
Role of Ion Channels in the Maintenance of Sperm Motility and Swimming Behavior in a Marine Teleost


Abstract
1. Introduction
2. Results
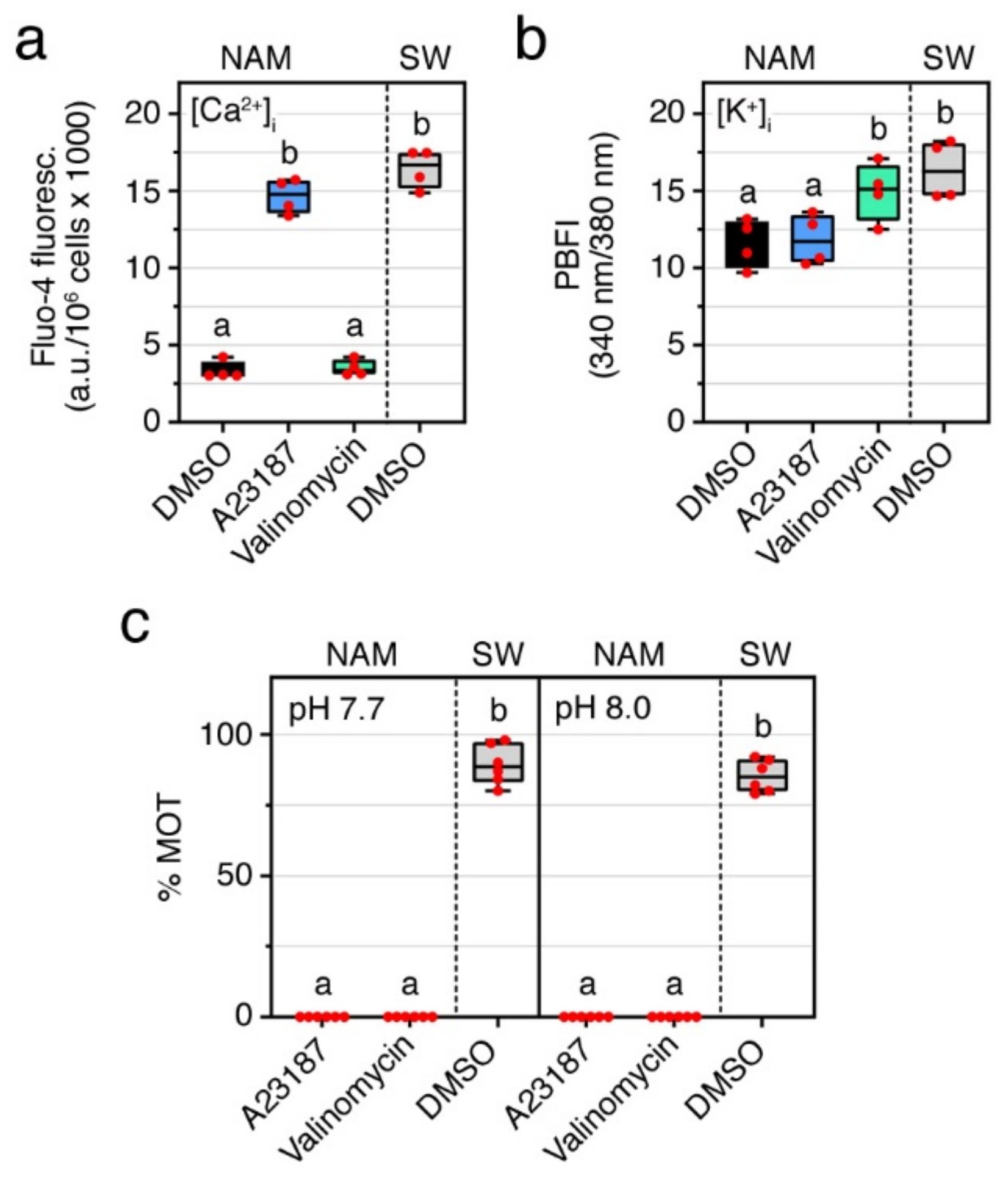
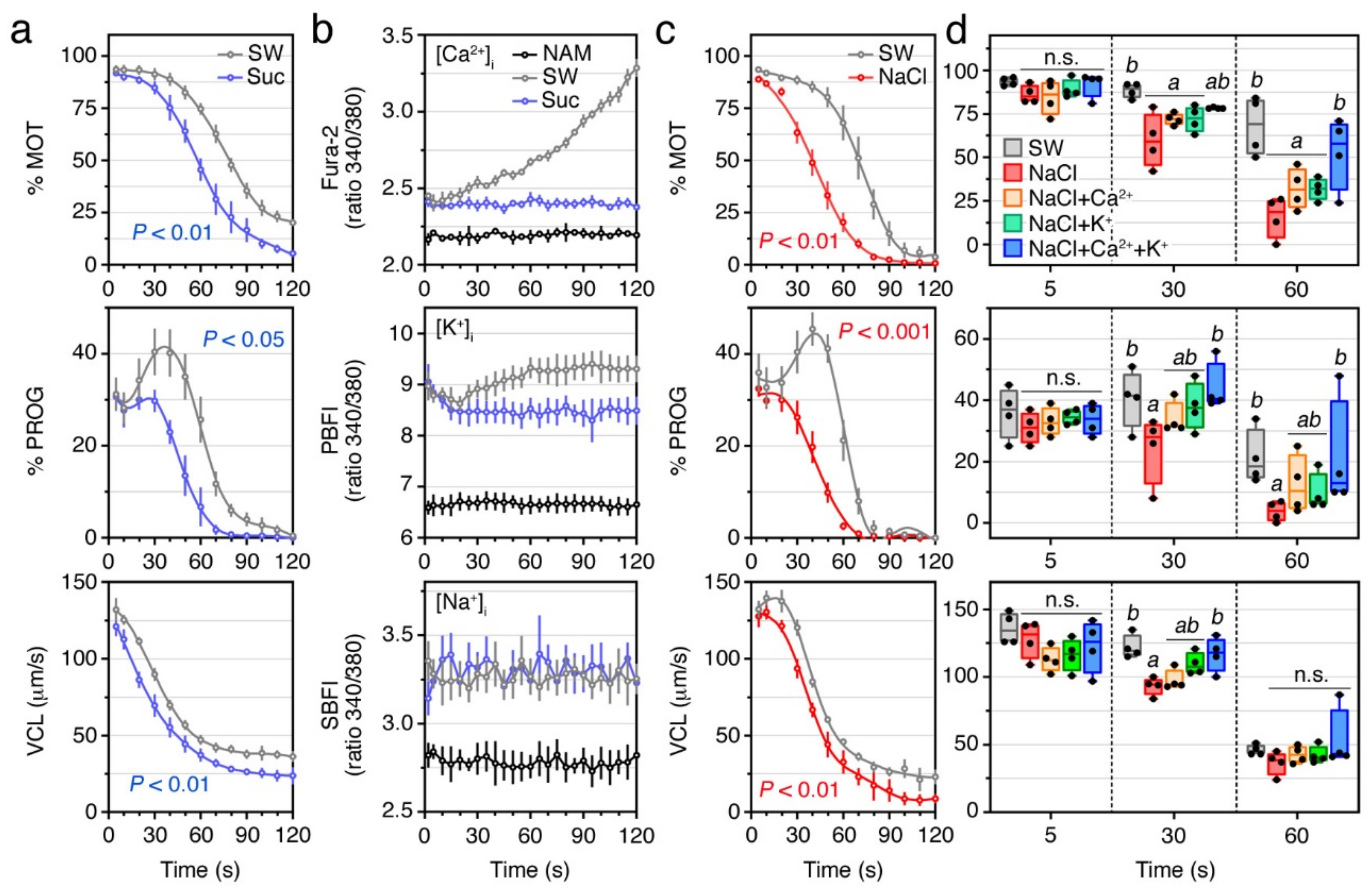
2.1. A Surge of Intracellular Ca2+, but Not of K+ or Na+, Is Necessary for Sperm Motility Activation in the Seabream
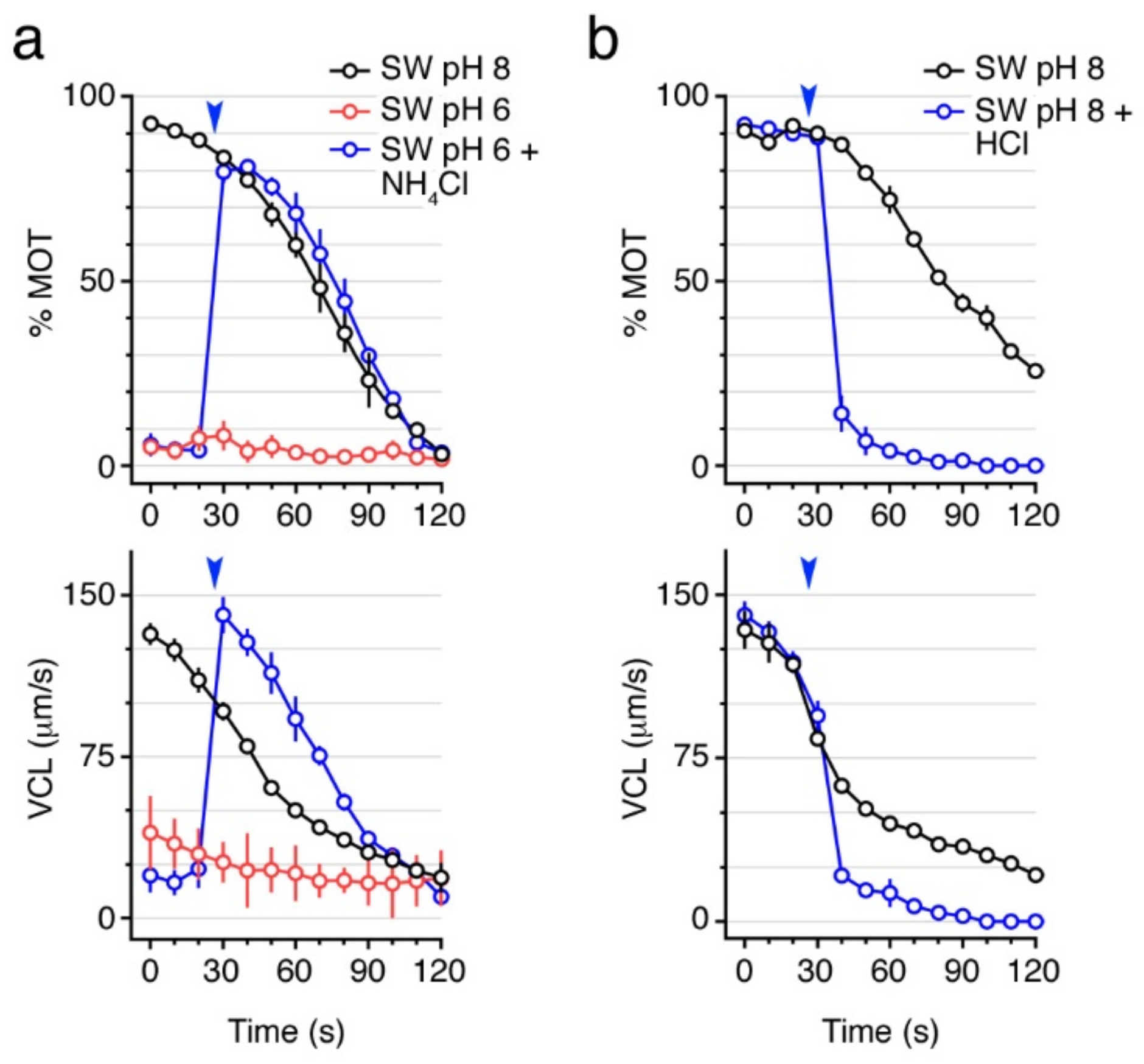
2.2. Basic pH Is Necessary to Activate and Maintain Sperm Motility
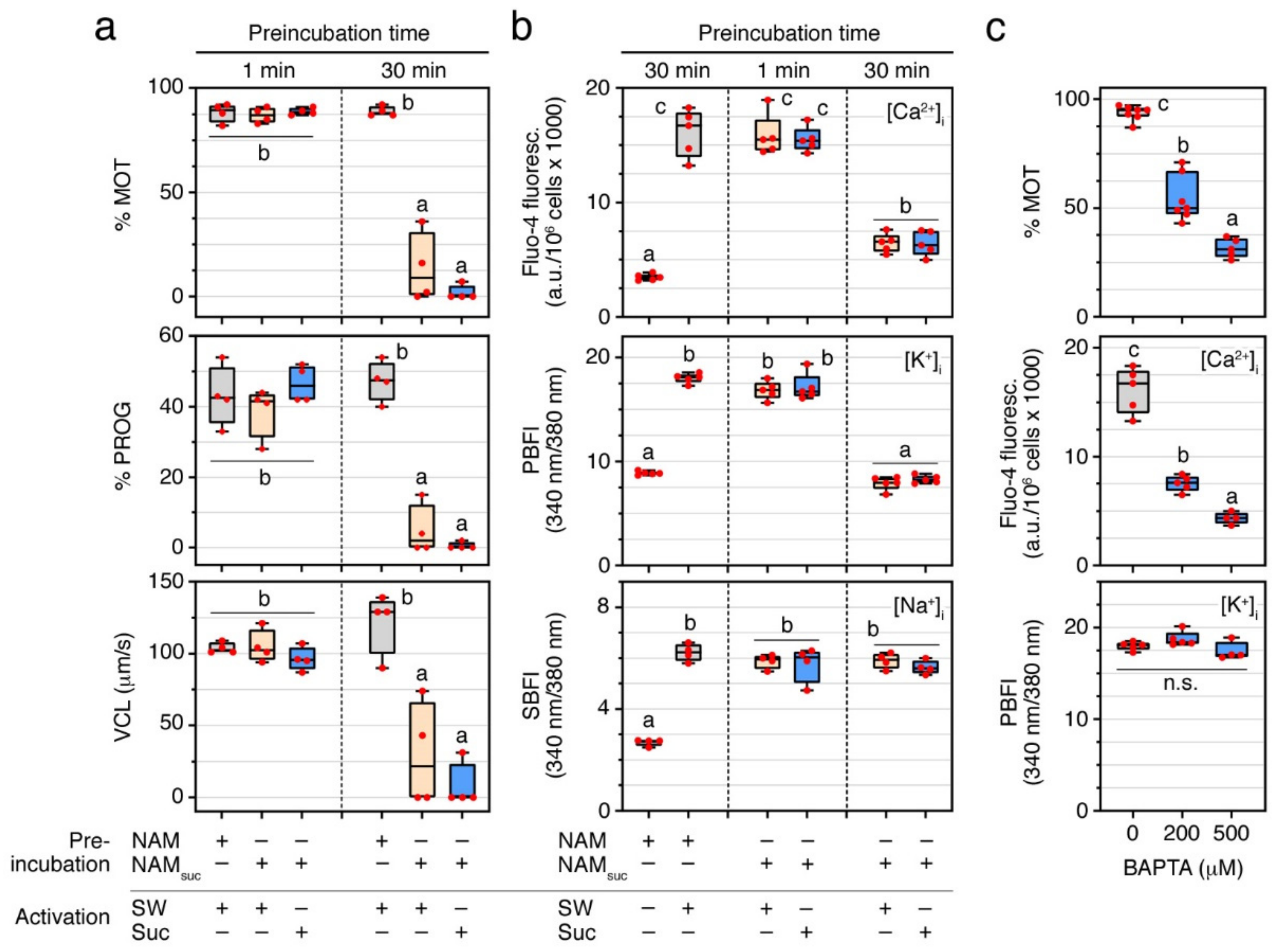
2.3. Ca2+ and pHi Are Not Sufficient to Trigger Sperm Motility
2.4. External Ca2+ and K+ Are Required for the Post Activation Maintenance of Sperm Motility
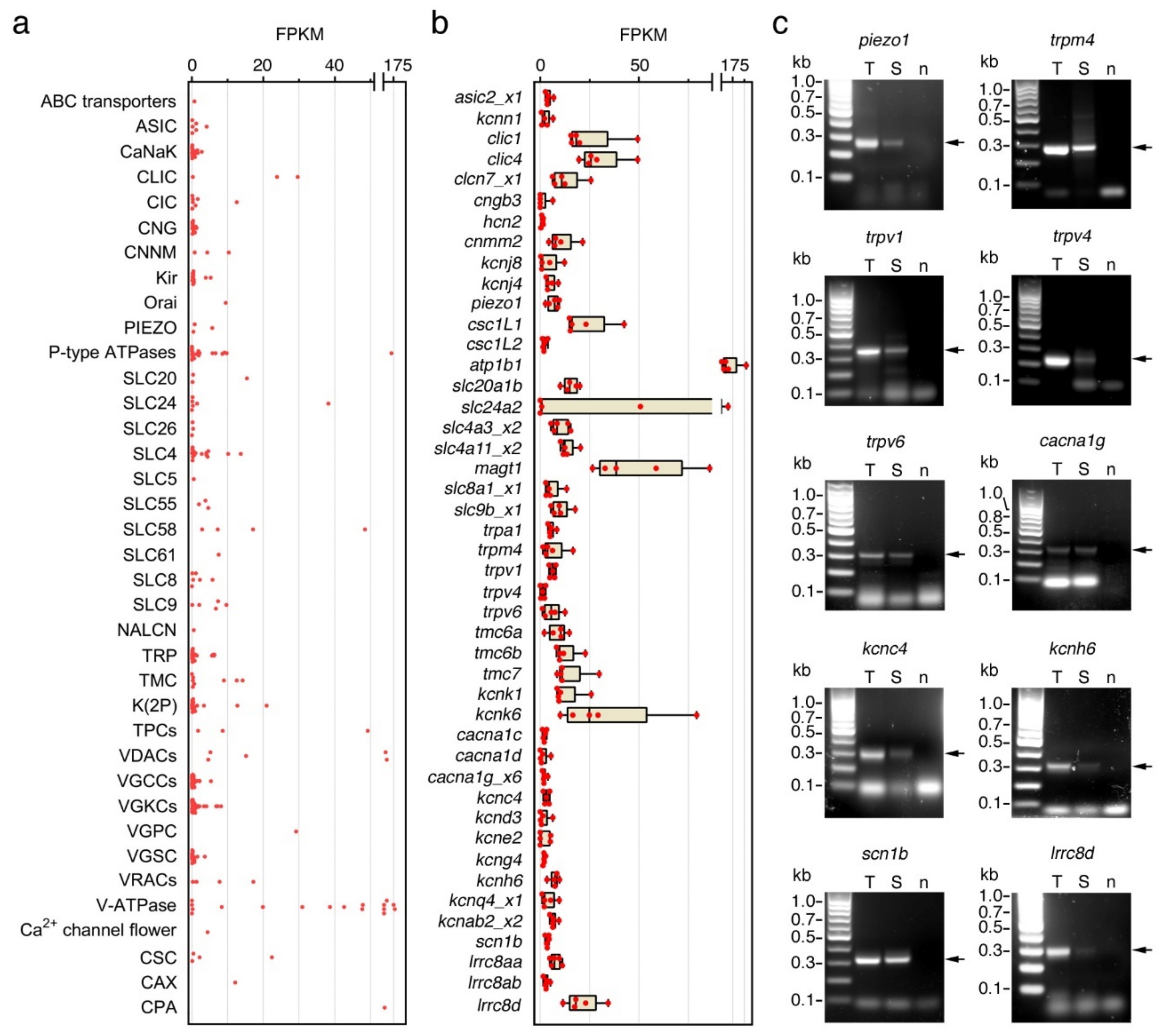
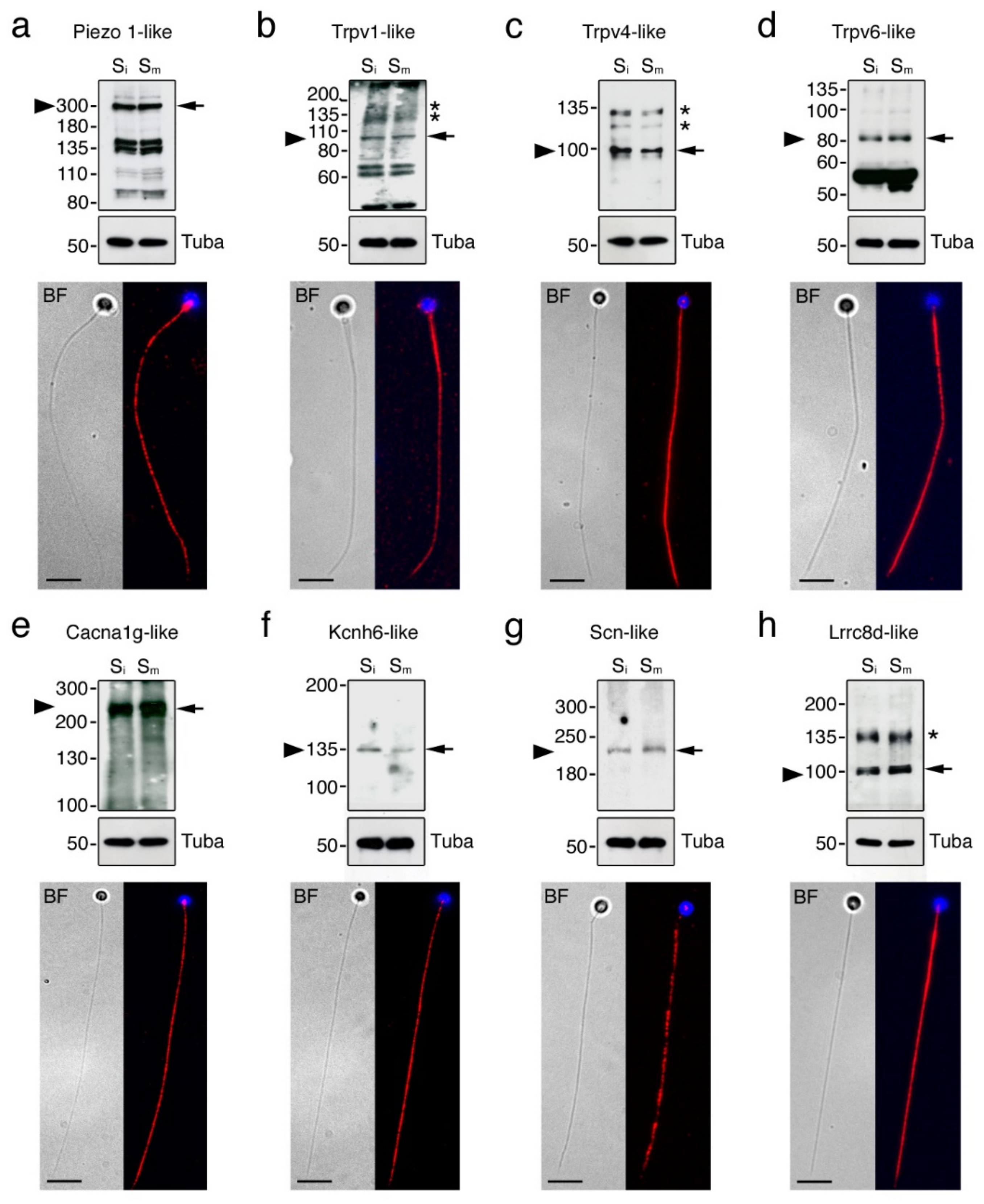
2.5. Seabream Spermatozoa Express Multiple Ion Channels
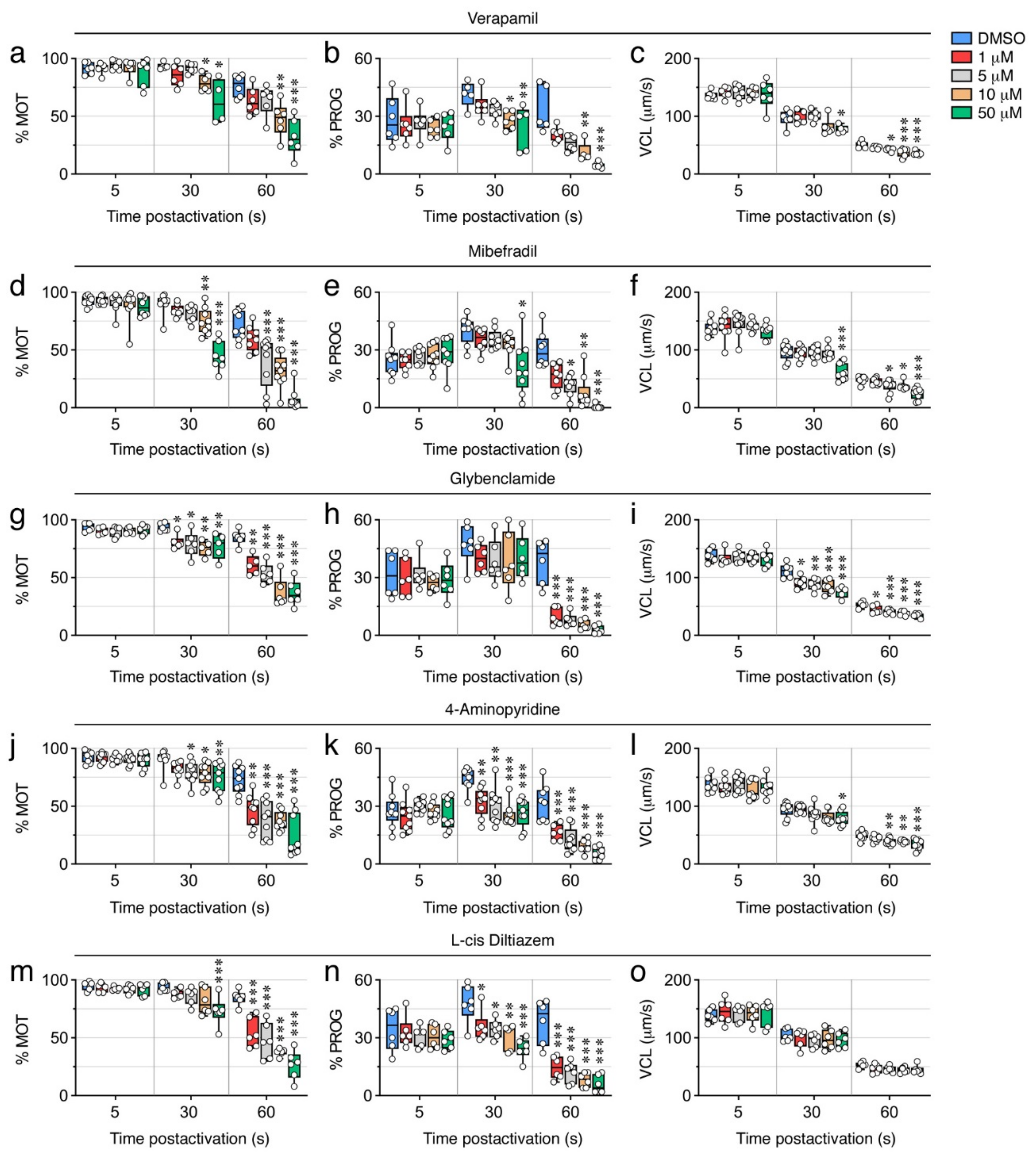
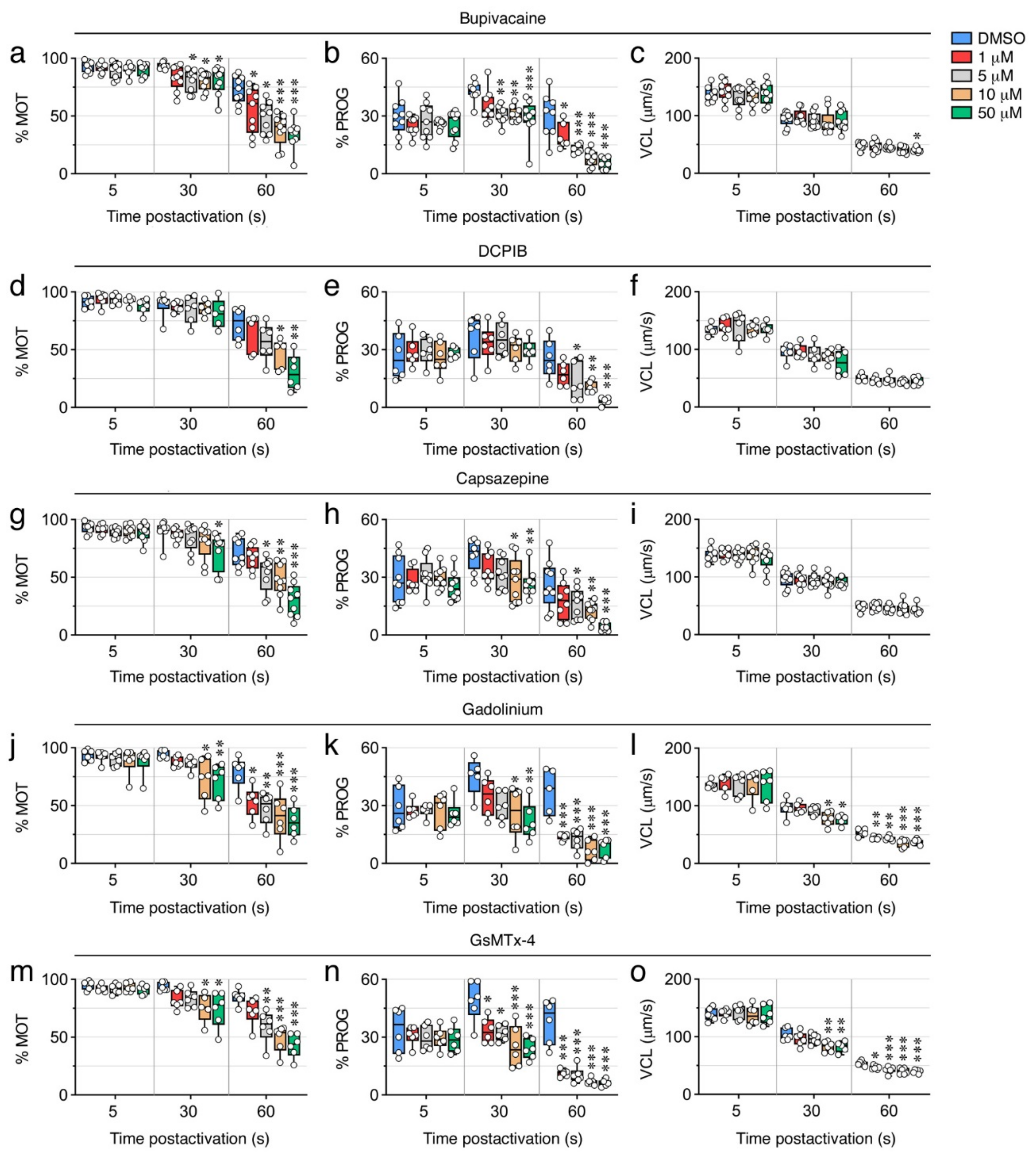
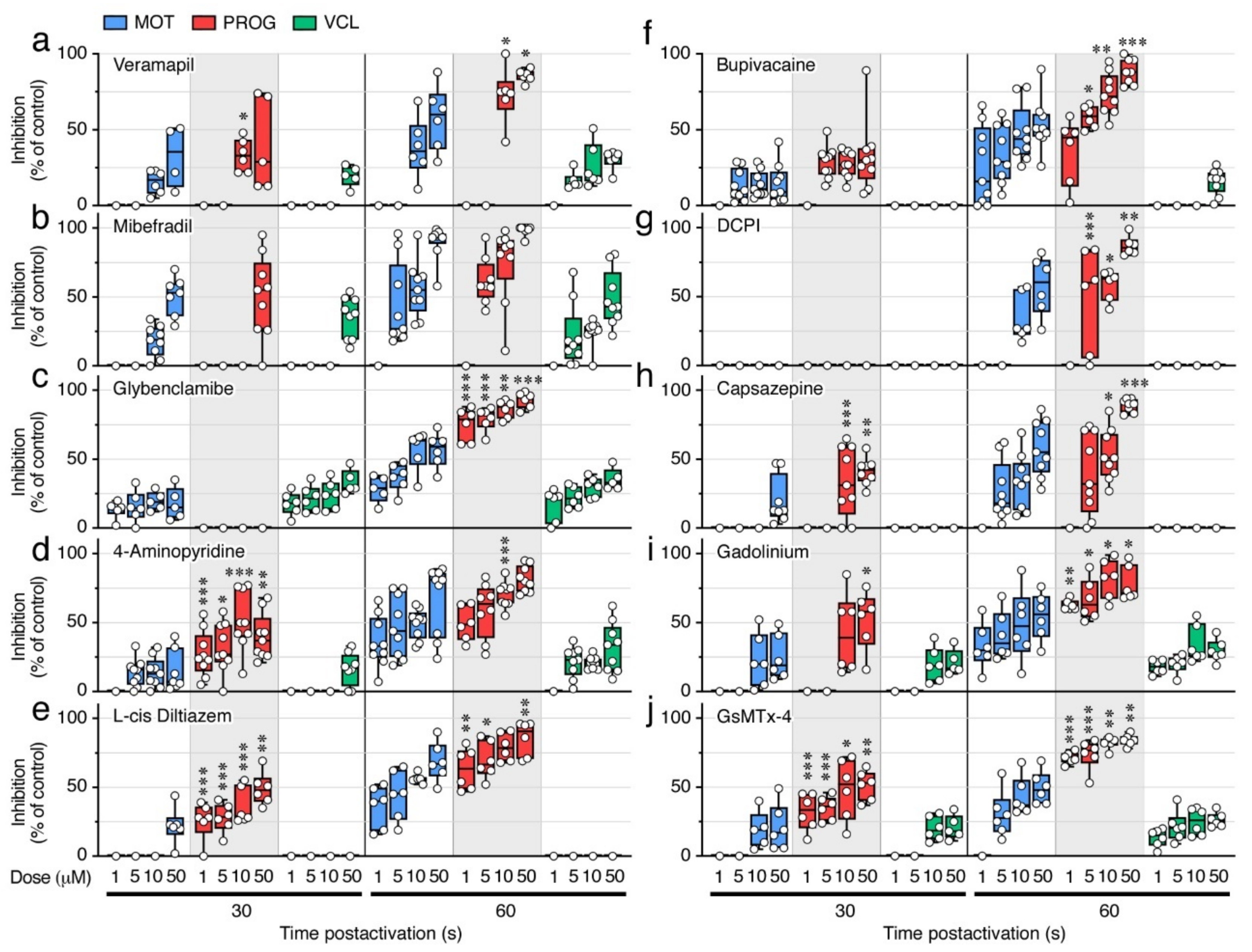
2.6. Ion Channel Blockers Affect the Motility of Post Activated Spermatozoa
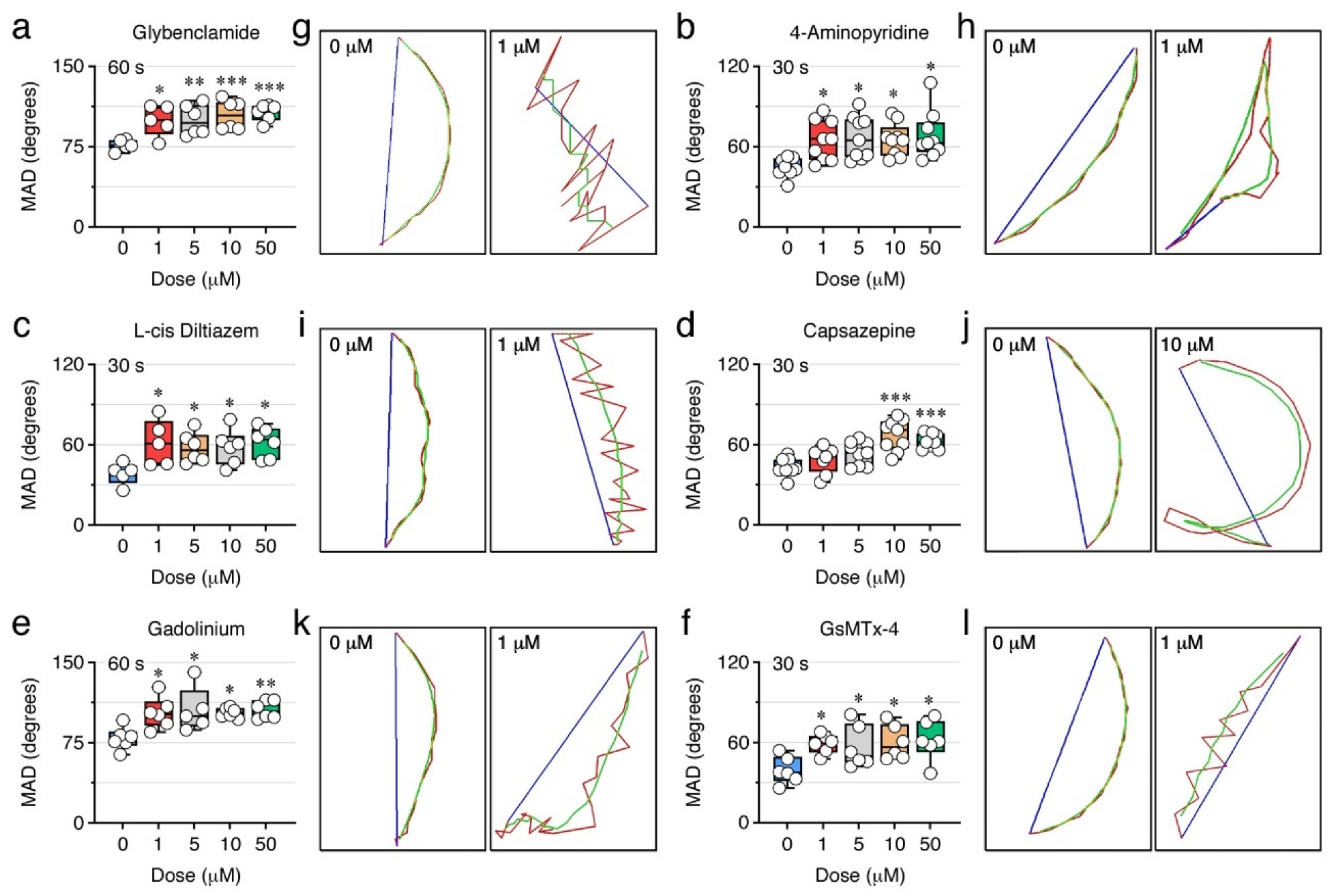
2.7. Ion Channels Can Control the Sperm Swimming Pattern
3. Discussion
4. Materials and Methods
4.1. Animals and Semen Collection
4.2. Reagents and Antibodies
4.3. Sperm Motility Assays
4.4. Measurement of Intracellular Ions
4.5. Meta-Analysis of a Seabream Sperm Transcriptome
4.6. RT-PCR Expression Analysis
4.7. Biochemical Fractionation of Spermatozoa and Protein Extraction
4.7.1. Total Sperm
4.7.2. Total Membrane
4.7.3. Isolation of Flagella
4.8. Immunoblotting
4.9. Immunofluorescence Microscopy
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pitnick, S.; Wolfner, M.F.; Dorus, S. Post-ejaculatory modifications to sperm (PEMS). Biol. Rev. Camb. Philos. Soc. 2020, 95, 365–392. [Google Scholar] [CrossRef] [PubMed]
- Alavi, S.M.H.; Cosson, J.; Bondarenko, O.; Linhart, O. Sperm motility in fishes: (III) diversity of regulatory signals from membrane to the axoneme. Theriogenology 2019, 136, 143–165. [Google Scholar] [CrossRef] [PubMed]
- Cosson, J.; Groison, A.; Suquet, M.; Fauvel, C.; Dreanno, C.; Billard, R. Studying sperm motility in marine fish: An overview on the state of the art. J. Appl. Ichthyol. 2008, 24, 460–486. [Google Scholar] [CrossRef]
