
New Insights and Potential Therapeutic Targeting of CB2 Cannabinoid Receptors in CNS Disorders


Abstract
:1. Introduction
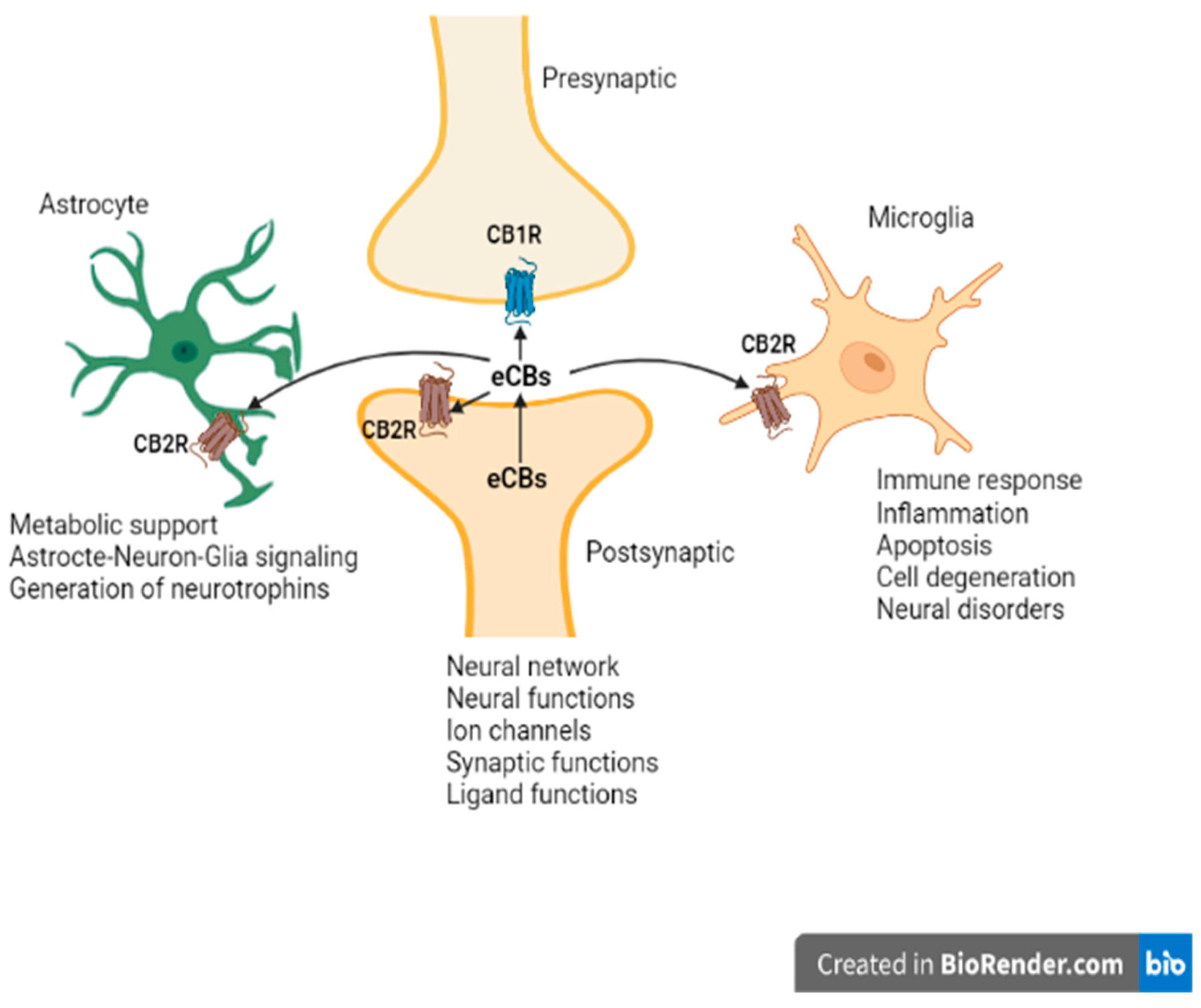
2. Overview of the ECS and New Insights into the Endocannabinoidome-eCBome
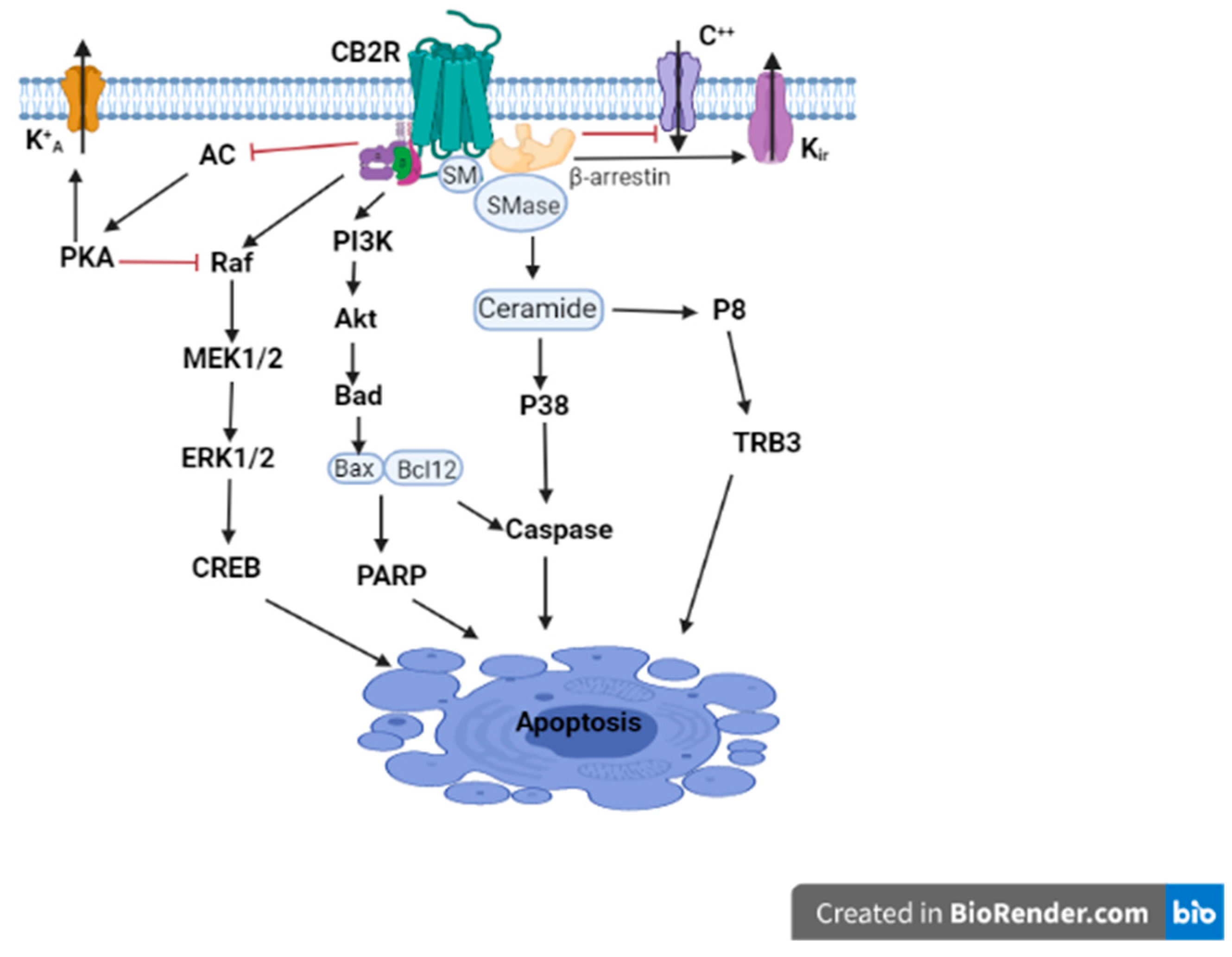
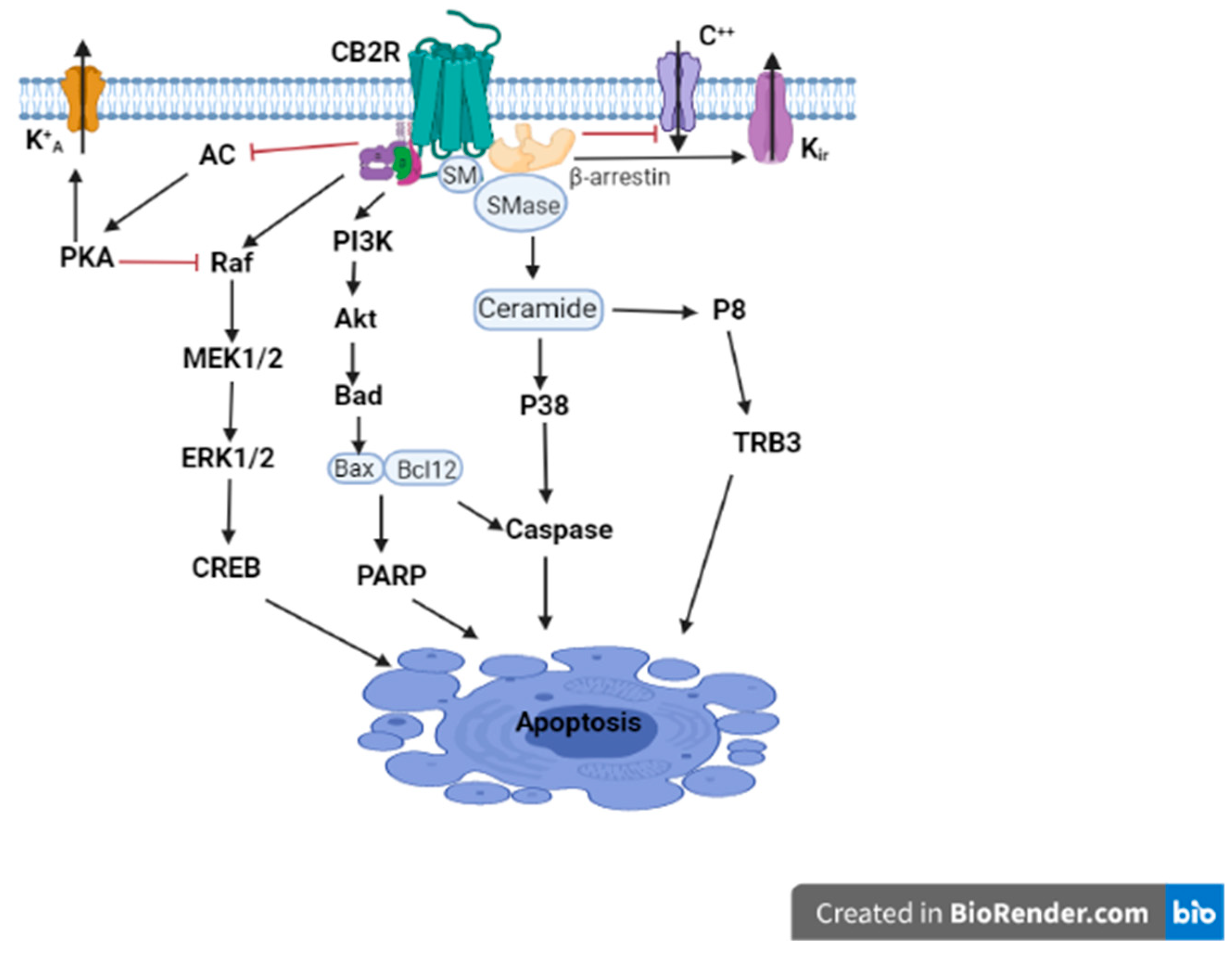
3. Unexpected CNS CB2R Expression and Function
4. Role of CB2Rs in CNS Disorders
4.1. Role of CB2Rs in Psychiatric Disorders
4.1.1. Anxiety-Related Disorders
4.1.2. Depression and CB2R-ECS
4.1.3. Schizophrenia and CB2R-ECS
4.1.4. CB2R-ECS in Addiction
4.1.5. CB2R-ECS in Autism Spectrum Disorder (ASD)
4.1.6. CB2R-ECS in Eating Disorders

{kind=link}
{kind=link}
| Disorder | Study Type | Model/Paradigm | CB2R Manipulation | Outcome | Reference |
|---|---|---|---|---|---|
| Anxiety/Anxiety-like behavior | Preclinical | Chronic mild stress/ Elevated plus maze | JWH015 | Induced angiogenesis | [18,42] |
| Marble burying | GW405833 | Induced anxiolysis | [73] | ||
| Chronic mild stress/ Zero maze | JWH015 | Reduced anxiety like behavior | [74] | ||
| AM630 | Increased anxiety-like behavior | ||||
| Light-dark box, Elevated plus maze | AM630 | Induced anxiogenesis and anxiolysis after acute and chronic administrations | [53] | ||
| CB2xP mice | No response to anxiogenic-like stimuli | ||||
| Light-dark box, Elevated plus maze | Cnr2−/− mice | Enhanced anxiety-like behavior | [56] | ||
| Light-dark box, Elevated plus maze, Forced swim and tail suspension | DAT-Cnr2 cKO mice, JWH133 | Increased anxiety-like behavior | [12] | ||
| Clinical | Children with anxiety | Significant relationship between rs2070956 polymorphism and treatment outcome | [75] | ||
| Depression | Preclinical | Tail suspension, novelty-suppressed feeding test, Chronic mild stress | CB2xP mice | Reduced immobility time | [72] |
| Light-dark box, Elevated plus maze | Cnr2−/− mice | Developed depressive-like behavior | [56] | ||
| Forced swim and tail suspension | DAT-Cnr2 cKO mice, JWH133 | Increased immobility time | [12] | ||
| Forced swim test | GW405833 | No change in time spent immobile | [77] | ||
| Chronic mild stress | JWH015 | Enhanced CB2R protein level | [17,45] | ||
| Chronic mild stress | CB2xP mice | Reduced CB2R mRNA | [72] | ||
| Clinical | Japanese depressive patients | High incidence of Q63R polymorphism of CB2R | [16] | ||
| Postmortem study | Reduced expression of CB2R gene | [81,82] | |||
| Schizophrenia | Preclinical | Early maternal deprivation in rats | Increased CB2R immunoreactivity in the hippocampus | [86] | |
| MK-801, methamphetamine | AM630 | Did not affect pre-pulse inhibition alone but enhanced MK-801 or methamphetamine induced effect | [119] | ||
| MK-801 | JWH015 | Enhanced pre-pulse impairment caused by MK-801 | [89] | ||
| Acoustic pre-pulse inhibition | Cnr2−/− mice | Decreased pre-pulse inhibition | [56,90] | ||
| MK-801, Pre-pulse inhibition | Cnr2−/− mice, AM630 | AM630 inhibited the ability of VU0467154 to reverse disruption of pre-pulse inhibition | [91] | ||
| Methylazoxymethanol acetate | Cannabidiol | Prevent schizophrenia-like deficits | [92,93] | ||
| Clinical | Schizophrenic patients | Significant reduction in the expression of CNR2 gene in peripheral mononuclear blood cells | [99] | ||
| Japanese schizophrenia patients | Significant association between CNR2 polymorphisms rs12744386 and rs2501432 | [119] | |||
| Chinese schizophrenic patients | Significant association between CNR2 polymorphisms rs22229579 and rs2501432 | [100] | |||
| Addiction | Preclinical | Self-administration, conditioned place preference | CB2xP | Decreased cocaine motor sensitization and self-administration | [102] |
| Open field test | Cnr2−/− mice | Enhanced cocaine motor sensitization | [56] | ||
| Open field, conditioned place preference | DAT-Cnr2 cKO mice | Increased psychostimulant induced motor sensitization and conditioned place preference | [55] | ||
| JWH133 | JWH133 inhibited cocaine and nicotine induced conditioned place preference | ||||
| Open field, conditioned place preference | Cnr2−/− mice, JWH133 | JWH133 blocked cocaine locomotion and self-administration | [41,103] | ||
| Drug self-administration under fixed and progressive ration | β-caryophyllene | Attenuated methamphetamine self-administration | [105] | ||
| Cnr2−/− mice | Blockage of β-caryophyllene induced reduction in methamphetamine self-administration | ||||
| Alcohol consumption and place preference | Cnr2−/− mice | Enhanced ethanol conditioned place preference | [106] | ||
| β-caryophyllene | Decreased ethanol consumption and preference | [107] | |||
| Chronic mild stress | JWH133 | Enhanced alcohol intake | [64,108] | ||
| Alcohol consumption | Increased amygdalar expression of CB2Rs | [109] | |||
| Clinical | Japanese alcoholic patients | Single nucleotide polymorphism R63Q in CNR2 gene | [64] | ||
| Autism Spectrum Disorder | Preclinical | BTBR T + tF/J | Increased level of CB2AR mRNA | [15] | |
| Clinical | Children with autism spectrum disorder | Up-regulation of expression of CNR2 gene in peripheral blood mononuclear cells | [113] | ||
| Eating Disorders | Clinical | Patients with eating disorders | No change in CB2R mRNA level in blood of the subjects | [117,118] | |
| Japanese patients with eating disorder | Link between CNR2 gene polymorphism and eating disorder | [119] |
4.2. Potential Role of CB2Rs in Neurologic and Neurodegenerative Disorders
4.2.1. CB2R-ECS in Alzheimer’s Disease (AD)
4.2.2. CB2R-ECS in Parkinson’s Disease (PD)
4.2.3. CB2R-ECS in Huntington’s Disease
4.2.4. CB2R-ECS in Multiple Sclerosis
4.2.5. CB2R-ECS in Amyotrophic Lateral Sclerosis
4.2.6. CB2R-ECS in Epilepsy
4.2.7. CB2R-ECS in Traumatic Brain Injury
| Disorder | Study Type | Model/Paradigm | CB2R Manipulation | Outcome | Reference |
|---|---|---|---|---|---|
| Alzheimer’s Disease | Preclinical | AβPP/PS1 transgenic mice | JWH133 | Reduced tau hyperphosphorylation, induced vasodilation | [128,129] |
| JWH133 | Enhanced brain glucose uptake | [130] | |||
| N-linoleyltyrosine | Protected neurons against Aβ injury | [131] | |||
| APPSw/Ind | JWH133 | CB2R activation blocks NMDA signaling in activated microglia | [132] | ||
| APP/PS1 mice | JWH015 | Improved novel object recognition, regulation in microglia-mediated neuroinflammation and dendritic complexity | [133] | ||
| Double transgenic APP/PS1 mice | 1-((3-benzyl-3-methyl-2,3-dihydro-1-benzofuran-6-yl) carbonyl) piperidine (MDA7) | Inhibited microglia activation, enhanced clearance of Aβ and decrease level of CB2R expression | [134,135] | ||
| Parkinson’s Disease | Preclinical | MPTP model | AM1241 | Prevented neurodegeneration | [145] |
| Lipopolysaccharide (LPS) model | Cnr2−/− | Enhanced activation of microglia | [150] | ||
| 6-hydroxydopamine (6-OHDA) | Δ9-THCV | Reduced motor inhibition and loss of TH-positive neurons caused by 6-OHDA, reduced CB2Rs up-regulation | [151] | ||
| Lipopolysaccharide (LPS) model | Exhibited a greater up-regulation of CB2Rs | ||||
| MPTP model | JWH015 | Protected MPTP induced neurodegeneration and suppress microglia activation | [152] | ||
| Rotenone (ROT) animal model | β-caryophyllene | Reduced oxidative stress and neuroinflammation | [153] | ||
| Clinical | PD patients | Elevated CB2Rs in microglia cells in the substantia nigra | [154] | ||
| Huntington’s Disease | Preclinical | Transgenic R6/2 mouse model | Elevated CB2R expression in the hippocampus, striatum and cerebellum | [158] | |
| Malonate rat model | Cnr2−/− | CB2R activation protected striatal neuron degeneration | [159] | ||
| R6/2 mice | Cnr2−/− | CB2R agonist suppresses motor deficits, synapse loss, and CNS inflammation | [160] | ||
