



Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases

Abstract
1. Introduction
2. Function and Structure of VTN and Its Receptor
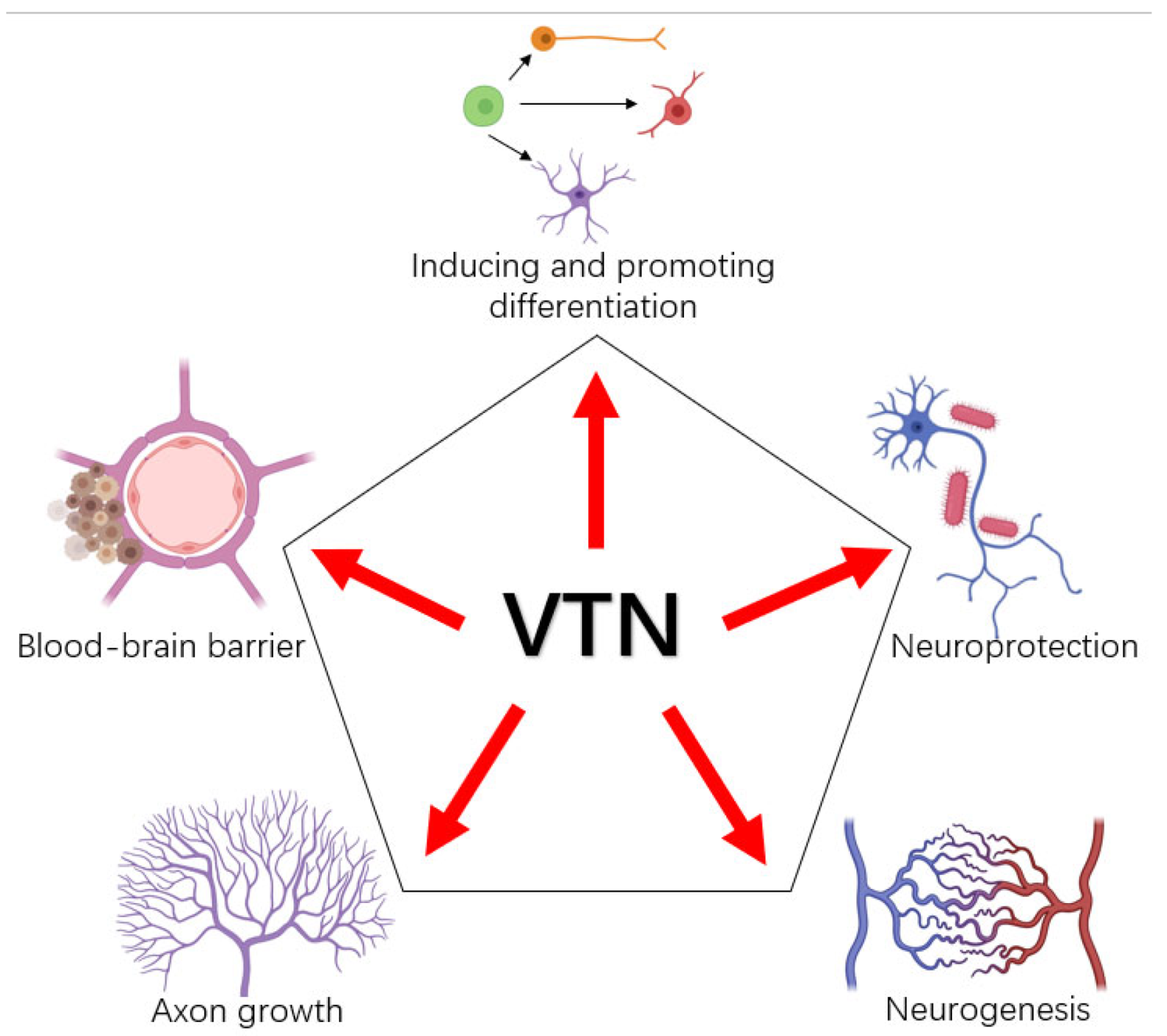
3. Functions of VTN in Neurons and Glial Cells
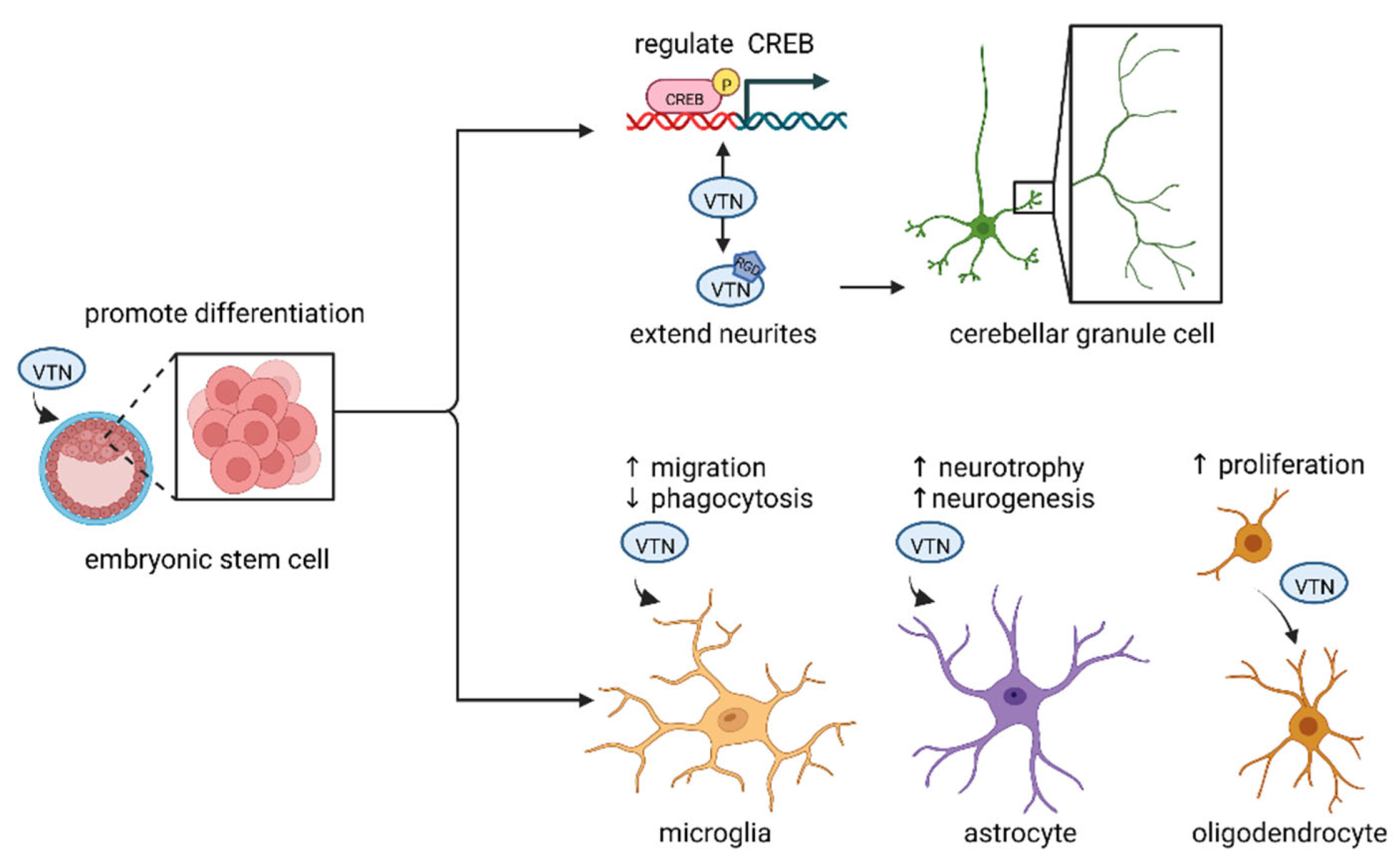
3.1. VTN and Neural Differentiation
3.1.1. VTN and Embryonic Stem Cells
3.1.2. VTN and Cerebellar Granule Cells
3.2. VTN and Axons
3.3. VTN and Glial Cells
3.3.1. VTN and Microglia
3.3.2. VTN and Astrocytes
3.3.3. VTN and Oligodendrocytes
4. VTN in Neurodegenerative Diseases
4.1. VTN and Age-Related Macular Degeneration
4.2. VTN and Alzheimer’s Disease
4.3. VTN and Multiple Sclerosis
4.4. VTN and Stroke

{kind=link}
{kind=link}
{kind=link}
| Diseases | Roles of VTN in Neurodegenerative Diseases | References |
|---|---|---|
| Age-related macular degeneration | a major component of drusen; involved in the pathogenesis of AMD | [65,66] |
| Alzheimer’s disease | exists in plaques and neurofibrillary tangles; inhibits the aggregation of β-peptide; regulates β-amyloid precursor protein; misfolding of VTN tends to amyloidosis | [18,70,72,73] |
| Multiple sclerosis | increased expression in active MS; regulates oligodendrocyte proliferation, survival and development | [75,76,77,78] |
| Stroke | inhibits post-stroke neurogenesis; aggravates neurological dysfunction | [80,81] |
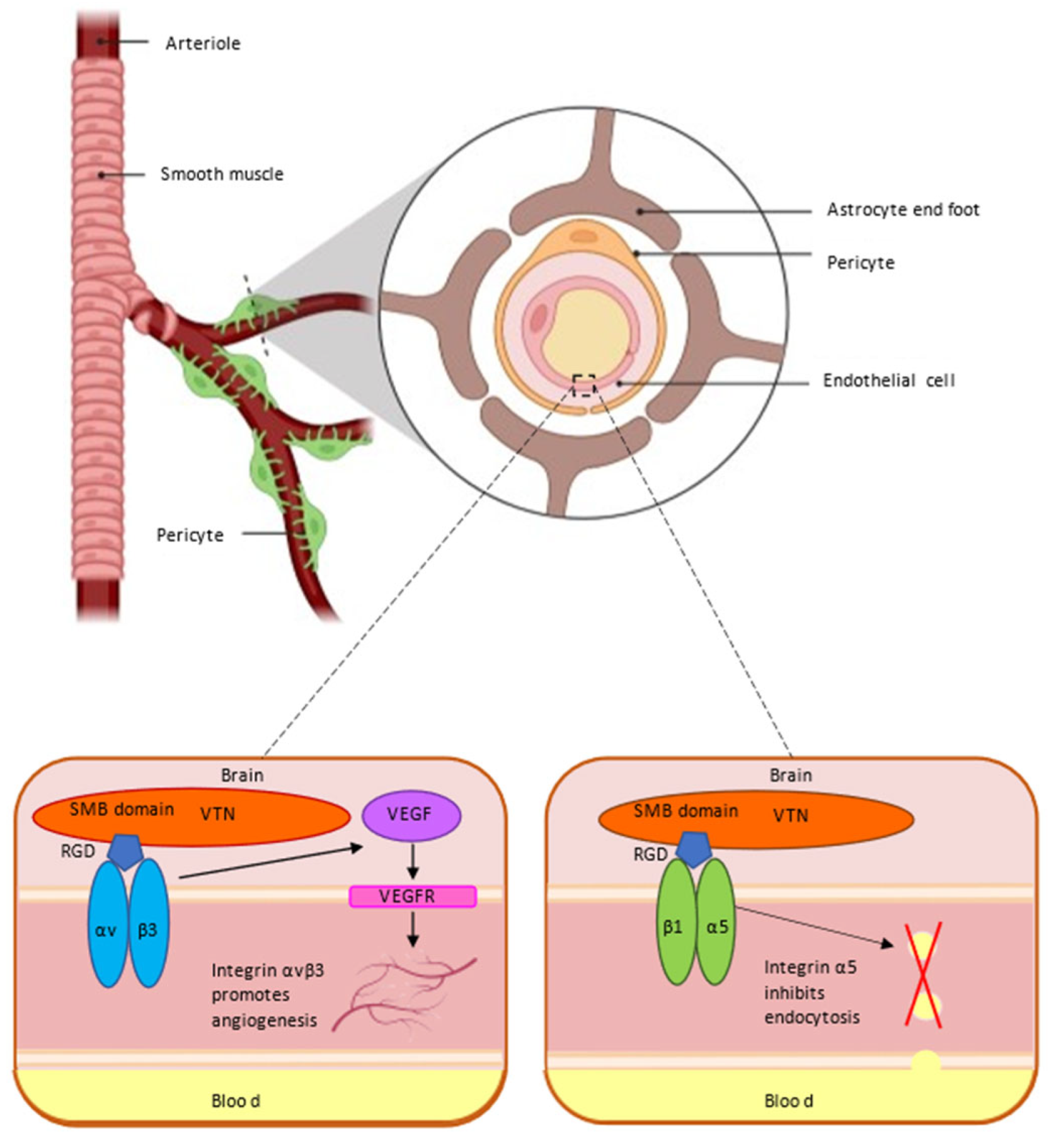
5. VTN and Blood–Brain Barrier
5.1. VTN Receptor αvβ3 Integrin and Blood–Brain Barrier
5.2. VTN Receptor α5 Integrin and Blood–Brain Barrier
6. Current Targeted Drugs for VTN and Its Receptors
6.1. Integrin Receptor Antagonists
6.2. Platelet Receptor Antagonists
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Valdoz, J.C.; Johnson, B.C.; Jacobs, D.J.; Franks, N.A.; Dodson, E.L.; Sanders, C.; Cribbs, C.G.; Van Ry, P.M. The ECM: To scaffold, or not to scaffold, that is the question. Int. J. Mol. Sci. 2021, 22, 12690. [Google Scholar] [CrossRef] [PubMed]
- Conlan, M.G.; Tomasini, B.R.; Schultz, R.L.; Mosher, D.F. Plasma vitronectin polymorphism in normal subjects and patients with disseminated intravascular coagulation. Blood 1988, 72, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Leavesley, D.I.; Kashyap, A.S.; Croll, T.; Sivaramakrishnan, M.; Shokoohmand, A.; Hollier, B.G.; Upton, Z. Vitronectin—Master controller or micromanager? IUBMB Life 2013, 65, 807–818. [Google Scholar] [CrossRef]
- Su, Y.-C.; Riesbeck, K. Vitronectin. In The Complement FactsBook; Elsevier: Amsterdam, The Netherlands, 2018; pp. 351–360. [Google Scholar]
- Preissner, K.T.; Seiffert, D. Role of vitronectin and its receptors in haemostasis and vascular remodeling. Thromb. Res. 1998, 89, 1–21. [Google Scholar] [CrossRef]
- Shen, J.; Zhu, Y.; Zhang, S.; Lyu, S.; Lyu, C.; Feng, Z.; Hoyle, D.L.; Wang, Z.Z.; Cheng, T. Vitronectin-activated αvβ3 and αvβ5 integrin signalling specifies haematopoietic fate in human pluripotent stem cells. Cell Prolif. 2021, 54, e13012. [Google Scholar] [CrossRef]
- Pitkänen, A.; Ndode-Ekane, X.E.; Łukasiuk, K.; Wilczynski, G.M.; Dityatev, A.; Walker, M.C.; Chabrol, E.; Dedeurwaerdere, S.; Vazquez, N.; Powell, E.M. Neural ECM and epilepsy. Prog. Brain Res. 2014, 214, 229–262. [Google Scholar] [CrossRef]
