
Vitamin D3 Treatment Alters Thyroid Functional Morphology in Orchidectomized Rat Model of Osteoporosis

 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Animals and Treatments
2.2. Sample Collection and Processing
2.3. Transmission Electron Microscopy (TEM)
2.4. Immunohistochemistry (IHC) and Immunofluorescence (IFC)
2.5. Quantification of IHC and Morphometric Analysis
2.6. Hormone Analysis
2.7. Statistical Analysis
3. Results
3.1. Thyroid Weight and Body Mass
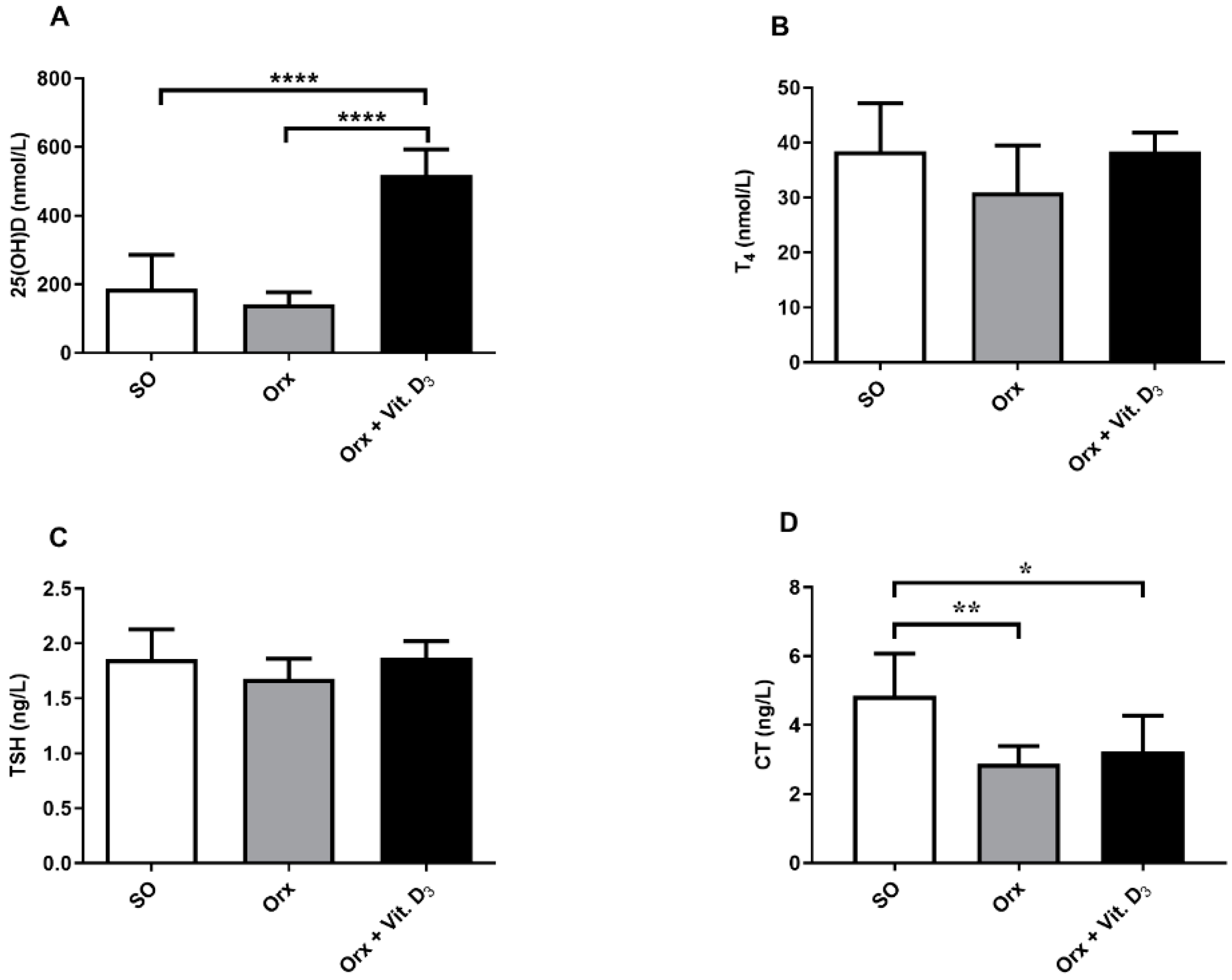
3.2. Serum Hormone Concentrations
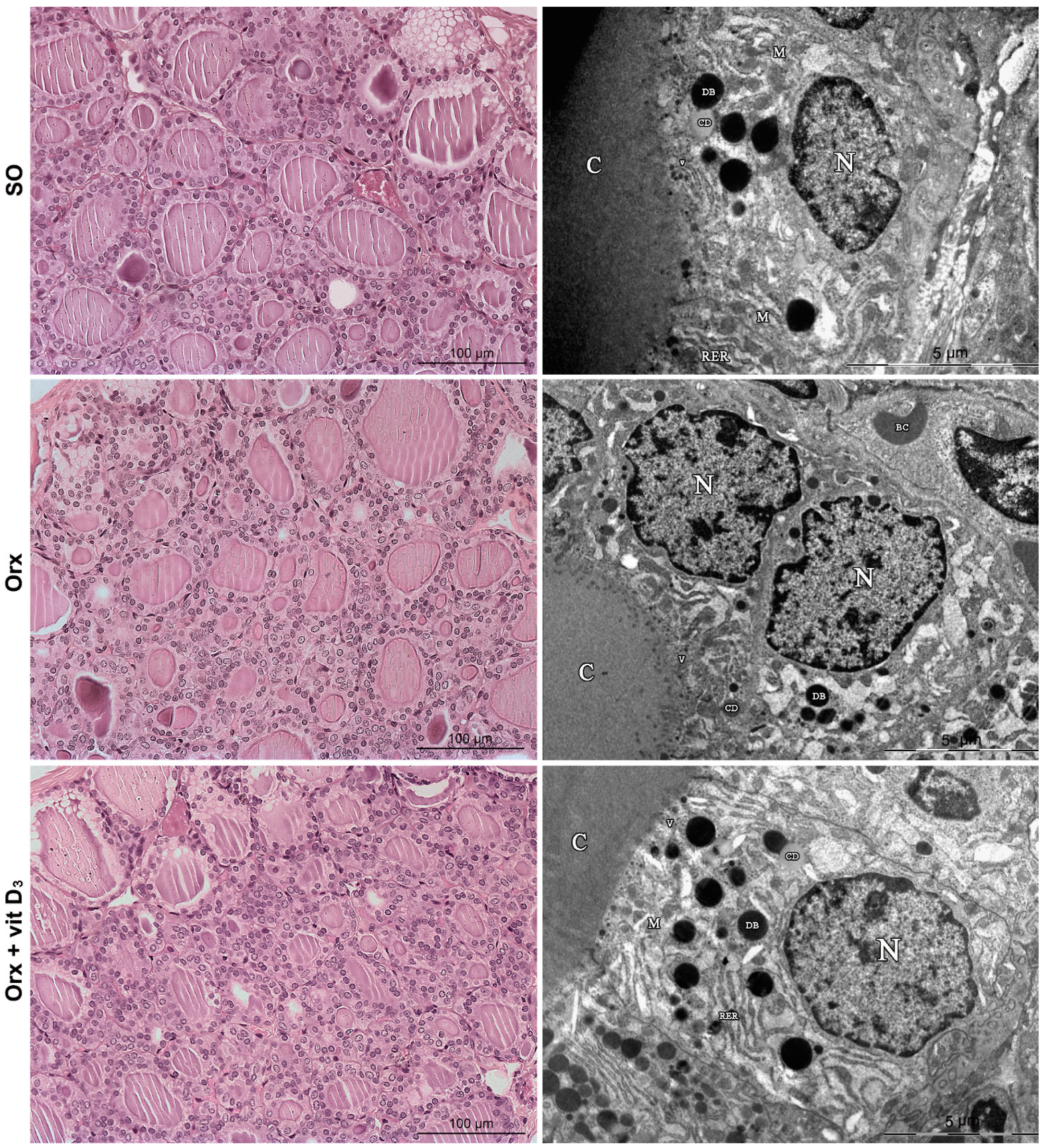
3.3. Histological and TEM Analysis of the Thyroid
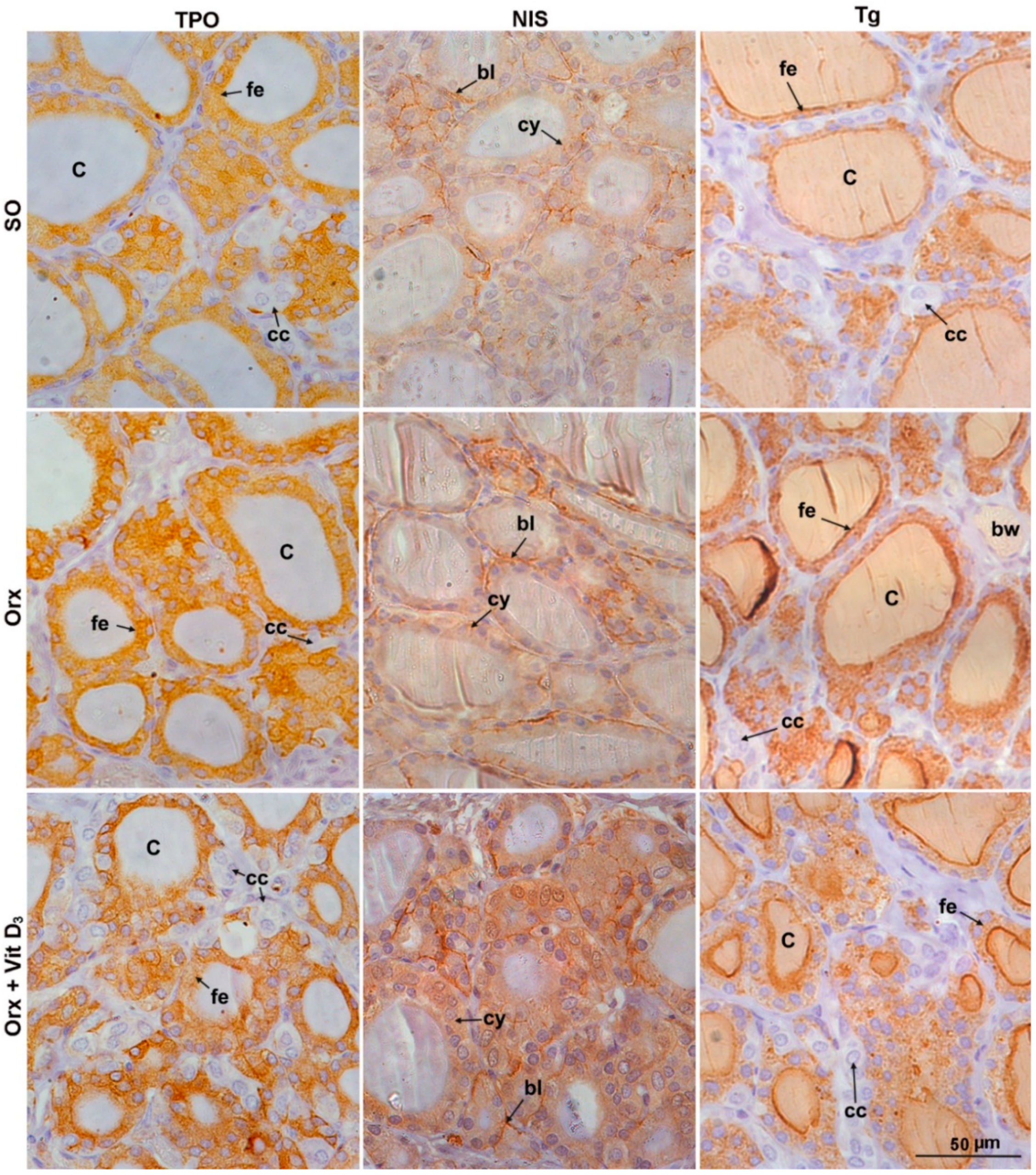
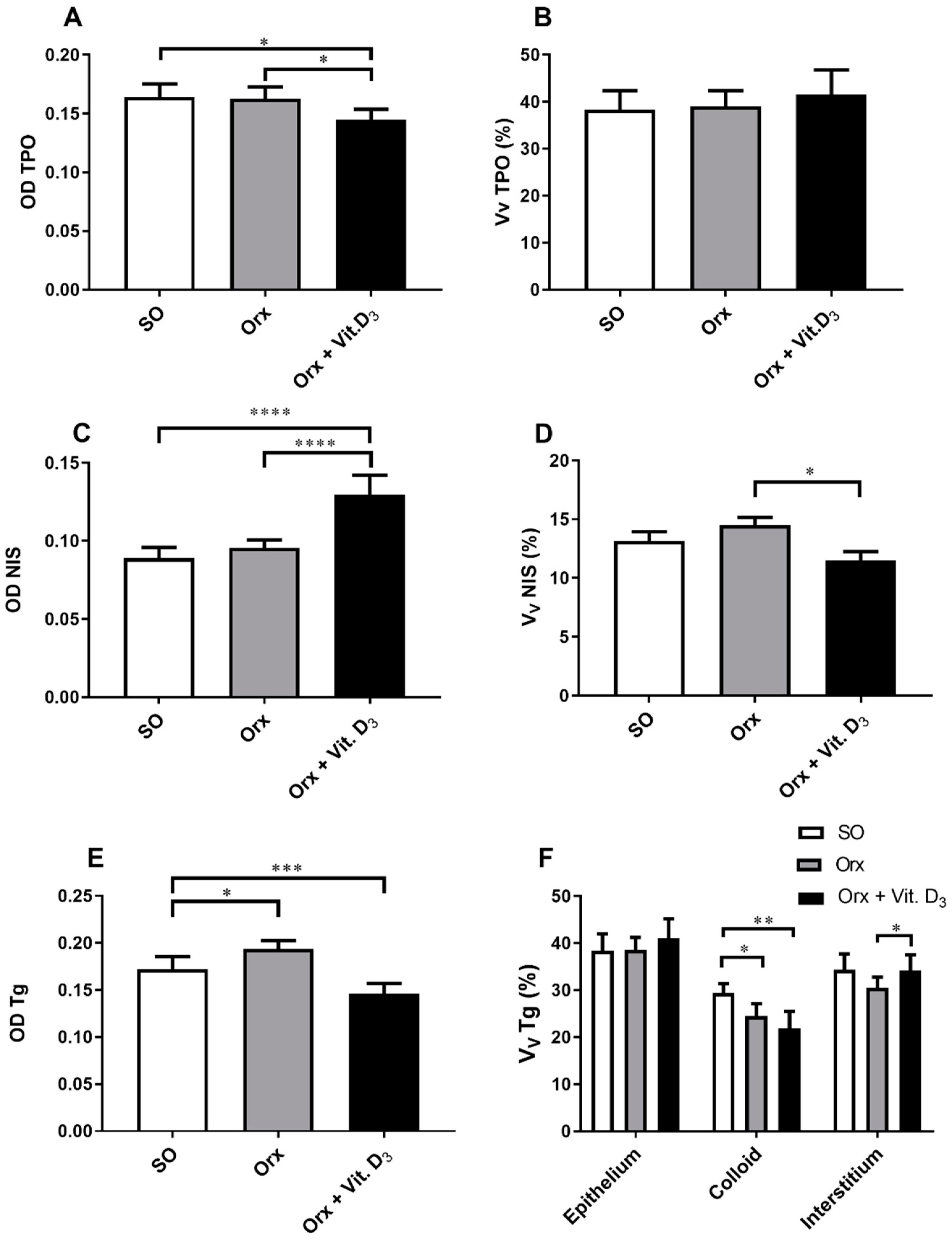
3.4. IHC Analysis of Thyroid-Specific Proteins
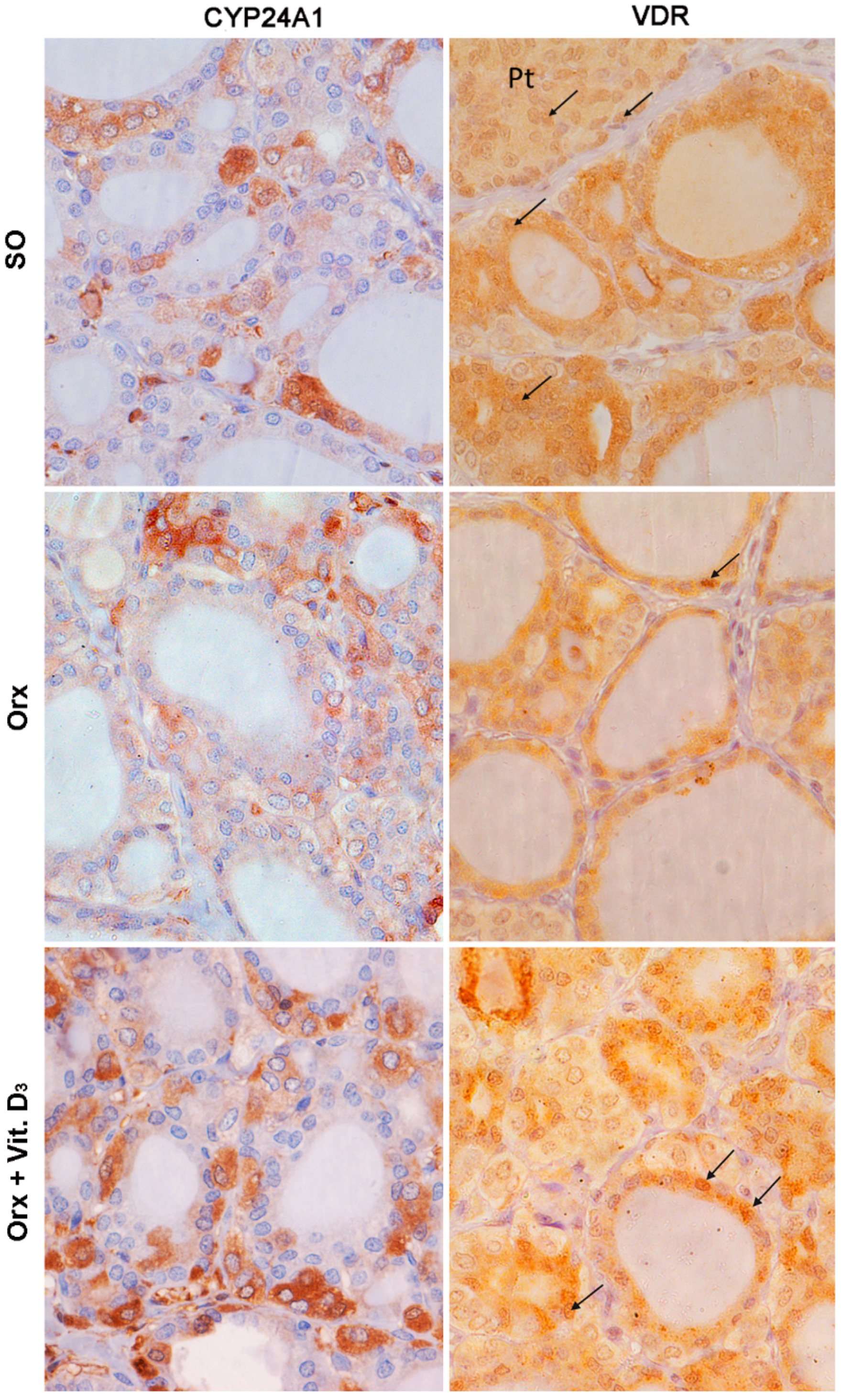
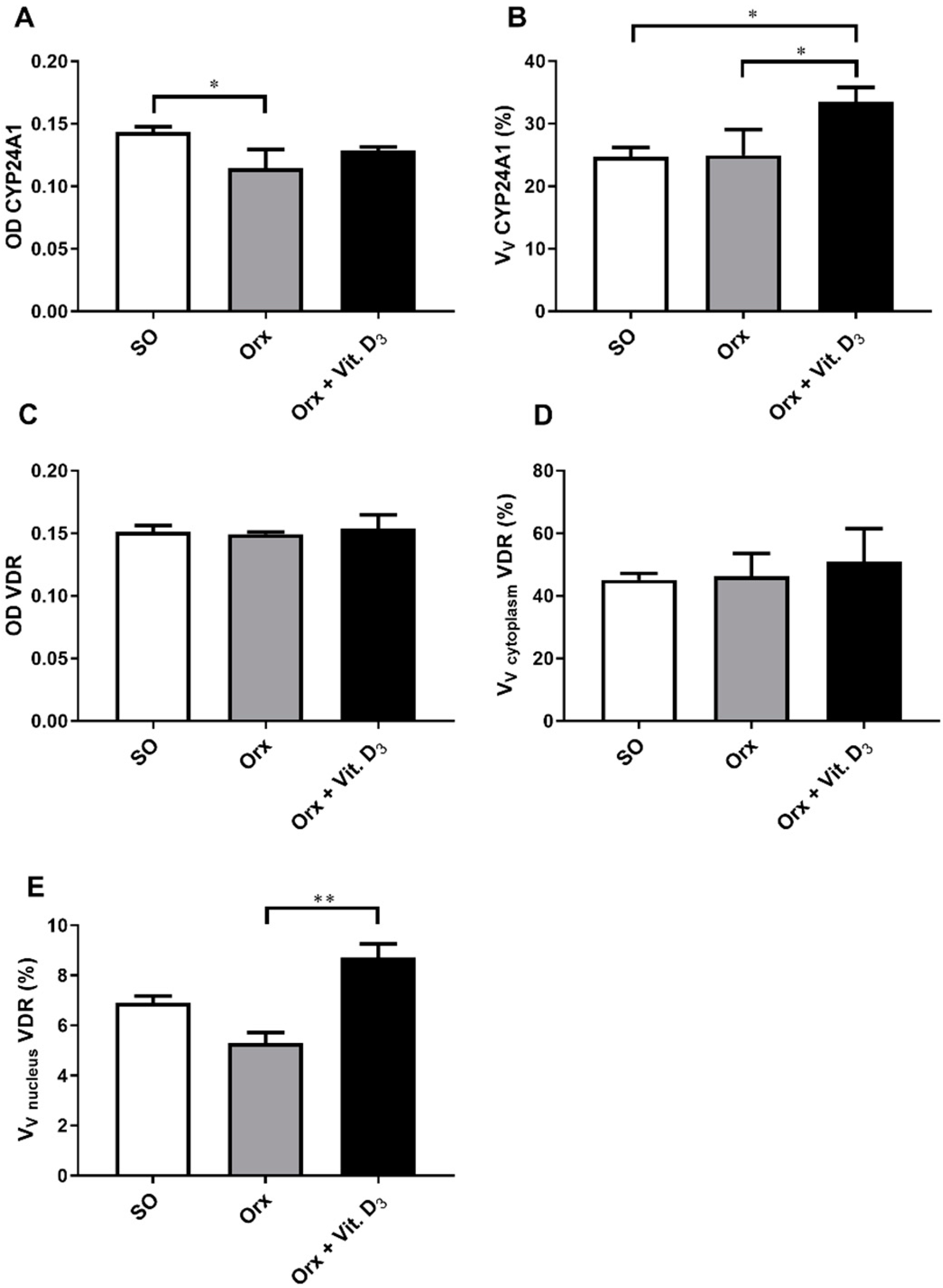
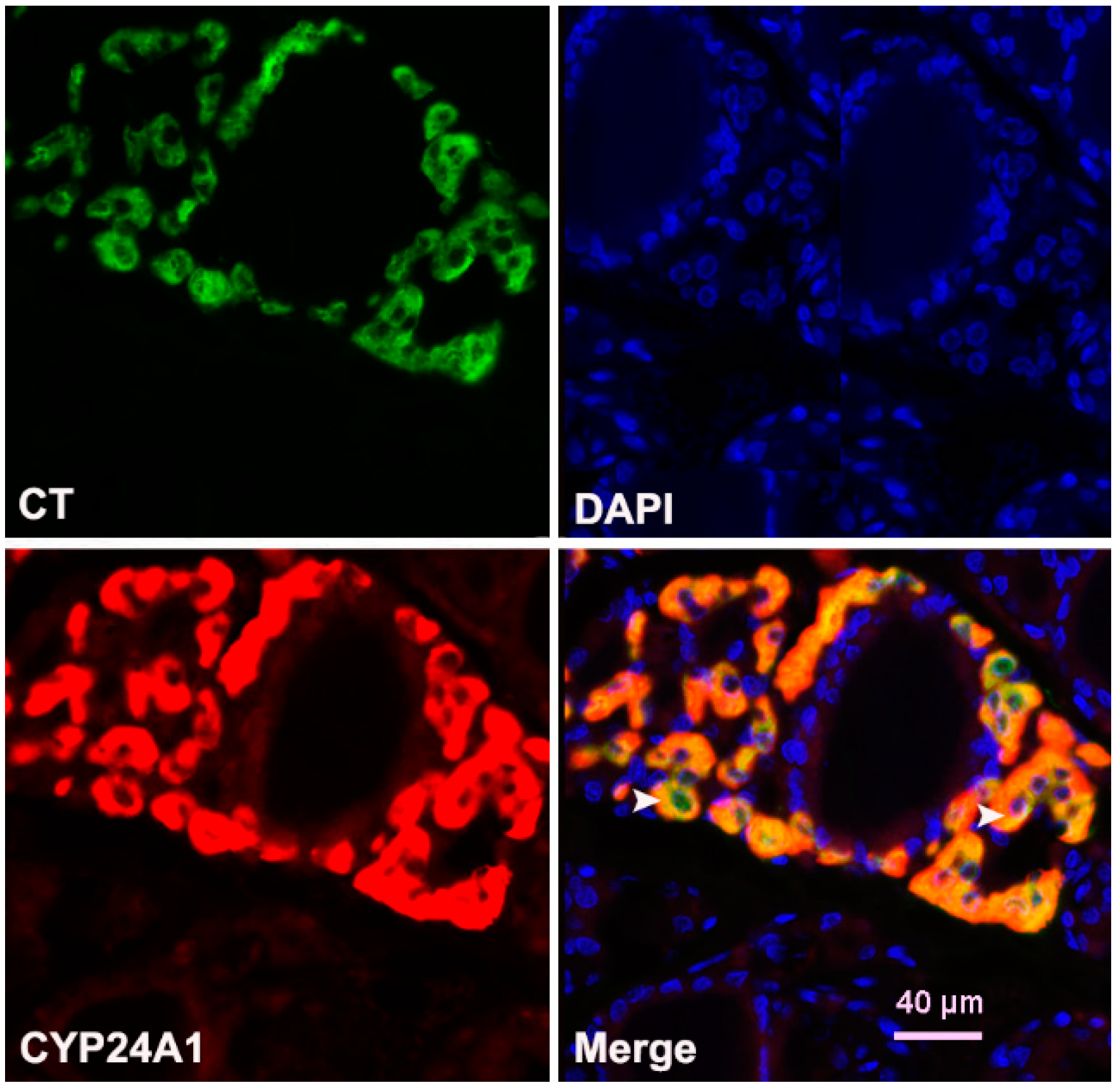
3.5. Immunohistochemical Expression of CYP24A1 and Vitamin D Receptor (VDR) in the Thyroid Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Perlman, R.L. Mouse models of human disease: An evolutionary perspective. Evol. Med. Public Health 2016, 2016, eow014. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, F.; Saba, A.; Zucchi, R. An update on vitamin d metabolism. Int. J. Mol. Sci. 2020, 21, 6573. [Google Scholar] [CrossRef]
- Wintermeyer, E.; Ihle, C.; Ehnert, S.; Stöckle, U.; Ochs, G.; de Zwart, P.; Flesch, I.; Bahrs, C.; Nussler, A.K. Crucial Role of Vitamin D in the Musculoskeletal System. Nutrients 2016, 8, 319. [Google Scholar] [CrossRef]
- Christakos, S.; Li, S.; De La Cruz, J.; Shroyer, N.F.; Criss, Z.K.; Verzi, M.P.; Fleet, J.C. Vitamin D and the intestine: Review and update. J. Steroid Biochem. Mol. Biol. 2020, 196, 105501. [Google Scholar] [CrossRef] [PubMed]