| R6/2 mice | Cnr2−/− | Increased CB2R expression in striatal microglia | [161] | ||
| Multiple Sclerosis | Preclinical | Experimental autoimmune encephalomyelitis (EAE) | Enhanced CB2R expression | [164] | |
| HU308 | Reduced symptoms, axonal loss and microglia activation | [165] | |||
| O-1966 | Reduced immune cell invasion and improved neurologic functions | [166] | |||
| β-caryophyllene | Inhibited activation of immune cells and diminished axonal demyelination | [167] | |||
| Theiler’s murine encephalomyelitis virus model | JWH015 | Improved the neurological deficits in a long-lasting way, induced anti-inflammatory response | [169,170] | ||
| Clinical | Iranian multiple sclerosis patients | Significant association between Q63R gene polymorphism and multiple sclerosis | [171] | ||
| Multiple sclerosis patients | Elevated CB2R expression in B cells | [172] | |||
| Human postmortem specimen | Enhanced microglia cells | [173] | |||
| Postmortem brain tissue | Enhanced expression of CB2Rs | [174] | |||
| Amyotrophic Lateral Sclerosis | Preclinical | TDP-43 transgenic mice | Up-regulation of CB2Rs | [179] | |
| G93A-SOD1 mice | AM1241 | Slowed motor neuron degeneration and preserved motor function | [176] | ||
| Delayed disease progression | [177] | ||||
| Sativex® | Increased level of CB2Rs | [180] | |||
| Clinical | Postmortem specimen | Up-regulation of CB2R | [173] | ||
| Epilepsy | Preclinical | Palmitoylethanolamide (PEA) | AM630 | Blocked almitoylethanolamide induced seizure | [188] |
| Pilocarpine, pentylenetetrazole and isoniazid-induced epileptic seizure models | β-caryophyllene | Improved seizure activity | [189] | ||
| Pentylenetetrazole (PTZ) or methyl-6,7-dimethoxy-4-ethyl-beta-carboline-3-carboxylate (DMCM) | HU308 | No antiseizure effect | [190] | ||
| AM630 | Did not increase seizure activity | ||||
| Pentylenetetrazole (PTZ) | AM1241 | Increased seizure intensity | [191] | ||
| Pentylenetetrazole (PTZ) | Cnr2−/− | Increased susceptibility to seizure | [192] | ||
| JWH133 | Did not alter seizure susceptibility | ||||
| Traumatic Brain Injury | Preclinical | Controlled cortical impact (CCI) | O-1966 | Attenuated blood-brain barrier disruption and neural degeneration | [194] |
| Induced acute neuroprotection | [195] | ||||
| Experimental closed-head injury (CHI) | HU-910 | Enhanced neuroprotection and neurobehavioral recovery | [196] | ||
| Experimental closed-head injury (CHI) | Increased expression of CB2Rs | [59] | |||
| Controlled cortical impact (CCI) | JWH133 | Reduced white matter injury | [197] |
5. Conclusions and Future Therapeutic Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Basavarajappa, B.S.; Shivakumar, M.; Joshi, V.; Subbanna, S. Endocannabinoid system in neurodegenerative disorders. J. Neurochem. 2017, 142, 624–648. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V. The endocannabinoidome as a substrate for noneuphoric phytocannabinoid action and gut microbiome dysfunction in neuropsychiatric disorders. Dialogues Clin. Neurosci. 2020, 22, 259–269. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Ross, R.A. Cannabinoid receptors and their ligands. Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Scotter, E.L.; Abood, M.E.; Glass, M. The endocannabinoid system as a target for the treatment of neurodegenerative disease. Br. J. Pharmacol. 2010, 160, 480–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Galiègue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carrière, D.; Carayon, P.; Bouaboula, M.; Shire, D.; LE Fur, G.; Casellas, P. Expression of Central and Peripheral Cannabinoid Receptors in Human Immune Tissues and Leukocyte Subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef]
- Brown, S.M.; Wager-Miller, J.; Mackie, K. Cloning and molecular characterization of the rat CB2 cannabinoid receptor. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2002, 1576, 255–264. [Google Scholar] [CrossRef]
- Griffin, G.; Wray, E.J.; Tao, Q.; McAllister, S.D.; Rorrer, W.K.; Aung, M.; Martin, B.R.; Abood, M.E. Evaluation of the cannabinoid CB2 receptor-selective antagonist, SR144528: Further evidence for cannabinoid CB2 receptor absence in the rat central nervous system. Eur. J. Pharmacol. 1999, 377, 117–125. [Google Scholar] [CrossRef]
- Poso, A.; Huffman, J.W. Targeting the cannabinoid CB2 receptor: Modelling and structural determinants of CB2 selective ligands. Br. J. Pharmacol. 2008, 153, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Brusco, A.; Tagliaferro, P.; Saez, T.; Onaivi, E.S. Postsynaptic localization of CB2 cannabinoid receptors in the rat hippocampus. Synapse 2008, 62, 944–949. [Google Scholar] [CrossRef]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Neuroscience: Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.R.; Canseco-Alba, A.; Zhang, H.Y.; Tagliaferro, P.; Chung, M.; Dennis, E.; Sanabria, B.; Schanz, N.; Escosteguy-Neto, J.C.; Ishiguro, H.; et al. Cannabinoid type 2 receptors in dopamine neurons inhibits psychomotor behaviors, alters anxiety, depression and alcohol preference. Sci. Rep. 2017, 7, 17410. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Navarrete, F.; Gasparyan, A.; Manzanares, J. Therapeutic potential of the cannabinoid receptor 2 in neuropsychiatry. Explor. Neuroprot. Ther. 2021, 1, 55–71. [Google Scholar] [CrossRef]
- Ghose, T. The Scientist Magazine. 2009. Available online: https://www.the-scientist.com/uncategorized/back-on-top-44122 (accessed on 1 June 2009).
- Liu, Q.R.; Pan, C.H.; Hishimoto, A.; Li, C.Y.; Xi, Z.X.; Llorente-Berzal, A.; Viveros, M.P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; et al. Species differences in cannabinoid receptor 2 (CNR2 gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Brain neuronal CB2 cannabinoid receptors in drug abuse and depression: From mice to human subjects. PLoS ONE 2008, 3, e1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Functional expression of brain neuronal CB2 cannabinoid receptors are involved in the effects of drugs of abuse and in depression. Ann. N. Y. Acad. Sci. 2008, 1139, 434–449. [Google Scholar] [CrossRef] [Green Version]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the presence and functional expression of cannabinoid CB2 receptors in brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gu, S.; Liu, Q.R. CNS effects of CB2 cannabinoid receptors: Beyond neuro-immuno-cannabinoid activity. J. Psychopharmacol. 2012, 26, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Alhouayek, M.; Muccioli, G.G. The endocannabinoid system in inflammatory bowel diseases: From pathophysiology to therapeutic opportunity. Trends Mol. Med. 2012, 18, 615–625. [Google Scholar] [CrossRef]
- Howlett, A.C. Cannabinoid receptor signaling. Handb. Exp. Pharmacol. 2005, 168, 53–79. [Google Scholar] [CrossRef]
- Glass, M.; Felder, C.C. Concurrent stimulation of cannabinoid CB1 and dopamine D2 receptors augments cAMP accumulation in striatal neurons: Evidence for a G(s) linkage to the CB1 receptor. J. Neurosci. 1997, 17, 5327–5333. [Google Scholar] [CrossRef] [Green Version]
- Turu, G.; Hunyady, L. Signal transduction of the CB1 cannabinoid receptor. J. Mol. Endocrinol. 2010, 44, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Marsicano, G.; Lutz, B. Expression of the cannabinoid receptor CB1 in distinct neuronal subpopulations in the adult mouse forebrain. Eur. J. Neurosci. 1999, 11, 4213–4225. [Google Scholar] [CrossRef] [PubMed]