- Liakka, K.A.; Autio-Harmainen, H. Distribution of the extracellular matrix proteins tenascin, fibronectin, and vitronectin in fetal, infant, and adult human spleens. J. Histochem. Cytochem. 1992, 40, 1203–1210. [Google Scholar] [CrossRef]
- Sumida, H.; Nakamura, H.; Satow, Y. Distribution of vitronectin in the embryonic chick heart during endocardial cell migration. Arch. Histol. Cytol. 1990, 53, 81–88. [Google Scholar] [CrossRef]
- Holmes, R. Preparation from human serum of an alpha-one protein which induces the immediate growth of unadapted cells in vitro. J. Cell Biol. 1967, 32, 297–308. [Google Scholar] [CrossRef]
- Preissner, K.T.; Jenne, D. Vitronectin: A new molecular connection in haemostasis. Thromb. Haemost. 1991, 66, 189–194. [Google Scholar] [CrossRef]
- Tollefsen, D.M.; Weigel, C.J.; Kabeer, M.H. The presence of methionine or threonine at position 381 in vitronectin is correlated with proteolytic cleavage at arginine 379. J. Biol. Chem. 1990, 265, 9778–9781. [Google Scholar] [CrossRef]
- Jeon, B.-M.; Yeon, G.-B.; Goo, H.-G.; Lee, K.E.; Kim, D.-S. PVDF nanofiber scaffold coated with a vitronectin peptide facilitates the neural differentiation of human embryonic stem cells. Dev. Reprod. 2020, 24, 135. [Google Scholar] [CrossRef]
- Park, W.U.; Yeon, G.-B.; Yu, M.-S.; Goo, H.-G.; Hwang, S.-H.; Na, D.; Kim, D.-S. A novel vitronectin peptide facilitates differentiation of oligodendrocytes from human pluripotent stem cells (Synthetic ecm for oligodendrocyte differentiation). Biology 2021, 10, 1254. [Google Scholar] [CrossRef]
- Abe, A.; Hashimoto, K.; Akiyama, A.; Iida, M.; Ikeda, N.; Hamano, A.; Watanabe, R.; Hayashi, Y.; Miyamoto, Y. αvβ5 integrin mediates the effect of vitronectin on the initial stage of differentiation in mouse cerebellar granule cell precursors. Brain Res. 2018, 1691, 94–104. [Google Scholar] [CrossRef]
- Sugahara, M.; Nakaoki, Y.; Yamaguchi, A.; Hashimoto, K.; Miyamoto, Y. Vitronectin is involved in the morphological transition of neurites in retinoic acid-induced neurogenesis of neuroblastoma cell line neuro2a. Neurochem. Res. 2019, 44, 1621–1635. [Google Scholar] [CrossRef]
- Bronfman, F.C.; Soto, C.; Tapia, L.; Tapia, V.; Inestrosa, N.C. Extracellular matrix regulates the amount of the β-amyloid precursor protein and its amyloidogenic fragments. J. Cell. Physiol. 1996, 166, 360–369. [Google Scholar] [CrossRef]
- Hall-Roberts, H.; Agarwal, D.; Obst, J.; Smith, T.B.; Monzón-Sandoval, J.; Di Daniel, E.; Webber, C.; James, W.S.; Mead, E.; Davis, J.B. TREM2 Alzheimer’s variant R47H causes similar transcriptional dysregulation to knockout, yet only subtle functional phenotypes in human iPSC-derived macrophages. Alzheimer’s Res. Ther. 2020, 12, 1–27. [Google Scholar] [CrossRef]
- Chillakuri, C.R.; Jones, C.; Mardon, H.J. Heparin binding domain in vitronectin is required for oligomerization and thus enhances integrin mediated cell adhesion and spreading. FEBS Lett. 2010, 584, 3287–3291. [Google Scholar] [CrossRef]
- Vasilyev, D.V.; Barish, M.E. Regulation of an inactivating potassium current (IA) by the extracellular matrix protein vitronectin in embryonic mouse hippocampal neurones. J. Physiol. 2003, 547, 859–871. [Google Scholar] [CrossRef]