- Darszon, A.; Nishigaki, T.; Beltran, C.; Treviño, C.L. Calcium channels in the development, maturation, and function of spermatozoa. Physiol. Rev. 2011, 91, 1305–1355. [Google Scholar] [CrossRef] [PubMed]
- Fechner, S.; Alvarez, L.; Bönigk, W.; Müller, A.; Berger, T.K.; Pascal, R.; Trötschel, C.; Poetsch, A.; Stölting, G.; Siegfried, K.R.; et al. A K+-selective CNG channel orchestrates Ca2+ signalling in zebrafish sperm. Elife 2015, 4, e07624. [Google Scholar] [CrossRef] [PubMed]
- Kaupp, U.B.; Strünker, T. Signaling in sperm: More different than similar. Trends Cell Biol. 2017, 27, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Brenker, C.; Goodwin, N.; Weyand, I.; Kashikar, N.D.; Naruse, M.; Krähling, M.; Müller, A.; Kaupp, U.B.; Strünker, T. The CatSper channel: A polymodal chemosensor in human sperm. EMBO J. 2012, 31, 1654–1665. [Google Scholar] [CrossRef]
- Nowicka-Bauer, K.; Szymczak-Cendlak, M. Structure and function of ion channels regulating sperm motility-an overview. Int. J. Mol. Sci. 2021, 22, 3259. [Google Scholar] [CrossRef]
- Dzyuba, V.; Cosson, J. Motility of fish spermatozoa: From external signaling to flagella response. Reprod. Biol. 2014, 14, 165–175. [Google Scholar] [CrossRef]
- Wobig, L.; Wolfenstetter, T.; Fechner, S.; Bönigk, W.; Körschen, H.G.; Jikeli, J.F.; Trötschel, C.; Feederle, R.; Kaupp, U.B.; Seifert, R.; et al. A family of hyperpolarization-activated channels selective for protons. Proc. Natl. Acad. Sci. USA 2020, 117, 13783–13791. [Google Scholar] [CrossRef]
- Lissabet, J.F.B.; Herrera Belén, L.; Lee-Estevez, M.; Risopatrón, J.; Valdebenito, I.; Figueroa, E.; Farías, J.G. The CatSper channel is present and plays a key role in sperm motility of the Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2020, 241, 110634. [Google Scholar] [CrossRef]
- Pérez, L.; Gallego, V.; Asturiano, J.F. Intracellular pH regulation and sperm motility in the European eel. Theriogenology 2020, 145, 48–58. [Google Scholar] [CrossRef]
- Zilli, L.; Schiavone, R.; Storelli, C.; Vilella, S. Molecular mechanism regulating axoneme activation in marine fish: A review. Int. Aquat. Res. 2012, 4, 2. [Google Scholar] [CrossRef][Green Version]