- Morris, H.A.; Anderson, P.H. Autocrine and paracrine actions of vitamin d. Clin. Biochem. Rev. 2010, 31, 129–138. [Google Scholar] [PubMed]
- Tashiro, K.; Abe, T.; Oue, N.; Yasui, W.; Ryoji, M. Characterization of Vitamin D-mediated induction of the CYP 24 transcription. Mol. Cell. Endocrinol. 2004, 226, 27–32. [Google Scholar] [CrossRef]
- Zhao, G.; Simpson, R.U. Membrane localization, Caveolin-3 association and rapid actions of vitamin D receptor in cardiac myocytes. Steroids 2010, 75, 555–559. [Google Scholar] [CrossRef]
- Hii, C.S.; Ferrante, A. The Non-Genomic Actions of Vitamin D. Nutrients 2016, 8, 135. [Google Scholar] [CrossRef]
- Horváth, H.C.; Lakatos, P.; Kósa, J.P.; Bácsi, K.; Borka, K.; Bises, G.; Nittke, T.; Hershberger, P.A.; Speer, G.; Kállay, E. The candidate oncogene CYP24A1: A potential biomarker for colorectal tumorigenesis. J. Histochem. Cytochem. 2010, 58, 277–285. [Google Scholar] [CrossRef]
- Clinckspoor, I.; Gérard, A.; Sande, V. The vitamin D receptor in thyroid development and function. Eur. Thyroid. J. 2012, 1, 168–175. [Google Scholar] [CrossRef]
- Danza, K.; Porcelli, L.; De Summa, S.; Di Fonte, R.; Pilato, B.; Lacalamita, R.; Serratì, S.; Azzariti, A.; Tommasi, S. The ERRα-VDR axis promotes calcitriol degradation and estrogen signaling in breast cancer cells, while VDR-CYP24A1-ERRα overexpression correlates with poor prognosis in patients with basal-like breast cancer. Mol. Oncol. 2021. [Google Scholar] [CrossRef]