- Harburger, D.S.; Calderwood, D.A. Integrin signalling at a glance. J. Cell Sci. 2009, 122, 159–163. [Google Scholar] [CrossRef]
- Preissner, T.K. Structure and Biological Role of Vitronectin. Annu. Rev. Cell Biol. 1991, 7, 275–310. [Google Scholar] [CrossRef]
- Takagi, J.; Petre, B.M.; Walz, T.; Springer, T.A. Global conformational rearrangements in integrin extracellular domains in outside-in and inside-out signaling. Cell 2002, 110, 599–611. [Google Scholar] [CrossRef]
- Zoppi, N.; Chiarelli, N.; Ritelli, M.; Colombi, M. Multifaced roles of the αvβ3 integrin in Ehlers–Danlos and arterial tortuosity syndromes’ dermal fibroblasts. Int. J. Mol. Sci. 2018, 19, 982. [Google Scholar] [CrossRef]
- Uehara, K.; Uehara, A. Integrin alphavbeta5 in endothelial cells of rat splenic sinus: An immunohistochemical and ultrastructural study. Cell Tissue Res. 2014, 356, 183–193. [Google Scholar] [CrossRef]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Mawaribuchi, S.; Aiki, Y.; Ikeda, N.; Ito, Y. mRNA and miRNA expression profiles in an ectoderm-biased substate of human pluripotent stem cells. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Seiffert, D.; Iruela-Arispe, M.L.; Sage, E.H.; Loskutoff, D.J. Distribution of vitronectin mRNA during murine development. Dev. Dyn. 1995, 203, 71–79. [Google Scholar] [CrossRef]
- Martínez-Morales, J.R.; Barbas, J.A.; Martí, E.; Bovolenta, P.; Edgar, D.; Rodríguez-Tébar, A. Vitronectin is expressed in the ventral region of the neural tube and promotes the differentiation of motor neurons. Development 1997, 124, 5139–5147. [Google Scholar] [CrossRef]
- Ikeda, Y. Expression of the two estrogen receptor (ER) subtypes, ERα and ERβ, during postnatal development of the rat cerebellum. In Proceedings of Annual Meeting of the Physiological Society of Japan; Physiological Society Of Japan: Tokyo, Japan, 2007; p. 044. [Google Scholar]
- Hashimoto, K.; Sakane, F.; Ikeda, N.; Akiyama, A.; Sugahara, M.; Miyamoto, Y. Vitronectin promotes the progress of the initial differentiation stage in cerebellar granule cells. Mol. Cell. Neurosci. 2016, 70, 76–85. [Google Scholar] [CrossRef]
- Wechsler-Reya, R.J. Caught in the matrix: How vitronectin controls neuronal differentiation. Trends Neurosci. 2001, 24, 680–682. [Google Scholar] [CrossRef]
- Pons, S.; Trejo, J.L.; Martínez-Morales, J.R.; Martí, E. Vitronectin regulates Sonic hedgehog activity during cerebellum development through CREB phosphorylation. Development 2001, 128, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Oishi, Y.; Hashimoto, K.; Abe, A.; Kuroda, M.; Fujii, A.; Miyamoto, Y. Vitronectin regulates the axon specification of mouse cerebellar granule cell precursors via αvβ5 integrin in the differentiation stage. Neurosci. Lett. 2021, 746, 135648. [Google Scholar] [CrossRef] [PubMed]
- Murase, S.I.; Hayashi, Y. Concomitant expression of genes encoding integrin αvβ5 heterodimer and vitronectin in growing parallel fibers of postnatal rat cerebellum: A possible role as mediators of parallel fiber elongation. J. Comp. Neurol. 1998, 397, 199–212. [Google Scholar] [CrossRef]
- Walker, D.; McGeer, P. Vitronectin expression in Purkinje cells in the human cerebellum. Neurosci. Lett. 1998, 251, 109–112. [Google Scholar] [CrossRef]
