
The Structural Interactions of Molecular and Fibrillar Collagen Type I with Fibronectin and Its Role in the Regulation of Mesenchymal Stem Cell Morphology and Functional Activity
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
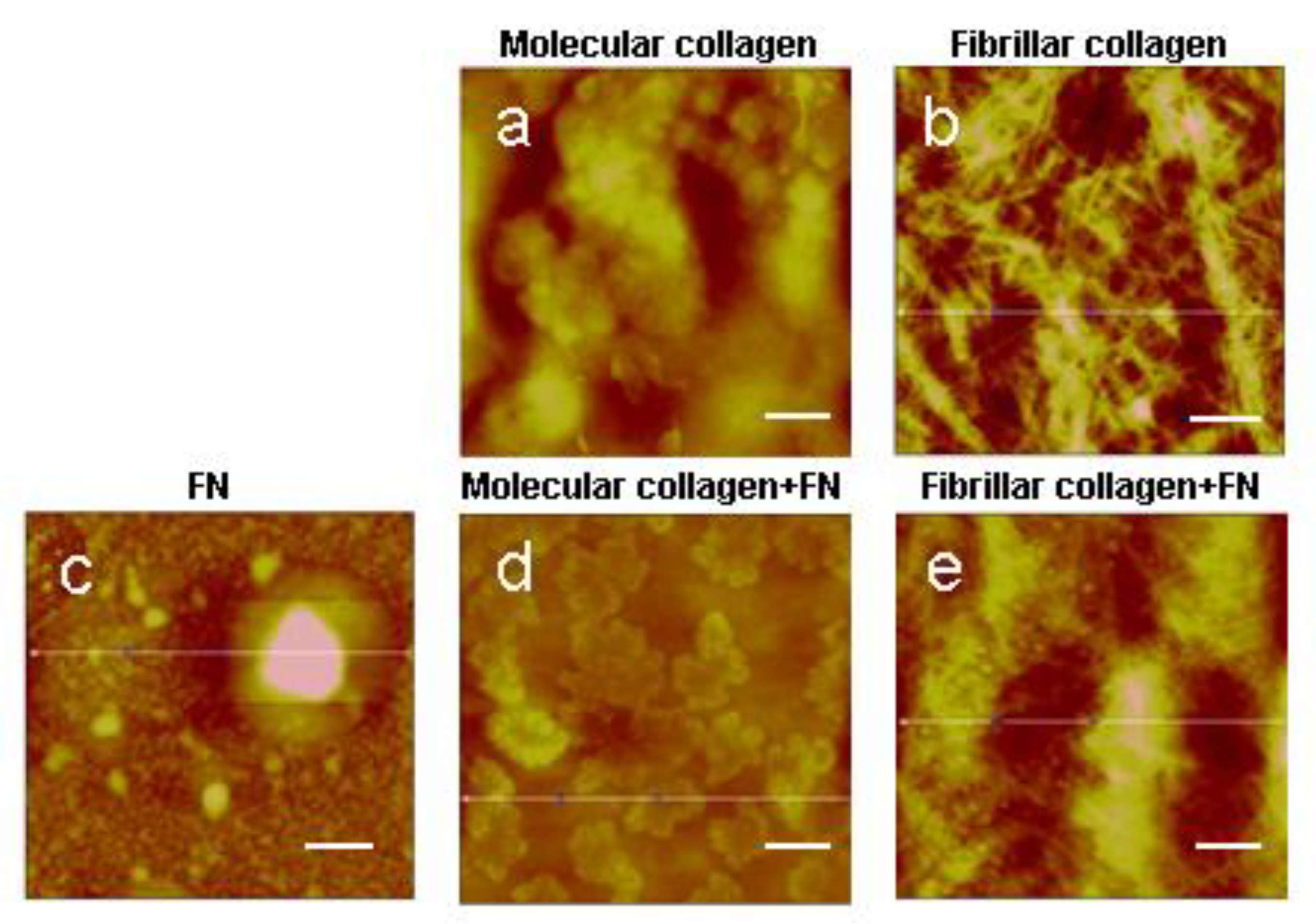
2.1. Atomic Force Microscopy (AFM)
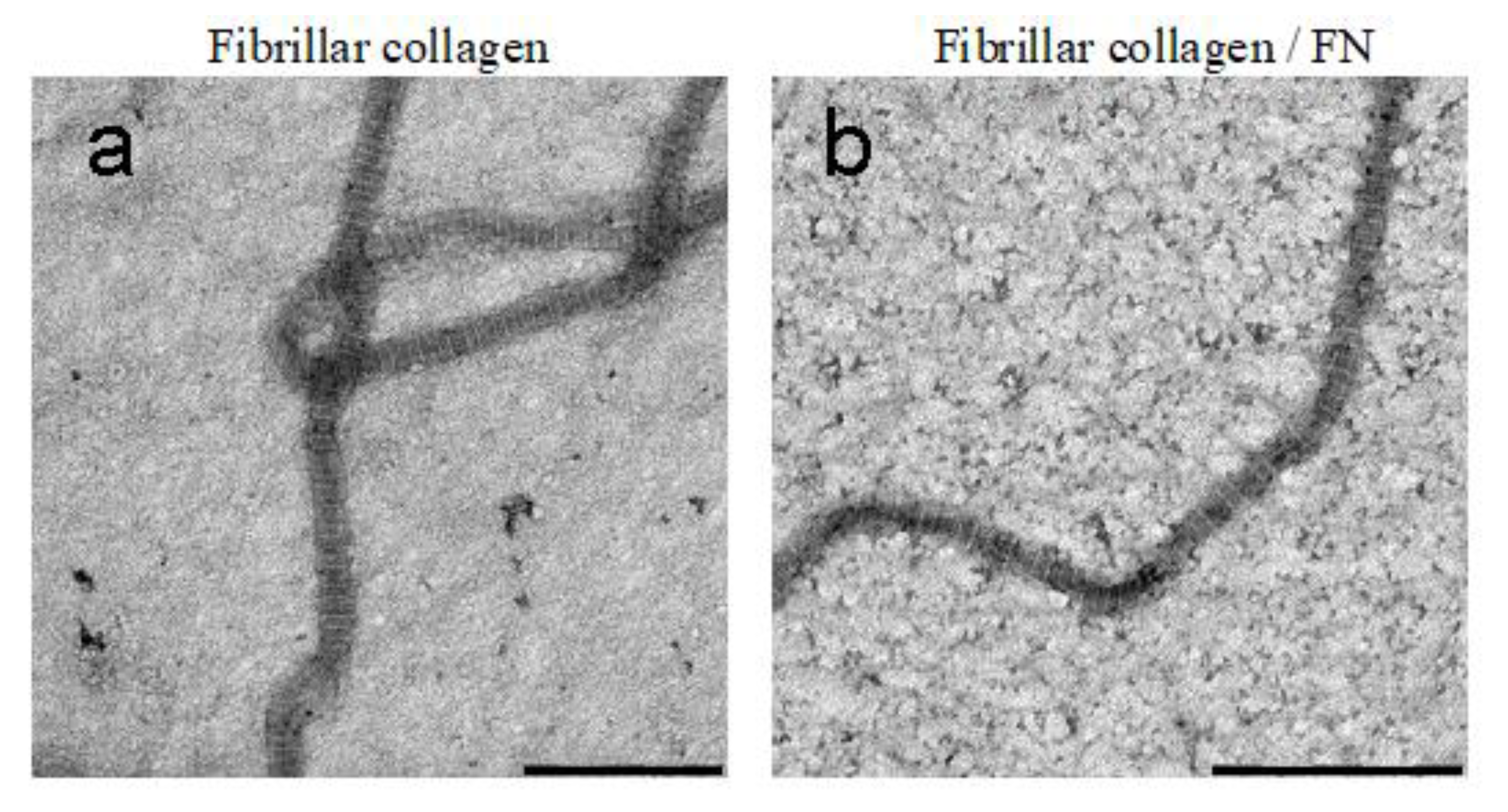
2.2. Transmission Electron Microscopy (TEM)
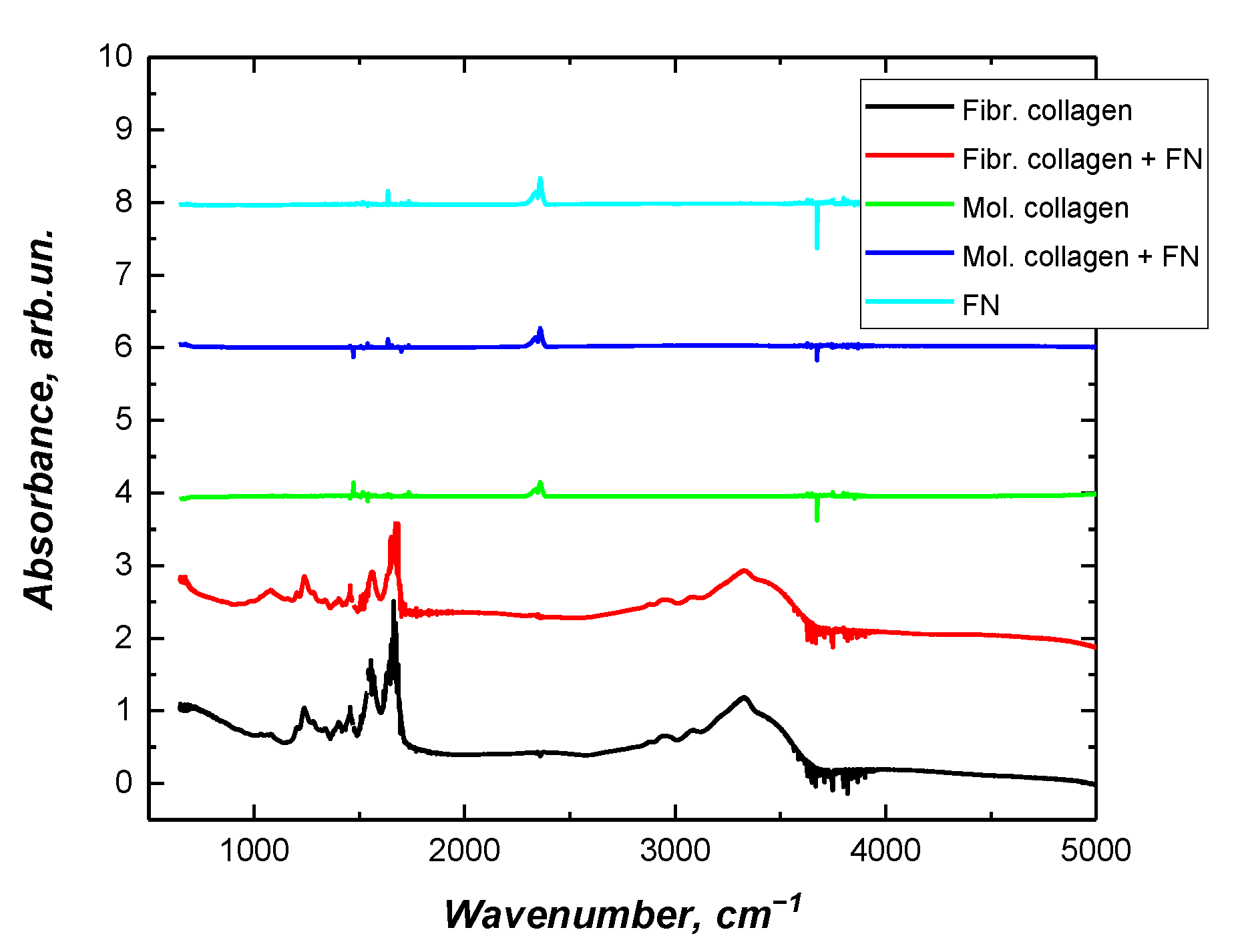
2.3. Fourier Transform Infrared Spectroscopy (FTIR)
2.4. Cell Interaction with Collagen—Fibronectin Surface
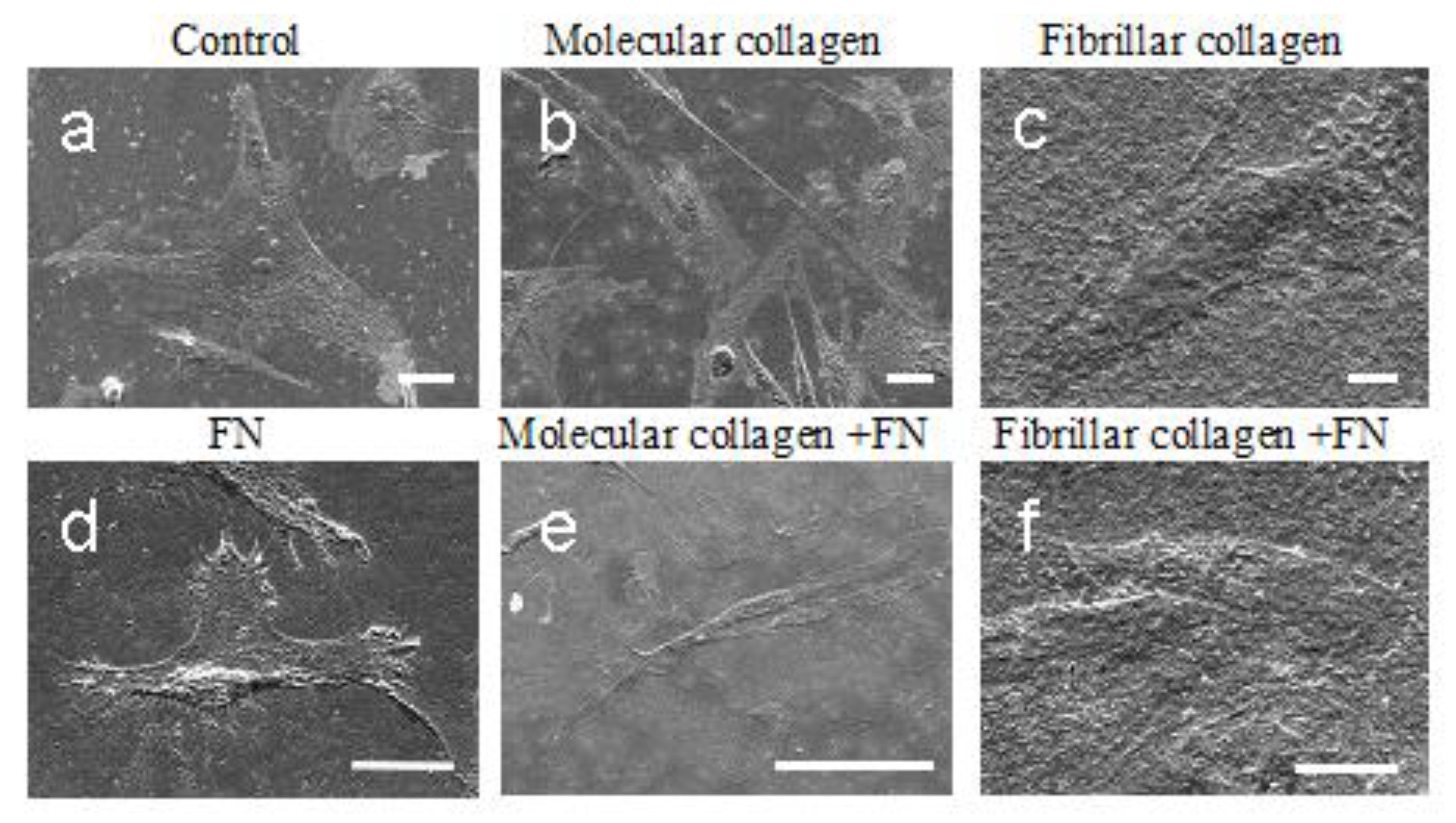
2.4.1. Scanning Electron Microscopy (SEM) Morphological Analysis
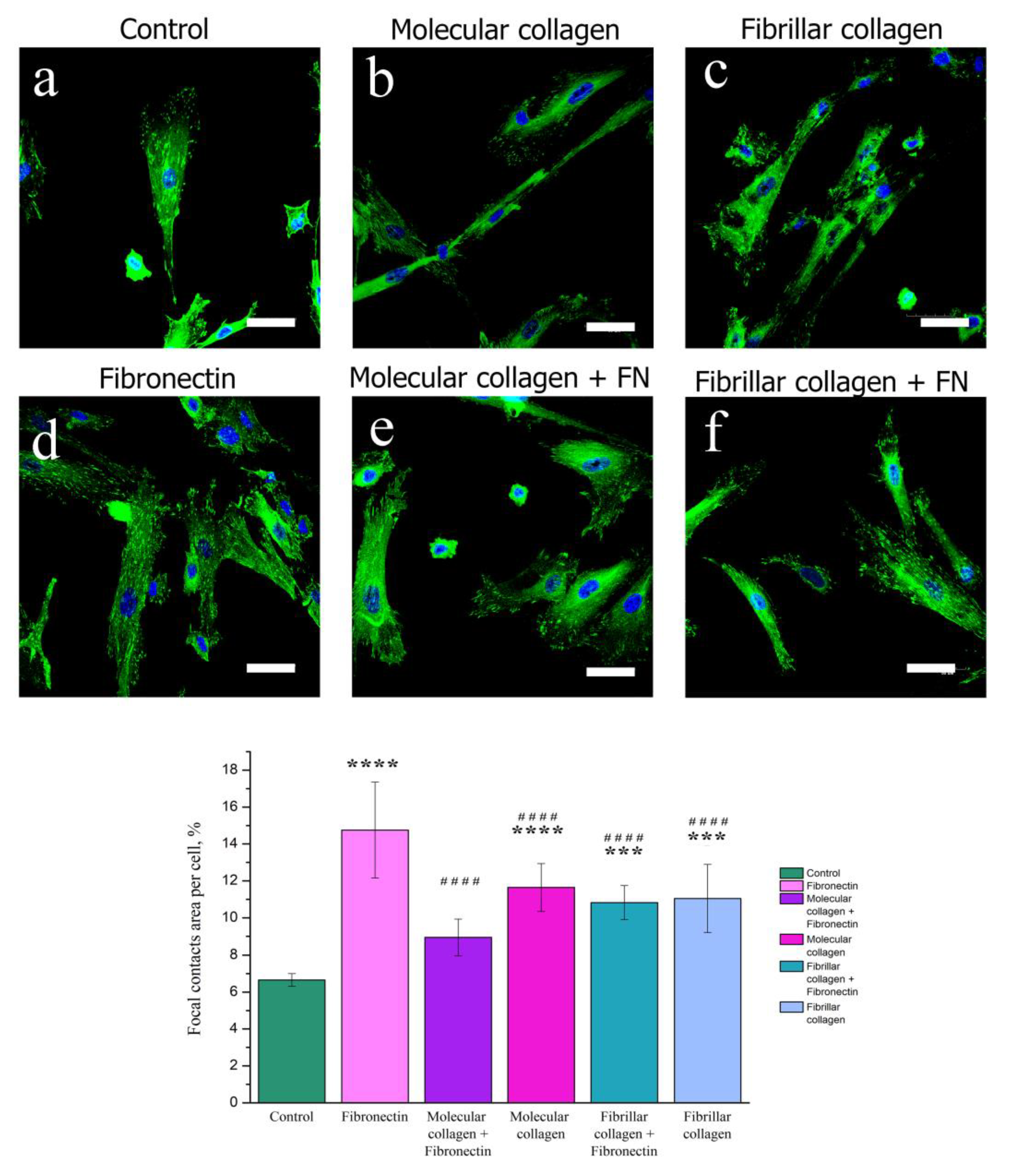
2.4.2. Focal Adhesion Contact Analysis by Confocal Fluorescence Microscopy
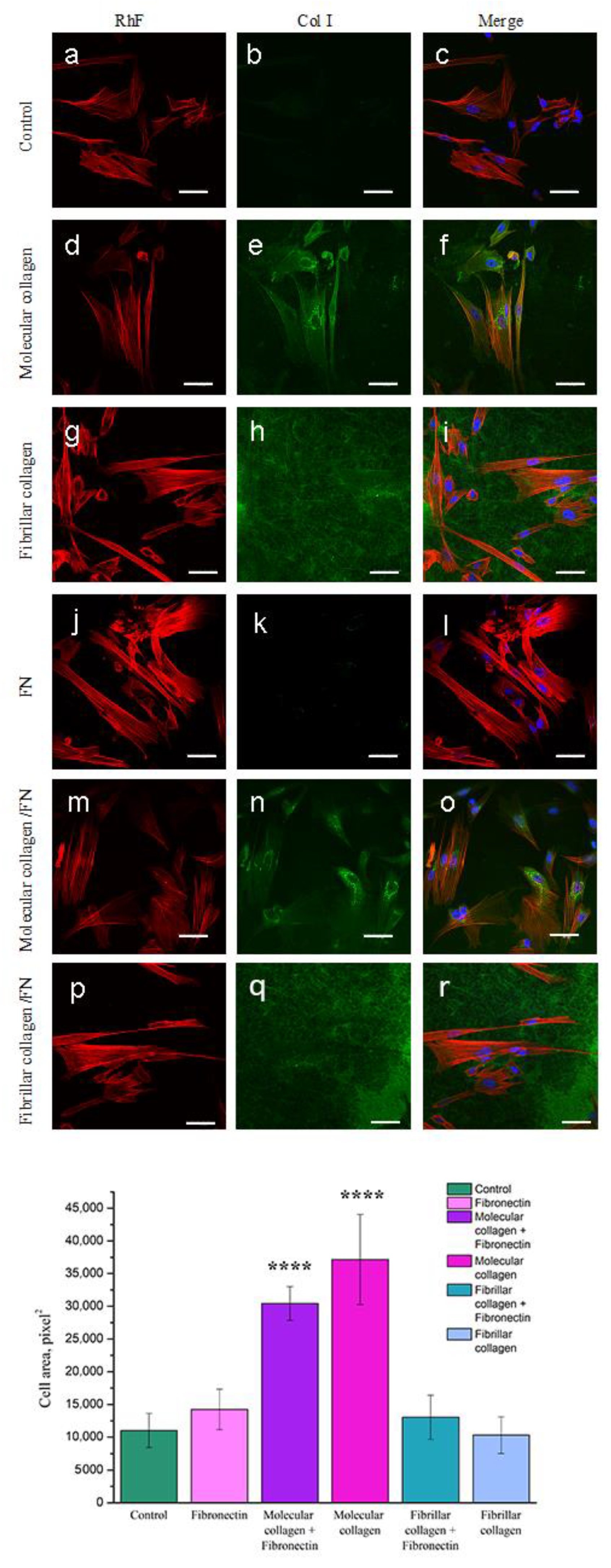
2.4.3. Spreading Analysis by Confocal Fluorescence Microscopy
3. Materials and Methods
3.1. Collagen Surface Formation
3.2. Atomic Force Microscopy
3.3. Transmission Electron Microscopy
3.4. Fourier Transform Infrared
3.5. Cell Cultivation
3.6. Scanning Electron Microscopy
3.7. Fluorescence Staining of MSCs
3.8. Cell and Focal Contacts Counts
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Assunção, M.; Dehghan-Baniani, D.; Yiu, C.H.K.; Später, T.; Beyer, S.; Blocki, A. Cell-Derived Extracellular Matrix for Tissue Engineering and Regenerative Medicine. Front. Bioeng. Biotechnol. 2020, 8, 602009. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Orgel, J.P.R.O.; San Antonio, J.D.; Antipova, O. Molecular and structural mapping of collagen fibril interactions. Connect. Tissue Res. 2011, 52, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Cowin, S.C. How is a tissue built? J. Biomech. Eng. 2000, 122, 553–569. [Google Scholar] [CrossRef] [PubMed]