- Clinckspoor, I.; Verlinden, L.; Mathieu, C.; Bouillon, R.; Verstuyf, A.; Decallonne, B. Vitamin D in thyroid tumorigenesis and development. Prog. Histochem. Cytochem. 2013, 48, 65–98. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Tirabassi, G.; Bizzaro, G.; Orio, F.; Paschou, S.A.; Vryonidou, A.; Balercia, G.; Shoenfeld, Y.; Colao, A. Vitamin D and thyroid disease: To D or not to D? Eur. J. Clin. Nutr. 2015, 69, 291–296. [Google Scholar] [CrossRef]
- Wang, S.; Wu, Y.; Zuo, Z.; Zhao, Y.; Wang, K. The effect of vitamin D supplementation on thyroid autoantibody levels in the treatment of autoimmune thyroiditis: A systematic review and a meta-analysis. Endocrine 2018, 59, 499–505. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, W.; Ma, C.; Zhao, Y.; Xiong, R.; Wang, H.; Chen, W.; Zheng, S.G. Immunomodulatory Function of Vitamin D and Its Role in Autoimmune Thyroid Disease. Front. Immunol. 2021, 12, 352. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.G.; Wakeley, S.E.; Hamel, F.G.; High, R.R.; Korch, C.; Goldner, W.S. Gene expression of vitamin d metabolic enzymes at baseline and in response to vitamin d treatment in thyroid cancer cell lines. Oncol. 2012, 83, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Yavropoulou, M.P.; Panagiotou, G.; Topouridou, K.; Karayannopoulou, G.; Koletsa, T.; Zarampoukas, T.; Goropoulos, A.; Chatzaki, E.; Yovos, J.G.; Pazaitou-Panayiotou, K. Vitamin D receptor and progesterone receptor protein and gene expression in papillary thyroid carcinomas: Associations with histological features. J. Endocrinol. Investig. 2017, 40, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.P.; Liane, K.M.; Bjorhovde, S.B.; Bjoro, T.; Torjesen, P.A.; Haug, E. Vitamin D Receptor Binding and Biological Effects of Cholecalciferol Analogues in Rat Thyroid Cells. J. Steroid Biochem. Mol. Biol. 1994, 50, 145–150. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D and immune function: Understanding common pathways. Curr. Osteoporos. Rep. 2009, 7, 58–63. [Google Scholar] [CrossRef]