- Terzian, A.L.; Drago, F.; Wotjak, C.T.; Micale, V. The dopamine and cannabinoid interaction in the modulation of emotions and cognition: Assessing the role of cannabinoid CB1 receptor in neurons expressing dopamine D1 receptors. Front. Behav. Neurosci. 2011, 5, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzian, A.L.B.; Micale, V.; Wotjak, C.T. Cannabinoid receptor type 1 receptors on GABAergic vs. glutamatergic neurons differentially gate sex-dependent social interest in mice. Eur. J. Neurosci. 2014, 40, 2293–2298. [Google Scholar] [CrossRef]
- Llorente-Berzal, A.; Terzian, A.L.B.; Di Marzo, V.; Micale, V.; Viveros, M.P.; Wotjak, C.T. 2-AG promotes the expression of conditioned fear via cannabinoid receptor type 1 on GABAergic neurons. Psychopharmacology 2015, 232, 2811–2825. [Google Scholar] [CrossRef] [PubMed]
- Micale, V.; Stepan, J.; Jurik, A.; Pamplona, F.A.; Marsch, R.; Drago, F.; Eder, M.; Wotjak, C.T. Extinction of avoidance behavior by safety learning depends on endocannabinoid signaling in the hippocampus. J. Psychiatr. Res. 2017, 90, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Brusco, A.; Tagliaferro, P.A.; Saez, T.; Onaivi, E.S. Ultrastructural localization of neuronal brain CB2 cannabinoid receptors. Ann. N. Y. Acad. Sci. 2008, 1139, 450–457. [Google Scholar] [CrossRef]
- Mechoulam, R.; Parker, L.A. The endocannabinoid system and the brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Thomas, B.F.; Zhang, Y. Overcoming the Psychiatric Side Effects of the Cannabinoid CB1 Receptor Antagonists: Current Approaches for Therapeutics Development. Curr. Top. Med. Chem. 2019, 19, 1418–1435. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.A.; Grieb, M.; Lutz, B. Central side-effects of therapies based on CB1 cannabinoid receptor agonists and antagonists: Focus on anxiety and depression. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.; Xu, F.; Taylor, D.H.; Zhao, J.F.; Wu, J. The impact of cannabinoid type 2 receptors (CB2Rs) in neuroprotection against neurological disorders. Acta Pharmacol. Sin. 2020, 41, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Leonard, C.M.; Ishiguro, H.; Zhang, P.W.; Lin, Z.; Akinshola, B.E.; Uhl, G.R. Endocannabinoids and cannabinoid receptor genetics. Prog. Neurobiol. 2002, 66, 307–344. [Google Scholar] [CrossRef]
- Joshi, N.; Onaivi, E.S. Endocannabinoid System Components: Overview and Tissue Distribution. Recent Adv. Cannabinoid Physiol. Pathol. 2019, 1162, 1–12. [Google Scholar]
- Jordan, C.J.; Xi, Z.X. Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neurosci. Biobehav. Rev. 2019, 98, 208–220. [Google Scholar] [CrossRef]
- Schatz, A.R.; Lee, M.; Condie, R.B.; Pulaski, J.T.; Kaminski, N.E. Cannabinoid receptors CB1 and CB2: A characterization of expression and adenylate cyclase modulation within the immune system. Toxicol. Appl. Pharmacol. 1997, 142, 278–287. [Google Scholar] [CrossRef]
- McCoy, K.L.; Matveyeva, M.; Carlisle, S.J.; Cabral, G.A. Cannabinoid inhibition of the processing of intact lysozyme by macrophages: Evidence for CB2 receptor participation. J. Pharmacol. Exp. Ther. 1999, 289, 1620–1625. [Google Scholar]
- Buckley, N.E.; McCoy, K.L.; Mezey, É.; Bonner, T.; Zimmer, A.; Felder, C.C.; Glass, M.; Zimmer, A. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB2 receptor. Eur. J. Pharmacol. 2000, 396, 141–149. [Google Scholar] [CrossRef]
- Gong, J.P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef]
- Xi, Z.X.; Peng, X.Q.; Li, X.; Song, R.; Zhang, H.Y.; Liu, Q.R.; Yang, H.J.; Bi, G.H.; Li, J.; Gardner, E.L. Brain cannabinoid CB2 receptors modulate cocaine’s actions in mice. Nat. Neurosci. 2011, 14, 1160–1168. [Google Scholar] [CrossRef] [Green Version]
- Onaivi, E.S. Neuropsychobiological evidence for the functional presence and expression of cannabinoid CB2 receptors in the brain. Neuropsychobiology 2007, 54, 231–246. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Liu, Q.R. Future perspectives: Cannabinoid CB2 receptor ligands and their therapeutic potential in mental diseases. In Cannabinoids in Neurologic and Mental Disease; Academic Press: Cambridge, MA, USA, 2015; pp. 425–444. ISBN 9780124171244. [Google Scholar]
- Chin, C.L.; Tovcimak, A.E.; Hradil, V.P.; Seifert, T.R.; Hollingsworth, P.R.; Chandran, P.; Zhu, C.Z.; Gauvin, D.; Pai, M.; Wetter, J.; et al. Differential effects of cannabinoid receptor agonists on regional brain activity using pharmacological MRI. Br. J. Pharmacol. 2008, 153, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onaivi, E.S.; Ishiguro, H.; Sejal, P.; Meozzi, P.A.; Myers, L.; Tagliaferro, P.; Hope, B.; Leonard, C.M.; Uhl, G.R.; Brusco, A.; et al. Methods to study the behavioral effects and expression of CB2 cannabinoid receptor and its gene transcripts in the chronic mild stress model of depression. Methods Mol. Med. 2006, 123, 291–298. [Google Scholar] [CrossRef]
- Chen, D.J.; Gao, M.; Gao, F.F.; Su, Q.X.; Wu, J. Brain cannabinoid receptor 2: Expression, function and modulation. Acta Pharmacol. Sin. 2017, 38, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Bosier, B.; Muccioli, G.G.; Hermans, E.; Lambert, D.M. Functionally selective cannabinoid receptor signalling: Therapeutic implications and opportunities. Biochem. Pharmacol. 2010, 80, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Roche, M.; Finn, D.P. Brain CB2 receptors: Implications for neuropsychiatric disorders. Pharmaceuticals 2010, 3, 2517–2533. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.Y.; Gao, M.; Liu, Q.R.; Bi, G.H.; Li, X.; Yang, H.J.; Gardner, E.L.; Wu, J.; Xi, Z.X. Cannabinoid CB2 receptors modulate midbrain dopamine neuronal activity and dopamine-related behavior in mice. Proc. Natl. Acad. Sci. USA 2014, 111, E5007–E5015. [Google Scholar] [CrossRef] [Green Version]
- Emadi, L.; Jonaidi, H.; Amir Abad, E.H. The role of central CB2 cannabinoid receptors on food intake in neonatal chicks. J. Comp. Physiol. A 2011, 197, 1143–1147. [Google Scholar] [CrossRef]
- Ignatowska-Jankowska, B.; Jankowski, M.M.; Swiergiel, A.H. Cannabidiol decreases body weight gain in rats: Involvement of CB2 receptors. Neurosci. Lett. 2011, 490, 82–84. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Geresu, B.; Canseco-Alba, A.; Sanabria, B.; Lin, Z.; Liu, Q.R.; Onaivi, E.S.; Engidawork, E. Involvement of CB2 receptors in the neurobehavioral effects of Catha edulis (Vahl) Endl. (khat) in mice. Molecules 2019, 24, 3164. [Google Scholar] [CrossRef] [Green Version]
- Canseco-Alba, A.; Schanz, N.; Sanabria, B.; Zhao, J.; Lin, Z.; Liu, Q.R.; Onaivi, E.S. Behavioral effects of psychostimulants in mutant mice with cell-type specific deletion of CB2 cannabinoid receptors in dopamine neurons. Behav. Brain Res. 2019, 360, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Alvaro, A.; Aracil-Fernández, A.; García-Gutiérrez, M.S.; Navarrete, F.; Manzanares, J. Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacology 2011, 36, 1489–1504. [Google Scholar] [CrossRef] [Green Version]
- Svíženská, I.H.; Brázda, V.; Klusáková, I.; Dubový, P. Bilateral Changes of Cannabinoid Receptor Type 2 Protein and mRNA in the Dorsal Root Ganglia of a Rat Neuropathic Pain Model. J. Histochem. Cytochem. 2013, 61, 529–547. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.J.; Reiner, D.; Shen, H.; Wu, K.J.; Liu, Q.R.; Wang, Y. Time-dependent protection of CB2 receptor agonist in stroke. PLoS ONE 2015, 10, e0132487. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodriguez, A.B.; Acaz-Fonseca, E.; Viveros, M.P.; Garcia-Segura, L.M. Changes in cannabinoid receptors, aquaporin 4 and vimentin expression after traumatic brain injury in adolescent male mice. Association with edema and neurological deficit. PLoS ONE 2015, 10, 0128782. [Google Scholar] [CrossRef] [Green Version]
