Abstract
Cancer-associated fibroblasts (CAFs) are an essential component of the tumour microenvironment. They represent a heterogeneous group of cells that are under the control of cancer cells and can reversely influence the cancer cell population. They affect the cancer cell differentiation status, and the migration and formation of metastases. This is achieved through the production of the extracellular matrix and numerous bioactive factors. IL-6 seems to play the central role in the communication of noncancerous and cancer cells in the tumour. This review outlines the role of exosomes in cancer cells and cancer-associated fibroblasts. Available data on the exosomal cargo, which can significantly intensify interactions in the tumour, are summarised. The role of exosomes as mediators of the dialogue between cancer cells and cancer-associated fibroblasts is discussed together with their therapeutic relevance. The functional unity of the paracrine- and exosome-mediated communication of cancer cells with the tumour microenvironment represented by CAFs is worthy of attention.
1. Introduction
Cancer ranks as a leading cause of death in many developed countries worldwide [1]. The growing burden of cancer incidence and mortality notably reflects the ageing of the population and changes in cancer risk factors. In positive or negative ways, many of these factors are associated with socioeconomic development. Importantly, cancer ranks as a leading barrier to increasing life expectancy in every country, according to the WHO [2].
Malignant tumours are highly complex tissues. In various ways, tumour morphogenesis is driven by mechanisms that are similar to those regulating the development of organs and tissues under normal physiological conditions [3]. To acquire a normal 3D structure, organs and tissues require a precise system of regulatory mechanisms. The normal architecture consequently contributes greatly to their correct function and maintenance in the long term. Therefore, structural and functional integrity is a critical prerequisite for the health of human body over its lifespan.
The process of malignant transformation is associated with an error in genetic information. This error may result from heredity or may be acquired during postnatal life. These acquired mutations may occur stochastically, by exposure to chemical/physical mutagens, due to viral infection, etc. Successful cancerogenesis usually requires many consecutively acquired mutations. Their conservation in the human genome may be facilitated by the failure of DNA repair mechanisms. At present, there are no doubts regarding the primary and critical role of these genetic factors in cancerogenesis. Cancer may arise from a single cell being transformed in a multi-step process.
Nevertheless, the tumour is not formed exclusively by these mutated cells. Moreover, the popular view of completely deregulated cancer growth seems to be somewhat naïve. Besides genetic defects, the malignant cell must be observed in and characterised by the context of the tissue environment.
There are numerous noncancerous cells located in the vicinity of mutated malignant cells, collectively forming the tumours. This non-malignant component in tumours is most frequently referred to as tumour stroma. Of note, the non-mutated genetic status of stromal cells does not necessarily imply their full functional normality. In recent years, it has become a widely accepted concept by many cancer biologists that good cells may become cancerous because of their exposure to a bad environment [4]. This notion was preceded by classical studies on the importance of senescence in the course of cancerogenesis [5,6]. Later, this concept of a dangerous neighbourhood was extended into clinical oncology with therapeutic implications for the cell interactome [7]. Conversely, the importance and power of a normal environment was depicted in seminal works on xenotransplanted teratomas [8], even in the 1970s. Surprisingly, the microenvironment was able to completely overdrive the malignant behaviour of teratoma cells, leading to their incorporation into normal structures of the embryo without signs of malignancy.
Therefore, the tissue microenvironment requires closer attention. Several principal stromal cell populations form the cancer cell niche. Namely, cancer-associated fibroblasts (CAFs) and various types of immune cells (e.g., cancer-associated macrophages) significantly influence the biological properties of cancerous cells. This may result in the maintenance of the low differentiation status of cancer stem cells, the increased proliferative capacity of cancer cells, and consequently their increased migratory capacity. These advantages contribute to clinically significant phenomena, such as the formation of lymphatic/distant metastases [9]. Cancer cells, in collaboration with their noncancerous counterparts in tumours, even influence the metabolic status through the production of biologically active factors with impact on the liver, striated muscle, and white fat that initiate wasting, resulting in the death of cancer patients [10].
These stromal components of tumours are the result of complex interactions across the tumour niche. With a certain level of hyperbole, the tumour can be seen as a selfish organ [11]. While normal organs execute their physiological functions, thus supporting the survival of the whole organism, the immature and poorly developed tumour expands selfishly at the expense of the organism. However, even at a rather primitive stage of their development, tumours still require vital regulations. Without tight stromal regulatory mechanisms, rapidly proliferating clones may very easily find themselves in a disastrous situation. Uncontrolled attempts to proliferate in the absence of proper metabolic resources could end in cell death. In a broader view, this scenario may remind us of the ecological phenomena of population extinction.
To conclude, the inclusion of population biology and community ecology can be inspiring for cancer biologists. Thinking of tumours as organs composed of a plethora of mutually interacting populations may allow us to better understand the processes governing cancer development and progress.
2. Fibroblasts Participate in Organ Morphogenesis
Fibroblasts participate in the 3D architecture formation of virtually all organs in the human body. Fibroblasts also significantly influence their correct functions. Speaking broadly, fibroblasts are the versatile spindle-shaped cells of mesenchymal origin that are primarily capable of both extracellular matrix (ECM) synthesis and ECM resorption via released proteolytic enzymes. Besides the production and maintenance of ECM and related molecules, fibroblasts are potent producers of various small signalling soluble molecules, e.g., cytokines and chemokines. However, fibroblast morphology in various tissues seems to be relatively uniform; there is an apparent positional memory of fibroblasts. This is governed by the genetic imprinting of the homeobox (HOX) family of transcription factors [12]. Similar fusiform morphology thus does not imply identical functional properties in fibroblasts of various origins.
The importance of fibroblast functions in tissues can be clearly evidenced in embryonic development. Surprisingly, fibroblasts act here quite uniformly in different tissues during their development. Stereotypically, the organ primordium, the epithelial bud, is surrounded by condensed mesenchymal cells in early development. The epithelium is usually separated from the mesenchyme by a thin sheath of connective tissue—the so-called basement membrane. This membrane excludes the direct physical contact of some populations; however, this membrane is permeable for small soluble molecules. The extensive exchange of molecules between the epithelium and mesenchyme across the basement membrane is essential for the successful development of the organ primordium. For example, this stereotypical model may be applied to the development of skin appendages, hairs, mammary gland, and also teeth [13,14,15]. Moreover, the early development of visceral organs, such as the lung [16], liver [17], and exocrine pancreas [18,19], follows a very similar morphogenetic scenario. As summarised in these review articles, the exchange of molecules, such as members of the BMP and FGF growth factor families, between the epithelium and mesenchyme is indispensable. Further, other factors, such as members of the TGF-β, IGF, and Wnt family ligands, as well as EGF and HGF, must also be highlighted in this context. The interaction of these factors via the appropriate membrane-bound receptors initiates the expression of specific transcription factors that control the genetic programme of the particular cell type differentiation, finally leading to organ development.
Relevant to the skin structure, several populations of fibroblast lineages have been identified in the last few years; mouse models proved to be particularly useful for that [20,21]. Papillary and reticular fibroblasts show different morphologies and molecular markers in vivo and in vitro, and they can also differ functionally. A very prominent fibroblast population is also found in the dermal papilla of hair follicles [22]. There is increasing evidence that deeply located fibroblasts present at the border with adipose subcutis also represent a very different subset of dermal fibroblasts [23]. Single-cell sequencing methods proved to be very useful for unravelling cell heterogeneity. However, the single-cell analysis-based literature on human skin remains relatively scarce [24,25]. Moreover, these transcriptomics-identified markers may not be easy to link with the fibroblast subpopulation markers necessary in diagnostic pathology. Additionally, the extrapolation of mouse data to humans is not straightforward because murine and human skin differ significantly across many morphological and functional features.
3. Fibroblasts as a Fundamental Bioactive Part of the Majority of Tumour Types
Tumours represent complex multicellular tissue/organ-like structures. The tumour cell type composition is known from histological research. Moreover, intercellular interactions could be easily observed and described using the established terms and principles of ecology. This approach facilitates a better understanding of the relation between the cancerous and noncancerous cell populations in primary tumours.
Intercellular interactions of various populations in the tumour also influence the spreading of cancer cells and the formation of metastases [26]. Even a highly malignant cell requires a safe harbour [27]. While the process of tumour formation and progression is relatively slow, it is possible for cancer cells to gradually gain control and rearrange the components of the tumour microenvironment through mutual and dynamic communication [28]. However, the migrating or circulating cancer cells require vital support to colonise their new niche and form metastases. It is very likely that the rate of success of these individual malignant cells at forming a new metastasis is remarkably low [29]. It is a well-fitting cellular embodiment of the classical seed and soil theory coined by Stephen Paget [30]. However, the soil does not passively accept the seeds. The failure of individual cells here is not due to the lack of their malignant features, but it is likely attributed to their meeting a non-permissive environment populated by non-cooperative cell types. With clinical relevance, most of the deaths in oncology may be attributed to metastatic disease [31].
The successful development of cancer in a patient’s body is, thus, in terms of Darwinian biology, a result of the co-evolution of the cancerous and noncancerous cells of the tumour. It has been well documented in many types of cancer that multiple steps of carcinogenesis leading to the cumulative effect of consequently acquired mutations (e.g., loss of key tumour suppressor genes and the overexpression of important oncogenes) in an individual cell will cause a normal cell to become a cancerous cell [32]. This ongoing evolution influences the fibroblasts in their vicinity. In this light, fibroblasts that positively influence the growth of cancer cells and their migration are of primary importance [33]. Consequently, fibroblasts modify the local immune response and thus stimulate the cancer-supporting function of the immune cells infiltrating the tumour site [34,35,36].
There is limited empirical data available on this interaction during the tumour progression. The theoretical literature in evolutionary studies usually highlights that interaction-dependent population size changes are likely the key to the full understanding of these co-evolutionary dynamics [37]. Thus, a more realistic view of the complex nature of tumour–stromal interactions is urgently needed.
It is likely assumption that the quantity of stromal cells is cancer type specific. It is well known that the proportion of CAFs and other stromal cells differs according to the type of solid tumour [38]. In some types of cancer, CAFs are numerous and extensively produce the extracellular matrix. This is a prominent feature in, e.g., highly malignant ductal adenocarcinoma of the pancreas, referred to as the desmoplastic type of tumour [39,40]. On the other hand, the proportion of stromal cells is usually somewhat modest in cutaneous malignant melanoma [34] or glioblastoma [41]. Despite their sparse quantity, the CAFs observed in these tumours are highly biologically active. Our knowledge remains sparse for other types of solid malignancies of non-epithelial origin, e.g., in sarcomas. Except for Ewing sarcoma [42] and osteosarcoma [43], the role of CAFs in sarcomas of different origins is unclear. Further, CAFs have not been extensively studied in haematological malignancies so far. However, there is evidence that cells with the properties of CAFs have been identified in both acute and chronic lymphocytic leukaemia, Hodgkin and follicular lymphoma, and multiple myeloma [44].
3.1. Origin of CAFs
The fibroblast populations in the vast majority of human body structures originate in the mesoderm germ layer. However, fibroblasts of the facial part of the head and part of the neck are raised from the neural crest; therefore, they are also referred to as ectomesenchyme [45,46]. Interestingly, there are significant differences in transcriptomic profiles between mesoderm-originated and ectomesenchyme-originated fibroblasts. This significant distinction is preserved even in adult life, as evidenced at the whole-genome scale [47].