- Rafati, N.; Chen, J.; Herpin, A.; Pettersson, M.E.; Han, F.; Feng, C.; Wallerman, O.; Rubin, C.J.; Péron, S.; Cocco, A.; et al. Reconstruction of the birth of a male sex chromosome present in Atlantic herring. Proc. Natl. Acad. Sci. USA 2020, 117, 24359–24368. [Google Scholar] [CrossRef]
- Boj, M.; Chauvigné, F.; Cerdà, J. Coordinated Action of Aquaporins Regulates Sperm Motility in a Marine Teleost. Biol. Reprod. 2015, 93, 40. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Harumi, T.; Matsubara, H.; Yan, W.; Yuan, S.; Hirohashi, N.; Iida, T.; Yamaha, E.; Arai, K.; Matsubara, T.; et al. Chemical and physical guidance of fish spermatozoa into the egg through the micropyle. Biol. Reprod. 2017, 96, 780–799. [Google Scholar] [CrossRef]
- Kholodnyy, V.; Dzyuba, B.; Gadêlha, H.; Cosson, J.; Boryshpolets, S. Egg-sperm interaction in sturgeon: Role of ovarian fluid. Fish Physiol. Biochem. 2021, 47, 653–669. [Google Scholar] [CrossRef]
- Kholodnyy, V.; Dzyuba, B.; Rodina, M.; Bloomfield-Gadêlha, H.; Yoshida, M.; Cosson, J.; Boryshpolets, S. Does the rainbow trout ovarian fluid promote the spermatozoon on its way to the egg? Int. J. Mol. Sci. 2021, 22, 9519. [Google Scholar] [CrossRef]
- Devigili, A.; Cattelan, S.; Gasparini, C. Sperm accumulation induced by the female reproductive fluid: Putative evidence of chemoattraction using a new tool. Cells 2021, 10, 2472. [Google Scholar] [CrossRef]
- Majhi, R.K.; Kumar, A.; Yadav, M.; Swain, N.; Kumari, S.; Saha, A.; Pradhan, A.; Goswami, L.; Saha, S.; Samanta, L.; et al. Thermosensitive ion channel TRPV1 is endogenously expressed in the sperm of a fresh water teleost fish (Labeo rohita) and regulates sperm motility. Channels 2013, 7, 483–492. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.; Wang, F.; Chen, C.; Zhang, P.; Song, D.; Luo, T.; Xu, H.; Zeng, X. Sperm motility modulated by Trpv1 regulates zebrafish fertilization. Theriogenology 2020, 151, 41–51. [Google Scholar] [CrossRef]
- Zilli, L.; Schiavone, R.; Storelli, C.; Vilella, S. Molecular mechanisms determining sperm motility initiation in two sparids (Sparus aurata and Lithognathus mormyrus). Biol. Reprod. 2008, 79, 356–366. [Google Scholar] [CrossRef]
- Boitano, S.; Omoto, C.K. Membrane hyperpolarization activates trout sperm without an increase in intracellular pH. J. Cell Sci. 1991, 98, 343–349. [Google Scholar] [CrossRef]