- Geresu, B.; Engidawork, E. Catha edulis F. (Khat) Reverses Haloperidol But Not Morphine Induced Motor Deficits Following Acute and Subacute Administration in Mice. Ethiop. Pharm. J. 2012, 28, 117–130. [Google Scholar] [CrossRef]
- Concannon, R.M.; Okine, B.N.; Finn, D.P.; Dowd, E. Differential upregulation of the cannabinoid CB2 receptor in neurotoxic and inflammation-driven rat models of Parkinson’s disease. Exp. Neurol. 2015, 269, 133–141. [Google Scholar] [CrossRef]
- Agudelo, M.; Yndart, A.; Morrison, M.; Figueroa, G.; Muñoz, K.; Samikkannu, T.; Nair, M.P. Differential expression and functional role of cannabinoid genes in alcohol users. Drug Alcohol Depend. 2013, 133, 789–793. [Google Scholar] [CrossRef] [Green Version]
- Rivera, P.; Miguéns, M.; Coria, S.M.; Rubio, L.; Higuera-Matas, A.; Bermúdez-Silva, F.J.; De Fonseca, F.R.; Suárez, J.; Ambrosio, E. Cocaine self-administration differentially modulates the expression of endogenous cannabinoid system-related proteins in the hippocampus of Lewis vs. Fischer 344 rats. Int. J. Neuropsychopharmacol. 2013, 16, 1277–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiguro, H.; Iwasaki, S.; Teasenfitz, L.; Higuchi, S.; Horiuchi, Y.; Saito, T.; Arinami, T.; Onaivi, E.S. Involvement of cannabinoid CB2 receptor in alcohol preference in mice and alcoholism in humans. Pharmacogenom. J. 2007, 7, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onaivi, E.S.; Carpio, O.; Ishiguro, H.; Schanz, N.; Uhl, G.R.; Benno, R. Behavioral effects of CB2 cannabinoid receptor activation and its influence on food and alcohol consumption. Ann. N. Y. Acad. Sci. 2008, 1139, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Onaivi, E.S.; Sharma, V. Cannabis for COVID-19: Can cannabinoids quell the cytokine storm? Futur. Sci. OA 2020, 6, FSO625. [Google Scholar] [CrossRef] [PubMed]
- Aso, E.; Ferrer, I. Cannabinoids for treatment of alzheimer’s disease: Moving toward the clinic. Front. Pharmacol. 2014, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Dittel, B.N. Unraveling the complexities of cannabinoid receptor 2 (CB2) immune regulation in health and disease. Immunol. Res. 2011, 51, 26–38. [Google Scholar] [CrossRef]
- Malfitano, A.M.; Basu, S.; Maresz, K.; Bifulco, M.; Dittel, B.N. What we know and do not know about the cannabinoid receptor 2 (CB2). Semin. Immunol. 2014, 26, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, B.; Marsicano, G.; Maldonado, R.; Hillard, C.J. The endocannabinoid system in guarding against fear, anxiety and stress. Nat. Rev. Neurosci. 2015, 16, 705–718. [Google Scholar] [CrossRef]
- Micale, V.; Di Marzo, V.; Sulcova, A.; Wotjak, C.T.; Drago, F. Endocannabinoid system and mood disorders: Priming a target for new therapies. Pharmacol. Ther. 2013, 138, 18–37. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, M.S.; Pérez-Ortiz, J.M.; Gutiérrez-Adán, A.; Manzanares, J. Depression-resistant endophenotype in mice overexpressing cannabinoid CB 2 receptors. Br. J. Pharmacol. 2010, 160, 1773–1784. [Google Scholar] [CrossRef] [Green Version]
- Valenzano, K.J.; Tafesse, L.; Lee, G.; Harrison, J.E.; Boulet, J.M.; Gottshall, S.L.; Mark, L.; Pearson, M.S.; Miller, W.; Shan, S.; et al. Pharmacological and pharmacokinetic characterization of the cannabinoid receptor 2 agonist, GW405833, utilizing rodent models of acute and chronic pain, anxiety, ataxia and catalepsy. Neuropharmacology 2005, 48, 658–672. [Google Scholar] [CrossRef]
- Ishiguro, H.; Horiuchi, Y.; Tabata, K.; Liu, Q.R.; Arinami, T.; Onaivi, E.S. Cannabinoid CB2 receptor gene and environmental interaction in the development of psychiatric disorders. Molecules 2018, 23, 1836. [Google Scholar] [CrossRef] [Green Version]
- Lester, K.J.; Coleman, J.R.I.; Roberts, S.; Keers, R.; Breen, G.; Bögels, S.; Creswell, C.; Hudson, J.L.; McKinnon, A.; Nauta, M.; et al. Genetic variation in the endocannabinoid system and response to Cognitive Behavior Therapy for child anxiety disorders. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2017, 174, 144–155. [Google Scholar] [CrossRef]
- Micale, V.; Tabiova, K.; Kucerova, J.; Drago, F. Role of the endocannabinoid system in depression: From preclinical to clinical evidence. In Cannabinoid Modulation of Emotion, Memory, and Motivation; Springer: New York, NY, USA, 2015; pp. 97–129. ISBN 9781493922949. [Google Scholar]
- Hu, B.; Doods, H.; Treede, R.D.; Ceci, A. Depression-like behaviour in rats with mononeuropathy is reduced by the CB2-selective agonist GW405833. Pain 2009, 143, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Mitchell, P.J. The validity of animal models of predisposition to depression. Behav. Pharmacol. 2002, 13, 169–188. [Google Scholar] [CrossRef]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef]
- Papp, M.; Moryl, E.; Willner, P. Pharmacological validation of the chronic mild stress model of depression. Eur. J. Pharmacol. 1996, 296, 129–136. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Navarrete, F.; Navarro, G.; Reyes-Resina, I.; Franco, R.; Lanciego, J.L.; Giner, S.; Manzanares, J. Alterations in Gene and Protein Expression of Cannabinoid CB2 and GPR55 Receptors in the Dorsolateral Prefrontal Cortex of Suicide Victims. Neurotherapeutics 2018, 15, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Salort, G.; Hernández-Hernández, E.; García-Fuster, M.J.; García-Sevilla, J.A. Regulation of cannabinoid CB1 and CB2 receptors, neuroprotective mTOR and pro-apoptotic JNK1/2 kinases in postmortem prefrontal cortex of subjects with major depressive disorder. J. Affect. Disord. 2020, 276, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, J.; Tabiova, K.; Drago, F.; Micale, V. Therapeutic potential of cannabinoids in schizophrenia. Recent Pat. CNS Drug Discov. 2014, 9, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, B.A.; De Bruin, N.M.W.J.; Van Den Kroonenburg, P.T.J.M.; Van Luijtelaar, E.L.J.M.; Cools, A.R. The effects of early maternal deprivation on auditory information processing in adult wistar rats. Biol. Psychiatry 2004, 55, 701–707. [Google Scholar] [CrossRef]
- Garner, B.; Wood, S.J.; Pantelis, C.; van den Buuse, M. Early maternal deprivation reduces prepulse inhibition and impairs spatial learning ability in adulthood: No further effect of post-pubertal chronic corticosterone treatment. Behav. Brain Res. 2007, 176, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Suárez, J.; Llorente, R.; Romero-Zerbo, S.Y.; Mateos, B.; Bermúdez-Silva, F.J.; De Fonseca, F.R.; Viveros, P. Early maternal deprivation induces gender-dependent changes on the expression of hippocampal CB1 and CB2 cannabinoid receptors of neonatal rats. Hippocampus 2009, 19, 623–632. [Google Scholar] [CrossRef]
- Errico, F.; Rossi, S.; Napolitano, F.; Catuogno, V.; Topo, E.; Fisone, G.; D’Aniello, A.; Centonze, D.; Usiello, A. D-aspartate prevents corticostriatal long-term depression and attenuates schizophrenia-like symptoms induced by amphetamine and MK-801. J. Neurosci. 2008, 28, 10404–10414. [Google Scholar] [CrossRef] [Green Version]
- Ishiguro, H.; Horiuchi, Y.; Ishikawa, M.; Koga, M.; Imai, K.; Suzuki, Y.; Morikawa, M.; Inada, T.; Watanabe, Y.; Takahashi, M.; et al. Brain Cannabinoid CB2 Receptor in Schizophrenia. Biol. Psychiatry 2010, 67, 974–982. [Google Scholar] [CrossRef]