As mentioned earlier, the cancer cell population undergoes the progressive multistage genetic alteration necessary to acquire fully malignant features. However, the CAFs are not primarily driven by acquired mutations. It is a vitally important task for proliferating malignant clones to initiate communication with the surrounding tissue in a timely manner. Without a permissive or even supportive environment, the clone will very likely not survive. Via this dialogue [48], cancer cells are able to recruit or even corrupt [49] the various cells around them, including tissue fibroblasts. In recent years, it has become widely recognised that epigenetic alterations are essential for regulating CAF bioactivity [50]. This local recruitment is the most widely recognised theory of the origin of CAFs, and it was also suggested as a potential target for therapeutic interventions in cancer [51].
However, CAFs may theoretically be descendants of cell types other than local fibroblasts. The epithelial–mesenchymal transition (EMT) represents a critically important developmental mechanism [52]. However, it is also one of the leading mechanisms in tumour spreading; it is necessary to initiate metastasis. Notably, epithelial cells undergoing EMT newly acquire the mesenchymal phenotype. This mesenchymal state is also associated with the capacity of cells to migrate. Therefore, it is a very attractive hypothesis that CAFs may originate from malignant cells via EMT [53,54]. This would represent a significant advantage for malignant cells, because the newly formed tumour would become self-sufficient at producing supportive stromal structures. This hypothesis is mainly supported by the observation of mutations in CAFs. However, should CAFs really originate from the tumour cell population, there would have to be a significant overlap in their mutational repertoire. As opposed by others, mutations in CAFs can also be explained by the broad-field cancerisation of mutagenic agents affecting both cancer cells and the surrounding mesenchyme [55,56]. In our own experience, we have not observed the concordant mutation of, e.g., BRAF V600E in melanomas and in isolated CAFs [57]. In other experiments, human cancer cells grafted to immunocompromised mice developed tumours, while the stroma was of host origin, as confirmed by a species-specific antibody recognising the human and not murine vimentin [58]. Based on these summarised data, the formation of CAFs from cancer cells via EMT in vivo is likely not a principal mechanism.
Other candidate cell types hypothesised as precursors of CAFs and factors stimulating their formation [59,60,61,62,63,64,65] are included in Table 1. Members of the TGF-β family seem to be the most frequently discussed in this context as inducers of the transition of precursor cells to myofibroblasts [66].

Table 1.
Cell types as potential precursors for CAF formation.
Significantly, a great deal of attention in the last few years has also been dedicated to mesenchymal stem cells (MSCs) and their ability to migrate to cancer sites. Besides the many unique properties of MSCs, these lowly differentiated cells can also differentiate to CAFs and gain their full properties. This behaviour, however, also substantiated the proposal of a new therapeutic strategy based on the loading of MSCs with oncolytic viruses to destroy cancer cells [67,68].
3.2. Markers of CAFs
Diagnostic pathology remains a vitally important discipline in any biomedical research, including on cancer. Immunohistochemical and molecular analysis further enhances our insight into the frequently subtle changes modifying the disease progression. Unfortunately, no specific marker reliably distinguishing CAFs from other fibroblasts has been discovered so far. The frequently mentioned notorious marker of tumour stroma expressed in CAFs is α-smooth muscle actin (SMA). However, not all of the fibroblasts in the tumour stroma in vivo or fibroblasts isolated from tumour stroma in vitro express this protein (Figure 1).
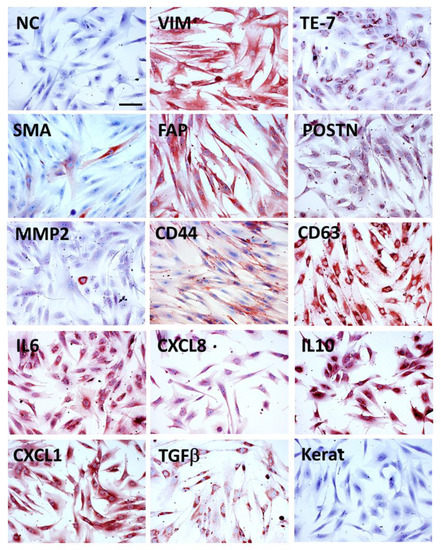
Figure 1.
CAFs isolated from human cutaneous basal cell carcinoma (immunocytochemical analysis, red AEC product indicates positive reaction, NC—negative control with isotype antibody, bar represents 100 μm). CAFs are expressing intermediate filament protein vimentin (VIM), and specific mesenchymal marker TE-7. SMA was detected only in a minority of cells, while FAP was present universally, and periostin (POSTN) was also found in virtually all cells. MMP2 was positive only in sporadic cells. CD44 (hyaluronic acid receptor) was widely expressed with marked positivity on the periphery of cells. Tetraspanin CD63 is associated with the membranes of intracellular vesicles and was detected in all cells as a distinct granular signal. CAFs produce high quantities of interleukins and chemokines, such as IL-6, CXCL-8, IL-10, CXCL-1, and TGF-β. Despite using pan-keratin reactive antibodies, no keratin positivity was observed in CAFs.
However, SMA expression is not a sufficient marker for mature myofibroblast identification [69]. The cytoskeletal distribution of SMA and the formation of stress fibres should also be considered. In fully mature myofibroblasts, the neo-expression of SMA-positive stress fibres is present. The absence of SMA within these fibres facilitates the distinction of proto-myofibroblasts from mature myofibroblasts. However, the significance of these steps of myofibroblast maturation and their role in tumour biology remains unclear. It has been suggested that the inhibition of SMA polymerisation could also have some therapeutic prospects [70].
For the sake of practicality, SMA expression across different types of tumours seems to be quite universal. However, the extent of the myofibroblastic population observed in various tumours varies greatly (Figure 2).
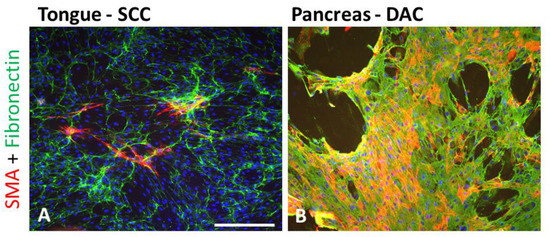
Figure 2.
CAFs isolated from squamous cell carcinoma (SCC) of the tongue (A) and ductal adenocarcinoma (DAC) (B) of the pancreas (desmoplastic form) express SMA (red) and produce fibronectin (green). Bar represents 100 μm.
Therefore, the identification of additional protein markers reflecting the activation of CAFs or their different origin may be helpful for recognising CAFs in diagnostic tissue sections or in cell culture [71,72,73,74]. These markers are listed in Table 2.
Table 2.
Markers of CAFs according to their origin.
Some of these markers, such as FAP, can be potentially employed in theranostic applications as a diagnostic and therapeutic tool [75,76].
3.3. Heterogeneity of CAFs
Recent progress in molecular biology methods, such as single-cell RNA sequencing, have facilitated deeper insights into tissue population heterogeneity. Using this powerful tool for distinct tumours and experimental situations, the subpopulations in cancer stroma, including CAFs, can be analysed in detail. Dermal fibroblasts cultured in 3D heterogeneous spheroids with melanoma cells demonstrated their ability to gain CAF-like properties. Notably, sun-damaged fibroblasts from aged donors transformed more readily than juvenile control fibroblasts [25]. Even in this simplistic 3D model, fibroblasts represent a heterogeneous population. One part of the CAFs predominantly produced the extracellular matrix (ECM). This is primarily consistent with the expected fibroblast role in the dermis. In contrast, another subpopulation predominantly produced inflammatory mediators. These factors, namely IL-6 and CXCL-8, are, to a great extent, produced in fibroblasts, with only a negligible quantity of ECM. Another subpopulation of fibroblasts observed in this model was driven by the influence of TGF-β signalling [25]. Similarly, other authors have demonstrated the heterogeneity of CAFs in clinical samples from different tumours or in animal experiments [75].
It was suggested that CAFs could be further subclassified into (a) immune CAFs that produce inflammatory mediators, (b) desmoplastic CAFs that secrete ECM, (c) contractile CAFs that have a high expression of SMA, and finally (d) aggressive CAFs that occur in aggressive tumours [76,77,78]. Notably, TGF-β signalling can drive several of these phenotypes, e.g., contractile CAFs and desmoplastic CAFs. Interestingly, the comparison of CAF heterogeneity demonstrated some similarities across different types of tumour [79,80,81]. This confirms earlier observations of the functional similarities among CAFs of various origins in vitro [82]. Further, it suggests that the transcriptional profile of CAFs can influence the biological properties of cancer in general, and it is worthy of attention and potentially also targeting. Based on these observations, it seems likely that a detailed analysis of CAF subtypes can be employed to refine diagnostics in the future. The practicality of this concept was tested in patients suffering from aggressive adenocarcinoma of the pancreas [83].
3.4. Role of CAFs in Cancer Biology
The cancer microenvironment, including CAFs, is to some extent similar to the site of a healing wound. SMA-expressing myofibroblasts are responsible for wound contraction and the production of bioactive factors that facilitate tissue integrity recovery [9,84,85]. The role of CAFs is quite similar to the role of myofibroblasts in the wound, but their functional activity is not time restricted [86]. It is known that wound healing and cancer share many important signalling pathways [87] and differ in temporal regulation. Sadly, we lack the proper understanding of the mechanism representing the master switch in CAFs.
It is known that CAFs can influence the differentiation status of cancer cells and the normal epithelium. Using CAFs in various in vitro models, it was demonstrated earlier that normal epithelial cells express markers characteristic of the immature cells of low differentiation status [88,89]. Moreover, when CAFs were co-cultured with other fibroblasts, these fibroblasts underwent phenotypic changes, and the differentiation potential of exposed fibroblasts was consequently comparable to mesenchymal stem cells [86].
Low differentiation status maintenance may be a critical advantage for tumour cells during, e.g., chemotherapy. The stem cell populations, including cancer stem cells, use several molecular transport mechanisms, causing their resistance to xenobiotics, including chemotherapy. Therefore, the maintenance of stemness may be a major cause of treatment failure. Besides that, the tumour microenvironment forms a safe harbour for cancer cells. The desmoplastic tumour niche may physically represent a site where cancer cells are exposed only to the tolerable doses of anticancer drugs [90].
Mediators of the intercellular communication between cancer cells and noncancerous cells, including CAFs, can also leak from the primary tumour site. Thus, CAFs can participate in the formation of premetastatic niches in distant body parts [10,57,91]. CAFs also significantly influence the migration of cancer cells, such as melanoma and glioblastoma cells, in vitro [92,93], resulting in a consequently increasing probability of colonising the new permissive site.
With an emphasis on the systemic clinical effect, elevated levels of these mediators are responsible for cancer patient wasting and preterminal cachexia [10,92,93]. As the activity of CAFs seems to be quite conserved, despite their origin and type of cancer, this seems to be worthy of targeting and therapeutic intervention.
3.4.1. Production and Remodelling of ECM
In the most classical view, fibroblasts are responsible for the maintenance of the structural integrity of connective tissues. Similar to other types of fibroblasts, CAFs also produce molecules of ECM, such as collagen, hyaluronic acid, fibronectin, periostin, and tenascin [94,95,96,97,98,99,100,101]. ECM production by CAFs seems to be more extensive than in normal fibroblasts, and it even reflects the severity of the disease.