- Bouslama-Oueghlani, L.; Wehrlé, R.; Doulazmi, M.; Chen, X.R.; Jaudon, F.; Lemaigre-Dubreuil, Y.; Rivals, I.; Sotelo, C.; Dusart, I. Purkinje cell maturation participates in the control of oligodendrocyte differentiation: Role of sonic hedgehog and vitronectin. PLoS ONE 2012, 7, e49015. [Google Scholar] [CrossRef]
- Felding-Habermann, B.; Cheresh, D.A. Vitronectin and its receptors. Curr. Opin. Cell Biol. 1993, 5, 864–868. [Google Scholar] [CrossRef]
- Hajj, G.N.; Santos, T.G.; Cook, Z.S.; Martins, V.R. Developmental expression of prion protein and its ligands stress-inducible protein 1 and vitronectin. J. Comp. Neurol. 2009, 517, 371–384. [Google Scholar] [CrossRef]
- Hajj, G.N.; Lopes, M.H.; Mercadante, A.F.; Veiga, S.S.; da Silveira, R.B.; Santos, T.G.; Ribeiro, K.C.; Juliano, M.A.; Jacchieri, S.G.; Zanata, S.M. Cellular prion protein interaction with vitronectin supports axonal growth and is compensated by integrins. J. Cell Sci. 2007, 120, 1915–1926. [Google Scholar] [CrossRef]
- Hansson, M.L.; Chatterjee, U.; Francis, J.; Arndt, T.; Broman, C.; Johansson, J.; Sköld, M.K.; Rising, A. Artificial spider silk supports and guides neurite extension in vitro. FASEB J. 2021, 35, e21896. [Google Scholar] [CrossRef]
- Wynn, T.A.; Barron, L.; Thompson, R.W.; Madala, S.K.; Wilson, M.S.; Cheever, A.W.; Ramalingam, T. Quantitative assessment of macrophage functions in repair and fibrosis. Curr. Protoc. Immunol. 2011, 93, 14–22. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, H. Microglia and their roles in response to injury of the central nervous system. Yixue Yanjiusheng Xuebao 2012, 25, 418–421. [Google Scholar] [CrossRef]
- Neher, J.J.; Neniskyte, U.; Zhao, J.-W.; Bal-Price, A.; Tolkovsky, A.M.; Brown, G.C. Inhibition of microglial phagocytosis is sufficient to prevent inflammatory neuronal death. J. Immunol. 2011, 186, 4973–4983. [Google Scholar] [CrossRef]
- Zhao, L.; Zabel, M.K.; Wang, X.; Ma, W.; Shah, P.; Fariss, R.N.; Qian, H.; Parkhurst, C.N.; Gan, W.B.; Wong, W.T. Microglial phagocytosis of living photoreceptors contributes to inherited retinal degeneration. EMBO Mol. Med. 2015, 7, 1179–1197. [Google Scholar] [CrossRef]
- Jeon, H.; Kim, J.-H.; Kim, J.-H.; Lee, W.-H.; Lee, M.-S.; Suk, K. Plasminogen activator inhibitor type 1 regulates microglial motility and phagocytic activity. J. Neuroinflamm. 2012, 9, 1–22. [Google Scholar] [CrossRef]
- Morel, L.; Chiang, M.S.R.; Higashimori, H.; Shoneye, T.; Iyer, L.K.; Yelick, J.; Tai, A.; Yang, Y. Molecular and functional properties of regional astrocytes in the adult brain. J. Neurosci. 2017, 37, 8706–8717. [Google Scholar] [CrossRef]
- Cohen-Salmon, M.; Slaoui, L.; Mazaré, N.; Gilbert, A.; Oudart, M.; Alvear-Perez, R.; Elorza-Vidal, X.; Chever, O.; Boulay, A.C. Astrocytes in the regulation of cerebrovascular functions. Glia 2021, 69, 817–841. [Google Scholar] [CrossRef]
- Augusto-Oliveira, M.; Arrifano, G.P.; Takeda, P.Y.; Lopes-Araújo, A.; Santos-Sacramento, L.; Anthony, D.C.; Verkhratsky, A.; Crespo-Lopez, M.E. Astroglia-specific contributions to the regulation of synapses, cognition and behaviour. Neurosci. Biobehav. Rev. 2020, 118, 331–357. [Google Scholar] [CrossRef]