- Takai, H.; Morisawa, M. Change in intracellular K+ concentration caused by external osmolality change regulates sperm motility of marine and freshwater teleosts. J. Cell Sci. 1995, 108, 1175–1181. [Google Scholar] [CrossRef]
- Márián, T.; Krasznai, Z.; Balkay, L.; Emri, M.; Trón, L. Role of extracellular and intracellular pH in carp sperm motility and modifications by hyperosmosis of regulation of the Na+/H+ exchanger. Cytometry 1997, 27, 374–382. [Google Scholar] [CrossRef]
- Tanaka, S.; Utoh, T.; Yamada, Y.; Horie, N.; Okamura, A.; Akazawa, A.; Mikawa, N.; Poka, H.; Kurokura, H. Role of sodium bicarbonate on the initiation of sperm motility in the Japanese eel. Fish. Sci. 2004, 70, 780–787. [Google Scholar] [CrossRef]
- Woolsey, J.; Ingermann, R.L. Acquisition of the potential for sperm motility insteelhead (Oncorhynchus mykiss): Effect of pH on dynein ATPase. Fish Physiol. Biochem. 2003, 19, 47–56. [Google Scholar] [CrossRef]
- Gallego, V.; Martínez-Pastor, F.; Mazzeo, I.; Peñaranda, D.S.; Herráez, M.P.; Asturiano, J.F. Intracellular changes in Ca2+, K+ and pH after sperm motility activation in the European eel (Anguilla anguilla): Preliminary results. Aquaculture 2014, 418–419, 155–158. [Google Scholar] [CrossRef]
- Oda, S.; Morisawa, M. Rises of intracellular Ca2+ and pH mediate the initiation of sperm motility by hyperosmolality in marine teleosts. Cell Motil. Cytoskelet. 1993, 25, 171–178. [Google Scholar] [CrossRef]
- Castro-Arnau, J.; Chauvigné, F.; Gómez-Garrido, J.; Esteve-Codina, A.; Dabad, M.; Alioto, T.; Finn, R.N.; Cerdà, J. Developmental RNA-Seq transcriptomics of haploid germ cells and spermatozoa uncovers novel pathways associated with teleost spermiogenesis. Sci. Rep. 2022, 12, 14162. [Google Scholar] [CrossRef]
- Napolitano, L.M.R.; Torre, V.; Marchesi, A. CNG channel structure, function, and gating: A tale of conformational flexibility. Pflugers Arch. 2021, 473, 1423–1435. [Google Scholar] [CrossRef]
- Ohta, H.; Kagawa, H.; Tanaka, H.; Unuma, T. Control by the environmental concentration of ions of the potential for motility in Japanese eel spermatozoa. Aquaculture 2001, 198, 339–351. [Google Scholar] [CrossRef]