On the other hand, stromal cells, including CAFs, also produce proteases [102,103]. This complexity of dynamic ECM production and degradation seems to be essential for tissue healing, but it is crucial for tumour spreading and metastasis formation as well [104,105,106]. In tissue samples, CAFs are usually observed at the leading edge of collectively migrating cancer cells, where they prepare a corridor, facilitating cancer cell migration and consequent metastatic spread [107].
3.4.2. Production of Bioactive Factors with Emphasis on IL-6
CAFs produce numerous bioactive factors, which have been reviewed extensively in numerous articles [10,108,109,110,111,112]. When comparing different tumour types, IL-6 seems to be widely expressed by CAFs in many, if not in virtually all, cancer types [51,113]. As shown in these articles, IL-6 participates in the control of the low level of differentiation of cancer cells and the regulation of their migratory activity. The systemic effect of IL-6 leads to the wasting of cancer patients. Further, after passing through the blood–brain barrier, IL-6 may have behavioural effects and induce depression [114].
As was demonstrated in vitro, the production of IL-6 by CAFs is precisely controlled in the co-culture of cell lines from the ductal cancer of the pancreas and CAFs isolated from the same cancer type [39]. However, IL-6 should also be seen as a principal component of the senescence-associated secretory phenotype in many cells [115]. Indeed, IL-6 production is tightly associated with ageing, and normal dermal fibroblasts from aged persons are thus functionally quite close to CAFs [116]. A very similar observation was reported in the heterogeneous spheroids prepared from melanoma cells and juvenile/adult dermal fibroblasts [25].
To elicit a cellular response, IL-6 must bind to and be recognised by a receptor [117]. Mechanistically, IL-6 is bound to IL-6R and associated with a signal transducer called glycoprotein GP-130. IL-6R is docked to the cell membrane; however, it can be proteolytically cleaved from the membrane and, thus, it becomes soluble (sIL-6R) and can circulate in body fluids. Both types of IL-6 receptors require cooperation with glycoprotein gp130 to initiate signalling via the phosphorylation of other interacting molecules. The effects of the membrane-bound and soluble IL-6R forms are not identical [118,119,120], and it is necessary to distinguish classical versus trans-signalling effects. Therefore, the action of IL-6 is understood to be context-dependent and highly complex.
The inhibition of IL-6 signalling via receptor interference can be achieved by, e.g., monoclonal antibodies. In clinics, tocilizumab was proven to be a highly effective therapy in rheumatoid arthritis; however, it failed to be sufficient as a cancer therapy [119]. Nevertheless, current cancer therapy is frequently based on combinations, and the simultaneous targeting of several mechanisms can be beneficial and affordable. Therefore, the targeting of IL-6 along with the targeting of other molecules seems to be worthy of attention [111,113,120]. We can hypothesise as to which factors should be hit along with IL-6. It was suggested elsewhere that the targeting of IL-6 and CXCL-8 could be promising [121]. More recently, Ham and co-workers [122] confirmed the prominent role of IL-6 and TGF-β in tumours of the gastrointestinal tract, where these two factors induce resistance to chemotherapy and actinotherapy.
4. Tumour Cell-Derived Exosomes in Cancer Biology
4.1. Definition of Exosome
Exosomes are small vesicles of 30–100 nm in diameter formed by a double lipid bilayer with the presence of several proteins, including tetraspanins—namely, CD9, CD63, and CD81 (Figure 3). Exosomes are released from cells via multivesicular body production. Their cargo can potentially be presented by almost 41,000 proteins, 5000 mRNA, and 3000 microRNAs (miRNAs). These data are continually updated in the exosome protein, RNA and lipid database (http://exocarta.org/; accessed 20 December 2021). Exosomes thus represent a powerful tool for the vertical transfer of information in a highly concentrated form [123,124,125].
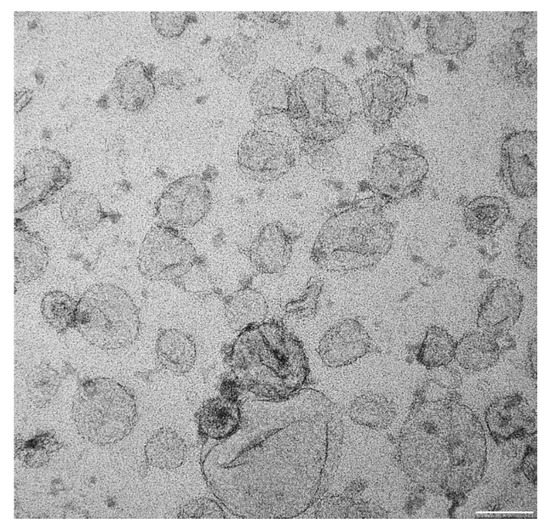
Figure 3.
Air-dried exosomes prepared from G361 melanoma cells cultured in exosome-depleted calf serum. Exosomes were adsorbed on a formvar membrane, fixed with 1% glutaraldehyde and contrasted by uranyl acetate, and detected using an electron microscope. The bar is 50 nm.
4.2. Effect of Cancer Cell-Derived Exosomes (CaExos) on the Function of CAFs
CaExos participate in the formation of the tumour microenvironment. In the broader sense, CaExos can facilitate the formation of the premetastatic niche in different types of tumours, including melanoma [126,127,128]. Exosomes can modulate various cells to prepare the microenvironment for cancer cell docking [129]. The majority of studies addressed questions regarding the role of exosomes in the mutual communication between cancer cells. However, the effect of CaExos on CAFs has not been so extensively elucidated. Indeed, CAFs can bind and internalise exosomes (Figure 4) and be subject to regulation by cancer cells.
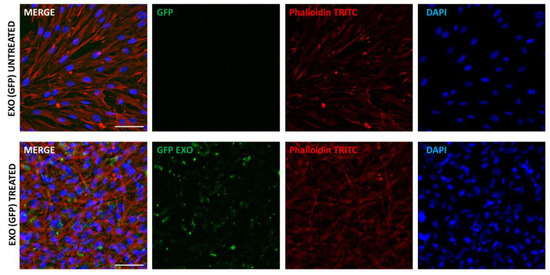
Figure 4.
CAFs from cutaneous basal cell carcinoma were treated by commercially available GFP-tagged exosomes for 12 h. Cells were consequently fixed by paraformaldehyde, permeabilised, and counterstained using phalloidin-TRITC (to show actin in the cytoplasm) and DAPI (to show DNA in the nuclei). Imaging was performed with a Leica Thunder Imager using the computational clearing technique to efficiently differentiate between the signal and background to visualise the distribution of GFP-tagged exosomes in CAFs. Bar represents 100 μm.
Melanoma-derived CaExos negatively influence the adhesion and proliferation of both the normal dermal fibroblasts and melanoma CAFs in a dose-dependent manner [130]. This finding reflects well the secretory senescent phenotype of the CAFs observed earlier [131,132]. This cellular phenotype is associated with the upregulation of miR-335 [133]. On the other hand, the in vitro invasiveness of both tested types of fibroblasts, i.e., normal dermal and CAFs, is stimulated in 3D collagen gels by the melanoma-derived CaExos. This may be understood to be a model of the invasive front mentioned earlier. Further, CaExos (melanoma) also stimulated the migration of melanoma cells from heterogeneous spheroids prepared from the melanoma cells and melanoma-isolated CAFs (Figure 5) in collagen gels [130].

Figure 5.
Melanoma cells migrate from the heterogeneous spheroids prepared from melanoma CAFs and melanoma cells (A,B). Melanoma-derived CaExo treatment facilitated migration (B). The blue circle demonstrates the position of the original spheroid, and the red circle highlights the leading edge of migration. To follow only the influence of melanoma-derived exosomes, the 3D collagen gel and culture medium contain only exosome-depleted serum. Bar represents 1 mm.
The expression profiles of normal dermal fibroblasts and melanoma-derived CAFs significantly differ after exposure to melanoma-derived CaExos. CAFs increase the expression of inflammation-promoting factors, including IL-1, IL-6, and CXCL-8, after CaExo treatment at the mRNA level, and a similar trend was also confirmed at the protein level. The normal dermal fibroblasts are also activated by CaExos, but their expression profile differs. In detail, normal dermal fibroblasts exposed to CaExo treatment mainly increased the expression of thrombospondin-1 at the protein level—this factor is important for tissue healing. Importantly, changes in the expression of IL-6 and CXCL-8 were insignificant at the protein level, although there were changes at the mRNA level [130]. The participation of melanoma-derived CaExos in the inflammation-supporting microenvironment was also observed by Lahav and co-workers [134]. The effects of CaExos on normal fibroblasts or mesenchymal stem cells as their precursors were also described in other model situations where normal cells can consequently acquire the properties of CAFs [135,136,137,138,139,140,141,142,143]. CaExos are also able to stimulate the endothelial cell–mesenchymal transition and the formation of CAFs from these precursors [144]. This activity seems to depend on the exosome cargo, mainly on miRNAs (Table 3).
Table 3.
Example of CaExo cargo molecules activating fibroblasts/mesenchymal stem cells/CAFs.
4.3. CAFs Produce Exosomes That Influence the Function of Cancer Cells
Cancer cells produce exosomes that significantly activate CAFs. A certain reciprocity can be easily expected here, and activated CAFs produce exosomes with a very significant impact on cancer cells. The effect of exosomes is enhanced by the low pH and hypoxia—these features are typical of the majority of solid tumours, independently of their origin [145,146,147]. The exosomes produced by CAFs notably influence the properties of cancer cells via a broad panel of mechanisms. This fosters stem cell-like properties in cancer cells, maintains their low differentiation status, and regulates their proliferation and migration. This is a base for the formation of metastases and resistance to chemo/actinotherapy. CAF-derived exosome efficiency mainly depends on the specific miRNAs in their cargo [73,122,136,148,149,150,151,152,153,154,155,156,157,158,159,160] (summarised in Table 4).
Table 4.
Effect of CAF-derived exosomes on cancer (selected examples).
4.4. Systemic Effects of Exosomes—Role in Cancer Wasting and Cachexia
Cancer wasting and cachexia are present in many patients in the terminal phase of malignant disease [161]. The terminal progression of the disease is associated with the production of cytokines, such as IL-6 and TNF-α. These mediators are produced by the cells of the cancer ecosystem [113] and leak to the circulating body fluids. Cancer cachexia is therefore characterised by systemic inflammation. Exosomes can also significantly participate in this deadly complication [162,163], mainly through their effect on the production of IL-6 [130]. However, non-coding RNAs seem to be critically important in the regulation of the metabolic changes leading to cachexia [163]. Mechanistically, skeletal muscle wasting as part of tumour-related cachexia in an animal model is associated with the presence of miR-125b-1-3P in the exosomes [164]. Similarly, structural and functional changes of white adipose tissue were observed in cachectic patients. This browning of adipose tissue may be linked to the effect of miR-146b-5p carried in exosomes [165]. This insidious syndrome dramatically impacts the quality of the remainder of the patient’s life, but is also associated with poor responses to therapy and decreased survival [166].
5. Conclusions
Tumours involve a complex cellular ecosystem with a multitude of intercellular interactions ongoing simultaneously. Malignant cells represent the principal population of tumours; however, stromal components forming the microenvironment cannot be neglected. Among many others, CAFs are one of the most common noncancerous cell types involved in forming tumours. CAFs are not homogeneous, and several functional subtypes can be distinguished in this cell population of mesenchymal origin. Their function is influenced by the cancer cells; however, CAFs also reversely influence the malignant population in many ways. CAFs continuously shape the landscape of the tumour microenvironment. Structurally, CAFs, as with any typical fibroblasts, produce and remodel ECM. However, their important function also lies in the paracrine production of a wide scale of bioactive factors. Significantly, CAFs produce molecules supporting the local inflammatory milieu that seems to be indispensable for cancer cell proliferation and invasiveness.
