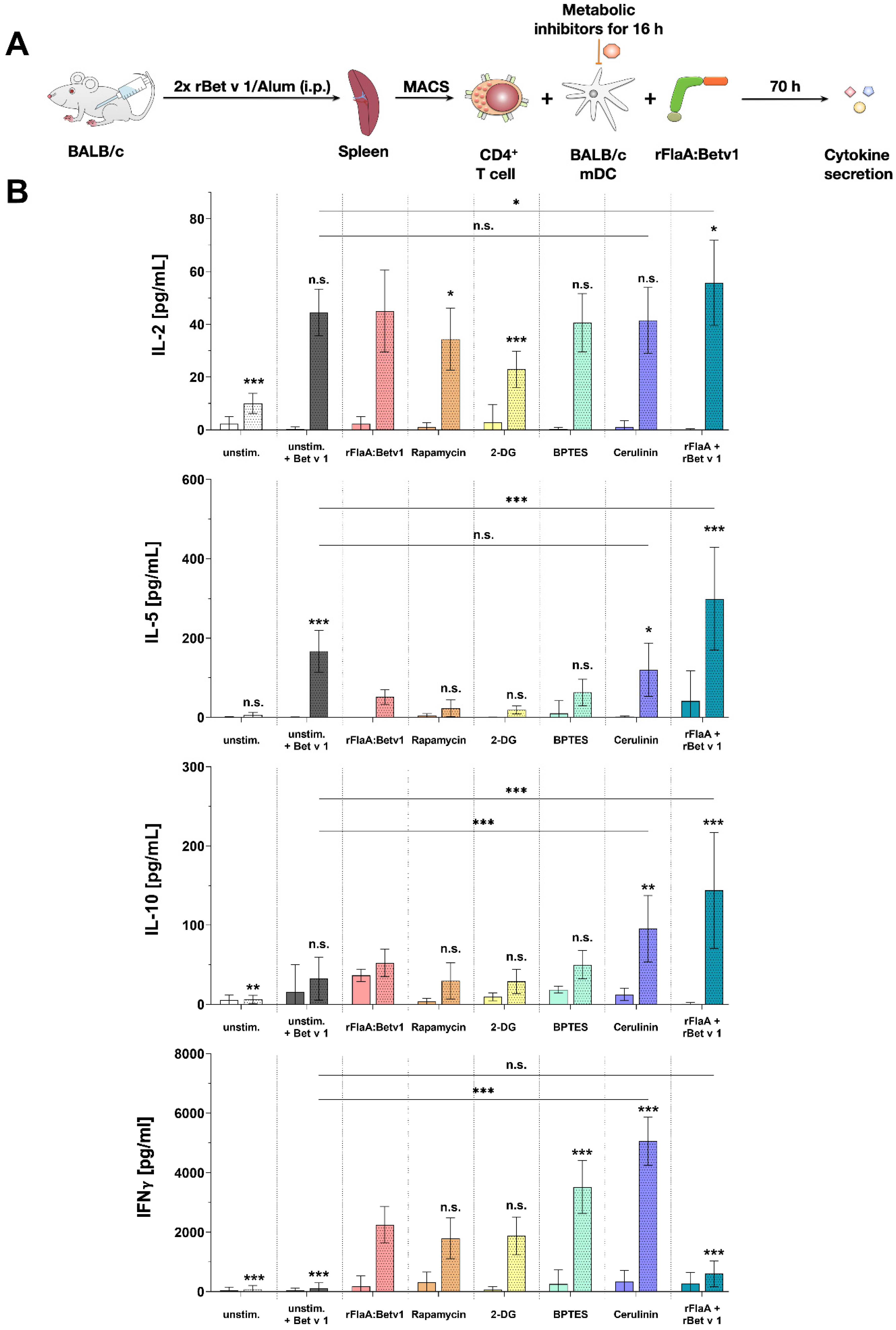
Finally, we investigated the contribution of the different mDC metabolic pathways to their capacity to modulate antigen-specific T cell responses. As reported before, the addition of rFlaA:Betv1 to rBet v 1-stimulated co-cultures was shown to suppress the Bet v 1-induced secretion of the TH2 cytokine IL-5 while maintaining IL-2, IFN-γ, and IL-10 secretion [
3]. Interestingly, only inhibition of fatty acid metabolism by cerulenin pretreatment of rFlaA:Betv1-stimulated mDCs reversed the suppression of IL-5 production while increasing secretion levels of both IL-10 and IFN-γ. Furthermore, IFN-γ secretion in co-cultures was also increased by inhibition of either mTOR, glucose, or amino acid metabolism. Collectively, these results suggest that fatty acid metabolism and its derived metabolites in mDCs with a “disrupted” Krebs cycle are highly important for the suppression of TH2 responses.