- von Bernhardi, R.; Eugenín-von Bernhardi, J.; Flores, B.; Eugenín León, J. Glial cells and integrity of the nervous system. Glial Cells Health Dis. CNS 2016, 949, 1–24. [Google Scholar] [CrossRef]
- Johnson, K.M.; Milner, R.; Crocker, S.J. Extracellular matrix composition determines astrocyte responses to mechanical and inflammatory stimuli. Neurosci. Lett. 2015, 600, 104–109. [Google Scholar] [CrossRef]
- Jia, C.; Keasey, M.P.; Malone, H.M.; Lovins, C.; Sante, R.R.; Razskazovskiy, V.; Hagg, T. Vitronectin from brain pericytes promotes adult forebrain neurogenesis by stimulating CNTF. Exp. Neurol. 2019, 312, 20–32. [Google Scholar] [CrossRef]
- Keasey, M.P.; Kang, S.S.; Lovins, C.; Hagg, T. Inhibition of a novel specific neuroglial integrin signaling pathway increases STAT3-mediated CNTF expression. Cell Commun. Signal. 2013, 11, 1–15. [Google Scholar] [CrossRef]
- Alizadeh, A.; Karimi-Abdolrezaee, S. Microenvironmental regulation of oligodendrocyte replacement and remyelination in spinal cord injury. J. Physiol. 2016, 594, 3539–3552. [Google Scholar] [CrossRef]
- Mathews, E.S.; Appel, B. Oligodendrocyte differentiation. Methods Cell Biol. 2016, 134, 69–96. [Google Scholar] [CrossRef]
- Soomro, S.H.; Jie, J.; Fu, H. Oligodendrocytes development and Wnt signaling pathway. Int. J. Hum. Anat 2018, 1, 17–35. [Google Scholar] [CrossRef]
- Baron, W.; Shattil, S.J. The oligodendrocyte precursor mitogen PDGF stimulates proliferation by activation of αvβ3 integrins. EMBO J. 2002, 21, 1957–1966. [Google Scholar] [CrossRef]
- Gil, J.-E.; Woo, D.-H.; Shim, J.-H.; Kim, S.-E.; You, H.-J.; Park, S.-H.; Paek, S.H.; Kim, S.-K.; Kim, J.-H. Vitronectin promotes oligodendrocyte differentiation during neurogenesis of human embryonic stem cells. FEBS Lett. 2009, 583, 561–567. [Google Scholar] [CrossRef]
- Shin, K.; Kent, J.E.; Singh, C.; Fujimoto, L.M.; Yu, J.; Tian, Y.; Im, W.; Marassi, F.M. Calcium and hydroxyapatite binding site of human vitronectin provides insights to abnormal deposit formation. Proc. Natl. Acad. Sci. USA 2020, 117, 18504–18510. [Google Scholar] [CrossRef]
- Fernandes, A.R.; Zielińska, A.; Sanchez-Lopez, E.; Dos Santos, T.; Garcia, M.L.; Silva, A.M.; Karczewski, J.; Souto, E.B. Exudative versus nonexudative age-related macular degeneration: Physiopathology and treatment options. Int. J. Mol. Sci. 2022, 23, 2592. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Reddy, S.T.; Hinton, D.R.; Kannan, R. Mechanisms of RPE senescence and potential role of αB crystallin peptide as a senolytic agent in experimental AMD. Exp. Eye Res. 2022, 215, 108918. [Google Scholar] [CrossRef]
- Waldstein, S.M.; Vogl, W.-D.; Bogunovic, H.; Sadeghipour, A.; Riedl, S.; Schmidt-Erfurth, U. Characterization of drusen and hyperreflective foci as biomarkers for disease progression in age-related macular degeneration using artificial intelligence in optical coherence tomography. JAMA Ophthalmol. 