Moreover, the inflammatory factors can leak from the primary tumour site via vessels and can also have a systemic effect. This can contribute to the formation of a premetastatic niche in distant tissues, which is critically important for successful metastatic spread. Concerning the whole body of the cancer patient, these factors also actively participate in the induction of cancer wasting and cachexia, which affects the patient’s treatment and decreases survival rates.
Exosomes represent a versatile tool for intercellular communication. CaExos facilitate the transition of precursor cells to fully cancer-promoting CAFs. CaExos significantly activate CAFs via the transmitted cargo or via miRNAs and proteins, such as TGF-β. Further, CaExos stimulate CAFs to produce a broad array of pro-inflammatory factors, e.g., IL-6 and CXCL-8. Conversely, exosomes produced by CAFs also have a distinct effect on cancer cells, which is mediated by the action of a wide panel of miRNAs in the exosomal cargo. Collectively, these miRNAs directly affect the important functions of cancer cells, such as differentiation, proliferation, and migration (Figure 6). These miRNAs also consequently induce resistance to chemotherapy and actinotherapy. This mechanism contributes to the failure of oncological therapy. Moreover, these miRNAs directly impact the metabolism of muscle fibres and adipocytes and, thus, adversely modify the terminal course of malignant disease: cancer wasting and cachexia.
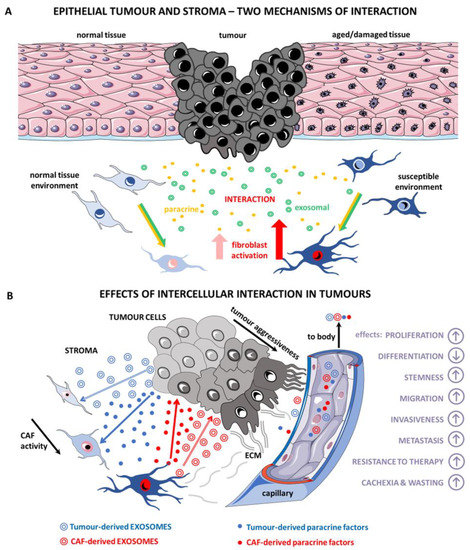
Figure 6.
Cancer cells interact with stromal CAFs via the paracrine secretion of bioactive substances, such as cytokines/growth factors, and also release exosomes. This leads to the formation of the permissive microenvironment necessary for tumour progression. The response of normal fibroblasts and aged/damaged fibroblasts to this stimulation is similar, but not identical (A) Tumour cells and CAFs use both modes of interaction (exosomal and paracrine) mutually in communication across the landscape of the tumour. These stimuli influence the biological properties of cancer cells, including the formation of metastases, and even exhibit systemic effects, such as wasting and cachexia (B) Graphics based on free Servier Medical Art image bank available at smart.servier.com. Servier Medical Art is licensed under a Creative Commons Attribution 3.0 Unported License.
This review demonstrates the functional unity of a paracrine and exosome-mediated dialogue between the cancer cells and the microenvironment represented by CAFs in the context of a tumour. In many clinical trials, the therapeutic intervention is based on monotherapy or targeting a single bioactive factor. Relevant to, e.g., IL-6 or its receptor, this therapeutic targeting was not as successful as expected. Perhaps targeting both subsystems of intercellular communication inside the cancer ecosystem, paracrine and exosome-mediated communications, simultaneously could be more fruitful.
Author Contributions
Conceptualisation, K.S.J.; preparation, K.S.J., J.B., L.L., M.K., M.V. and Y.Z.N.; writing—review and editing, artwork K.S.J. and L.L.; funding acquisition, K.S.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Grant Agency of the Czech Republic, project No. 19-05048S, by the Ministry of Education, Youth and Sports of the Czech Republic, project “Centre for Tumour Ecology—Research of the Cancer Microenvironment Supporting Cancer Growth and Spread”, No. CZ.02.1.01/0.0/0.0/16_019/0000785.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available in authors’ archive.
Acknowledgments
We acknowledge the Electron Microscopy Core Facility, Institute of Molecular Genetics, Academy of Sciences of the Czech Republic, Prague, Czech Republic for support with obtaining electron microscopic pictures of exosomes. The authors are grateful to Šárka Takáčová for English and grammar revision. We also acknowledge the long-term generous support of League Against Cancer (LPR Praha).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ASC-1 | Neutral amino acid transporter SLC7A10 |
| CAFs | Cancer-associated fibroblasts |
| CaExos | Cancer cell-derived exosomes |
| CCL | Chemokine (C-C motif) ligand |
| CXCL | Chemokine (C-X-C motif) ligand |
| DDR2 | Discoidin domain receptor 2 |
| ECM | Extracellular matrix |
| EGF | Epidermal growth factor |
| EMT | Epithelial–mesenchymal transition |
| FAP | Fibroblast activation protein |
| FGF | Fibroblast growth factor |
| FSP1 | Fibroblast-specific protein 1 |
| HDGF | Hepatoma-derived growth factor |
| HGF | Hepatic growth factor |
| IGF | Insulin-like growth factor |
| IL | Interleukin |
| KERAT | Keratin |
| miR | microRNA |
| MMP | Matrix metalloproteinase |
| NG2 | Neural/glial antigen 2 |
| PDGF-BB | Platelet-derived growth factor BB |
| PDGFRα | Platelet-derived growth factor receptor α |
| PD-L1 | Programmed cell death ligand 1 |
| RAB3B | Ras-related protein Rab-3B |
| SMA | α-Smooth muscle actin |
| TGF | Transforming growth factor |
| TNFα | Tumour necrosis factor α |
| WNT | Wingless and Int signalization |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Nakasone, E.S.; Werb, Z. Tumors as Organs: Complex Tissues That Interface with the Entire Organism. Dev. Cell 2010, 18, 884–901. [Google Scholar] [CrossRef]
- Chia, S.B.; Degregori, J. Cancer Cells Build a Bad Neighbourhood in the Gut. Nature 2021, 594, 340–341. [Google Scholar] [CrossRef]
- Campisi, J. Senescent Cells, Tumor Suppression, and Organismal Aging: Good Citizens, Bad Neighbors. Cell 2005, 120, 513–522. [Google Scholar] [CrossRef]
- Krtolica, A.; Parrinello, S.; Lockett, S.; Desprez, P.Y.; Campisi, J. Senescent Fibroblasts Promote Epithelial Cell Growth and Tumorigenesis: A Link between Cancer and Aging. Proc. Natl. Acad. Sci. USA 2001, 98, 12072–12077. [Google Scholar] [CrossRef] [PubMed]
- Fadiel, A.; Eichenbaum, K.D.; Xia, Y. Cell Interactome: Good Neighbors or Bad Neighbors. Biosci. Hypotheses 2008, 1, 255. [Google Scholar] [CrossRef][Green Version]
- Mintz, B.; Illmensee, K. Normal Genetically Mosaic Mice Produced from Malignant Teratocarcinoma Cells. Proc. Natl. Acad. Sci. USA 1975, 72, 3585–3589. [Google Scholar] [CrossRef]
- Lacina, L.; Plzak, J.; Kodet, O.; Szabo, P.; Chovanec, M.; Dvorankova, B.; Smetana, K., Jr. Cancer Microenvironment: What Can We Learn from the Stem Cell Niche. Int. J. Mol. Sci. 2015, 16, 24094–24110. [Google Scholar] [CrossRef] [PubMed]
- Kodet, O.; Kučera, J.; Strnadová, K.; Dvořánková, B.; Štork, J.; Lacina, L.; Smetana, K. Cutaneous Melanoma Dissemination Is Dependent on the Malignant Cell Properties and Factors of Intercellular Crosstalk in the Cancer Microenvironment (Review). Int. J. Oncol. 2020, 57, 619–630. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Firestein, S.G.; Budd, R.C.; Gabriel, S.E.; McInnes, I.B.; O’Dell, J.R. Firestein: Kelley’s Textbook of Rheumatology, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Mikkola, M.L. Genetic Basis of Skin Appendage Development. Semin. Cell Dev. Biol. 2007, 18, 225–236. [Google Scholar] [CrossRef]
- Avagliano, A.; Fiume, G.; Ruocco, M.R.; Martucci, N.; Vecchio, E.; Insabato, L.; Russo, D.; Accurso, A.; Masone, S.; Montagnani, S.; et al. Influence of Fibroblasts on Mammary Gland Development, Breast Cancer Microenvironment Remodeling, and Cancer Cell Dissemination. Cancers 2020, 12, 1697. [Google Scholar] [CrossRef]
- Macias, H.; Hinck, L. Mammary Gland Development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef]
- Schittny, J.C. Development of the Lung. Cell Tissue Res. 2017, 367, 427–444. [Google Scholar] [CrossRef]
- Duncan, S.A. Mechanisms Controlling Early Development of the Liver. Mech. Dev. 2003, 120, 19–33. [Google Scholar] [CrossRef]
- Jørgensen, M.C.; Ahnfelt-Rønne, J.; Hald, J.; Madsen, O.D.; Serup, P.; Hecksher-Sørensen, J. An Illustrated Review of Early Pancreas Development in the Mouse. Endocr. Rev. 2007, 28, 685–705. [Google Scholar] [CrossRef]
- Gittes, G.K. Developmental Biology of the Pancreas: A Comprehensive Review. Dev. Biol. 2009, 326, 4–35. [Google Scholar] [CrossRef]
- Driskell, R.R. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Watt, F.M. Understanding Fibroblast Heterogeneity in the Skin. Trends Cell Biol. 2015, 25, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Clavel, C.; Rendl, M.; Watt, F.M. Hair Follicle Dermal Papilla Cells at a Glance. J. Cell Sci. 2011, 124, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Haydont, V.; Neiveyans, V.; Perez, P.; Busson, É.; Lataillade, J.J.; Asselineau, D.; Fortunel, N.O. Fibroblasts from the Human Skin Dermo-Hypodermal Junction Are Distinct from Dermal Papillary and Reticular Fibroblasts and from Mesenchymal Stem Cells and Exhibit a Specific Molecular Profile Related to Extracellular Matrix Organization and Modeling. Cells 2020, 9, 368. [Google Scholar] [CrossRef] [PubMed]