- Kadler, K.E.; Hill, A.; Canty-Laird, E.G. Collagen fibrillogenesis: Fibronectin, integrins, and minor collagens as organizers and nucleators. Curr. Opin. Cell Biol. 2008, 20, 495–501. [Google Scholar] [CrossRef]
- Rieu, C.; Picaut, L.; Mosser, G.; Trichet, L. From Tendon Injury to Collagen-based Tendon Regeneration: Overview and Recent Advances. Curr. Pharm. Des. 2017, 23, 3483–3506. [Google Scholar] [CrossRef]
- Kadler, K.E.; Holmes, D.F.; Trotter, J.A.; Chapman, J.A. Review Artice: Collagen fibril formation Karl. Biochem. J. 1996, 316, 1–11. [Google Scholar] [CrossRef]
- Kadler, K.E.; Hojima, Y.; Prockop, D.J. Assembly of collagen fibrils de novo by cleavage of the type I pC-collagen with procollagen C-proteinase. Assay of critical concentration demonstrates that collagen self-assembly is a classical example of an entropy-driven process. J. Biol. Chem. 1987, 262, 15696–15701. [Google Scholar] [CrossRef]
- Di Lullo, G.A.; Sweeney, S.M.; Körkkö, J.; Ala-Kokko, L.; San Antonio, J.D. Mapping the ligand-binding sites and disease-associated mutations on the most abundant protein in the human, type I collagen. J. Biol. Chem. 2002, 277, 4223–4231. [Google Scholar] [CrossRef]
- Somaiah, C.; Kumar, A.; Mawrie, D.; Sharma, A.; Patil, S.D.; Bhattacharyya, J.; Swaminathan, R.; Jaganathan, B.G. Collagen promotes higher adhesion, survival and proliferation of mesenchymal stem cells. PLoS ONE 2015, 10, e0145068. [Google Scholar] [CrossRef]
- Nashchekina, Y.A.; Yudintceva, N.M.; Nikonov, P.O.; Ivanova, E.A.; Smagina, L.V.; Voronkina, I.V. Effect of Concentration of Collagen Gel on Functional Activity of Bone Marrow Mesenchymal Stromal Cells. Bull. Exp. Biol. Med. 2017, 163, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Kuboki, Y. Osteoblast-related gene expression of bone marrow cells during the osteoblastic differentiation induced by type I collagen. J. Biochem. 