
Anion-Exchange Chromatography at the Service of Gene Therapy: Baseline Separation of Full/Empty Adeno-Associated Virus Capsids by Screening of Conditions and Step Gradient Elution Mode

 , and
, and
Abstract
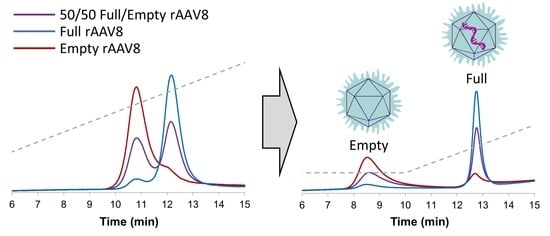
1. Introduction
2. Results and Discussion
2.1. Preliminary Selection of Column Hardware and Evaluation of Sample Stability
2.2. Optimization of the Mobile Phase Conditions
2.3. Optimization of the Step Gradient Method
2.3.1. Development of a Step Gradient Method
2.3.2. Method Validation by Full/Empty Quantification
3. Materials and Methods
3.1. Chemical and Reagents
3.2. Sample Preparation
3.3. Instrumentation and Experimental Conditions
3.3.1. Generic Linear Gradient
3.3.2. Optimized Linear Gradient
3.3.3. Optimized Linear Gradient including an Isocratic Step
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charpentier, E.; Doudna, J.A. Rewriting a genome. Nature 2013, 495, 50–51. [Google Scholar] [CrossRef] [PubMed]
- Dammes, N.; Peer, D. Paving the Road for RNA Therapeutics. Trends Pharmacol. Sci. 2020, 41, 755–775. [Google Scholar] [CrossRef]
- High, K.A.; Roncarolo, M.G. Gene Therapy. N. Engl. J. Med. 2019, 381, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Keeler, A.M.; Flotte, T.R. Recombinant Adeno-Associated Virus Gene Therapy in Light of Luxturna (and Zolgensma and Glybera): Where Are We, and How Did We Get Here? Annu. Rev. Virol. 2019, 6, 601–621. [Google Scholar] [CrossRef] [PubMed]
- Burger, C.; Gorbatyuk, O.S.; Velardo, M.J.; Peden, C.S.; Williams, P.; Zolotukhin, S.; Reier, P.J.; Mandel, R.J.; Muzyczka, N. Recombinant AAV Viral Vectors Pseudotyped with Viral Capsids from Serotypes 1, 2, and 5 Display Differential Efficiency and Cell Tropism after Delivery to Different Regions of the Central Nervous System. Mol. Ther. 2004, 10, 302–317. [Google Scholar] [CrossRef] [PubMed]
- White, S.J.; Nicklin, S.A.; Büning, H.; Brosnan, M.J.; Leike, K.; Papadakis, E.D.; Hallek, M.; Baker, A.H. Targeted Gene Delivery to Vascular Tissue In Vivo by Tropism-Modified Adeno-Associated Virus Vectors. Circulation 2004, 109, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Kaplitt, M.G.; Leone, P.; Samulski, R.J.; Xiao, X.; Pfaff, D.W.; O’Malley, K.L.; During, M.J. Long-term gene expression and phenotypic correction using adeno-associated virus vectors in the mammalian brain. Nat. Genet. 1994, 8, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Straus, S.E.; Sebring, E.D.; Rose, J.A. Concatemers of alternating plus and minus strands are intermediates in adenovirus-associated virus DNA synthesis. Proc. Natl. Acad. Sci. USA 1976, 73, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Dismuke, D.; Tenenbaum, L.; Samulski, R. Biosafety of Recombinant Adeno-associated Virus Vectors. Curr. Gene Ther. 2014, 13, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, M.D.; Kyöstiö, S.R.; Kotin, R.M.; Owens, R.A. Adeno-associated virus (AAV) Rep proteins mediate complex formation between AAV DNA and its integration site in human DNA. Proc. Natl. Acad. Sci. USA 1994, 91, 5808–5812. [Google Scholar] [CrossRef] [PubMed]
- Maguire, A.M.; Bennett, J.; Aleman, E.M.; Leroy, B.P.; Aleman, T.S. Clinical Perspective: Treating RPE65-Associated Retinal Dystrophy. Mol. Ther. 2021, 29, 442–463. [Google Scholar] [CrossRef] [PubMed]
- Chen, H. Adeno-associated virus vectors for human gene therapy. World J. Med. Genet. 2015, 5, 28. [Google Scholar] [CrossRef]
- Flotte, T.R. Empty Adeno-Associated Virus Capsids: Contaminant or Natural Decoy? Hum. Gene Ther. 2017, 28, 147–148. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Li, M.; Zhong, L.; Su, Q.; Li, J.; Li, S.; He, R.; Zhang, Y.; Hendricks, G.; Wang, J.; et al. Empty virions in AAV8 vector preparations reduce transduction efficiency and may cause total viral particle dose-limiting side effects. Mol. Ther.-Methods Clin. Dev. 2014, 1, 9. [Google Scholar] [CrossRef]
- FDA Guidance for Industry: Chemistry, Manufacturing, and Control (CMC) Information for Human Gene Therapy Investigational New Drug Applications (INDs). Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/chemistry-manufacturing-and-control-cmc-information-human-gene-therapy-investigational-new-drug (accessed on 7 June 2022).