- Philippeos, C.; Telerman, S.B.; Oulès, B.; Pisco, A.O.; Shaw, T.J.; Elgueta, R.; Lombardi, G.; Driskell, R.R.; Soldin, M.; Lynch, M.D.; et al. Spatial and Single-Cell Transcriptional Profiling Identifies Functionally Distinct Human Dermal Fibroblast Subpopulations. J. Investig. Dermatol. 2018, 138, 811–825. [Google Scholar] [CrossRef]
- Novotný, J.; Strnadová, K.; Dvořánková, B.; Kocourková, Š.; Jakša, R.; Dundr, P.; Pačes, V.; Smetana, K.; Kolář, M.; Lacina, L. Single-Cell RNA Sequencing Unravels Heterogeneity of the Stromal Niche in Cutaneous Melanoma Heterogeneous Spheroids. Cancers 2020, 12, 3324. [Google Scholar] [CrossRef] [PubMed]
- Kareva, I. What Can Ecology Teach Us about Cancer? Transl. Oncol. 2011, 4, 266–270. [Google Scholar] [CrossRef]
- Hirata, E.; Girotti, M.R.; Viros, A.; Hooper, S.; Spencer-Dene, B.; Matsuda, M.; Larkin, J.; Marais, R.; Sahai, E. Intravital Imaging Reveals How BRAF Inhibition Generates Drug-Tolerant Microenvironments with High Integrin Β1/FAK Signaling. Cancer Cell 2015, 27, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Beacham, E.C.D.A. Stromagenesis: The Changing Face of Fibroblastic Microenvironments during Tumor Progression. Semin. Cancer Biol. 2005, 15, 329–341. [Google Scholar] [CrossRef]
- Rodrigues, P.; Vanharanta, S. Circulating Tumor Cells: Come Together, Right Now, over Metastasis. Cancer Discov. 2019, 9, 22. [Google Scholar] [CrossRef]
- Paget, S. THE DISTRIBUTION OF SECONDARY GROWTHS IN CANCER OF THE BREAST. Lancet 1889, 133, 571–573. [Google Scholar] [CrossRef]
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of Deaths from Cancer Caused by Metastases? Cancer Med. 2019, 8, 5574. [Google Scholar] [CrossRef]
- Thompson, A.M.; Steel, C.M.; Chetty, U.; Carter, D.C. Evidence for the Multistep Theory of Carcinogenesis in Human Breast Cancer. Breast 1992, 1, 29–34. [Google Scholar] [CrossRef]
- Lacina, L.; Čoma, M.; Dvořánková, B.; Kodet, O.; Melegová, N.; Gál, P.; Smetana, K. Evolution of Cancer Progression in the Context of Darwinism. Anticancer. Res. 2019, 39, 1–16. [Google Scholar] [CrossRef]
- Lacina, L.; Kodet, O.; Dvořánková, B.; Szabo, P.; Smetana, K. Ecology of Melanoma Cell. Histol. Histopathol. 2018, 33, 247–254. [Google Scholar] [CrossRef]
- Josson, S.; Matsuoka, Y.; Chung, L.W.K.; Zhau, H.E.; Wang, R. Tumor-Stroma Co-Evolution in Prostate Cancer Progression and Metastasis. Semin. Cell Dev. Biol. 2010, 21, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Dujon, A.M.; Aktipis, A.; Alix-Panabières, C.; Amend, S.R.; Boddy, A.M.; Brown, J.S.; Capp, J.P.; DeGregori, J.; Ewald, P.; Gatenby, R.; et al. Identifying Key Questions in the Ecology and Evolution of Cancer. Evol. Appl. 2021, 14, 877–892. [Google Scholar] [CrossRef]
- Papkou, A.; Gokhale, C.S.; Traulsen, A.; Schulenburg, H. Host–Parasite Coevolution: Why Changing Population Size Matters. Zoology 2016, 119, 330–338. [Google Scholar] [CrossRef]
- Becht, E.; Giraldo, N.A.; Lacroix, L.; Buttard, B.; Elarouci, N.; Petitprez, F.; Selves, J.; Laurent-Puig, P.; Sautès-Fridman, C.; Fridman, W.H.; et al. Estimating the Population Abundance of Tissue-Infiltrating Immune and Stromal Cell Populations Using Gene Expression. Genome Biol. 2016, 17, 218. [Google Scholar] [CrossRef] [PubMed]
- Novák, Š.; Kolář, M.; Szabó, A.; Vernerová, Z.; Lacina, L.; Strnad, H.; Šáchová, J.; Hradilová, M.; Havránek, J.; Španko, M.; et al. Desmoplastic Crosstalk in Pancreatic Ductal Adenocarcinoma Is Reflected by Different Responses of Panc-1, MIAPaCa-2, PaTu-8902, and CAPAN-2 Cell Lines to Cancer-Associated/Normal Fibroblasts. Cancer Genom. Proteom. 2021, 18, 221–243. [Google Scholar] [CrossRef] [PubMed]
- Norton, J.; Foster, D.; Chinta, M.; Titan, A.; Longaker, M. Pancreatic Cancer Associated Fibroblasts (CAF): Under-Explored Target for Pancreatic Cancer Treatment. Cancers 2020, 12, 1347. [Google Scholar] [CrossRef]
- Balaziova, E.; Vymola, P.; Hrabal, P.; Mateu, R.; Zubal, M.; Tomas, R.; Netuka, D.; Kramar, F.; Zemanova, Z.; Svobodova, K.; et al. Fibroblast Activation Protein Expressing Mesenchymal Cells Promote Glioblastoma Angiogenesis. Cancers 2021, 13, 3304. [Google Scholar] [CrossRef]
- Pachva, M.C.; Lai, H.; Jia, A.; Rouleau, M.; Sorensen, P.H. Extracellular Vesicles in Reprogramming of the Ewing Sarcoma Tumor Microenvironment. Front. Cell Dev. Biol. 2021, 9, 726205. [Google Scholar] [CrossRef]
- Song, Y.J.; Xu, Y.; Deng, C.; Zhu, X.; Fu, J.; Chen, H.; Lu, J.; Xu, H.; Song, G.; Tang, Q.; et al. Gene Expression Classifier Reveals Prognostic Osteosarcoma Microenvironment Molecular Subtypes. Front. Immunol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Raffaghello, L.; Vacca, A.; Pistoia, V.; Ribatti, D. Cancer Associated Fibroblasts in Hematological Malignancies. Oncotarget 2015, 6, 2589–2603. [Google Scholar] [CrossRef] [PubMed]
- Blentic, A.; Tandon, P.; Payton, S.; Walshe, J.; Carney, T.; Kelsh, R.N.; Mason, I.; Graham, A. The Emergence of Ectomesenchyme. Dev. Dyn. 2008, 237, 592–601. [Google Scholar] [CrossRef]
- LeBleu, V.S.; Neilson, E.G. Origin and Functional Heterogeneity of Fibroblasts. FASEB J. 2020, 34, 3519–3536. [Google Scholar] [CrossRef]
- Živicová, V.; Lacina, L.; Mateu, R.; Smetana, K.; Kavková, R.; Krejcí, E.D.; Grim, M.; Kvasilová, A.; Borský, J.; Strnad, H.; et al. Analysis of Dermal Fibroblasts Isolated from Neonatal and Child Cleft Lip and Adult Skin: Developmental Implications on Reconstructive Surgery. Int. J. Mol. Med. 2017, 40, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yang, J.; Liu, J.; Wang, Y.; Mu, J.; Zeng, Q.; Deng, S.; Zhou, H. Signaling Pathways in Cancer-Associated Fibroblasts and Targeted Therapy for Cancer. Signal Transduct. Target. Ther. 2021, 6, 218. [Google Scholar] [CrossRef] [PubMed]
- Bukkuri, A.; Adler, F.R. Viewing Cancer through the Lens of Corruption: Using Behavioral Ecology to Understand Cancer. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Lee, Y.T.; Tan, Y.J.; Falasca, M.; Oon, C.E. Cancer-Associated Fibroblasts: Epigenetic Regulation and Therapeutic Intervention in Breast Cancer. Cancers 2020, 12, 2949. [Google Scholar] [CrossRef] [PubMed]
- Lacina, L.; Brábek, J.; Král, V.; Kodet, O.; Smetana, K. Interleukin-6: A Molecule with Complex Biological Impact in Cancer. Histol. Histopathol. 2019, 34, 125–136. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Petersen, O.W.; Nielsen, H.L.; Gudjonsson, T.; Villadsen, R.; Rank, F.; Niebuhr, E.; Bissell, M.J.; Rønnov-Jessen, L. Epithelial to Mesenchymal Transition in Human Breast Cancer Can Provide a Nonmalignant Stroma. Am. J. Pathol. 2003, 162, 391–402. [Google Scholar] [CrossRef]
- Kopantzev, E.P.; Vayshlya, N.A.; Kopantseva, M.R.; Egorov, V.I.; Pikunov, M.; Zinovyeva, M.V.; Vinogradova, T.V.; Zborovskaya, I.B.; Sverdlov, E.D. Cellular and Molecular Phenotypes of Proliferating Stromal Cells from Human Carcinomas. Br. J. Cancer 2010, 102, 1533–1540. [Google Scholar] [CrossRef]
- Polyak, K.; Haviv, I.; Campbell, I.G. Co-Evolution of Tumor Cells and Their Microenvironment. Trends Genet. 2009, 25, 30–38. [Google Scholar] [CrossRef]
- Haviv, I.; Polyak, K.; Qiu, W.; Hu, M.; Campbell, I. Origin of Carcinoma Associated Fibroblasts. Cell Cycle (Georget. Tex.) 2009, 8, 589–595. [Google Scholar] [CrossRef]
- Kodet, O.; Dvořánková, B.; Bendlová, B.; Sýkorová, V.; Krajsová, I.; Štork, J.; Kučera, J.; Szabo, P.; Strnad, H.; Kolář, M.; et al. Microenvironment-Driven Resistance to B-Raf Inhibition in a Melanoma Patient Is Accompanied by Broad Changes of Gene Methylation and Expression in Distal Fibroblasts. Int. J. Mol. Med. 2018, 41, 2687–2703. [Google Scholar] [CrossRef] [PubMed]
- Dvořánková, B.; Smetana, K.; Říhová, B.; Kučera, J.; Mateu, R.; Szabo, P. Cancer-Associated Fibroblasts Are Not Formed from Cancer Cells by Epithelial-to-Mesenchymal Transition in Nu/Nu Mice. Histochem. Cell Biol. 2015, 143, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Gunaydin, G.; Kesikli, S.A.; Guc, D. Cancer Associated Fibroblasts Have Phenotypic and Functional Characteristics Similar to the Fibrocytes That Represent a Novel MDSC Subset. Oncoimmunology 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.; Muñoz-Durango, N.; Riedel, C.A.; Echeverría, C.; Kalergis, A.M.; Cabello-Verrugio, C.; Simon, F. Endothelial-to-Mesenchymal Transition: Cytokine-Mediated Pathways That Determine Endothelial Fibrosis under Inflammatory Conditions. Cytokine Growth Factor Rev. 2017, 33, 41–54. [Google Scholar] [CrossRef]
- Ganguly, D.; Chandra, R.; Karalis, J.; Teke, M.; Aguilera, T.; Maddipati, R.; Wachsmann, M.B.; Ghersi, D.; Siravegna, G.; Zeh, H.J.; et al. Cancer-Associated Fibroblasts: Versatile Players in the Tumor Microenvironment. Cancers 2020, 12, 2652. [Google Scholar] [CrossRef]