2020, 138, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Mullins, R.F.; Russell, S.R.; Anderson, D.H.; Hageman, G.S. Drusen associated with aging and age-related macular degeneration contain proteins common to extracellular deposits associated with atherosclerosis, elastosis, amyloidosis, and dense deposit disease. FASEB J. 2000, 14, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Hageman, G.S.; Mullins, R.F.; Russell, S.R.; Johnson, L.V.; Anderson, D.H. Vitronectin is a constituent of ocular drusen and the vitronectin gene is expressed in human retinal pigmented epithelial cells. FASEB J. 1999, 13, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Biasella, F.; Plössl, K.; Karl, C.; Weber, B.H.; Friedrich, U. Altered protein function caused by AMD-associated variant rs704 links vitronectin to disease pathology. Investig. Ophthalmol. Vis. Sci. 2020, 61, 2. [Google Scholar] [CrossRef]
- Biasella, F.; Strunz, T.; Kiel, C.; Weber, B.H.; Friedrich, U.; Consortium, I.A.G. Vitronectin and its interaction with PAI-1 suggests a functional link to vascular changes in AMD pathobiology. Cells 2022, 11, 1766. [Google Scholar] [CrossRef]
- Scheltens, P.; Strooper, B.D.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; Flier, W.M.V.D. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Masters, C.L.; Bateman, R.; Blennow, K.; Rowe, C.C.; Sperling, R.A.; Cummings, J.L. Alzheimer’s disease. Nat. Rev. Dis. Prim. 2015, 1, 15056. [Google Scholar] [CrossRef]
- Akiyama, H.; Kawamata, T.; Dedhar, S.; McGeer, P. Immunohistochemical localization of vitronectin, its receptor and beta-3 integrin in Alzheimer brain tissue. J. Neuroimmunol. 1991, 32, 19–28. [Google Scholar] [CrossRef]
- Eikelenboom, P.; Zhan, S.; Kamphorst, W.; Van der Valk, P.; Rozemuller, J. Cellular and substrate adhesion molecules (integrins) and their ligands in cerebral amyloid plaques in Alzheimer’s disease. Virchows Arch. 1994, 424, 421–427. [Google Scholar] [CrossRef]
- Geraghty, N.J.; Satapathy, S.; Kelly, M.; Cheng, F.; Lee, A.; Wilson, M.R. Expanding the family of extracellular chaperones: Identification of human plasma proteins with chaperone activity. Protein Sci. 2021, 30, 2272–2286. [Google Scholar] [CrossRef]
- Shin, T.M.; Isas, J.M.; Hsieh, C.-L.; Kayed, R.; Glabe, C.G.; Langen, R.; Chen, J. Formation of soluble amyloid oligomers and amyloid fibrils by the multifunctional protein vitronectin. Mol. Neurodegener. 2008, 3, 1–12. [Google Scholar] [CrossRef]
- Gironi, M.; Arnò, C.; Comi, G.; Penton-Rol, G.; Furlan, R. Multiple sclerosis and neurodegenerative diseases. In Immune Rebalancing; Elsevier: Amsterdam, The Netherlands, 2016; pp. 63–84. [Google Scholar]
- O’Meara, R.W.; Michalski, J.-P.; Kothary, R. Integrin signaling in oligodendrocytes and its importance in CNS myelination. J. Signal Transduct. 2011, 2011, 354091. [Google Scholar] [CrossRef]
- Psenicka, M.W.; Smith, B.C.; Tinkey, R.A.; Williams, J.L. Connecting Neuroinflammation and Neurodegeneration in Multiple Sclerosis: Are Oligodendrocyte Precursor Cells a Nexus of Disease? Front. Cell. Neurosci. 2021, 15, 221. [Google Scholar] [CrossRef]
- Ghorbani, S.; Yong, V.W. The extracellular matrix as modifier of neuroinflammation and remyelination in multiple sclerosis. Brain 2021, 144, 1958–1973. [Google Scholar] [CrossRef]
- Sobel, R.A.; Chen, M.; Maeda, A.; Hinojoza, J.R. Vitronectin and integrin vitronectin receptor localization in multiple sclerosis lesions. J. Neuropathol. Exp. Neurol. 1995, 54, 202–213. [Google Scholar] [CrossRef]