- Khella, R.; Short, J.L.; Malone, D.T. CB2 receptor agonism reverses MK-801-induced disruptions of prepulse inhibition in mice. Psychopharmacology 2014, 231, 3071–3087. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Ortega-Álvaro, A.; Busquets-García, A.; Pérez-Ortiz, J.M.; Caltana, L.; Ricatti, M.J.; Brusco, A.; Maldonado, R.; Manzanares, J. Synaptic plasticity alterations associated with memory impairment induced by deletion of CB2 cannabinoid receptors. Neuropharmacology 2013, 73, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.J.; Wilson, J.M.; Remke, D.H.; Mahmood, M.S.; Uddin, M.J.; Wess, J.; Patel, S.; Marnett, L.J.; Niswender, C.M.; Jones, C.K.; et al. Antipsychotic-like Effects of M4 Positive Allosteric Modulators Are Mediated by CB2 Receptor-Dependent Inhibition of Dopamine Release. Neuron 2016, 91, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Stark, T.; Ruda-Kucerova, J.; Iannotti, F.A.; D’Addario, C.; Di Marco, R.; Pekarik, V.; Drazanova, E.; Piscitelli, F.; Bari, M.; Babinska, Z.; et al. Peripubertal cannabidiol treatment rescues behavioral and neurochemical abnormalities in the MAM model of schizophrenia. Neuropharmacology 2019, 146, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Stark, T.; Di Bartolomeo, M.; Di Marco, R.; Drazanova, E.; Platania, C.B.M.; Iannotti, F.A.; Ruda-Kucerova, J.; D’Addario, C.; Kratka, L.; Pekarik, V.; et al. Altered dopamine D3 receptor gene expression in MAM model of schizophrenia is reversed by peripubertal cannabidiol treatment. Biochem. Pharmacol. 2020, 177, 114004. [Google Scholar] [CrossRef]
- Waldo Zuardi, A.; Alexandre, S.; Crippa, J.E.C.; Hallak, J.; Bhattacharyya, S.; Atakan, Z.; Martin-Santos, R.K.; McGuire, P.; Silveira Guimaraes, F. A Critical Review of the Antipsychotic Effects of Cannabidiol: 30 Years of a Translational Investigation. Curr. Pharm. Des. 2012, 18, 5131–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, T.; Di Martino, S.; Drago, F.; Wotjak, C.T.; Micale, V. Phytocannabinoids and schizophrenia: Focus on adolescence as a critical window of enhanced vulnerability and opportunity for treatment. Pharmacol. Res. 2021, 174, 105938. [Google Scholar] [CrossRef] [PubMed]
- Galaj, E.; Xi, Z.X. Possible receptor mechanisms underlying cannabidiol effects on addictive-like behaviors in experimental animals. Int. J. Mol. Sci. 2021, 22, 134. [Google Scholar] [CrossRef]
- Scarante, F.F.; Ribeiro, M.A.; Almeida-Santos, A.F.; Guimarães, F.S.; Campos, A.C. Glial Cells and Their Contribution to the Mechanisms of Action of Cannabidiol in Neuropsychiatric Disorders. Front. Pharmacol. 2021, 11, 2258. [Google Scholar] [CrossRef]
- Navarro, G.; Gonzalez, A.; Sánchez-Morales, A.; Casajuana-Martin, N.; Gómez-Ventura, M.; Cordomí, A.; Busqué, F.; Alibés, R.; Pardo, L.; Franco, R. Design of Negative and Positive Allosteric Modulators of the Cannabinoid CB2 Receptor Derived from the Natural Product Cannabidiol. J. Med. Chem. 2021, 64, 9354–9364. [Google Scholar] [CrossRef] [PubMed]
- Bioque, M.; García-Bueno, B.; MacDowell, K.S.; Meseguer, A.; Saiz, P.A.; Parellada, M.; Gonzalez-Pinto, A.; Rodriguez-Jimenez, R.; Lobo, A.; Leza, J.C.; et al. Peripheral endocannabinoid system dysregulation in first-episode psychosis. Neuropsychopharmacology 2013, 38, 2568–2577. [Google Scholar] [CrossRef]
- Tong, D.; He, S.; Wang, L.; Jin, L.; Si, P.; Cheng, X. Association of single-nucleotide polymorphisms in the cannabinoid receptor 2 gene with schizophrenia in the Han Chinese population. J. Mol. Neurosci. 2013, 51, 454–460. [Google Scholar] [CrossRef]
- Manzanares, J.; Cabañero, D.; Puente, N.; García-Gutiérrez, M.S.; Grandes, P.; Maldonado, R. Role of the endocannabinoid system in drug addiction. Biochem. Pharmacol. 2018, 157, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aracil-Fernández, A.; Trigo, J.M.; García-Gutiérrez, M.S.; Ortega-Álvaro, A.; Ternianov, A.; Maldonado, R.; Manzanares, J. P.6.d.008 Decreased cocaine motor sensitisation and self-administration in mice overexpressing cannabinoid CB2 receptors. Eur. Neuropsychopharmacol. 2011, 21, S586–S587. [Google Scholar] [CrossRef]
- Lopes, J.B.; Bastos, J.R.; Costa, R.B.; Aguiar, D.C.; Moreira, F.A. The roles of cannabinoid CB1 and CB2 receptors in cocaine-induced behavioral sensitization and conditioned place preference in mice. Psychopharmacology 2020, 237, 385–394. [Google Scholar] [CrossRef]
- Blanco-Calvo, E.; Rivera, P.; Arrabal, S.; Vargas, A.; Pavón, F.J.; Serrano, A.; Castilla-Ortega, E.; Galeano, P.; Rubio, L.; Suárez, J.; et al. Pharmacological blockade of either cannabinoid CB1 or CB2 receptors prevents both cocaine-induced conditioned locomotion and cocaine-induced reduction of cell proliferation in the hippocampus of adult male rat. Front. Integr. Neurosci. 2014, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- He, X.H.; Galaj, E.; Bi, G.H.; He, Y.; Hempel, B.; Wang, Y.L.; Gardner, E.L.; Xi, Z.X. β-caryophyllene, an FDA-Approved Food Additive, Inhibits Methamphetamine-Taking and Methamphetamine-Seeking Behaviors Possibly via CB2 and Non-CB2 Receptor Mechanisms. Front. Pharmacol. 2021, 12, 722476. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Álvaro, A.; Ternianov, A.; Aracil-Fernández, A.; Navarrete, F.; García-Gutiérrez, M.S.; Manzanares, J. Role of cannabinoid CB2 receptor in the reinforcing actions of ethanol. Addict. Biol. 2015, 20, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Al Mansouri, S.; Ojha, S.; Al Maamari, E.; Al Ameri, M.; Nurulain, S.M.; Bahi, A. The cannabinoid receptor 2 agonist, β-caryophyllene, reduced voluntary alcohol intake and attenuated ethanol-induced place preference and sensitivity in mice. Pharmacol. Biochem. Behav. 2014, 124, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Pradier, B.; Erxlebe, E.; Markert, A.; Rácz, I. Interaction of cannabinoid receptor 2 and social environment modulates chronic alcohol consumption. Behav. Brain Res. 2015, 287, 163–171. [Google Scholar] [CrossRef]
- Sánchez-Marín, L.; Flores-López, M.; Pastor, A.; Gavito, A.L.; Suárez, J.; de la Torre, R.; Pavón, F.J.; Rodríguez de Fonseca, F.; Serrano, A. Acute stress and alcohol exposure during adolescence result in an anxious phenotype in adulthood: Role of altered glutamate/endocannabinoid transmission mechanisms. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 113, 110460. [Google Scholar] [CrossRef]
- Navarrete, F.; García-Gutiérrez, M.S.; Gasparyan, A.; Navarro, D.; Manzanares, J. CB2 receptor involvement in the treatment of substance use disorders. Biomolecules 2021, 11, 1556. [Google Scholar] [CrossRef]
- Do, L.L.T.N. American Psychiatric Association Diagnostic and Statistical Manual of Mental Disorders (DSM-IV). In Encyclopedia of Child Behavior and Development; Springer: New York, NY, USA, 2011; pp. 84–85. [Google Scholar]
- Chakrabarti, B.; Persico, A.; Battista, N.; Maccarrone, M. Endocannabinoid Signaling in Autism. Neurotherapeutics 2015, 12, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Siniscalco, D.; Sapone, A.; Giordano, C.; Cirillo, A.; De Magistris, L.; Rossi, F.; Fasano, A.; Bradstreet, J.J.; Maione, S.; Antonucci, N. Cannabinoid receptor type 2, but not type 1, is up-regulated in peripheral blood mononuclear cells of children affected by autistic disorders. J. Autism Dev. Disord. 2013, 43, 2686–2695. [Google Scholar] [CrossRef]
- Pietropaolo, S.; Marsicano, G. The role of the endocannabinoid system as a therapeutic target for autism spectrum disorder: Lessons from behavioral studies on mouse models. Neurosci. Biobehav. Rev. 2021, in press. [Google Scholar] [CrossRef]