One shortcoming of our study is that we did not analyze the CD4+ T cells used for the co-culture experiments in more detail. While we observed an induction of both TH1 and TH2-cytokine secretion after re-stimulation of the T cells ex vivo, showing that the sensitization induced Bet v 1-specific T cells, we cannot comment on the frequency of allergen-specific T cells. Furthermore, we did not analyze a potential induction of regulatory T cells by the rFlaA:Betv1-stimulated mDCs. These parameters will be the focus of future studies.
Although we have only examined acute mDC activation in our study, the strong activation of mDCs by the fusion protein in conjunction with the distinct changes in mDC metabolism (which were comparable to other well-established TLR-ligands already described to induce trained immune responses) allows us to speculate that rFlaA:Betv1 may be able to induce trained immune responses. Investigating if such trained immune responses are indeed induced and may mediate heterologous protection against other antigens upon in vivo application of flagellin-containing fusion proteins will be a focus of future studies.
4. Material and Methods
4.1. Mice
BALB/c and C57Bl/6 (Jackson Laboratories, Bar Harbor, ME, USA) were bred under specific pathogen-free (SPF) conditions at the animal facility of the Paul-Ehrlich-Institut. All animal experiments were performed according to the German animal protection law (granting authority: RP Darmstadt, Germany; approval no. F107/1049).
4.2. Generation of Recombinant Proteins
rFlaA from
L. monocytogenes (accession no: NC_003210) was generated according to Schülke et al. [
36], recombinant birch pollen allergen Bet v 1 (accession no: X15877.1) according to the method of Siebeneicher et al. [
37], and the fusion protein of rFlaA and rBet v 1 (rFlaA:Betv1) was generated according to Schülke et al. [
3] using cDNA fusion with cDNAs of both rFlaA and rBet v 1 as templates.
4.3. TLR-Ligands
LPS was obtained from Sigma-Aldrich (derived from Salmonella abortus equi, L5886, Steinheim, Germany). All other used TLR-ligands (FSL-1, poly I:C, MPLA, and R848) were obtained from InvivoGen (Toulouse, France). rFlaA and rFlaA:Betv1 were generated recombinantly (see above). TLR-ligands were used, if not stated otherwise, in the following concentrations: FSL-1 1 µg/mL; poly I:C 10 µg/mL, LPS 10 µg/mL; MPLA 10 µg/mL; rFlaA 17.4 µg/mL; rFlaA:Betv1 27.4 µg/mL; or R848 1 µg/mL. mDCs were stimulated with the different TLR-ligands for either 24 or 72 h as independent stimulations and analyzed for cytokine secretion.
4.4. Differentiation and In Vitro Stimulation of mDCs
Mouse mDCs were generated as described previously [
36]. Bone marrow cells from either C57Bl/6 or BALB/c mice were cultured in culture medium (RPMI1640, Gibco, Karlsruhe, Germany, supplemented with 10% FCS, 1 mM sodium pyruvate, 10 mM HEPES, penicillin (100 U/mL), streptomycin (100 µg/mL), and 0.1 mM 2-mercaptoethanol, and 100 ng/mL recombinant mouse GM-CSF) for 8 days, and medium was changed every 2 days. On day 8, mDCs were harvested by pipetting and either analyzed by Metabolic Flux Analysis (details see below) or seeded at a concentration of 5 × 10
5 cells/mL in 24-well plates (Thermo Scientific, Dreieich, Germany) in culture medium without the supplementation of GM-CSF.
4.5. ELISA Measurements
Cytokine concentrations in supernatants were analyzed by ELISA using the following antibody combinations: IL-2: purified anti-mouse IL-2 (clone: JES6–1A12, 1:500) and biotin anti-mouse IL-2 (JES6–5H4, 1:500, both BioLegend, San Diego, CA, USA); IL-10: mouse IL-10 ELISA Development Kit (#900-T53, PeproTech, Hamburg, Germany); IL-1β: purified anti-mouse IL-1β (clone: B122, 1:500) and biotin anti-mouse IL-1β (polyclonal, 1:500); IL-5: purified anti-mouse IL-5 (clone: TRFK5, 1:500) and biotin anti-mouse IL-5 (clone: TRFK4, 1:500); IL-12: purified anti-mouse IL-12p70 (clone: C18.2, 1:500) and biotin anti-mouse IL-12p70 (clone: C17.8, 1:500); IFN-γ: purified anti-mouse IFN-γ (clone: XMG1.2, 1:1000) and biotin anti-mouse IFN-γ (clone: R4-6A2, 1:500); IL-13: purified anti-mouse IL-13 (clone: eBio13A; 1:500) and biotin anti-mouse IL-13 (clone: eBio1316H, 1:250, all eBioscience, Frankfurt, Germany).