- Johnston, S.C.; Mendis, S.; Mathers, C.D. Global variation in stroke burden and mortality: Estimates from monitoring, surveillance, and modelling. Lancet Neurol. 2009, 8, 345–354. [Google Scholar] [CrossRef]
- Jia, C.; Malone, H.M.; Keasey, M.P.; Lovins, C.; Elam, J.; Hagg, T. Blood vitronectin induces detrimental brain interleukin-6 and correlates with outcomes after stroke only in female mice. Stroke 2020, 51, 1587–1595. [Google Scholar] [CrossRef]
- Jia, C.; Keasey, M.P.; Malone, H.M.; Lovins, C.; Hagg, T. Vitronectin mitigates stroke-increased neurogenesis only in female mice and through FAK-regulated IL-6. Exp. Neurol. 2020, 323, 113088. [Google Scholar] [CrossRef]
- Chojdak-Łukasiewicz, J.; Dziadkowiak, E.; Zimny, A.; Paradowski, B. Cerebral small vessel disease: A review. Adv. Clin. Exp. Med. 2021, 30, 349–356. [Google Scholar] [CrossRef]
- Rajani, R.M.; Williams, A. Endothelial cell–oligodendrocyte interactions in small vessel disease and aging. Clin. Sci. 2017, 131, 369–379. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood–brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Xiao, Z.J.; Yang, B.; Lan, Z.; Fang, F. Blood-brain barrier: More contributor to disruption of central nervous system homeostasis than victim in neurological disorders. Front. Neurosci. 2020, 14, 764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, L.; An, C.; Wang, R.; Yang, L.; Yu, W.; Li, P.; Gao, Y. The blood brain barrier in cerebral ischemic injury–Disruption and repair. Brain Hemorrhages 2020, 1, 34–53. [Google Scholar] [CrossRef]
- Langen, U.H.; Ayloo, S.; Gu, C. Development and cell biology of the blood-brain barrier. Annu. Rev. Cell Dev. Biol. 2019, 35, 591. [Google Scholar] [CrossRef]
- Brooks, P.C.; Clark, R.A.; Cheresh, D.A. Requirement of vascular integrin αvβ3 for angiogenesis. Science 1994, 264, 569–571. [Google Scholar] [CrossRef]
- Okada, Y.; Copeland, B.R.; Hamann, G.F.; Koziol, J.A.; Cheresh, D.A.; Del Zoppo, G. Integrin alphavbeta3 is expressed in selected microvessels after focal cerebral ischemia. Am. J. Pathol. 1996, 149, 37. [Google Scholar] [CrossRef]
- McCarty, J.H.; Lacy-Hulbert, A.; Charest, A.; Bronson, R.T.; Crowley, D.; Housman, D.; Savill, J.; Roes, J.; Hynes, R.O. Selective ablation of αv integrins in the central nervous system leads to cerebral hemorrhage, seizures, axonal degeneration and premature death. Development 2005, 132, 165–176. [Google Scholar] [CrossRef]
- Byzova, T.V.; Goldman, C.K.; Pampori, N.; Thomas, K.A.; Bett, A.; Shattil, S.J.; Plow, E.F. A mechanism for modulation of cellular responses to VEGF: Activation of the integrins. Mol. Cell 2000, 6, 851–860. [Google Scholar] [CrossRef]
- De Arcangelis, A.; Georges-Labouesse, E. Integrin and ECM functions: Roles in vertebrate development. Trends Genet. 2000, 16, 389–395. [Google Scholar] [CrossRef]
- Wu, C.-C.; Wang, L.-C.; Su, Y.-T.; Wei, W.-Y.; Tsai, K.-J. Synthetic α5β1 integrin ligand PHSRN is proangiogenic and neuroprotective in cerebral ischemic stroke. Biomaterials 2018, 185, 142–154. [Google Scholar] [CrossRef]
- Ayloo, S.; Lazo, C.G.; Sun, S.; Zhang, W.; Cui, B.; Gu, C. Pericyte-to-endothelial cell signaling via vitronectin-integrin regulates blood-CNS barrier. Neuron 2022, 110, 1641–1655.e6. [Google Scholar] [CrossRef]