- Di Marzo, V.; Petrosino, S. Endocannabinoids and the regulation of their levels in health and disease. Curr. Opin. Lipidol. 2007, 18, 129–140. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Monteleone, P.; Matias, I.; Martiadis, V.; De Petrocellis, L.; Maj, M.; Di Marzo, V. Blood levels of the endocannabinoid anandamide are increased in anorexia nervosa and in binge-eating disorder, but not in bulimia nervosa. Neuropsychopharmacology 2005, 30, 1216–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frieling, H.; Albrecht, H.; Jedtberg, S.; Gozner, A.; Lenz, B.; Wilhelm, J.; Hillemacher, T.; de Zwaan, M.; Kornhuber, J.; Bleich, S. Elevated cannabinoid 1 receptor mRNA is linked to eating disorder related behavior and attitudes in females with eating disorders. Psychoneuroendocrinology 2009, 34, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, H.; Carpio, O.; Horiuchi, Y.; Shu, A.; Higuchi, S.; Schanz, N.; Benno, R.; Arinami, T.; Onaivi, E.S. A nonsynonymous polymorphism in cannabinoid CB2 receptor gene is associated with eating disorders in humans and food intake is modified in mice by its ligands. Synapse 2010, 64, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Ali, T.; Park, H.Y.; Badshah, H.; Rehman, S.U.; Kim, M.O. Neuroprotective Effect of Fisetin Against Amyloid-Beta-Induced Cognitive/Synaptic Dysfunction, Neuroinflammation, and Neurodegeneration in Adult Mice. Mol. Neurobiol. 2017, 54, 2269–2285. [Google Scholar] [CrossRef]
- Schmöle, A.C.; Lundt, R.; Toporowski, G.; Hansen, J.N.; Beins, E.; Halle, A.; Zimmer, A. Cannabinoid Receptor 2-Deficiency Ameliorates Disease Symptoms in a Mouse Model with Alzheimer’s Disease-Like Pathology. J. Alzheimer’s Dis. 2018, 64, 379–392. [Google Scholar] [CrossRef]
- Vázquez, C.; Tolón, R.M.; Grande, M.T.; Caraza, M.; Moreno, M.; Koester, E.C.; Villaescusa, B.; Ruiz-Valdepeñas, L.; Fernández-Sánchez, F.J.; Cravatt, B.F.; et al. Endocannabinoid regulation of amyloid-induced neuroinflammation. Neurobiol. Aging 2015, 36, 3008–3019. [Google Scholar] [CrossRef]
- Van Der Stelt, M.; Mazzola, C.; Esposito, G.; Matias, I.; Petrosino, S.; De Filippis, D.; Micale, V.; Steardo, L.; Drago, F.; Iuvone, T.; et al. Endocannabinoids and β-amyloid-induced neurotoxicity in vivo: Effect of pharmacological elevation of endocannabinoid levels. Cell. Mol. Life Sci. 2006, 63, 1410–1424. [Google Scholar] [CrossRef]
- Benito, C.; Núñez, E.; Tolón, R.M.; Carrier, E.J.; Rábano, A.; Hillard, C.J.; Romero, J. Cannabinoid CB2 Receptors and Fatty Acid Amide Hydrolase Are Selectively Overexpressed in Neuritic Plaque-Associated Glia in Alzheimer’s Disease Brains. J. Neurosci. 2003, 23, 11136–11141. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, B.G.; Blázquez, C.; Gómez Del Pulgar, T.; Guzmán, M.; De Ceballos, M.L. Prevention of Alzheimer’s disease pathology by cannabinoids: Neuroprotection mediated by blockade of microglial activation. J. Neurosci. 2005, 25, 1904–1913. [Google Scholar] [CrossRef] [Green Version]
- Solas, M.; Francis, P.T.; Franco, R.; Ramirez, M.J. CB2 receptor and amyloid pathology in frontal cortex of Alzheimer’s disease patients. Neurobiol. Aging 2013, 34, 805–808. [Google Scholar] [CrossRef]
- Koppel, J.; Vingtdeux, V.; Marambaud, P.; d’Abramo, C.; Jimenez, H.; Stauber, M.; Friedman, R.; Davies, P. CB2 receptor deficiency increases amyloid pathology and alters tau processing in a transgenic mouse model of Alzheimer’s disease. Mol. Med. 2014, 20, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aso, E.; Juvés, S.; Maldonado, R.; Ferrer, I. CB2 cannabinoid receptor agonist ameliorates alzheimer-like phenotype in AβPP/PS1 mice. J. Alzheimer’s Dis. 2013, 35, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Dorado, J.; Villalba, N.; Prieto, D.; Brera, B.; Martín-Moreno, A.M.; Tejerina, T.; de Ceballos, M.L. Vascular dysfunction in a transgenic model of Alzheimer’s disease: Effects of CB1R and CB2R cannabinoid agonists. Front. Neurosci. 2016, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Köfalvi, A.; Lemos, C.; Martín-Moreno, A.M.; Pinheiro, B.S.; García-García, L.; Pozo, M.A.; Valério-Fernandes, Â.; Beleza, R.O.; Agostinho, P.; Rodrigues, R.J.; et al. Stimulation of brain glucose uptake by cannabinoid CB2 receptors and its therapeutic potential in Alzheimer’s disease. Neuropharmacology 2016, 110, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, C.M.; Zheng, Q.X.; Zhou, Y.; Liu, Y.T.; Gong, L.P.; Zeng, Y.C.; Liu, S. N-linoleyltyrosine exerts neuroprotective effects in APP/PS1 transgenic mice via cannabinoid receptor-mediated autophagy. J. Pharmacol. Sci. 2021, 147, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santisteban, R.; Lillo, A.; Lillo, J.; Rebassa, J.-B.; Contestí, J.S.; Saura, C.A.; Franco, R.; Navarro, G. N-Methyl-D-aspartate (NMDA) and cannabinoid CB2 receptors form functional complexes in cells of the central nervous system: Insights into the therapeutic potential of neuronal and microglial NMDA receptors. Alzheimers Res. Ther. 2021, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shi, J.; Wang, B.; Li, J.; Jia, H. CB2 cannabinoid receptor agonist ameliorates novel object recognition but not spatial memory in transgenic APP/PS1 mice. Neurosci. Lett. 2019, 707, 134286. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bie, B.; Yang, H.; Xu, J.J.; Brown, D.L.; Naguib, M. Activation of the CB2 receptor system reverses amyloid-induced memory deficiency. Neurobiol. Aging 2013, 34, 791–804. [Google Scholar] [CrossRef]
- Wu, J.; Hocevar, M.; Foss, J.F.; Bie, B.; Naguib, M. Activation of CB2 receptor system restores cognitive capacity and hippocampal Sox2 expression in a transgenic mouse model of Alzheimer’s disease. Eur. J. Pharmacol. 2017, 811, 12–20. [Google Scholar] [CrossRef]
- Sheng, W.S.; Hu, S.; Min, X.; Cabral, G.A.; Lokensgard, J.R.; Peterson, P.K. Synthetic cannabinoid WIN55,212-2 inhibits generation of inflammatory mediators by IL-1β-stimulated human astrocytes. Glia 2005, 49, 211–219. [Google Scholar] [CrossRef]
- Ehrhart, J.; Obregon, D.; Mori, T.; Hou, H.; Sun, N.; Bai, Y.; Klein, T.; Fernandez, F.; Tan, J.; Shytle, D. Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation. J. Neuroinflamm. 2005, 2, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aso, E.; Andrés-Benito, P.; Carmona, M.; Maldonado, R.; Ferrer, I. Cannabinoid receptor 2 participates in amyloid-β processing in a mouse model of Alzheimer’s disease but plays a minor role in the therapeutic properties of a cannabis-based medicine. J. Alzheimer’s Dis. 2016, 51, 489–500. [Google Scholar] [CrossRef] [Green Version]
- de Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Branchi, I.; D’Andrea, I.; Armida, M.; Cassano, T.; Pèzzola, A.; Potenza, R.L.; Morgese, M.G.; Popoli, P.; Alleva, E. Nonmotor symptoms in Parkinson’s disease: Investigating early-phase onset of behavioral dysfunction in the 6-hydroxydopamine-lesioned rat model. J. Neurosci. Res. 2008, 86, 2050–2061. [Google Scholar] [CrossRef] [PubMed]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the: Substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar] [CrossRef]
- Ouchi, Y.; Yoshikawa, E.; Sekine, Y.; Futatsubashi, M.; Kanno, T.; Ogusu, T.; Torizuka, T. Microglial activation and dopamine terminal loss in early Parkinson’s disease. Ann. Neurol. 2005, 57, 168–175. [Google Scholar] [CrossRef]
- Gerhard, A.; Pavese, N.; Hotton, G.; Turkheimer, F.; Es, M.; Hammers, A.; Eggert, K.; Oertel, W.; Banati, R.B.; Brooks, D.J. In vivo imaging of microglial activation with [11C](R)-PK11195 PET in idiopathic Parkinson’s disease. Neurobiol. Dis. 2006, 21, 404–412. [Google Scholar] [CrossRef]
- Shi, J.; Cai, Q.; Zhang, J.; He, X.; Liu, Y.; Zhu, R.; Jin, L. AM1241 alleviates MPTP-induced Parkinson’s disease and promotes the regeneration of DA neurons in PD mice. Oncotarget 2017, 8, 67837–67850. [Google Scholar] [CrossRef] [Green Version]
- Centonze, D.; Finazzi-Agrò, A.; Bernardi, G.; Maccarrone, M. The endocannabinoid system in targeting inflammatory neurodegenerative diseases. Trends Pharmacol. Sci. 2007, 28, 180–187. [Google Scholar] [CrossRef]
