
Competitive Sperm-Marked Beetles for Monitoring Approaches in Genetic Biocontrol and Studies in Reproductive Biology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
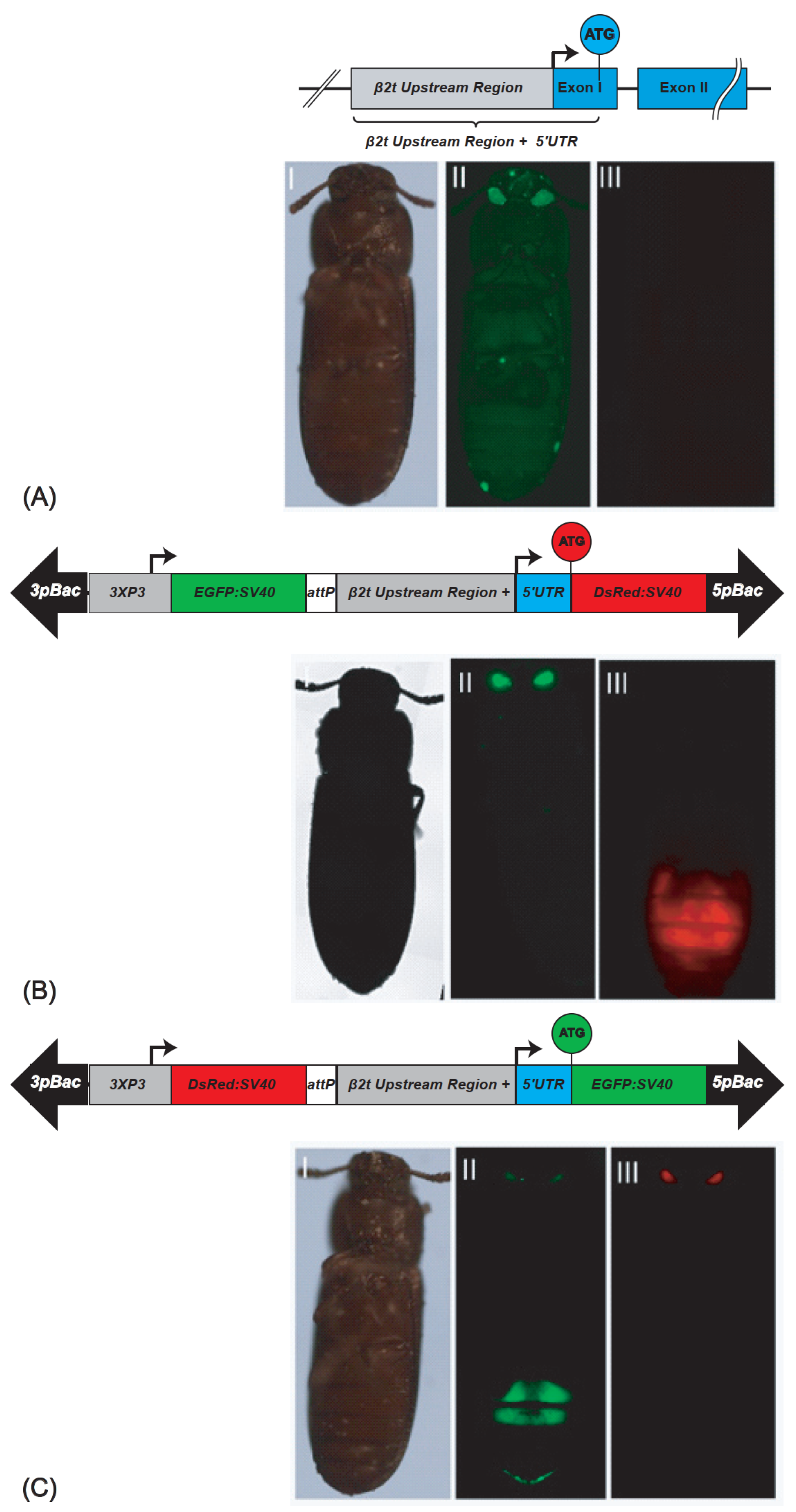
2.1. Generation of Different Tribolium Castaneum Sperm-Marking Lines
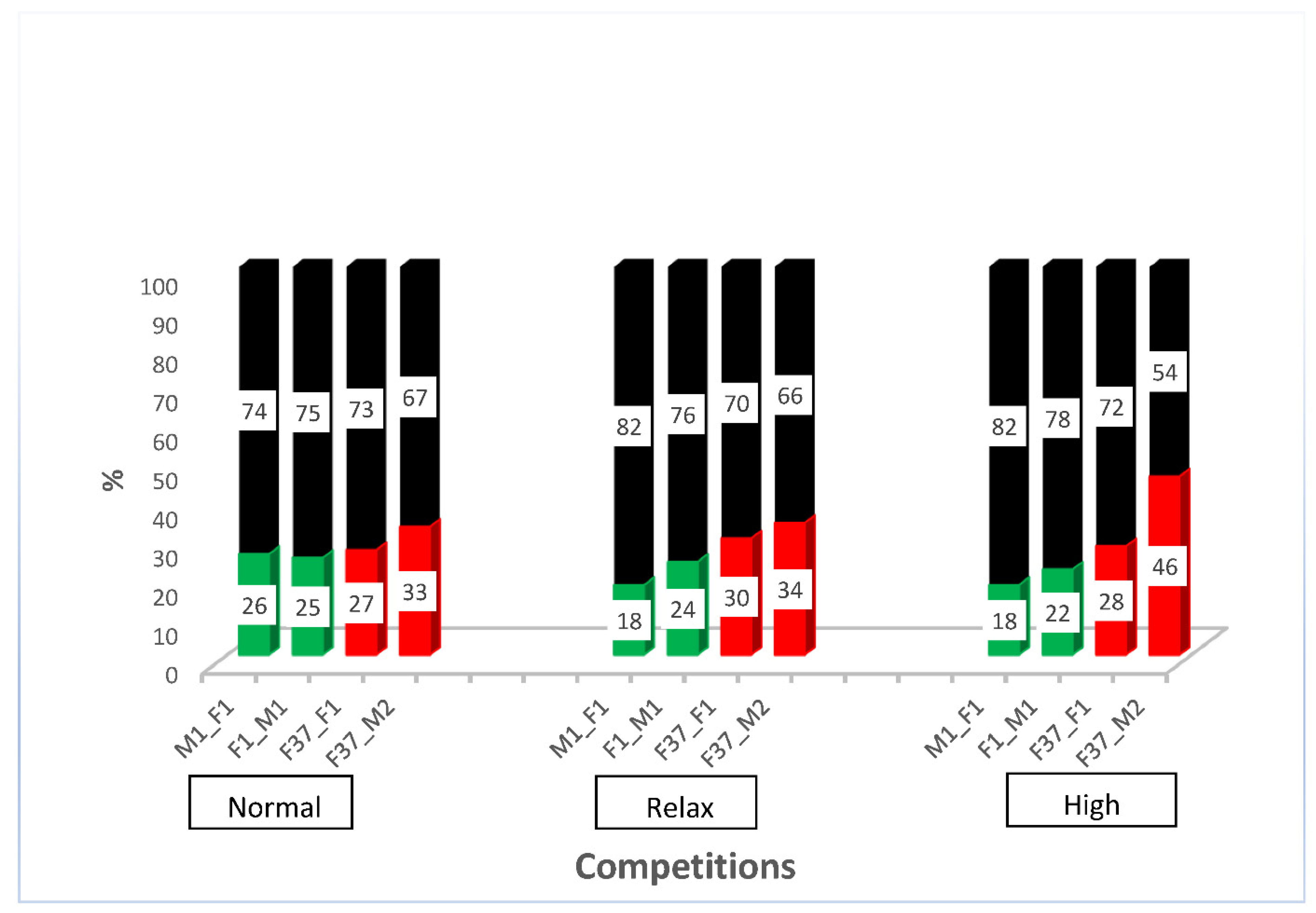
2.2. Competitiveness of Sperm-Marking Lines
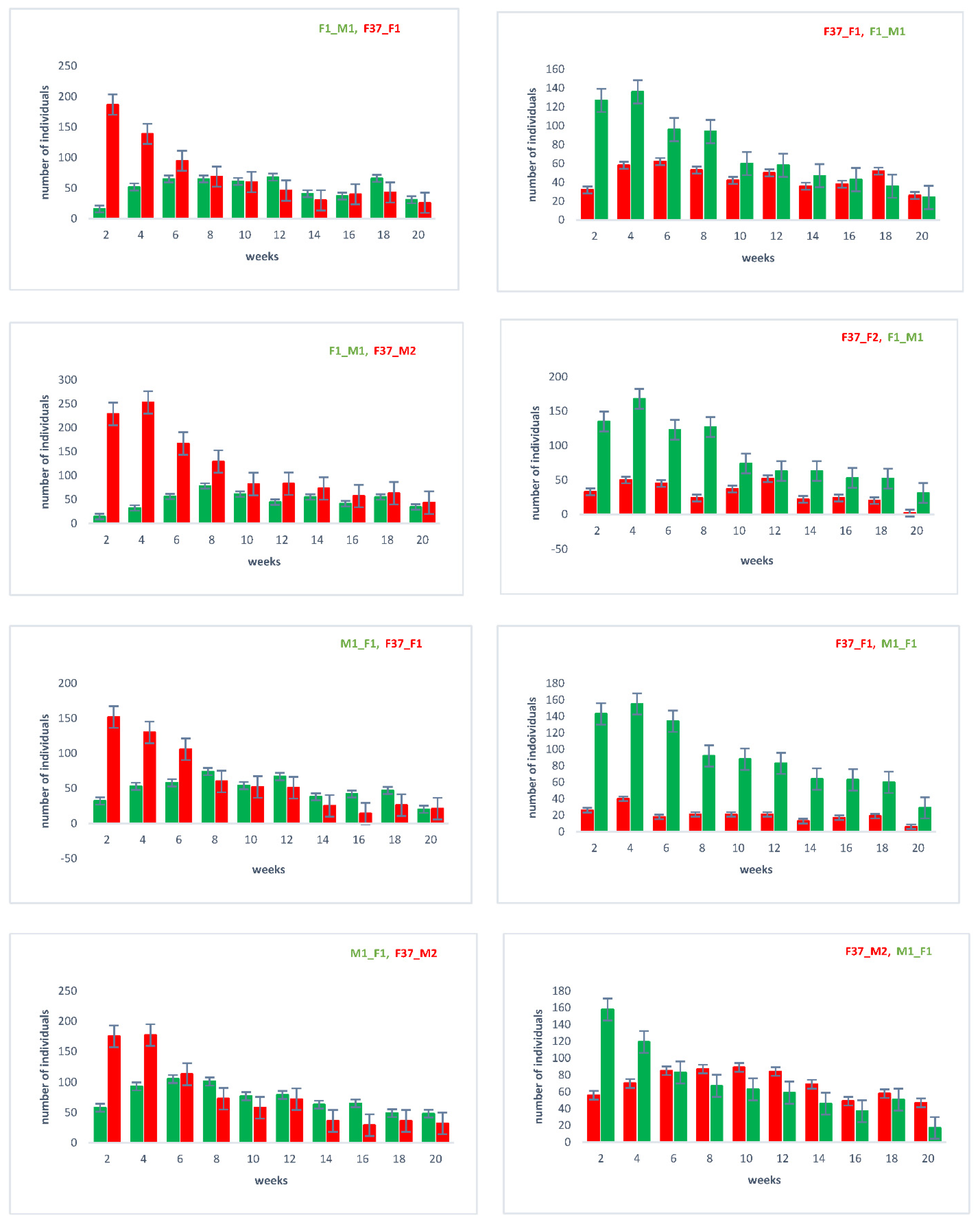
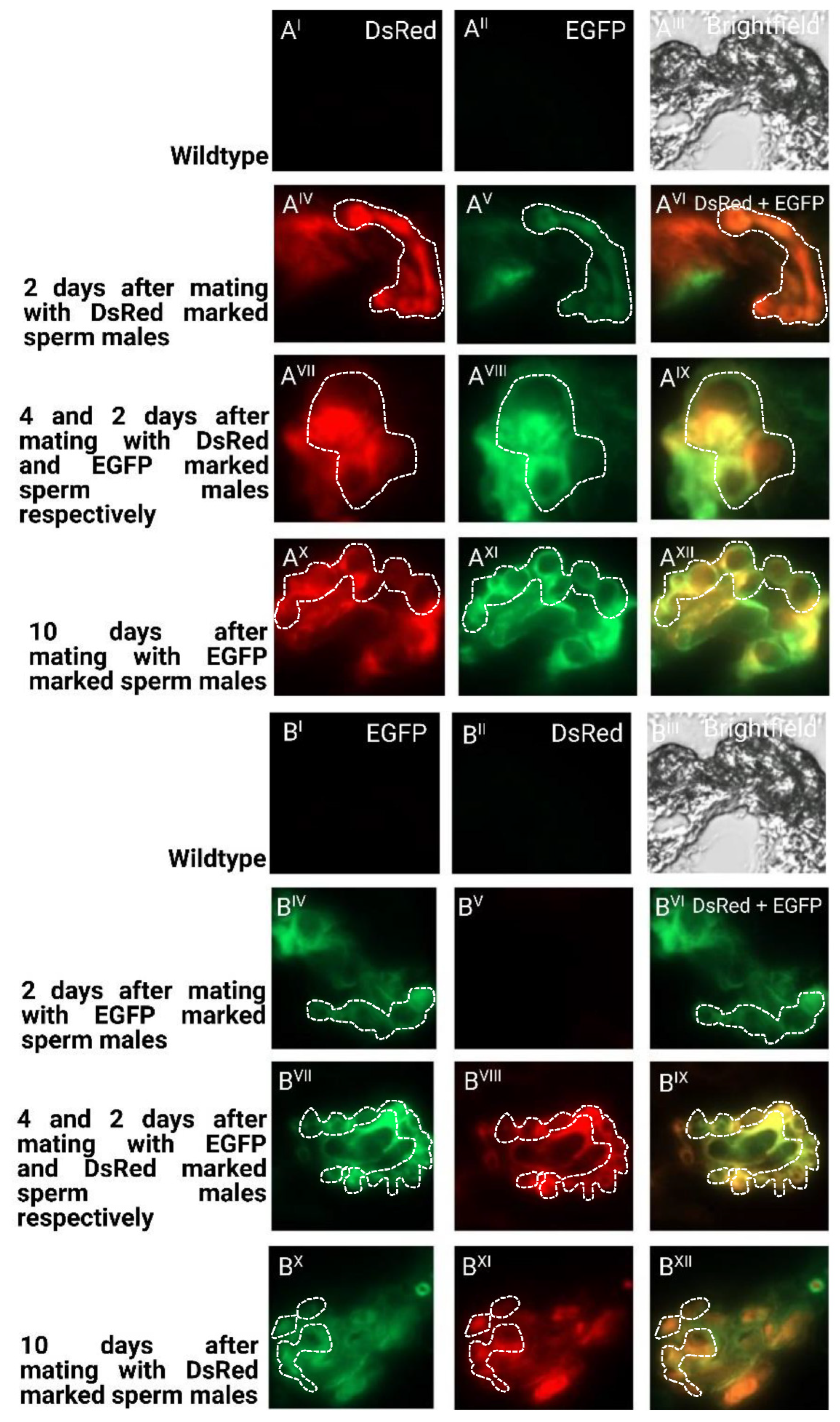
2.3. Sperm Use in Consecutive Matings
3. Discussion
4. Materials and Methods
4.1. Beetle Culture
4.2. Isolation of the β2t Promoter Region and Cloning of the Transformation Vectors
4.3. Germline Transformation and Strain Establishment
4.4. Competitive Assays
4.5. Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scolari, F.; Yuval, B.; Gomulski, L.M.; Schetelig, M.F.; Gabrieli, P.; Bassetti, F.; Wimmer, E.A.; Malacrida, A.R.; Gasperi, G. Polyandry in the medfly—Shifts in paternity mediated by sperm stratification and mixing. BMC Genet. 2014, 15, S10. [Google Scholar] [CrossRef]
- Catteruccia, F.; Benton, J.P.; Crisanti, A. An Anopheles transgenic sexing strain for vector control. Nat. Biotechnol. 2005, 23, 1414–1417. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.C.; Walter, M.F.; Hice, R.H.; O’Brochta, D.A.; Atkinson, P.W. Testis-specific expression of the beta2 tubulin promoter of Aedes aegypti and its application as a genetic sex-separation marker. Insect Mol. Biol. 