2001, 129, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Nashchekina, Y.A.; Alexandrova, S.A.; Nikonov, P.; Ivankova, E.I.; Yudin, V.E.; Blinova, M.I.; Mikhailova, N.A. Study of the Osteoindictive Properties of Protein-Modified Polylactide Scaffolds. Bull. Exp. Biol. Med. 2019, 167, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Parisi, L.; Toffoli, A.; Ghezzi, B.; Mozzoni, B.; Lumetti, S.; Macaluso, G.M. A glance on the role of fibronectin in controlling cell response at biomaterial interface. Jpn. Dent. Sci. Rev. 2020, 56, 50–55. [Google Scholar] [CrossRef]
- Baneyx, G.; Baugh, L.; Vogel, V. Fibronectin extension and unfolding within cell matrix fibrils controlled by cytoskeletal tension. Proc. Natl. Acad. Sci. USA 2002, 99, 5139–5143. [Google Scholar] [CrossRef]
- Schwarzbauer, J.E.; DeSimone, D.W. Fibronectins, their fibrillogenesis, and in vivo functions. Cold Spring Harb. Perspect. Biol. 2011, 3, a005041. [Google Scholar] [CrossRef]
- George, E.L.; Baldwin, H.S.; Hynes, R.O. Fibronectins are essential for heart and blood vessel morphogenesis but are dispensable for initial specification of precursor cells. Blood 1997, 90, 3073–3081. [Google Scholar] [CrossRef]
- Sottile, J.; Hocking, D.C. Fibronectin Polymerization Regulates the Composition and Stability of Extracellular Matrix Fibrils and Cell-Matrix Adhesions. Mol. Biol. Cell 2002, 13, 3546–3559. [Google Scholar] [CrossRef]
- García, A.J.; Vega, M.D.; Boettiger, D. Modulation of cell proliferation and differentiation through substrate- dependent changes in fibronectin conformation. Mol. Biol. Cell 1999, 10, 785–798. [Google Scholar] [CrossRef]