4.6. Flow Cytometry
rFlaA:Betv1-induced mDC activation was analyzed by flow cytometry using anti-mouse PE-conjugated CD40 (clone: 1C10, dilution: 1 to 100), PE-conjugated CD69 (clone: H1.2F3, dilution: 1 to 150), FITC-conjugated CD80 (clone: 16–10A1, dilution: 1 to 50, all eBiosciences, Frankfurt, Germany). Additionally, cells were stained with anti-mouse pacific blue-conjugated CD11b (clone: M1/70.15, dilution: 1 to 50, Invitrogen, Thermo Fisher Scientific), allophycocyanin-conjugated CD11c (clone: HL3, dilution: 1 to 500, BD Bioscience), and PE-Cy5-conjugated B220 (clone: RA3–6B2, dilution: 1 to 100, BD Bioscience). The FITC- or PE-intensity of CD11b+CD11c+B220− cells (mDCs) was quantified by flow cytometry using a FORTESSA flow cytometer (BD Bioscience). Geometrical mean fluorescence intensities were calculated and normalized to unstimulated cells. Data were analyzed using FlowJo V.7 (Treestar Inc., Ashland, OR, USA).
4.7. Metabolic Inhibitor Experiments
If indicated, mDCs were pretreated for 90 min with either 0.1 or 1 µg/mL antimycin A from Streptomyces sp. (electron transport chain complex III inhibitor), 0.02, 0.2, or 2 µM oligomycin from Streptomyces diastatochromogenes (inhibitor of ATP synthase), 1 µM NS-398 (COX-2 inhibitor, involved in the conversion of arachidonic acid to prostaglandin H2), 0.1, 1, or 10 nM rapamycin (mTOR inhibitor), 2, 10, or 20 µM BPTES (glutaminase inhibitor), 0.2, 1, or 2 µg/mL cerulenin (inhibitor of fatty acid synthase), 10, 50, or 100 µM etomoxir (inhibitor of carnitine palmitoyltransferase 1 (CPT1a) involved in fatty acid oxidation, all Sigma-Aldrich, Steinheim, Germany), 0.1, 0.5, or 1 mM 2-deoxyglucose (2-DG, hexokinase-2 inhibitor, Carl-Roth Laboratory Supplies, Karlsruhe, Germany), 0.1, 1, or 10 µM YC-I (post-transcriptional inhibitor of HIF-1α expression), or 1, or 10 µM dimethyl-bisphenol A (DBA, HIF-1α inhibitor, promoting degradation of HIF-1α protein, both Abcam). Subsequently, mDCs were either left unstimulated or stimulated as indicated for either 24 h (to determine cytokine secretion by ELISA or expression of surface markers by flow cytometry) or 72 h (determination of cytokine secretion, Warburg Effect, glucose consumption, and antimicrobial activity). Stimulation was done with either 17.4 µg/mL rFlaA + 10 µg/mL rBet v 1, or 27.4 µg/mL rFlaA:Betv1. LPS (10 µg/mL, L5886, Sigma Aldrich, Taufkirchen, Germany) was used as a positive control. The toxicity of the used inhibitors was determined using the fixable viability dye eFlour780 (Thermo Fisher Scientific, data not shown). Inhibitor concentrations showing toxic effects were excluded from the analyses.
4.8. Western Blot
1.5 × 106 mDCs were seeded and starved for 2 h in RPMI plus 1% of FBS in 6-well plates (Thermo Scientific), and then stimulated with either 10 µg/mL LPS, 17.4 µg/mL rFlaA + 10 µg/mL rBet v 1, or 27.4 µg/mL rFlaA:Betv1 for the indicated durations. Cells were harvested, lysed with lysis buffer (62.5 mM Tris-HCl (pH 6.8), 2% w/v SDS, 10% glycerol, 50 mM DTT, 0.01% w/v bromophenol blue), and proteins were separated by SDS-polyacrylamide gel followed by transfer to nitrocellulose membranes. The expression of hypoxia-inducible factor 1α (HIF-1α) was analyzed using: 1st Ab: anti-HIF-1α antibody (Cell Signaling Technologies (CST), Cat #14179S, 1:1000) and 2nd Ab: anti-rabbit IgG, HRP-conjugated antibody (CST, Cat #7074, 1:5000). β-tubulin was detected using: HRP-conjugated anti-β-tubulin (CST, Cat #5346, 1:5000). Images were captured with an iBright™ CL1500 system (Thermo Fischer Scientific), and the intensities of band were quantified using ImageJ software (imagej. nih.gov, version: 1.52a).
4.9. Generation of TH2-Biased rBet v 1-Specifc CD4+ T Cells
For the generation of Bet v 1-specific, TH2-biased CD4+ T cells, BALB/c mice were sensitized by two i.p. injections of 10 µg rBet v 1 plus 2 mg aluminum hydroxide (alum; InjectTM Alum, Thermo Scientific, Dreieich, Germany) two weeks apart. One week after the second sensitization, mice were sacrificed, and splenocytes were used for CD4+ T cell isolation using the CD4 T Cell Isolation Kit according to the manufacturer’s recommendations (Miltenyi Biotec, Bergisch Gladbach, Germany).