2007, 16, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Schetelig, M.F.; Bertin, S.; Malacrida, A.R.; Gasperi, G.; Wimmer, E.A. Fluorescent sperm marking to improve the fight against the pest insect Ceratitis capitata (Wiedemann; Diptera: Tephritidae). New Biotechnol. 2008, 25, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Zimowska, G.J.; Nirmala, X.; Handler, A.M. The beta2-tubulin gene from three tephritid fruit fly species and use of its promoter for sperm marking. Insect Biochem. Mol. Biol. 2009, 39, 508–515. [Google Scholar] [CrossRef]
- Ahmed, H.M.M.; Hildebrand, L.; Wimmer, E.A. Improvement and use of CRISPR/Cas9 to engineer a sperm-marking strain for the invasive fruit pest Drosophila suzukii. BMC Biotechnol. 2019, 19, 85. [Google Scholar] [CrossRef] [PubMed]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. (Eds.) Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Klassen, W.; Curtis, C.F.; Hendrichs, J. History of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 1–44. [Google Scholar]
- Enkerlin, W.R. Impact of Fruit Fly Control Programmes using the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 979–1006. [Google Scholar]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Enkerlin, W.R.; Lopez, L.; Celedonio, H. Increased accuracy in discrimination between captured wild unmarked and released dye-marked adults in fruit fly (Diptera: Tephritidae) sterile released programs. J. Econ. Entomol. 1996, 89, 946–949. [Google Scholar] [CrossRef]