- Erat, M.C.; Schwarz-Linek, U.; Pickford, A.R.; Farndale, R.W.; Campbell, I.D.; Vakonakis, I. Implications for collagen binding from the crystallographic structure of fibronectin 6FnI1-2FnII7FnI. J. Biol. Chem. 2010, 285, 33764–33770. [Google Scholar] [CrossRef]
- Erat, M.C.; Sladek, B.; Campbell, I.D.; Vakonakis, I. Structural analysis of collagen type I interactions with human fibronectin reveals a cooperative binding mode. J. Biol. Chem. 2013, 288, 17441–17450. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, C.A.; Dalecki, D.; Hocking, D.C. Regional Fibronectin and Collagen Fibril Co-Assembly Directs Cell Proliferation and Microtissue Morphology. PLoS ONE 2013, 8, e77316. [Google Scholar] [CrossRef] [PubMed]
- Vaheri, A.; Kurkinen, M.; Lehto, V.P.; Linder, E.; Timpl, R. Codistribution of pericellular matrix proteins in cultured fibroblasts and loss in transformation: Fibronectin and procollagen. Proc. Natl. Acad. Sci. USA 1978, 75, 4944–4948. [Google Scholar] [CrossRef] [PubMed]
- Shved, Y.A.; Kukhareva, L.B.; Zorin, I.M.; Blinova, M.I.; Bilibin, A.Y.; Pinaev, G.P. Interaction of cultured skin cells with the polylactide matrix coved with different collagen structural isoforms. Cell Tissue Biol. 2007, 1, 89–95. [Google Scholar] [CrossRef]
- Gold, L.I.; Pearlstein, E. Fibronectin-collagen binding and requirement during cellular adhesion. Biochem. J. 1980, 186, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Pearlstein, E. Plasma membrane glycoprotein which mediates adhesion of fibroblasts to collagen. Nature 1976, 262, 497–500. [Google Scholar] [CrossRef]