- Vieira, I.H.; Rodrigues, D.; Paiva, I. Vitamin D and Autoimmune Thyroid Disease-Cause, Consequence, or a Vicious Cycle? Nutrients 2020, 12, 2791. [Google Scholar] [CrossRef]
- Williams, G.R.; Bassett, J.H.D. Thyroid diseases and bone health. J. Endocrinol. Investig. 2018, 41, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Blouin, S.; Libouban, H.; Moreau, M.F.; Chappard, D. Orchidectomy models of osteoporosis. Methods Mol. Biol. 2008, 455, 125–134. [Google Scholar] [CrossRef]
- Filipović, B.; Šošić-Jurjević, B.; Ajdžanović, V.; Trifunović, S.; Manojlović-Stojanoski, M.; Ristić, N.; Nestorović, N.; Milošević, V.; Sekulić, M. The effect of orchidectomy on thyroid C cells and bone histomorphometry in middle-aged rats. Histochem. Cell Biol. 2007, 128, 153–159. [Google Scholar] [CrossRef]
- Gunness, M.; Orwoll, E. Early induction of alterations in cancellous and cortical bone histology after orchiectomy in mature rats. J. Bone Miner. Res. 1995, 10, 1735–1744. [Google Scholar] [CrossRef]
- Libouban, H.; Moreau, M.F.; Legrand, E.; Audran, M.; Baslé, M.F.; Chappard, D. Comparison of histomorphometric descriptors of bone architecture with dual-energy X-ray absorptiometry for assessing bone loss in the orchidectomized rat. Osteoporos. Int. 2002, 13, 422–428. [Google Scholar] [CrossRef]
- Erben, R.G.; Eberle, J.; Stahr, K.; Goldberg, M. Androgen deficiency induces high turnover osteopenia in aged male rats: A sequential histomorphometric study. J. Bone Miner. Res. 2000, 15, 1085–1098. [Google Scholar] [CrossRef]