- Iyoshi, S.; Yoshihara, M.; Nakamura, K.; Sugiyama, M.; Koya, Y.; Kitami, K.; Uno, K.; Mogi, K.; Tano, S.; Tomita, H.; et al. Pro-Tumoral Behavior of Omental Adipocyte-Derived Fibroblasts in Tumor Microenvironment at the Metastatic Site of Ovarian Cancer. Int. J. Cancer 2021, 149, 1961–1972. [Google Scholar] [CrossRef]
- Louault, K.; Li, R.R.; De Clerck, Y.A. Cancer-Associated Fibroblasts: Understanding Their Heterogeneity. Cancers 2020, 12, 3108. [Google Scholar] [CrossRef]
- Manoukian, P.; Bijlsma, M.; van Laarhoven, H. The Cellular Origins of Cancer-Associated Fibroblasts and Their Opposing Contributions to Pancreatic Cancer Growth. Front. Cell Dev. Biol. 2021, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Fang, Q.; Liu, P.; Ma, D.; Cao, S.; Zhang, L.; Chen, Q.; Hu, T.; Wang, J. Mesenchymal Stem Cells With Cancer-Associated Fibroblast-Like Phenotype Stimulate SDF-1/CXCR4 Axis to Enhance the Growth and Invasion of B-Cell Acute Lymphoblastic Leukemia Cells Through Cell-to-Cell Communication. Front. Cell Dev. Biol. 2021, 9, 9. [Google Scholar] [CrossRef]
- Dvořánková, B.; Szabo, P.; Lacina, L.; Gal, P.; Uhrova, J.; Zima, T.; Kaltner, H.; André, S.; Gabius, H.J.; Sykova, E.; et al. Human Galectins Induce Conversion of Dermal Fibroblasts into Myofibroblasts and Production of Extracellular Matrix: Potential Application in Tissue Engineering and Wound Repair. Cells Tissues Organs 2011, 194, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Xia, Q.; Muhammad, T.; Liu, L.; Meng, X.; Bars-Cortina, D.; Khan, A.A.; Huang, Y.; Dong, L. Glioblastoma Therapy: Rationale for a Mesenchymal Stem Cell-Based Vehicle to Carry Recombinant Viruses. Stem Cell Rev. Rep. 2021, 1–21. [Google Scholar] [CrossRef]
- Moreno, R. Mesenchymal Stem Cells and Oncolytic Viruses: Joining Forces against Cancer. J. ImmunoTherapy Cancer 2021, 9, e001684. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.; Woods, E.L.; Dally, J.; Kong, D.; Steadman, R.; Moseley, R.; Midgley, A.C. Myofibroblasts: Function, Formation, and Scope of Molecular Therapies for Skin Fibrosis. Biomolecules 2021, 11, 1095. [Google Scholar] [CrossRef] [PubMed]
- Mifková, A.; Kodet, O.; Szabo, P.; Kučera, J.; Dvořánková, B.; André, S.; Koripelly, G.; Gabius, H.J.; Lehn, J.M.; Smetana, K. Synthetic Polyamine BPA-C8 Inhibits TGF-Β1-Mediated Conversion of Human Dermal Fibroblast to Myofibroblasts and Establishment of Galectin-1-Rich Extracellular Matrix in Vitro. ChemBioChem 2014, 15, 1465–1470. [Google Scholar] [CrossRef]
- Han, C.; Liu, T.; Yin, R. Biomarkers for Cancer-Associated Fibroblasts. Biomark. Res. 2020, 8, 64. [Google Scholar] [CrossRef]
- Boyd, L.N.C.; Andini, K.D.; Peters, G.J.; Kazemier, G.; Giovannetti, E. Heterogeneity and Plasticity of Cancer-Associated Fibroblasts in the Pancreatic Tumor Microenvironment. Semin. Cancer Biol. 2021, S1044-579X(21)00056-0. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Zhang, Q.; Liu, B.; Cheng, Y.; Zhang, Y.; Sun, Y.; Liu, J. Exosomal MiR-590-3p Derived from Cancer-Associated Fibroblasts Confers Radioresistance in Colorectal Cancer. Mol. Ther.-Nucleic Acids 2021, 24, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.S.; Kanugula, S.S.; Sudhir, S.; Pereira, M.P.; Jain, S.; Aghi, M.K. The Role of Cancer-Associated Fibroblasts in Tumor Progression. Cancers 2021, 13, 1399. [Google Scholar] [CrossRef] [PubMed]
- Busek, P.; Mateu, R.; Zubal, M.; Kotackova, L.; Sedo, A. Targeting Fibroblast Activation Protein in Cancer-Prospects and Caveats. Front. Biosci.-Landmark 2018, 23, 1933–1968. [Google Scholar]
- Šimková, A.; Bušek, P.; Šedo, A.; Konvalinka, J. Molecular Recognition of Fibroblast Activation Protein for Diagnostic and Therapeutic Applications. Biochim. Biophys. Acta-Proteins Proteom. 2020, 1868, 140409. [Google Scholar] [CrossRef] [PubMed]
- Irvine, A.F.; Waise, S.; Green, E.W.; Stuart, B.; Thomas, G.J. Characterising Cancer-Associated Fibroblast Heterogeneity in Non-Small Cell Lung Cancer: A Systematic Review and Meta-Analysis. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Miyashita, N.; Saito, A. Organ Specificity and Heterogeneity of Cancer-Associated Fibroblasts in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 10973. [Google Scholar] [CrossRef]
- Simon, T.; Salhia, B. Cancer Associated Fibroblast Subpopulations with Diverse and Dynamic Roles in the Tumor Microenvironment. Mol. Cancer Res. 2021. [Google Scholar] [CrossRef]
- Watt, D.M.; Morton, J.P. Heterogeneity in Pancreatic Cancer Fibroblasts—TGFβ as a Master Regulator? Cancers 2021, 13, 4984. [Google Scholar] [CrossRef]
- Pradhan, R.N.; Krishnamurty, A.T.; Fletcher, A.L.; Turley, S.J.; Müller, S. A Bird’s Eye View of Fibroblast Heterogeneity: A Pan-Disease, Pan-Cancer Perspective. Immunol. Rev. 2021, 302, 299–320. [Google Scholar] [CrossRef]
- Dvořánková, B.; Szabo, P.; Lacina, L.; Kodet, O.; Matouškové, E.; Smetana, K.; Matoušková, E.; Smetana, K. Fibroblasts Prepared from Different Types of Malignant Tumors Stimulate Expression of Luminal Marker Keratin 8 in the EM-G3 Breast Cancer Cell Line. Histochem. Cell Biol. 2012, 137, 679–685. [Google Scholar] [CrossRef]
- Geng, X.; Chen, H.; Zhao, L.; Hu, J.; Yang, W.; Li, G.; Cheng, C.; Zhao, Z.; Zhang, T.; Li, L.; et al. Cancer-Associated Fibroblast (CAF) Heterogeneity and Targeting Therapy of CAFs in Pancreatic Cancer. Front. Cell Dev. Biol. 2021, 9, 655152. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F. Tumors: Wounds That Do Not Heal. Cancer Immunol. Res. 2009, 315, 1650–1659. [Google Scholar] [CrossRef] [PubMed]
- Smetana, K.; Szabo, P.; Gál, P.; André, S.; Gabius, H.J.; Kodet, O.; Dvořánková, B. Emerging Role of Tissue Lectins as Microenvironmental Effectors in Tumors and Wounds. Histol. Histopathol. 2015, 30, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Szabó, P.; Kolář, M.; Dvořánková, B.; Lacina, L.; Štork, J.; Vlček, Č.; Strnad, H.; Tvrdek, M.; Smetana, K., Jr. Mouse 3T3 Fibroblasts under the Influence of Fibroblasts Isolated from Stroma of Human Basal Cell Carcinoma Acquire Properties of Multipotent Stem Cells. Biol. Cell 2011, 103, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Krejčí, E.; Dvořánková, B.; Szabo, P.; Naňka, O.; Strnad, H.; Kodet, O.; Lacina, L.; Kolář, M.; Smetana, K. Fibroblasts as Drivers of Healing and Cancer Progression: From In Vitro Experiments to Clinics; CRC Press: Boca Raton, FL, USA, 2016; ISBN 9781498704656. [Google Scholar]
- Lacina, L.; Smetana, K., Jr.; Dvořánková, B.; Pytlík, R.; Kideryová, L.; Kučerová, L.; Plzáková, Z.; Štork, J.; Gabius, H.-J.; André, S. Stromal Fibroblasts from Basal Cell Carcinoma Affect Phenotype of Normal Keratinocytes. Br. J. Dermatol. 2007, 156, 819–829. [Google Scholar] [CrossRef]
- Lacina, L.; Dvořánkova, B.; Smetana, K., Jr.; Chovanec, M.; Plzǎk, J.; Tachezy, R.; Kideryovǎ, L.; Kučerová, L.; Čada, Z.; Bouček, J.; et al. Marker Profiling of Normal Keratinocytes Identifies the Stroma from Squamous Cell Carcinoma of the Oral Cavity as a Modulatory Microenvironment in Co-Culture. Int. J. Radiat. Biol. 2007, 83, 837–848. [Google Scholar] [CrossRef]
- Ishii, T.; Suzuki, A.; Kuwata, T.; Hisamitsu, S.; Hashimoto, H.; Ohara, Y.; Yanagihara, K.; Mitsunaga, S.; Yoshino, T.; Kinoshita, T.; et al. Drug-Exposed Cancer-Associated Fibroblasts Facilitate Gastric Cancer Cell Progression Following Chemotherapy. Gastric Cancer 2021, 24, 810–822. [Google Scholar] [CrossRef] [PubMed]
- Kučera, J.; Strnadová, K.; Dvořánková, B.; Lacina, L.; Krajsová, I.; Štork, J.; Kovářová, H.; Skalníková, H.K.; Vodička, P.; Motlík, J.; et al. Serum Proteomic Analysis of Melanoma Patients with Immunohistochemical Profiling of Primary Melanomas and Cultured Cells: Pilot Study. Oncol. Rep. 2019, 42, 1793–1804. [Google Scholar] [CrossRef]
- Loumaye, A.; Thissen, J.P. Biomarkers of Cancer Cachexia. Clin. Biochem. 2017, 50, 1281–1288. [Google Scholar] [CrossRef]
- Cehreli, R.; Yavuzsen, T.; Ates, H.; Akman, T.; Ellidokuz, H.; Oztop, I. Can Inflammatory and Nutritional Serum Markers Predict Chemotherapy Outcomes and Survival in Advanced Stage Nonsmall Cell Lung Cancer Patients? BioMed Res. Int. 2019, 2019, 1–8. [Google Scholar] [CrossRef]
- Zivicova, V.; Gal, P.; Mifkova, A.; Novak, S.; Kaltner, H.; Kolar, M.; Strnad, H.; Sachova, J.; Hradilova, M.; Chovanec, M.; et al. Detection of Distinct Changes in Gene-Expression Profiles in Specimens of Tumors and Transition Zones of Tenascin-Positive/-Negative Head and Neck Squamous Cell Carcinoma. Anticancer. Res. 2018, 38, 1279–1290. [Google Scholar] [CrossRef]
- Libring, S.; Shinde, A.; Chanda, M.K.; Nuru, M.; George, H.; Saleh, A.M.; Abdullah, A.; Kinzer-Ursem, T.L.; Calve, S.; Wendt, M.K.; et al. The Dynamic Relationship of Breast Cancer Cells and Fibroblasts in Fibronectin Accumulation at Primary and Metastatic Tumor Sites. Cancers 2020, 12, 1270. [Google Scholar] [CrossRef]