- Pérez, L.; Vílchez, M.C.; Gallego, V.; Morini, M.; Peñaranda, D.S.; Asturiano, J.F. Role of calcium on the initiation of sperm motility in the European eel. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 191, 98–106. [Google Scholar] [CrossRef]
- Vílchez, M.C.; Morini, M.; Peñaranda, D.S.; Gallego, V.; Asturiano, J.F.; Pérez, L. Role of potassium and pH on the initiation of sperm motility in the European eel. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 203, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Vílchez, M.C.; Morini, M.; Peñaranda, D.S.; Gallego, V.; Asturiano, J.F.; Pérez, L. Sodium affects the sperm motility in the European eel. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 198, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Perchec-Poupard, G.; Gatti, J.L.; Cosson, J.; Jeulin, C.; Fierville, F.; Billard, R. Effects of extracellular environment on the osmotic signal transduction involved inactivation of motility of carp spermatozoa. J. Reprod. Fertil. 1997, 110, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Nishigaki, T.; José, O.; González-Cota, A.L.; Romero, F.; Treviño, C.L.; Darszon, A. Intracellular pH in sperm physiology. Biochem. Biophys. Res. Commun. 2014, 450, 1149–1158. [Google Scholar] [CrossRef]
- King, S.M.; Otter, T.; Witman, G.B. Purification and characterization of Chlamydomonas flagellar dyneins. Methods Enzymol. 1986, 134, 291–306. [Google Scholar]
- Bondarenko, V.; Cosson, J. Structure and beating behavior of the sperm motility apparatus in aquatic animals. Theriogenology 2019, 135, 152–163. [Google Scholar] [CrossRef]
- Gatti, J.L.; Billard, R.; Christen, R. Ionic regulation of the plasma membrane potential of rainbow trout (Salmo gairdneri) spermatozoa: Role in the initiation of sperm motility. J. Cell. Physiol. 1990, 143, 546–554. [Google Scholar] [CrossRef]
- Billard, R.; Cosson, J.; Crim, L.W. Motility of fresh and aged halibut sperm. Aquat. Living Resour. 1993, 6, 67–75. [Google Scholar] [CrossRef]
- Chauvaud, L.; Cosson, J.; Suquet, M.; Billard, R. Sperm motility in turbot, Scophtalmus maximus: Initiation of movement and changes with time of swimming characteristics. Environ. Biol. Fish 1995, 43, 341–349. [Google Scholar] [CrossRef]