- Hagler, J.R.; Jackson, C.G. Methods for marking insects: Current techniques and future prospects. Annu. Rev. Entomol. 2001, 46, 511–543. [Google Scholar] [CrossRef] [PubMed]
- Lance, D.R.; McInnis, D.O. Biological Basis of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 113–142. [Google Scholar]
- Brown, S.J.; Shippy, T.D.; Miller, S.; Bolognesi, R.; Beeman, R.W.; Lorenzen, M.D.; Bucher, G.; Wimmer, E.A.; Klingler, M. The red flour beetle, Tribolium castaneum (Coleoptera): A model for studies of development and pest biology. In Emerging Model Organisms; CSH Protoc: Cold Spring Harbor, NY, USA, 2009. [Google Scholar] [CrossRef]
- Fedina, T.Y.; Lewis, S.M. Evaluating Cryptic Female Choice in Highly Promiscous Tribolium Beetles. In Cryptic Female Choice in Arthropods; Peretti, A.V., Aisenberg, A., Eds.; Springer: Cham, Switzerland, 2015; pp. 431–459. [Google Scholar]
- Siebert, K.S.; Lorenzen, M.D.; Brown, S.J.; Park, Y.; Beeman, R.W. Tubulin superfamily genes in Tribolium castaneum and the use of a Tubulin promoter to drive transgene expression. Insect Biochem. Mol. Biol. 2008, 38, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Michiels, F.; Gasch, A.; Kaltschmidt, B.; Renkawitz-Pohl, R. A 14 bp promoter element directs the testis specificity of the Drosophila beta 2 tubulin gene. EMBO J. 1989, 8, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Horn, C.; Wimmer, E. A