- Mercier, I.; Lechaire, J.P.; Desmouliere, A.; Gaill, F.; Aumailley, M. Interactions of human skin fibroblasts with monomeric or fibrillar collagens induce different organization of the cytoskeleton. Exp. Cell Res. 1996, 225, 245–256. [Google Scholar] [CrossRef]
- Nashchekina, Y.; Nikonov, P.; Mikhailova, N.; Nashchekin, A. Collagen scaffolds treated by hydrogen peroxide for cell cultivation. Polymers 2021, 13, 4134. [Google Scholar] [CrossRef]
- Velling, T.; Risteli, J.; Wennerberg, K.; Mosher, D.F.; Johansson, S. Polymerization of type I and III collagens is dependent on fibronectin and enhanced by integrins α11β1 and α2β1. J. Biol. Chem. 2002, 277, 37377–37381. [Google Scholar] [CrossRef]
- Li, S.; Van Den Diepstraten, C.; D’Souza, S.J.; Chan, B.M.C.; Pickering, J.G. Vascular smooth muscle cells orchestrate the assembly of type I collagen via α2β1 integrin, RhoA, and fibronectin polymerization. Am. J. Pathol. 2003, 163, 1045–1056. [Google Scholar] [CrossRef]
- Paten, J.A.; Martin, C.L.; Wanis, J.T.; Siadat, S.M.; Figueroa-Navedo, A.M.; Ruberti, J.W.; Deravi, L.F. Molecular Interactions between Collagen and Fibronectin: A Reciprocal Relationship that Regulates De Novo Fibrillogenesis. Chem 2019, 5, 2126–2145. [Google Scholar] [CrossRef]
- Williams, E.C.; Janmey, P.A.; Ferry, J.D.; Mosher, D.F. Conformational states of fibronectin. Effects of pH, ionic strength, and collagen binding. J. Biol. Chem. 1982, 257, 14973–14978. [Google Scholar] [CrossRef]
- Ingham, K.C.; Brew, S.A.; Isaacs, B.S. Interaction of fibronectin and its gelatin-binding domains with fluorescent-labeled chains of type 1 collagen. J. Biol. Chem. 1988, 263, 4624–4628. [Google Scholar] [CrossRef]
- Speranza, M.L.; Valentini, G.; Calligaro, A. Influence of Fibronectin on the Fibrillogenesis of Type I and Type III Collagen. Top. Catal. 1987, 7, 115–123. [Google Scholar] [CrossRef]