- Sošić-Jurjević, B.; Filipović, B.; Renko, K.; Ajdžanović, V.; Manojlović-Stojanoski, M.; Milošević, V.; Köhrle, J. Orchidectomy of middle-aged rats decreases liver deiodinase 1 and pituitary deiodinase 2 activity. J. Endocrinol. 2012, 215, 247–256. [Google Scholar] [CrossRef][Green Version]
- Živanović, J.; Jarić, I.; Ajdžanović, V.; Mojić, M.; Miler, M.; Šošić-Jurjević, B.; Milošević, V.; Filipović, B. Daidzein upregulates anti-aging protein Klotho and NaPi 2a cotransporter in a rat model of the andropause. Ann. Anat. 2019, 221, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Ajdžanović, V.; Šošić-Jurjević, B.; Filipović, B.; Trifunović, S.; Manojlović-Stojanoski, M.; Sekulić, M.; Milošević, V. Genistein-induced histomorphometric and hormone secreting changes in the adrenal cortex in middle-aged rats. Exp. Biol. Med. 2009, 234, 148–156. [Google Scholar] [CrossRef]
- Šošić-Jurjevic, B.; Filipović, B.; Renko, K.; Miler, M.; Trifunović, S.; Ajdžanovič, V.; Köhrle, J.; Milošević, V. Testosterone and estradiol treatments differently affect pituitary-thyroid axis and liver deiodinase 1 activity in orchidectomized middle-aged rats. Exp. Gerontol. 2015, 72, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Bjelobaba, I.; Yan, Z.; Kucka, M.; Tomić, M.; Stojilkovic, S.S. Expression and roles of pannexins in ATP release in the pituitary gland. Endocrinology 2011, 152, 2342–2352. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.R.; Aspray, T.J. The role of vitamin D in maintaining bone health in older people. Ther. Adv. Musculoskelet. Dis. 2017, 9, 89–95. [Google Scholar] [CrossRef]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Evaluation, treatment, and prevention of vitamin D deficiency: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Shepard, R.M.; Deluca, H.F. Plasma concentrations of vitamin D3 and its metabolites in the rat as influenced by vitamin D3 or 25-hydroxyvitamin D3 intakes. Arch. Biochem. Biophys. 1980, 202, 43–53. [Google Scholar] [CrossRef]