- Shohami, E.; Cohen-Yeshurun, A.; Magid, L.; Algali, M.; Mechoulam, R. Endocannabinoids and traumatic brain injury. Br. J. Pharmacol. 2011, 163, 1402–1410. [Google Scholar] [CrossRef] [Green Version]
- Pertwee, R.G. Targeting the endocannabinoid system with cannabinoid receptor agonists: Pharmacological strategies and therapeutic possibilities. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3353–3363. [Google Scholar] [CrossRef]
- Micale, V.; Mazzola, C.; Drago, F. Endocannabinoids and neurodegenerative diseases. Pharmacol. Res. 2007, 56, 382–392. [Google Scholar] [CrossRef]
- Gómez-Gálvez, Y.; Palomo-Garo, C.; Fernández-Ruiz, J.; García, C. Potential of the cannabinoid CB2 receptor as a pharmacological target against inflammation in Parkinson’s disease. Prog. Neuro-Psychopharmacolo. Biol. Psychiatry 2016, 64, 200–208. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Palomo-Garo, C.; García-Arencibia, M.; Ramos, J.A.; Pertwee, R.G.; Fernández-Ruiz, J. Symptom-relieving and neuroprotective effects of the phytocannabinoid Δ 9-THCV in animal models of Parkinson’s disease. Br. J. Pharmacol. 2011, 163, 1495–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, D.A.; Martinez, A.A.; Seillier, A.; Koek, W.; Acosta, Y.; Fernandez, E.; Strong, R.; Lutz, B.; Marsicano, G.; Roberts, J.L.; et al. WIN55,212-2, a cannabinoid receptor agonist, protects against nigrostriatal cell loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. Eur. J. Neurosci. 2009, 29, 2177–2186. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Azimullah, S.; Haque, M.E.; Ojha, S.K. Cannabinoid type 2 (CB2) receptors activation protects against oxidative stress and neuroinflammation associated dopaminergic neurodegeneration in rotenone model of parkinson’s disease. Front. Neurosci. 2016, 10, 321. [Google Scholar] [CrossRef] [Green Version]
- García, M.C.; Cinquina, V.; Palomo-Garo, C.; Rábano, A.; Fernández-Ruiz, J. Identification of CB2 receptors in human nigral neurons that degenerate in Parkinson’s disease. Neurosci. Lett. 2015, 587, 1–4. [Google Scholar] [CrossRef]
- Eddings, C.R.; Arbez, N.; Akimov, S.; Geva, M.; Hayden, M.R.; Ross, C.A. Pridopidine protects neurons from mutant-huntingtin toxicity via the sigma-1 receptor. Neurobiol. Dis. 2019, 129, 118–129. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J. The endocannabinoid system as a target for the treatment of motor dysfunction. Br. J. Pharmacol. 2009, 156, 1029–1040. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, F.; Capparelli, E.; Abate, C.; Colabufo, N.A.; Contino, M. Perspectives of Cannabinoid Type 2 Receptor (CB2R) Ligands in Neurodegenerative Disorders: Structure-Affinity Relationship (SAfiR) and Structure-Activity Relationship (SAR) Studies. J. Med. Chem. 2017, 60, 9913–9931. [Google Scholar] [CrossRef]
- Haider, A.; Spinelli, F.; Herde, A.M.; Mu, B.; Keller, C.; Margelisch, M.; Weber, M.; Schibli, R.; Mu, L.; Ametamey, S.M. Evaluation of 4-oxo-quinoline-based CB2 PET radioligands in R6/2 chorea huntington mouse model and human ALS spinal cord tissue. Eur. J. Med. Chem. 2018, 145, 746–759. [Google Scholar] [CrossRef]
- Sagredo, O.; González, S.; Aroyo, I.; Pazos, M.R.; Benito, C.; Lastres-Becker, I.; Romero, J.P.; Tolón, R.M.; Mechoulam, R.; Brouillet, E.; et al. Cannabinoid CB2 receptor agonists protect the striatum against malonate toxicity: Relevance for Huntington’s disease. Glia 2009, 57, 1154–1167. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, J.; Truong, J.; Bouchard, K.; Dunkelberger, D.; Desrayaud, S.; Moussaoui, S.; Tabrizi, S.J.; Stella, N.; Muchowski, P.J. Cannabinoid receptor 2 signaling in peripheral immune cells modulates disease onset and severity in mouse models of Huntington’s disease. J. Neurosci. 2012, 32, 18259–18268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazuelos, J.; Aguado, T.; Pazos, M.R.; Julien, B.; Carrasco, C.; Resel, E.; Sagredo, O.; Benito, C.; Romero, J.; Azcoitia, I.; et al. Microglial CB2 cannabinoid receptors are neuroprotective in Huntington’s disease excitotoxicity. Brain 2009, 132, 3152–3164. [Google Scholar] [CrossRef] [Green Version]
- Bejargafshe, M.J.; Hedayati, M.; Zahabiasli, S.; Tahmasbpour, E.; Rahmanzadeh, S.; Nejad-Moghaddam, A. Safety and efficacy of stem cell therapy for treatment of neural damage in patients with multiple sclerosis. Stem Cell Investig. 2019, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, E.D.; Dutra, R.C. Cannabinoid receptors as therapeutic targets for autoimmune diseases: Where do we stand? Drug Discov. Today 2019, 24, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Maresz, K.; Carrier, E.J.; Ponomarev, E.D.; Hillard, C.J.; Dittel, B.N. Modulation of the cannabinoid CB2 receptor in microglial cells in response to inflammatory stimuli. J. Neurochem. 2005, 95, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Palazuelos, J.; Davoust, N.; Julien, B.; Hatterer, E.; Aguado, T.; Mechoulam, R.; Benito, C.; Romero, J.; Silva, A.; Guzmán, M.; et al. The CB2 cannabinoid receptor controls myeloid progenitor trafficking: Involvement in the pathogenesis of an animal model of multiple sclerosis. J. Biol. Chem. 2008, 283, 13320–13324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Martin, B.R.; Adler, M.W.; Razdan, R.J.; Kong, W.; Ganea, D.; Tuma, R.F. Modulation of cannabinoid receptor activation as a neuroprotective strategy for EAE and stroke. J. Neuroimmune Pharmacol. 2009, 4, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Alberti, T.B.; Barbosa, W.L.R.; Vieira, J.L.F.; Raposo, N.R.B.; Dutra, R.C. (−)-β-caryophyllene, a CB2 receptor-selective phytocannabinoid, suppresses motor paralysis and neuroinflammation in a murine model of multiple sclerosis. Int. J. Mol. Sci. 2017, 18, 691. [Google Scholar] [CrossRef] [PubMed]
- Loría, F.; Petrosino, S.; Mestre, L.; Spagnolo, A.; Correa, F.; Hernangómez, M.; Guaza, C.; Di Marzo, V.; Docagne, F. Study of the regulation of the endocannabinoid system in a virus model of multiple sclerosis reveals a therapeutic effect of palmitoylethanolamide. Eur. J. Neurosci. 2008, 28, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Arévalo-Martín, Á.; Vela, J.M.; Molina-Holgado, E.; Borrell, J.; Guaza, C. Therapeutic action of cannabinoids in a murine model of multiple sclerosis. J. Neurosci. 2003, 23, 2511–2516. [Google Scholar] [CrossRef] [PubMed]
- Arevalo-Martin, A.; Molina-Holgado, E.; Guaza, C. A CB 1/CB 2 receptor agonist, WIN 55,212-2, exerts its therapeutic effect in a viral autoimmune model of multiple sclerosis by restoring self-tolerance to myelin. Neuropharmacology 2012, 63, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Tahamtan, A.; Rezaiy, S.; Samadizadeh, S.; Moradi, A.; Tabarraei, A.; Javid, N.; Oladnabi, M.; Naeimi, M.H. Cannabinoid CB2 Receptor Functional Variation (Q63R) Is Associated with Multiple Sclerosis in Iranian Subjects. J. Mol. Neurosci. 2020, 70, 26–31. [Google Scholar] [CrossRef]
- Sánchez López, A.J.; Román-Vega, L.; Ramil Tojeiro, E.; Giuffrida, A.; García-Merino, A. Regulation of cannabinoid receptor gene expression and endocannabinoid levels in lymphocyte subsets by interferon-β: A longitudinal study in multiple sclerosis patients. Clin. Exp. Immunol. 2015, 179, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Yiangou, Y.; Facer, P.; Durrenberger, P.; Chessell, I.P.; Naylor, A.; Bountra, C.; Banati, R.R.; Anand, P. COX-2, CB2 and P2X7-immunoreactivities are increased in activated microglial cells/macrophages of multiple sclerosis and amyotrophic lateral sclerosis spinal cord. BMC Neurol. 2006, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Benito, C.; Romero, J.P.; Tolón, R.M.; Clemente, D.; Docagne, F.; Hillard, C.J.; Guaza, C.; Romero, J. Cannabinoid CB1 and CB2 receptors and fatty acid amide hydrolase are specific markers of plaque cell subtypes in human multiple sclerosis. J. Neurosci. 2007, 27, 2396–2402. [Google Scholar] [CrossRef] [Green Version]