versatile vector set for animal transgenesis. Dev. Genes Evol. 2001, 210, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Horn, C.; Schmid, B.G.M.; Pogoda, F.S.; Wimmer, E.A. Fluorescent transformation markers for insect transgenesis. Insect Biochem. Mol. Biol. 2002, 32, 1221–1235. [Google Scholar] [CrossRef]
- Berghammer, A.J.; Klingler, M.; AWimmer, E. A universal marker for transgenic insects. Nature 1999, 402, 370–371. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Jakes, E.; Myles, K.M.; Adelman, Z.N. The β2Tubulin, Rad50-ATPase and enolase cis-regulatory regions mediate male germline expression in Tribolium castaneum. Sci. Rep. 2021, 11, 18131. [Google Scholar] [CrossRef] [PubMed]
- Fedina, T.Y.; Lewis, S.M. Female influence over offspring paternity in the red flour beetle Tribolium castaneum. Proc. R. Soc. London B 2004, 271, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Droge-Young, E.M.; Belote, J.M.; Perez, G.S.; Pitnick, S. Resolving mechanisms of short-term competitive fertilization success in the red flour beetle. J. Insect Physiol. 2016, 93–94, 1–10. [Google Scholar] [CrossRef]
- Bloch Qazi, M.C.; Herbeck, J.T.; Lewis, S.M. Mechanisms of sperm transfer and storage in the red flour beetle (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 1996, 89, 892–897. [Google Scholar] [CrossRef]