- Zhu, J.G.; Ochalek, J.T.; Kaufmann, M.; Jones, G.; DeLuca, H.F. CYP2R1 is a major, but not exclusive, contributor to 25-hydroxyvitamin D production in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 15650–15655. [Google Scholar] [CrossRef]
- Sagartz, J.E.; Ozaki, A.; Capen, C.C. Phagocytosis of fluorescent beads by rat thyroid follicular cells (FRTL-5): Comparison with iodide trapping as an index of functional activity of thyrocytes in vitro. Toxicol. Pathol. 1995, 23, 635–643. [Google Scholar] [CrossRef]
- Schmidt, F.; Wolf, R.; Baumann, L.; Braunbeck, T. Ultrastructural Alterations in Thyrocytes of Zebrafish (Danio rerio) after Exposure to Propylthiouracil and Perchlorate. Toxicol. Pathol. 2017, 45, 649–662. [Google Scholar] [CrossRef]
- Gérard, A.C.; Denef, J.F.; Many, M.C.; Gathy, P.; de Burbure, C.; van den Hove, M.F.; Coppée, F.; Ledent, C.; Colin, I.M. Relationships between cell division, expression of growth factors and microcirculation in the thyroids of Tg-A2aR transgenic mice and patients with Graves’ disease. J. Endocrinol. 2003, 177, 269–277. [Google Scholar] [CrossRef][Green Version]
- Lamberg-Allardt, C.; Valtonen, E. 1,25-Dihydroxycholecalciferol attenuates thyrotropin stimulated iodide accumulation in rat thyroid follicular FRTL-5 cells by reducing iodide porter number. Biochem. Biophys. Res. Commun. 1992, 182, 1435–1439. [Google Scholar] [CrossRef]
- Berg, J.P.; Torjesen, P.A.; Haug1, E. Dihydroxyvitamin D3 Attenuates TSH and 8-(4-Chlorophenylthio)-cAMP-Stimulated Growth and Iodide Uptake by Rat Thyroid Cells (FRTL-5). Thyroid 1993, 3, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.; Eastman, C. Goitre—Causes, investigation and management. Aust. Fam. Physician 2012, 41, 573–576. [Google Scholar]