- Schnödt, M.; Büning, H. Improving the Quality of Adeno-Associated Viral Vector Preparations: The Challenge of Product-Related Impurities. Hum. Gene Ther. Methods 2017, 28, 101–108. [Google Scholar] [CrossRef]
- Fu, X.; Chen, W.-C.; Argento, C.; Clarner, P.; Bhatt, V.; Dickerson, R.; Bou-Assaf, G.; Bakhshayeshi, M.; Lu, X.; Bergelson, S.; et al. Analytical Strategies for Quantification of Adeno-Associated Virus Empty Capsids to Support Process Development. Hum. Gene Ther. Methods 2019, 30, 144–152. [Google Scholar] [CrossRef]
- Gimpel, A.L.; Katsikis, G.; Sha, S.; Maloney, A.J.; Hong, M.S.; Nguyen, T.N.T.; Wolfrum, J.; Springs, S.L.; Sinskey, A.J.; Manalis, S.R.; et al. Analytical methods for process and product characterization of recombinant adeno-associated virus-based gene therapies. Mol. Ther.-Methods Clin. Dev. 2021, 20, 740–754. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, B.; Yarbrough, J.; Domsic, J.; Bennett, A.; Bothner, B.; Kozyreva, O.G.; Samulski, R.J.; Muzyczka, N.; McKenna, R.; Agbandje-McKenna, M. Structure and Dynamics of Adeno-Associated Virus Serotype 1 VP1-Unique N-Terminal Domain and Its Role in Capsid Trafficking. J. Virol. 2013, 87, 4974–4984. [Google Scholar] [CrossRef] [PubMed]
- Khatwani, S.L.; Pavlova, A.; Pirot, Z. Anion-exchange HPLC assay for separation and quantification of empty and full capsids in multiple adeno-associated virus serotypes. Mol. Ther.-Methods Clin. Dev. 2021, 21, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Urabe, M.; Xin, K.-Q.; Obara, Y.; Nakakura, T.; Mizukami, H.; Kume, A.; Okuda, K.; Ozawa, K. Removal of Empty Capsids from Type 1 Adeno-Associated Virus Vector Stocks by Anion-Exchange Chromatography Potentiates Transgene Expression. Mol. Ther. 2006, 13, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, R.; Argento, C.; Pieracci, J.; Bakhshayeshi, M. Separating Empty and Full Recombinant Adeno-Associated Virus Particles Using Isocratic Anion Exchange Chromatography. Biotechnol. J. 2021, 16, 2000015. [Google Scholar] [CrossRef] [PubMed]
- Kaludov, N.; Handelman, B.; Chiorini, J.A. Scalable Purification of Adeno-Associated Virus Type 2, 4, or 5 Using Ion-Exchange Chromatography. Hum. Gene Ther. 2002, 13, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Lock, M.; Alvira, M.R.; Wilson, J.M. Analysis of Particle Content of Recombinant Adeno-Associated Virus Serotype 8 Vectors by Ion-Exchange Chromatography. Hum. Gene Ther. Methods 2012, 23, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.R.; Stadalius, M.A.; Quarry, M.A. Gradient elution in reversed-phase HPLC-separation of macromolecules. Anal. Chem. 1983, 55, 1412A–1430A. [Google Scholar] [CrossRef]
- Fekete, S.; Bobály, B.; Nguyen, J.M.; Beck, A.; Veuthey, J.-L.; Wyndham, K.; Lauber, M.A.; Guillarme, D. Use of Ultrashort Columns for Therapeutic Protein Separations. Part 1: Theoretical Considerations and Proof of Concept. Anal. Chem. 2021, 93, 1277–1284. [Google Scholar] [CrossRef]