- Fedina, T.Y.; Lewis, S.M. Female mate choice across mating stages and between sequential mates in flour beetles. J. Evol. Biol. 2007, 20, 2138–2143. [Google Scholar] [CrossRef] [PubMed]
- Bloch Qazi, M.C.; Aprille, J.R.; Lewis, S.M. Female role in sperm storage in the red flour beetle, Tribolium castaneum. Comp. Biochem. Physiol. A 1998, 120, 641–647. [Google Scholar] [CrossRef]
- Edvardsson, M.; Arnqvist, G. Copulatory courtship and cryptic female choice in red flour beetles Tribolium castaneum. Proc. R. Soc. London B 2000, 267, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.M.; Kobel, A.; Fedina, T.; Beeman, R.W. Sperm stratification and paternity success in red flour beetles. Physiol. Entomol. 2005, 30, 303–307. [Google Scholar] [CrossRef]
- Schmitt-Engel, C.; Schultheis, D.; Schwirz, J.; Ströhlein, N.; Troelenberg, N.; Majumdar, U.; Dao, V.A.; Grossmann, D.; Richter, T.; Tech, M.; et al. The iBeetle large-scale RNAi screen reveals gene functions for insect development and physiology. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schetelig, M.F.; Caceres, C.; Zacharopoulou, A.; Franz, G.; Wimmer, E.A. Conditional embryonic lethality to improve the sterile insect technique in Ceratitis capitata(Diptera: Tephritidae). BMC Biol. 2009, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Eckermann, K.N.; Ahmed, H.M.M.; KaramiNejadRanjbar, M.; Dippel, S.; Ogaugwu, C.E.; Kitzmann, P.; Isah, M.D.; Wimmer, E.A. Hyperactive piggyBac transposase improves transformation