- Sapudom, J.; Müller, C.D.; Nguyen, K.T.; Martin, S.; Anderegg, U.; Pompe, T. Matrix Remodeling and Hyaluronan Production by Myofibroblasts and Cancer-Associated Fibroblasts in 3d Collagen Matrices. Gels 2020, 6, 33. [Google Scholar] [CrossRef]
- Hashimoto, M.; Uesugi, N.; Osakabe, M.; Yanagawa, N.; Otsuka, K.; Kajiwara, Y.; Ueno, H.; Sasaki, A.; Sugai, T. Expression Patterns of Microenvironmental Factors and Tenascin-C at the Invasive Front of Stage II and III Colorectal Cancer: Novel Tumor Prognostic Markers. Front. Oncol. 2021, 11, 690816. [Google Scholar] [CrossRef] [PubMed]
- Kay, E.J.; Koulouras, G.; Zanivan, S. Regulation of Extracellular Matrix Production in Activated Fibroblasts: Roles of Amino Acid Metabolism in Collagen Synthesis. Front. Oncol. 2021, 11, 719922. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, V.; Chao, L.; Woodley, D.T.; Murrell, D.; Kim, M.; O’Toole, E.A.; Ly, A.; Cogan, J.; Mosallaei, D.; Wysong, A.; et al. Over-Expression of Stromal Periostin Correlates with Poor Prognosis of Cutaneous Squamous Cell Carcinomas. Exp. Dermatol. 2021, 30, 698–704. [Google Scholar] [CrossRef]
- Schwörer, S.; Pavlova, N.N.; Cimino, F.V.; King, B.; Cai, X.; Sizemore, G.M.; Thompson, C.B. Fibroblast Pyruvate Carboxylase Is Required for Collagen Production in the Tumour Microenvironment. Nat. Metab. 2021, 3, 1484–1499. [Google Scholar] [CrossRef] [PubMed]
- Sueyama, T.; Kajiwara, Y.; Mochizuki, S.; Shimazaki, H.; Shinto, E.; Hase, K.; Ueno, H. Periostin as a Key Molecule Defining Desmoplastic Environment in Colorectal Cancer. Virchows Arch. 2021, 478, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Eiro, N.; Cid, S.; Fraile, M.; Cabrera, J.R.; Gonzalez, L.O.; Vizoso, F.J. Analysis of the Gene Expression Profile of Stromal Pro-Tumor Factors in Cancer-Associated Fibroblasts from Luminal Breast Carcinomas. Diagnostics 2020, 10, 865. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Akasu, R.; Miyazaki, A. Calpain-Associated Proteolytic Regulation of the Stromal Microenvironment in Cancer. Curr. Pharm. Des. 2021, 27, 3128–3138. [Google Scholar] [CrossRef]
- Hassona, Y.; Cirillo, N.; Heesom, K.; Parkinson, E.K.; Prime, S.S. Senescent Cancer-Associated Fibroblasts Secrete Active MMP-2 That Promotes Keratinocyte Dis-Cohesion and Invasion. Br. J. Cancer 2014, 111, 1230–1237. [Google Scholar] [CrossRef]
- Chang, J.; Chaudhuri, O. Beyond Proteases: Basement Membrane Mechanics and Cancer Invasion. J. Cell Biol. 2019, 218, 2456–2469. [Google Scholar] [CrossRef]
- Conti, S.; Kato, T.; Park, D.; Sahai, E.; Trepat, X.; Labernadie, A. CAFs and Cancer Cells Co-Migration in 3D Spheroid Invasion Assay. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 2179. [Google Scholar] [CrossRef]
- Miyazaki, K.; Togo, S.; Okamoto, R.; Idiris, A.; Kumagai, H.; Miyagi, Y. Collective Cancer Cell Invasion in Contact with Fibroblasts through Integrin-A5β1/Fibronectin Interaction in Collagen Matrix. Cancer Sci. 2020, 111, 4381–4392. [Google Scholar] [CrossRef]
- Mishra, P.; Banerjee, D.; Ben-Baruch, A. Chemokines at the Crossroads of Tumor-Fibroblast Interactions That Promote Malignancy. J. Leukoc. Biol. 2011, 89, 31–39. [Google Scholar] [CrossRef]
- Jobe, N.P.; Živicová, V.; Mifková, A.; Rösel, D.; Dvořánková, B.; Kodet, O.; Strnad, H.; Kolář, M.; Šedo, A.; Smetana, K.; et al. Fibroblasts Potentiate Melanoma Cells in Vitro Invasiveness Induced by UV-Irradiated Keratinocytes. Histochem. Cell Biol. 2018, 149, 503–516. [Google Scholar] [CrossRef]
- Asokan, S.; Bandapalli, O.R. CXCL8 Signaling in the Tumor Microenvironment. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 1302. [Google Scholar] [CrossRef]
- Španko, M.; Strnadová, K.; Pavlíček, A.J.; Szabo, P.; Kodet, O.; Valach, J.; Dvořánková, B.; Smetana, K.; Lacina, L. Il-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives. Int. J. Mol. Sci. 2021, 22, 1027. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed]
- Brábek, J.; Jakubek, M.; Vellieux, F.; Novotný, J.; Kolář, M.; Lacina, L.; Szabo, P.; Strnadová, K.; Rösel, D.; Dvořánková, B.; et al. Interleukin-6: Molecule in the Intersection of Cancer, Ageing and COVID-19. Int. J. Mol. Sci. 2020, 21, 7937. [Google Scholar] [CrossRef]
- Shimura, Y.; Kurosawa, H.; Tsuchiya, M.; Sawa, M.; Kaneko, H.; Liu, L.; Makino, Y.; Nojiri, H.; Iwase, Y.; Kaneko, K.; et al. Serum Interleukin 6 Levels Are Associated with Depressive State of the Patients with Knee Osteoarthritis Irrespective of Disease Severity. Clin. Rheumatol. 2017, 36, 2781–2787. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.P.; Desprez, P.Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Strnadova, K.; Sandera, V.; Dvorankova, B.; Kodet, O.; Duskova, M.; Smetana, K.; Lacina, L. Skin Aging: The Dermal Perspective. Clin. Dermatol. 2019, 37. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Interleukin-6 Signalling in Health and Disease. F1000Research 2020, 9, 1013. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Il-6 Trans-Signaling via the Soluble IL-6 Receptor: Importance for the Proinflammatory Activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, K.; Ito, T.; Ozaki, Y.; Amuro, H.; Tanaka, A.; Nishizawa, T.; Son, Y.; Inaba, M.; Nomura, S. Serum Interleukin 6 before and after Therapy with Tocilizumab Is a Principal Biomarker in Patients with Rheumatoid Arthritis. J. Rheumatol. 2013, 40, 1074–1081. [Google Scholar] [CrossRef]
- Plzák, J.; Bouček, J.; Bandúrová, V.; Kolář, M.; Hradilová, M.; Szabo, P.; Lacina, L.; Chovanec, M.; Smetana, K. The Head and Neck Squamous Cell Carcinoma Microenvironment as a Potential Target for Cancer Therapy. Cancers 2019, 11, 440. [Google Scholar] [CrossRef]
- Jobe, N.P.; Rösel, D.; Dvořánková, B.; Kodet, O.; Lacina, L.; Mateu, R.; Smetana, K.; Brábek, J. Simultaneous Blocking of IL-6 and IL-8 Is Sufficient to Fully Inhibit CAF-Induced Human Melanoma Cell Invasiveness. Histochem. Cell Biol. 2016, 146, 205–217. [Google Scholar] [CrossRef]
- Ham, I.H.; Lee, D.; Hur, H. Cancer-Associated Fibroblast-Induced Resistance to Chemotherapy and Radiotherapy in Gastrointestinal Cancers. Cancers 2021, 13, 1172. [Google Scholar] [CrossRef]
- Fu, H.; Yang, H.; Zhang, X.; Xu, W. The Emerging Roles of Exosomes in Tumor–Stroma Interaction. J. Cancer Res. Clin. Oncol. 2016, 142, 1897–1907. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Dudás, J.; Riechelmann, H.; Skvortsova, I.I. The Role of Exosomes in Cancer Metastasis. Semin. Cancer Biol. 2017, 44, 170–181. [Google Scholar] [CrossRef]
- Liu, J.; Ren, L.; Li, S.; Li, W.; Zheng, X.; Yang, Y.; Fu, W.; Yi, J.; Wang, J.; Du, G. The Biology, Function, and Applications of Exosomes in Cancer. Acta Pharm. Sin. B 2021, 11, 2783–2797. [Google Scholar] [CrossRef] [PubMed]
- Dror, S.; Sander, L.; Schwartz, H.; Sheinboim, D.; Barzilai, A.; Dishon, Y.; Apcher, S.; Golan, T.; Greenberger, S.; Barshack, I.; et al. Melanoma MiRNA Trafficking Controls Tumour Primary Niche Formation. Nat. Cell Biol. 2016, 18, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Weidle, U.H.; Birzele, F.; Kollmorgen, G.; Rüger, R. The Multiple Roles of Exosomes in Metastasis. Cancer Genom. Proteom. 2017, 14, 1–16. [Google Scholar] [CrossRef]
- Feng, W.; Dean, D.C.; Hornicek, F.J.; Shi, H.; Duan, Z. Exosomes Promote Pre-Metastatic Niche Formation in Ovarian Cancer. Mol. Cancer 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef]
- Strnadová, K.; Pfeiferová, L.; Přikryl, P.; Dvořánková, B.; Vlčák, E.; Frýdlová, J.; Vokurka, M.; Novotný, J.; Šáchová, J.; Hradilová, M.; et al. Exosomes Produced by Melanoma Cells Significantly Influence the Biological Properties of Normal and Cancer-Associated Fibroblasts. Histochem. Cell Biol. 2021, 1–20. [Google Scholar] [CrossRef]
- Lee, J.S.; Yoo, J.E.; Kim, H.; Rhee, H.; Koh, M.J.; Nahm, J.H.; Choi, J.S.; Lee, K.H.; Park, Y.N. Tumor Stroma with Senescence-Associated Secretory Phenotype in Steatohepatitic Hepatocellular Carcinoma. PLoS ONE 2017, 12, e0171922. [Google Scholar] [CrossRef]
- Yasuda, T.; Koiwa, M.; Yonemura, A.; Miyake, K.; Kariya, R.; Kubota, S.; Yokomizo-Nakano, T.; Yasuda-Yoshihara, N.; Uchihara, T.; Itoyama, R.; et al. Inflammation-Driven Senescence-Associated Secretory Phenotype in Cancer-Associated Fibroblasts Enhances Peritoneal Dissemination. Cell Rep. 2021, 34, 108779. [Google Scholar] [CrossRef]
- Kabir, T.D.; Leigh, R.J.; Tasena, H.; Mellone, M.; Coletta, R.D.; Parkinson, E.K.; Prime, S.S.; Thomas, G.J.; Paterson, I.C.; Zhou, D.; et al. A MiR-335/COX-2/PTEN Axis Regulates the Secretory Phenotype of Senescent Cancer-Associated Fibroblasts. Aging 2016, 8, 1608–1635. [Google Scholar] [CrossRef]