- Ludolph, A.C.; Meyer, T.; Riepe, M.W. The role of excitotoxicity in ALS what is the evidence? J. Neurol. 2000, 247, 7–16. [Google Scholar] [CrossRef]
- Shoemaker, J.L.; Seely, K.A.; Reed, R.L.; Crow, J.P.; Prather, P.L. The CB2 cannabinoid agonist AM-1241 prolongs survival in a transgenic mouse model of amyotrophic lateral sclerosis when initiated at symptom onset. J. Neurochem. 2007, 101, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Moore, D.H.; Makriyannis, A.; Abood, M.E. AM1241, a cannabinoid CB2 receptor selective compound, delays disease progression in a mouse model of amyotrophic lateral sclerosis. Eur. J. Pharmacol. 2006, 542, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Espejo-Porras, F.; García-Toscano, L.; Rodríguez-Cueto, C.; Santos-García, I.; de Lago, E.; Fernandez-Ruiz, J. Targeting glial cannabinoid CB2 receptors to delay the progression of the pathological phenotype in TDP-43 (A315T) transgenic mice, a model of amyotrophic lateral sclerosis. Br. J. Pharmacol. 2019, 176, 1585–1600. [Google Scholar] [CrossRef] [PubMed]
- Espejo-Porras, F.; Piscitelli, F.; Verde, R.; Ramos, J.A.; Di Marzo, V.; de Lago, E.; Fernández-Ruiz, J. Changes in the endocannabinoid signaling system in CNS structures of TDP-43 transgenic mice: Relevance for a neuroprotective therapy in TDP-43-related disorders. J. Neuroimmune Pharmacol. 2015, 10, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Martet, M.; Espejo-Porras, F.; Fernández-Ruiz, J.; De Lago, E. Changes in endocannabinoid receptors and enzymes in the spinal cord of sod1g93a transgenic mice and evaluation of a sativex®-like combination of phytocannabinoids: Interest for future therapies in amyotrophic lateral sclerosis. CNS Neurosci. Ther. 2014, 20, 809–815. [Google Scholar] [CrossRef]
- Ji, X.; Zeng, Y.; Wu, J. The CB2 receptor as a novel therapeutic target for epilepsy treatment. Int. J. Mol. Sci. 2021, 22, 8961. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, J.; Huang, S.; Zhu, W.; Wang, Y.; Chen, O.; Xue, J. Neuroprotective effects of isoliquiritigenin against cognitive impairment via suppression of synaptic dysfunction, neuronal injury, and neuroinflammation in rats with kainic acid-induced seizures. Int. Immunopharmacol. 2019, 72, 358–366. [Google Scholar] [CrossRef]
- Sugaya, Y.; Yamazaki, M.; Uchigashima, M.; Kobayashi, K.; Watanabe, M.; Sakimura, K.; Kano, M. Crucial Roles of the Endocannabinoid 2-Arachidonoylglycerol in the Suppression of Epileptic Seizures. Cell Rep. 2016, 16, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.S.; Stella, N.; Catterall, W.A.; Westenbroek, R.E. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 11229–11234. [Google Scholar] [CrossRef] [Green Version]
- Devinsky, O.; Patel, A.D.; Cross, J.H.; Villanueva, V.; Wirrell, E.C.; Privitera, M.; Greenwood, S.M.; Roberts, C.; Checketts, D.; VanLandingham, K.E.; et al. Effect of Cannabidiol on Drop Seizures in the Lennox–Gastaut Syndrome. N. Engl. J. Med. 2018, 378, 1888–1897. [Google Scholar] [CrossRef] [Green Version]
- Bialer, M.; Perucca, E. Does cannabidiol have antiseizure activity independent of its interactions with clobazam? An appraisal of the evidence from randomized controlled trials. Epilepsia 2020, 61, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Wu, J. Cannabis, cannabinoid receptors, and endocannabinoid system: Yesterday, today, and tomorrow. Acta Pharmacol. Sin. 2019, 40, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Aghaei, I.; Rostampour, M.; Shabani, M.; Naderi, N.; Motamedi, F.; Babaei, P.; Khakpour-Taleghani, B. Palmitoylethanolamide attenuates PTZ-induced seizures through CB1 and CB2 receptors. Epilepsy Res. 2015, 117, 23–28. [Google Scholar] [CrossRef] [PubMed]
- da Silva Oliveira, G.L.; da Silva, J.C.C.L.; dos Santos, C.L.; da Silva, A.P.; Feitosa, C.M.; de Castro Almeida, F.R. Anticonvulsant, Anxiolytic and Antidepressant Properties of the β-caryophyllene in Swiss Mice: Involvement of Benzodiazepine-GABAAergic, Serotonergic and Nitrergic Systems. Curr. Mol. Pharmacol. 2020, 14, 36–51. [Google Scholar] [CrossRef]
- Huizenga, M.N.; Wicker, E.; Beck, V.C.; Forcelli, P.A. Anticonvulsant effect of cannabinoid receptor agonists in models of seizures in developing rats. Epilepsia 2017, 58, 1593–1602. [Google Scholar] [CrossRef]
- de Carvalho, C.R.; Hoeller, A.A.; Franco, P.L.C.; Martini, A.P.S.; Soares, F.M.S.; Lin, K.; Prediger, R.D.; Whalley, B.J.; Walz, R. The cannabinoid CB2 receptor-specific agonist AM1241 increases pentylenetetrazole-induced seizure severity in Wistar rats. Epilepsy Res. 2016, 127, 160–167. [Google Scholar] [CrossRef]
- Shapiro, L.; Wong, J.C.; Escayg, A. Reduced cannabinoid 2 receptor activity increases susceptibility to induced seizures in mice. Epilepsia 2019, 60, 2359–2369. [Google Scholar] [CrossRef]
- Chodobski, A.; Zink, B.J.; Szmydynger-Chodobska, J. Blood-Brain Barrier Pathophysiology in Traumatic Brain Injury. Transl. Stroke Res. 2011, 2, 492–516. [Google Scholar] [CrossRef] [Green Version]
- Amenta, P.S.; Jallo, J.I.; Tuma, R.F.; Elliott, M.B. A cannabinoid type 2 receptor agonist attenuates blood-brain barrier damage and neurodegeneration in a murine model of traumatic brain injury. J. Neurosci. Res. 2012, 90, 2293–2305. [Google Scholar] [CrossRef] [PubMed]
- Elliott, M.B.; Tuma, R.F.; Amenta, P.S.; Barbe, M.F.; Jallo, J.I. Acute effects of a selective cannabinoid-2 receptor agonist on neuroinflammation in a model of traumatic brain injury. J. Neurotrauma 2011, 28, 973–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magid, L.; Heymann, S.; Elgali, M.; Avram, L.; Cohen, Y.; Liraz-Zaltsman, S.; Mechoulam, R.; Shohami, E. Role of CB2 Receptor in the Recovery of Mice after Traumatic Brain Injury. J. Neurotrauma 2019, 36, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Luo, Q.; Shang, B.; Yang, X.; Zhang, Y.; Pan, Q.; Wu, N.; Tang, W.; Du, D.; Sun, X.; et al. Selective activation of cannabinoid receptor-2 reduces white matter injury via PERK signaling in a rat model of traumatic brain injury. Exp. Neurol. 2022, 347, 113899. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hua, T.; Vemuri, K.; Ho, J.H.; Wu, Y.; Wu, L.; Popov, P.; Benchama, O.; Zvonok, N.; Locke, K.; et al. Crystal Structure of the Human Cannabinoid Receptor CB2. Cell 2019, 176, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kibret, B.G.; Ishiguro, H.; Horiuchi, Y.; Onaivi, E.S. New Insights and Potential Therapeutic Targeting of CB2 Cannabinoid Receptors in CNS Disorders. Int. J. Mol. Sci. 2022, 23, 975. https://doi.org/10.3390/ijms23020975
Kibret BG, Ishiguro H, Horiuchi Y, Onaivi ES. New Insights and Potential Therapeutic Targeting of CB2 Cannabinoid Receptors in CNS Disorders. International Journal of Molecular Sciences. 2022; 23(2):975. https://doi.org/10.3390/ijms23020975
Chicago/Turabian StyleKibret, Berhanu Geresu, Hiroki Ishiguro, Yasue Horiuchi, and Emmanuel S. Onaivi. 2022. "New Insights and Potential Therapeutic Targeting of CB2 Cannabinoid Receptors in CNS Disorders" International Journal of Molecular Sciences 23, no. 2: 975. https://doi.org/10.3390/ijms23020975
APA StyleKibret, B. G., Ishiguro, H., Horiuchi, Y., & Onaivi, E. S. (2022). New Insights and Potential Therapeutic Targeting of CB2 Cannabinoid Receptors in CNS Disorders. International Journal of Molecular Sciences, 23(2), 975. https://doi.org/10.3390/ijms23020975