- Effer, B.; Figueroa, E.; Augsburger, A.; Valdebenito, I. Sperm biology of Merluccius australis: Sperm structure, semen characteristics and effects of pH, temperature and osmolality on sperm motility. Aquaculture 2013, 408–409, 147–151. [Google Scholar] [CrossRef]
- Krasznai, Z.; Morisawa, M.; Krasznai, Z.T.; Morisawa, S.; Inaba, K.; Bazsane, Z.K.; Rubovszky, B.; Bodnar, B.; Borsos, A.; Marian, T. Gadolinium, a mechano-sensitivechannel blocker, inhibits osmosis-initiated motility of sea- and freshwaterfish sperm, but does not affect human or ascidian sperm motility. Cell Motil. Cytoskelet. 2003, 55, 232–243. [Google Scholar] [CrossRef]
- Boitano, S.; Omoto, C.K. Trout sperm swimming patterns and role of intracellular Ca++. Cell Motil. Cytoskelet. 1992, 21, 74–82. [Google Scholar] [CrossRef]
- Wehner, F.; Bondarava, M.; ter Veld, F.; Endl, E.; Nürnberger, H.R.; Li, T. Hypertonicity-induced cation channels. Acta Physiol. 2006, 187, 21–25. [Google Scholar] [CrossRef]
- Cosson, J.; Groison, A.L.; Suquet, M.; Fauvel, C.; Dreanno, C.; Billard, R. Marine fish spermatozoa: Racing ephemeral swimmers. Reproduction 2008, 136, 277–294. [Google Scholar] [CrossRef]
- Zilli, L.; Schiavone, R.; Chauvigné, F.; Cerdà, J.; Storelli, C.; Vilella, S. Evidence for the involvement of aquaporins in sperm motility activation of the teleost gilthead sea bream (Sparus aurata). Biol. Reprod. 2009, 81, 880–888. [Google Scholar] [CrossRef]
- Krasznai, Z.; Morisawa, M.; Morisawa, S.; Krasznai, Z.T.; Tron, L.; Marian, T. Role of ion channels and membrane potential in the initiation of carp sperm motility. Aquat. Living Resour. 2003, 16, 445–449. [Google Scholar] [CrossRef]
- Majhi, R.K.; Kumar, A.; Giri, S.C.; Goswami, C. Differential expression and localization of TRPV channels in the mature sperm of Anas platyrhynchos. Reprod. Domest. Anim. 2020, 55, 1619–1628. [Google Scholar] [CrossRef]
- Ramal-Sanchez, M.; Bernabò, N.; Valbonetti, L.; Cimini, C.; Taraschi, A.; Capacchietti, G.; Machado-Simoes, J.; Barboni, B. Role and modulation of TRPV1 in mammalian spermatozoa: An updated review. Int. J. Mol. Sci. 2021, 22, 4306. [Google Scholar] [CrossRef]