efficiency in diverse insect species. Insect Biochem. Mol. Biol. 2018, 98, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://coleoguy.blogspot.com/2011/10/dissecting-tribolium-genitalia.html (accessed on 1 October 2019).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isah, M.D.; Atika, B.; Dippel, S.; Ahmed, H.M.M.; Wimmer, E.A. Competitive Sperm-Marked Beetles for Monitoring Approaches in Genetic Biocontrol and Studies in Reproductive Biology. Int. J. Mol. Sci. 2022, 23, 12594. https://doi.org/10.3390/ijms232012594
Isah MD, Atika B, Dippel S, Ahmed HMM, Wimmer EA. Competitive Sperm-Marked Beetles for Monitoring Approaches in Genetic Biocontrol and Studies in Reproductive Biology. International Journal of Molecular Sciences. 2022; 23(20):12594. https://doi.org/10.3390/ijms232012594
Chicago/Turabian StyleIsah, Musa Dan’azumi, Bibi Atika, Stefan Dippel, Hassan M. M. Ahmed, and Ernst A. Wimmer. 2022. "Competitive Sperm-Marked Beetles for Monitoring Approaches in Genetic Biocontrol and Studies in Reproductive Biology" International Journal of Molecular Sciences 23, no. 20: 12594. https://doi.org/10.3390/ijms232012594
APA StyleIsah, M. D., Atika, B., Dippel, S., Ahmed, H. M. M., & Wimmer, E. A. (2022). Competitive Sperm-Marked Beetles for Monitoring Approaches in Genetic Biocontrol and Studies in Reproductive Biology. International Journal of Molecular Sciences, 23(20), 12594. https://doi.org/10.3390/ijms232012594