- Balla, B.; Tobiás, B.; Kósa, J.P.; Podani, J.; Horváth, P.; Nagy, Z.; Horányi, J.; Járay, B.; Székely, E.; Krenács, L.; et al. Vitamin D-neutralizing CYP24A1 expression, oncogenic mutation states and histological findings of human papillary thyroid cancer. J. Endocrinol. Investig. 2015, 38, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Baier, R.; Grauer, A.; Lazaretti-Castrot, M.; Ziegler, R.; Raue, F. Differential effects of 1,25-dihydroxyvitamin D3 on cell proliferation and calcitonin gene expression. Endocrinology 1994, 135, 2006–2011. [Google Scholar] [CrossRef] [PubMed]
- Felsenfeld, A.J.; Levine, B.S. Calcitonin, the forgotten hormone: Does it deserve to be forgotten? Clin. Kidney J. 2015, 8, 180–187. [Google Scholar] [CrossRef]
- Berg, J.P.; Haug, E. Vitamin D: A Hormonal Regulator of the cAMP Signaling Pathway. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Ongphiphadhanakul, B.; Ebner, S.A.; Fang, S.L.; Lombardi, A.; Baran, D.T.; Braverman, L.E. 1,25-Dihydroxycholecalciferol modulates 3H-thymidine incorporation in FRTL5 cells. J. Cell. Biochem. 1992, 49, 304–309. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Manufacturer | Cat. Number | Origin | Dilution |
|---|---|---|---|---|
| TPO | Santa Cruz, Italy | sc-376876 | Mouse | 1:400 |
| NIS | Acris, Germany | EUD4101 | Rabbit | 1:600 |
| Tg | Dako, Denmark | A0251 | Rabbit | 1:500 |
| CYP24A1 | Santa Cruz Biotech Inc., Italy | sc-66851 | Rabbit | 1:100 |
| VDR | Abcam, UK | Ab 3508 | Rabbit | 1:1000 |
| CT | Dako, Denmark | A576 | Rabbit | 1:300 |
| Anti-mouse, HRP labeled | Abcam, UK | Ab6820 | Donkey | 1:200 |