- Fekete, S.; Beck, A.; Veuthey, J.-L.; Guillarme, D. Proof of Concept to Achieve Infinite Selectivity for the Chromatographic Separation of Therapeutic Proteins. Anal. Chem. 2019, 91, 12954–12961. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, J.M.; Liu, X.; DeLoffi, M.; Murisier, A.; Fekete, S.; Guillarme, D.; Lauber, M.A. Aptamer-based immunoaffinity LC-MS using an ultra-short column for rapid attomole level quantitation of intact mAbs. J. Chromatogr. B 2021, 1173, 122694. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Name | Provider | Column Hardware | Column Dimensions (mm) | Particle Size (µm) |
|---|---|---|---|---|---|
| PP | ProPac SAX-10 | Thermo | Nonporous PEEK | 50 × 4 | 10 |
| PS | ProSwift SAX-1S | Thermo | Monolith PEEK-lined SS | 50 × 4.6 | - |
| AX | Bio-SAX | Agilent | Nonporous PEEK | 50 × 4.6 | 5 |
| QS | TSKgel Q-STAT | Tosoh | Nonporous SS tubing, PEEK frits | 100 × 4.6 | 7 |
| Buffer Name | Acronym | MW (g/mol) | pKa (25 °C) | Structure |
|---|---|---|---|---|
| 2-Amino-2-methyl-1,3-propanediol | AMPD | 105.15 | 8.8 |  |
| 1,3-Bis[tris(hydroxymethyl)methylamino]propane | BTP | 282.33 | 9.0 |  |
| N-(1,1-Dimethyl-2-hydroxyethyl)-3-amino-2-hydroxypropanesulfonic acid | AMPSO | 227.28 | 9.0 |  |
| 2-(Cyclohexylamino)ethanesulfonic acid | CHES | 207.29 | 9.5 |  |
| 3-(Cyclohexylamino)-2-hydroxy-1-propanesulfonic acid | CAPSO | 237.32 | 9.6 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aebischer, M.K.; Gizardin-Fredon, H.; Lardeux, H.; Kochardt, D.; Elger, C.; Haindl, M.; Ruppert, R.; Guillarme, D.; D’Atri, V. Anion-Exchange Chromatography at the Service of Gene Therapy: Baseline Separation of Full/Empty Adeno-Associated Virus Capsids by Screening of Conditions and Step Gradient Elution Mode. Int. J. Mol. Sci. 2022, 23, 12332. https://doi.org/10.3390/ijms232012332
Aebischer MK, Gizardin-Fredon H, Lardeux H, Kochardt D, Elger C, Haindl M, Ruppert R, Guillarme D, D’Atri V. Anion-Exchange Chromatography at the Service of Gene Therapy: Baseline Separation of Full/Empty Adeno-Associated Virus Capsids by Screening of Conditions and Step Gradient Elution Mode. International Journal of Molecular Sciences. 2022; 23(20):12332. https://doi.org/10.3390/ijms232012332
Chicago/Turabian StyleAebischer, Megane K., Hugo Gizardin-Fredon, Honorine Lardeux, Dominik Kochardt, Carsten Elger, Markus Haindl, Raphael Ruppert, Davy Guillarme, and Valentina D’Atri. 2022. "Anion-Exchange Chromatography at the Service of Gene Therapy: Baseline Separation of Full/Empty Adeno-Associated Virus Capsids by Screening of Conditions and Step Gradient Elution Mode" International Journal of Molecular Sciences 23, no. 20: 12332. https://doi.org/10.3390/ijms232012332
APA StyleAebischer, M. K., Gizardin-Fredon, H., Lardeux, H., Kochardt, D., Elger, C., Haindl, M., Ruppert, R., Guillarme, D., & D’Atri, V. (2022). Anion-Exchange Chromatography at the Service of Gene Therapy: Baseline Separation of Full/Empty Adeno-Associated Virus Capsids by Screening of Conditions and Step Gradient Elution Mode. International Journal of Molecular Sciences, 23(20), 12332. https://doi.org/10.3390/ijms232012332