- Mundt, N.; Spehr, M.; Lishko, P.V. TRPV4 is the temperature-sensitive ion channel of human sperm. eLife 2018, 7, e35853. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Sucharita, S.; Majhi, R.K.; Tiwari, A.; Ghosh, A.; Pradhan, S.K.; Patra, B.K.; Dash, R.R.; Nayak, R.N.; Giri, S.C.; et al. TRPA1 is selected as a semi-conserved channel during vertebrate evolution due to its involvement in spermatogenesis. Biochem. Biophys. Res. Commun. 2019, 512, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, X.; Ye, H.; Gao, W.; Pu, X.; Yang, Z. Distribution profiles of transient receptor potential melastatin- and vanilloid-related channels in rat spermatogenic cells and sperm. Mol. Biol. Rep. 2010, 37, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Arias, R.D.; Vívenes, C.Y.; Camejo, M.I.; Piñero, S.; Proverbio, T.; Martínez, E.; Marín, R.; Proverbio, F. ATPases, ion exchangers and human sperm motility. Reproduction 2015, 149, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, D.S.; Swain, D.K.; Shah, N.; Yadav, H.P.; Nakade, U.P.; Singh, V.K.; Nigam, R.; Yadav, S.; Garg, S.K. Functional and molecular characterization of voltage gated sodium channel Nav 1.8 in bull spermatozoa. Theriogenology 2017, 90, 210–218. [Google Scholar] [CrossRef]
- Candenas, L.; Pinto, F.M.; Cejudo-Román, A.; González-Ravina, C.; Fernández-Sánchez, M.; Pérez-Hernández, N.; Irazusta, J.; Subirán, N. Veratridine-sensitive Na+ channels regulate human sperm fertilization capacity. Life Sci. 2018, 196, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Chauvigné, F.; Boj, M.; Vilella, S.; Finn, R.N.; Cerdà, J. Subcellular localization of selectively permeable aquaporins in the male germ line of a marine teleost reveals spatial redistribution in activated spermatozoa. Biol. Reprod. 2013, 89, 37. [Google Scholar] [CrossRef]
- Chauvigné, F.; Boj, M.; Finn, R.N.; Cerdà, J. Mitochondrial aquaporin-8-mediated hydrogen peroxide transport is essential for teleost spermatozoon motility. Sci. Rep. 2015, 5, 7789. [Google Scholar] [CrossRef]
- Harding, S.D.; Armstrong, J.F.; Faccenda, E.; Southan, C.; Alexander, S.P.H.; Davenport, A.P.; Pawson, A.J.; Spedding, M.; Davies, J.A. NC-IUPHAR. The IUPHAR/BPS guide to PHARMACOLOGY in 2022: Curating pharmacology for COVID-19, malaria and antibacterials. Nucleic Acids Res. 2022, 50, D1282–D1294. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound. | NAM | NAMsuc | NAM(-Ca2+,-K+) | NAM(-Ca2+) | NAM(-K+) |
|---|---|---|---|---|---|