- Gener Lahav, T.; Adler, O.; Zait, Y.; Shani, O.; Amer, M.; Doron, H.; Abramovitz, L.; Yofe, I.; Cohen, N.; Erez, N. Melanoma-Derived Extracellular Vesicles Instigate Proinflammatory Signaling in the Metastatic Microenvironment. Int. J. Cancer 2019, 145, 2521–2534. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. Exosome and Mesenchymal Stem Cell Cross-Talk in the Tumor Microenvironment. Semin. Immunol. 2018, 35, 69–79. [Google Scholar] [CrossRef]
- Zhou, X.; Yan, T.; Huang, C.; Xu, Z.; Wang, L.; Jiang, E.; Wang, H.; Chen, Y.; Liu, K.; Shao, Z.; et al. Melanoma Cell-Secreted Exosomal MiR-155-5p Induce Proangiogenic Switch of Cancer-Associated Fibroblasts via SOCS1/JAK2/STAT3 Signaling Pathway. J. Exp. Clin. Cancer Res. 2018, 37, 242. [Google Scholar] [CrossRef]
- Hu, T.; Hu, J. Melanoma-Derived Exosomes Induce Reprogramming Fibroblasts into Cancer-Associated Fibroblasts via Gm26809 Delivery. Cell Cycle 2019, 18, 3085–3094. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Geng, Y. Exosomes Derived from Chronic Lymphocytic Leukaemia Cells Transfer MiR-146a to Induce the Transition of Mesenchymal Stromal Cells into Cancer-Associated Fibroblasts. J. Biochem. 2020, 168, 491–498. [Google Scholar] [CrossRef]
- Yang, S.S.; Ma, S.; Dou, H.; Liu, F.; Zhang, S.Y.; Jiang, C.; Xiao, M.; Huang, Y.X. Breast Cancer-Derived Exosomes Regulate Cell Invasion and Metastasis in Breast Cancer via MiR-146a to Activate Cancer Associated Fibroblasts in Tumor Microenvironment. Exp. Cell Res. 2020, 391, 111983. [Google Scholar] [CrossRef]
- Huang, Q.; Hsueh, C.Y.; Shen, Y.J.; Guo, Y.; Huang, J.M.; Zhang, Y.F.; Li, J.Y.; Gong, H.L.; Zhou, L. Small Extracellular Vesicle-Packaged TGFβ1 Promotes the Reprogramming of Normal Fibroblasts into Cancer-Associated Fibroblasts by Regulating Fibronectin in Head and Neck Squamous Cell Carcinoma. Cancer Lett. 2021, 517. [Google Scholar] [CrossRef]
- Shelton, M.; Anene, C.A.; Nsengimana, J.; Roberts, W.; Newton-Bishop, J.; Boyne, J.R. The Role of CAF Derived Exosomal MicroRNAs in the Tumour Microenvironment of Melanoma. Biochim. Biophys. Acta-Rev. Cancer 2021, 1875, 188456. [Google Scholar] [CrossRef]
- Shu, S.; Matsuzaki, J.; Want, M.Y.; Conway, A.; Benjamin-Davalos, S.; Allen, C.L.; Koroleva, M.; Battaglia, S.; Odunsi, A.; Minderman, H.; et al. An Immunosuppressive Effect of Melanoma-Derived Exosomes on NY-ESO-1 Antigen-Specific Human CD8+ T Cells Is Dependent on IL-10 and Independent of BRAFV600E Mutation in Melanoma Cell Lines. Immunol. Investig. 2020, 49, 744–757. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Y.; Chang, X.; Ba, X.; Hu, N.; Liu, Q.; Fang, L.; Wang, Z. Melanoma-Derived Exosomes Endow Fibroblasts with an Invasive Potential via Mir-21 Target Signaling Pathway. Cancer Manag. Res. 2020, 12, 12965–12974. [Google Scholar] [CrossRef]
- Yeon, J.H.; Jeong, H.E.; Seo, H.; Cho, S.; Kim, K.; Na, D.; Chung, S.; Park, J.; Choi, N.; Kang, J.Y. Cancer-Derived Exosomes Trigger Endothelial to Mesenchymal Transition Followed by the Induction of Cancer-Associated Fibroblasts. Acta Biomater. 2018, 76, 146–153. [Google Scholar] [CrossRef]
- Deep, G.; Panigrahia, G.K. Hypoxia-Induced Signaling Promotes Prostate Cancer Progression: Exosomes Role as Messenger of Hypoxic Response in Tumor Microenvironmen. Crit. Rev. Oncog. 2015, 20, 419–434. [Google Scholar] [CrossRef]
- Boussadia, Z.; Lamberti, J.; Mattei, F.; Pizzi, E.; Puglisi, R.; Zanetti, C.; Pasquini, L.; Fratini, F.; Fantozzi, L.; Felicetti, F.; et al. Acidic Microenvironment Plays a Key Role in Human Melanoma Progression through a Sustained Exosome Mediated Transfer of Clinically Relevant Metastatic Molecules. J. Exp. Clin. Cancer Res. 2018, 37, 245. [Google Scholar] [CrossRef]
- Xi, L.; Peng, M.; Liu, S.; Liu, Y.; Wan, X.; Hou, Y.; Qin, Y.; Yang, L.; Chen, S.; Zeng, H.; et al. Hypoxia-Stimulated ATM Activation Regulates Autophagy-Associated Exosome Release from Cancer-Associated Fibroblasts to Promote Cancer Cell Invasion. J. Extracell. Vesicles 2021, 10, e12146. [Google Scholar] [CrossRef]
- Melnik, B.C. MiR-21: An Environmental Driver of Malignant Melanoma? J. Transl. Med. 2015, 13, 1–16. [Google Scholar] [CrossRef]
- Dou, D.; Ren, X.; Han, M.; Xu, X.; Ge, X.; Gu, Y.; Wang, X. Cancer-Associated Fibroblasts-Derived Exosomes Suppress Immune Cell Function in Breast Cancer via the MiR-92/PD-L1 Pathway. Front. Immunol. 2020, 11, 2026. [Google Scholar] [CrossRef]
- Guo, L.; Li, B.; Yang, J.; Shen, J.; Ji, J.; Miao, M. Fibroblast-Derived Exosomal MicroRNA-369 Potentiates Migration and Invasion of Lung Squamous Cell Carcinoma Cells via NF1-Mediated MAPK Signaling Pathway. Int. J. Mol. Med. 2020, 46, 595–608. [Google Scholar] [CrossRef]
- Shan, G.; Gu, J.; Zhou, D.; Li, L.; Cheng, W.; Wang, Y.; Tang, T.; Wang, X. Cancer-Associated Fibroblast-Secreted Exosomal MiR-423-5p Promotes Chemotherapy Resistance in Prostate Cancer by Targeting GREM2 through the TGF-β Signaling Pathway. Exp. Mol. Med. 2020, 52, 1809–1822. [Google Scholar] [CrossRef]
- Chen, P.Y.; Wei, W.F.; Wu, H.Z.; Fan, L.S.; Wang, W. Cancer-Associated Fibroblast Heterogeneity: A Factor That Cannot Be Ignored in Immune Microenvironment Remodeling. Front. Immunol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, K.; Lu, X.; Wang, Y.; Chen, J. Cancer-Associated Fibroblasts-Derived Exosomes Promote Lung Cancer Progression by OIP5-AS1/ MiR-142-5p/ PD-L1 Axis. Mol. Immunol. 2021, 140, 47–58. [Google Scholar] [CrossRef]
- Kunou, S.; Shimada, K.; Takai, M.; Sakamoto, A.; Aoki, T.; Hikita, T.; Kagaya, Y.; Iwamoto, E.; Sanada, M.; Shimada, S.; et al. Exosomes Secreted from Cancer-Associated Fibroblasts Elicit Anti-Pyrimidine Drug Resistance through Modulation of Its Transporter in Malignant Lymphoma. Oncogene 2021, 40, 3989–4003. [Google Scholar] [CrossRef]
- Luo, G.; Zhang, Y.; Wu, Z.; Zhang, L.; Liang, C.; Chen, X. Exosomal LINC00355 Derived from Cancer-Associated Fibroblasts Promotes Bladder Cancer Cell Resistance to Cisplatin by Regulating MiR-34b-5p/ABCB1 Axis. Acta Biochim. Biophys. Sin. 2021, 53, 558–566. [Google Scholar] [CrossRef]
- Yin, H.; Yu, S.; Xie, Y.; Dai, X.; Dong, M.; Sheng, C.; Hu, J. Cancer-Associated Fibroblasts-Derived Exosomes Upregulate MicroRNA-135b-5p to Promote Colorectal Cancer Cell Growth and Angiogenesis by Inhibiting Thioredoxin-Interacting Protein. Cell. Signal. 2021, 84, 110029. [Google Scholar] [CrossRef]
- Yugawa, K.; Yoshizumi, T.; Mano, Y.; Itoh, S.; Harada, N.; Ikegami, T.; Kohashi, K.; Oda, Y.; Mori, M. Cancer-Associated Fibroblasts Promote Hepatocellular Carcinoma Progression through Downregulation of Exosomal MiR-150-3p. Eur. J. Surg. Oncol. 2021, 47, 384–393. [Google Scholar] [CrossRef]
- Zhang, H.W.; Shi, Y.; Liu, J.-B.; Wang, H.M.; Wang, P.Y.; Wu, Z.J.; Li, L.; Gu, L.P.; Cao, P.S.; Wang, G.R.; et al. Cancer-Associated Fibroblast-Derived Exosomal MicroRNA-24-3p Enhances Colon Cancer Cell Resistance to MTX by down-Regulating CDX2/HEPH Axis. J. Cell. Mol. Med. 2021, 25, 3699–3713. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, P.; Li, H.X. CAFs-Derived Exosomal MiRNA-130a Confers Cisplatin Resistance of NSCLC Cells through PUM2-Dependent Packaging. Int. J. Nanomed. 2021, 16, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Du, J.; Min, Z.; Ma, L.; Zhang, W.; Zhu, W.; Liu, Y. Carcinoma-Associated Fibroblasts Derived Exosomes Modulate Breast Cancer Cell Stemness through Exonic CircHIF1A by MiR-580-5p in Hypoxic Stress. Cell Death Discov. 2021, 7, 1–15. [Google Scholar] [CrossRef]
- White, J.P. IL-6, Cancer and Cachexia: Metabolic Dysfunction Creates the Perfect Storm. Transl. Cancer Res. 2017, 6, S280–S285. [Google Scholar] [CrossRef]
- Yamagata, A.S.; Freire, P.P. Are Cachexia-Associated Tumors TransmitTERS of ER Stress. Biochem. Soc. Trans. 2021, 49, 1841–1853. [Google Scholar] [CrossRef]
- Kottorou, A.; Dimitrakopoulos, F.I.; Tsezou, A. Non-Coding RNAs in Cancer-Associated Cachexia: Clinical Implications and Future Perspectives. Transl. Oncol. 2021, 14, 101101. [Google Scholar] [CrossRef]
- Miao, C.; Zhang, W.; Feng, L.; Gu, X.; Shen, Q.; Lu, S.; Fan, M.; Li, Y.; Guo, X.; Ma, Y.; et al. Cancer-Derived Exosome MiRNAs Induce Skeletal Muscle Wasting by Bcl-2-Mediated Apoptosis in Colon Cancer Cachexia. Mol. Ther.-Nucleic Acids 2021, 24, 923–938. [Google Scholar] [CrossRef] [PubMed]
- Di, W.; Zhang, W.; Zhu, B.; Li, X.; Tang, Q.; Zhou, Y. Colorectal Cancer Prompted Adipose Tissue Browning and Cancer Cachexia through Transferring Exosomal MiR-146b-5p. J. Cell. Physiol. 2021, 236, 5399–5410. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, T.; Terracina, K.P.; Raza, A.; Matsubara, H.; Takabe, K. Cancer Cachexia, Mechanism and Treatment. World J. Gastrointest. Oncol. 2015, 7, 17–29. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).