- Jakobsen, R.J.; Brown, L.L.; Hutson, T.B.; Fink, D.J.; Veis, A. Intermolecular interactions in collagen self-assembly as revealed by Fourier transform infrared spectroscopy. Science 1983, 220, 1288–1290. [Google Scholar] [CrossRef] [PubMed]
- Nashchekina, Y.A.; Starostina, A.A.; Trusova, N.A.; Sirotkina, M.Y.; Lihachev, A.I.; Nashchekin, A.V. Molecular and fibrillar structure collagen analysis by FTIR spectroscopy. J. Phys. Conf. Ser. 2020, 1697, 012053. [Google Scholar] [CrossRef]
- Muyonga, J.H.; Cole, C.G.B.; Duodu, K.G. Fourier transform infrared (FTIR) spectroscopic study of acid soluble collagen and gelatin from skins and bones of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 86, 325–332. [Google Scholar] [CrossRef]
- Musiime, M.; Chang, J.; Hansen, U.; Kadler, K.E.; Zeltz, C.; Gullberg, D. Collagen assembly at the cell surface: Dogmas revisited. Cells 2021, 10, 662. [Google Scholar] [CrossRef]
- Graham, J.; Raghunath, M.; Vogel, V. Fibrillar fibronectin plays a key role as nucleator of collagen i polymerization during macromolecular crowding-enhanced matrix assembly. Biomater. Sci. 2019, 7, 4519–4535. [Google Scholar] [CrossRef]
- Kubow, K.E.; Vukmirovic, R.; Zhe, L.; Klotzsch, E.; Smith, M.L.; Gourdon, D.; Luna, S.; Vogel, V. Mechanical forces regulate the interactions of fibronectin and collagen i in extracellular matrix. Nat. Commun. 2015, 6, 8026. [Google Scholar] [CrossRef] [PubMed]
- Tamkun, J.W.; DeSimone, D.W.; Fonda, D.; Patel, R.S.; Buck, C.; Horwitz, A.F.; Hynes, R.O. Structure of integrin, a glycoprotein involved in the transmembrane linkage between fibronectin and actin. Cell 1986, 46, 271–282. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Hynes, R.O. The emergence of integrins: A personal and historical perspective. Matrix Biol. 2004, 23, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Zeltz, C.; Orgel, J.; Gullberg, D. Molecular composition and function of integrin-based collagen glues-Introducing Colinbris. Biochim. Biophys. Acta-Gen. Subj. 2014, 1840, 2533–2548. [Google Scholar] [CrossRef] [PubMed]
- Bourgot, I.; Primac, I.; Louis, T.; Noël, A.; Maquoi, E. Reciprocal Interplay Between Fibrillar Collagens and Collagen-Binding Integrins: Implications in Cancer Progression and Metastasis. Front. Oncol. 2020, 10, 1488. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Hartig, R.; Delahay, R.M.; Rohde, M.; Brandt, S.; Conradi, J.; Takahashi, S.; Smolka, A.J.; Sewald, N.; Backert, S. A small fibronectin-mimicking protein from bacteria induces cell spreading and focal adhesion formation. J. Biol. Chem. 2010, 285, 23515–23526. [Google Scholar] [CrossRef]
- Leiss, M.; Beckmann, K.; Girós, A.; Costell, M.; Fässler, R. The role of integrin binding sites in fibronectin matrix assembly in vivo. Curr. Opin. Cell Biol. 2008, 20, 502–507. [Google Scholar] [CrossRef]
- Nagata, K.; Humphries, M.J.; Olden, K.; Yamada, K.M. Collagen can modulate cell interactions with fibronectin. J. Cell Biol. 1985, 101, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Dumbauld, D.W.; Lee, T.T.; Singh, A.; Scrimgeour, J.; Gersbach, C.A.; Zamir, E.A.; Fu, J.; Chen, C.S.; Curtis, J.E.; Craig, S.W.; et al. How vinculin regulates force transmission. Proc. Natl. Acad. Sci. USA 2013, 110, 9788–9793. [Google Scholar] [CrossRef]
- Nardone, G.; Oliver-De La Cruz, J.; Vrbsky, J.; Martini, C.; Pribyl, J.; Skládal, P.; Pešl, M.; Caluori, G.; Pagliari, S.; Martino, F.; et al. YAP regulates cell mechanics by controlling focal adhesion assembly. Nat. Commun. 2017, 8, 15321. [Google Scholar] [CrossRef] [PubMed]