| NaCl | 75 | - | 81.5 | 80 | 75.6 |
| KCl | 1.5 | - | - | 1.5 | - |
| MgCl2 | 12.9 | - | 12.9 | 12.9 | 12.9 |
| CaCl2 | 2.65 | - | - | - | 2.65 |
| NaHCO3 | 20 | - | 20 | 20 | 20 |
| Glucose | 4.4 | - | 4.4 | 4.4. | 4.4 |
| Bovine serum albumin | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 |
| Sucrose | - | 280 | - | - | - |
| pH | 7.7 | 7.7 | 7.7 | 7.7 | 7.7 |
| mOsm/kg | 280 | 280 | 280 | 280 | 280 |
| Compound | Selectivity |
|---|---|
| Verapamil | High voltage-activated (L-type) Ca2+ channels. |
| Mibefradil | Transient, low-voltage-activated (T-type) Ca2+ channels. |
| Glybenclamide | ATP-sensitive K+ channels. |
| 4-Aminopyridine | Voltage-dependent K+ channels. |
| L-cis Diltiazem | Cyclic nucleotide-gated channels (nonselective monovalent and divalent cations). |
| Bupivacaine | Voltage-gated Na+ channels. |
| DCPIB | Volume-regulated anion channels (VRAC) (transport of Cl−, taurine, glutamate). |
| Capsazepine | Transient receptor potential vanilloid (TRPV) channels (highly Ca2+ selective). |
| Gadolinium | Stretch-activated ion channels (SACs) (Ca2+, Na+ and K+ transport). |
| GsMTx-4 | Cationic mechanosensitive channels (MSCs) and SACs (Ca2+, Na+ and K+ transport). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Arnau, J.; Chauvigné, F.; Cerdà, J. Role of Ion Channels in the Maintenance of Sperm Motility and Swimming Behavior in a Marine Teleost. Int. J. Mol. Sci. 2022, 23, 12113. https://doi.org/10.3390/ijms232012113
Castro-Arnau J, Chauvigné F, Cerdà J. Role of Ion Channels in the Maintenance of Sperm Motility and Swimming Behavior in a Marine Teleost. International Journal of Molecular Sciences. 2022; 23(20):12113. https://doi.org/10.3390/ijms232012113
Chicago/Turabian StyleCastro-Arnau, Júlia, François Chauvigné, and Joan Cerdà. 2022. "Role of Ion Channels in the Maintenance of Sperm Motility and Swimming Behavior in a Marine Teleost" International Journal of Molecular Sciences 23, no. 20: 12113. https://doi.org/10.3390/ijms232012113
APA StyleCastro-Arnau, J., Chauvigné, F., & Cerdà, J. (2022). Role of Ion Channels in the Maintenance of Sperm Motility and Swimming Behavior in a Marine Teleost. International Journal of Molecular Sciences, 23(20), 12113. https://doi.org/10.3390/ijms232012113