4.10. mDC:TC Co-Cultures
First, 1.6 × 10
5 BALB/c mDCs were seeded in 48-well plates (Thermo Scientific). mDCs were treated with different metabolic inhibitors (5 nM rapamycin, 1 µM BPTES, 0.5 mM 2-deoxyglucose, or 0.2 µg/mL cerulenin) for 16 h. Subsequently, plates were washed by centrifugation at 1200 rpm for 5 min at 4 °C, media were removed carefully through pipetting, and 250 µL of fresh RPMI culture media was added. T cells were isolated from rBet v 1/alum-sensitized BALB/c mice as described above and 6.3 × 10
5 CD4
+ T cells were added to the mDCs. Co-cultures were stimulated with either 10 µg/mL LPS or 13.7 µg/mL rFlaA:Betv1. Co-cultures were subsequently either left unstimulated or re-stimulated with 2 µg/mL of rBet v 1 for 72 h. The total culture volume was adjusted to 500 µL. Cytokine secretion in culture supernatants was determined by ELISA. Please note that unstimulated and rBet v 1-stimulated controls shown in
Figure 6 were shared with a previously published dataset [
38] for 3R reasons, while all other results contained in
Figure 6 were previously unpublished.
4.11. Determination of Antimicrobial Activity
To determine the antimicrobial activity of supernatants derived from mDCs after 72 h of culture, overnight cultures (ONC) of Escherichia coli (E. coli) K12 were prepared. To achieve this, 10 mL of Luria-Bertani (LB) medium was inoculated with 5 μL of an E. coli K12 glycerol stock in a 50 mL falcon tube and incubated overnight at 180 rpm and 37 °C. The next day, the OD600nm of the ONC was measured and adjusted to an OD600nm of 0.75 with LB medium. 75 μL of the adjusted ONC were added to each well of a flat bottom 96-well plate except for 4 wells that only contained LB medium as a blank. After the addition of 75 μL of samples to the respective ONC wells, the plates were incubated for 120 min (80 rpm, 37 °C). Subsequently, the OD600nm was measured with a SpectraMaxPlus384 microplate reader (Molecular Devices, San Jose, CA, USA). For determination of bacterial growth via colony counting, ONCs of E. coli K12 were adjusted to an OD600nm of 1.0 with LB medium. Then, 50 µL of bacteria was mixed with 50 µL of supernatant and incubated for 2.5 h at 37 °C in a thermomixer at 300 rpm. Bacteria were subsequently plated on LB agar plates in 1 to 10,000 dilutions, incubated overnight at 37 °C, and colonies were counted manually.
4.12. Glucose Measurement and Analysis of the Warburg Effect
Glucose concentrations in the harvested cell culture supernatants 72 h post-stimulation were determined using the Glucose (GO) Assay Kit (Sigma-Aldrich) according to [
3]. The metabolic rate was calculated as the percentage of glucose consumption. For this, the measured glucose concentration in the samples was subtracted from the glucose concentration in the stock media (RPMI1640, 2 mg/mL). The Warburg Effect in culture media was determined as 1/OD570 nm, normalized to unstimulated controls according to [
3].
4.13. Metabolic Flux Analysis
For metabolic flux analysis, 8 × 104 mDCs per well were seeded in Seahorse XF96 cell culture microplates (V3-PS, TC-treated, Agilent, Santa Clara, CA, USA). The next day, the medium was exchanged and cells were stimulated as indicated with the different inhibitors, proteins, TLR ligands, or controls. Seahorse XF Real-Time ATP rate assays were performed according to the manufacturer’s recommendations (Agilent). Post measurement, samples were normalized to total protein content via BCA (Thermo Fischer Scientific, Dreieich, Germany) and analyzed using Wave Desktop Software (Agilent) and Graphpad Prism v8 (Graphpad Software, Inc, San Diego, CA, USA). ATP rate assays were analyzed using the respective report generator sheet according to the manufacturer’s recommendations (Agilent).
4.14. Statistical Analysis
Statistical analysis was performed with GraphPad Prism (GraphPad Prism version 9.2.0 for Windows, GraphPad Software, San Diego, CA, USA) for Windows using two-way ANOVA tests adjusted for multiple comparisons according to either Dunnett’s multiple comparison Test (
Figure 6; Repos.
Figures S1B, S2B and S5B), uncorrected Fisher’s LSD (Repos.
Figure S1C,D), or Tukey’s Test (all other figures). For statistically significant results, the following convention was used: *
p < 0.05, **
p < 0.01, and ***
p < 0.001.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