- Askew, B.C.; Furuya, T.; Edwards, D.S. Ocular distribution and pharmacodynamics of SF0166, a topically administered αvβ3 integrin antagonist, for the treatment of retinal diseases. J. Pharmacol. Exp. Ther. 2018, 366, 244–250. [Google Scholar] [CrossRef]
- Su, H.; Gorodny, N.; Gomez, L.F.; Gangadharmath, U.B.; Mu, F.; Chen, G.; Walsh, J.C.; Szardenings, K.; Berman, D.S.; Kolb, H.C. Atherosclerotic plaque uptake of a novel integrin tracer 18F-Flotegatide in a mouse model of atherosclerosis. J. Nucl. Cardiol. 2014, 21, 553–562. [Google Scholar] [CrossRef]
- Hu, T.-T.; Vanhove, M.; Porcu, M.; Van Hove, I.; Van Bergen, T.; Jonckx, B.; Barbeaux, P.; Vermassen, E.; Feyen, J.H. The potent small molecule integrin antagonist THR-687 is a promising next-generation therapy for retinal vascular disorders. Exp. Eye Res. 2019, 180, 43–52. [Google Scholar] [CrossRef]
- Chen, H.; Qiao, J.; Li, Q.; Deng, J.; Tan, Z.; Guo, T.; Li, W. Safety, pharmacokinetic and pharmacodynamic studies of batifiban injection following single-and multiple-dose administration to healthy Chinese subjects. J. Huazhong Univ. Sci. Technol. [Med. Sci.] 2009, 29, 12–18. [Google Scholar] [CrossRef]
- Coller, B.S.; Scudder, L.E.; Beer, J.; Gold, H.K.; Folts, J.D.; Cavagnaro, J.; Jordan, R.; Wagner, C.; Iuliucci, J.; Knight, D. Monoclonal Antibodies to Platelet Glycoprotein IIb/IIIa as Antithrombotic Agents a. Ann. N. Y. Acad. Sci. 1991, 614, 193–213. [Google Scholar] [CrossRef]
- Jbara, M.; Bhogal, S.; Bajaj, K.; Chhabra, L. Abciximab-induced delayed profound thrombocytopaenia. Case Rep. 2017, 2017, bcr-2017-219379. [Google Scholar] [CrossRef]
- Zhu, X.; Cao, G. Safety of Glycoprotein IIb-IIIa Inhibitors Used in Stroke-Related Treatment: A Systematic Review and Meta-Analysis. Clin. Appl. Thromb. Hemost. 2020, 26, 1076029620942594. [Google Scholar] [CrossRef]



| Name | Function | Efficacy | Company |
|---|---|---|---|
| SF0166 | αvβ3 integrin inhibitor | effect | Scifulor |
| Flotegatide | RGD peptide | effect | Siemens Medical Solutions |
| Thr-687 | integrin antagonist | ongoing | Oxurion |
| Battifiban | platelet GPIIb/IIIa receptor antagonist | effect | DongRui (Hangzhou) Medical Technology |
| Abciximab | platelet GPIIb/IIIa receptor antagonist | effect | Janssen Biologics BV |
| Tirofiban | platelet GPIIb/IIIa receptor antagonist | effect | Medicure Pharma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruzha, Y.; Ni, J.; Quan, Z.; Li, H.; Qing, H. Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 12387. https://doi.org/10.3390/ijms232012387
Ruzha Y, Ni J, Quan Z, Li H, Qing H. Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases. International Journal of Molecular Sciences. 2022; 23(20):12387. https://doi.org/10.3390/ijms232012387
Chicago/Turabian StyleRuzha, Yelizhati, Junjun Ni, Zhenzhen Quan, Hui Li, and Hong Qing. 2022. "Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases" International Journal of Molecular Sciences 23, no. 20: 12387. https://doi.org/10.3390/ijms232012387
APA StyleRuzha, Y., Ni, J., Quan, Z., Li, H., & Qing, H. (2022). Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases. International Journal of Molecular Sciences, 23(20), 12387. https://doi.org/10.3390/ijms232012387