- Horzum, U.; Ozdil, B.; Pesen-Okvur, D. Step-by-step quantitative analysis of focal adhesions. MethodsX 2014, 1, 56–59. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nashchekina, Y.; Nikonov, P.; Prasolov, N.; Sulatsky, M.; Chabina, A.; Nashchekin, A. The Structural Interactions of Molecular and Fibrillar Collagen Type I with Fibronectin and Its Role in the Regulation of Mesenchymal Stem Cell Morphology and Functional Activity. Int. J. Mol. Sci. 2022, 23, 12577. https://doi.org/10.3390/ijms232012577
Nashchekina Y, Nikonov P, Prasolov N, Sulatsky M, Chabina A, Nashchekin A. The Structural Interactions of Molecular and Fibrillar Collagen Type I with Fibronectin and Its Role in the Regulation of Mesenchymal Stem Cell Morphology and Functional Activity. International Journal of Molecular Sciences. 2022; 23(20):12577. https://doi.org/10.3390/ijms232012577
Chicago/Turabian StyleNashchekina, Yuliya, Pavel Nikonov, Nikita Prasolov, Maksim Sulatsky, Alina Chabina, and Alexey Nashchekin. 2022. "The Structural Interactions of Molecular and Fibrillar Collagen Type I with Fibronectin and Its Role in the Regulation of Mesenchymal Stem Cell Morphology and Functional Activity" International Journal of Molecular Sciences 23, no. 20: 12577. https://doi.org/10.3390/ijms232012577
APA StyleNashchekina, Y., Nikonov, P., Prasolov, N., Sulatsky, M., Chabina, A., & Nashchekin, A. (2022). The Structural Interactions of Molecular and Fibrillar Collagen Type I with Fibronectin and Its Role in the Regulation of Mesenchymal Stem Cell Morphology and Functional Activity. International Journal of Molecular Sciences, 23(20), 12577. https://doi.org/10.3390/ijms232012577