| Anti-rabbit, HRP labeled | Dako, Denmark | P0399 | Swine | 1:200 |
| Group | SO | Orx | Orx + Vit. D3 | Statistical Significance |
|---|---|---|---|---|
| Absolute thyroid weight (mg) | 40.00 ± 7.69 | 44.33 ± 5.43 | 41.00 ± 5.90 | n.s. |
| Relative thyroid weight | 0.049 ± 0.007 | 0.059 ± 0.009 | 0.063 ± 0.01 | n.s. vs. Orx p = 0.08 vs. SO |
| Body mass (g) | 683.3 ± 52.8 | 675.0 ± 63.5 | 665.0 ± 68.9 | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šošić-Jurjević, B.; Trifunović, S.; Živanović, J.; Ajdžanović, V.; Miler, M.; Ristić, N.; Filipović, B. Vitamin D3 Treatment Alters Thyroid Functional Morphology in Orchidectomized Rat Model of Osteoporosis. Int. J. Mol. Sci. 2022, 23, 791. https://doi.org/10.3390/ijms23020791
Šošić-Jurjević B, Trifunović S, Živanović J, Ajdžanović V, Miler M, Ristić N, Filipović B. Vitamin D3 Treatment Alters Thyroid Functional Morphology in Orchidectomized Rat Model of Osteoporosis. International Journal of Molecular Sciences. 2022; 23(2):791. https://doi.org/10.3390/ijms23020791
Chicago/Turabian StyleŠošić-Jurjević, Branka, Svetlana Trifunović, Jasmina Živanović, Vladimir Ajdžanović, Marko Miler, Nataša Ristić, and Branko Filipović. 2022. "Vitamin D3 Treatment Alters Thyroid Functional Morphology in Orchidectomized Rat Model of Osteoporosis" International Journal of Molecular Sciences 23, no. 2: 791. https://doi.org/10.3390/ijms23020791
APA StyleŠošić-Jurjević, B., Trifunović, S., Živanović, J., Ajdžanović, V., Miler, M., Ristić, N., & Filipović, B. (2022). Vitamin D3 Treatment Alters Thyroid Functional Morphology in Orchidectomized Rat Model of Osteoporosis. International Journal of Molecular Sciences, 23(2), 791. https://doi.org/10.3390/ijms23020791