
Hippocampal Expression of Cytochrome P450 1B1 in Penetrating Traumatic Brain Injury


Abstract
:1. Introduction
2. Results
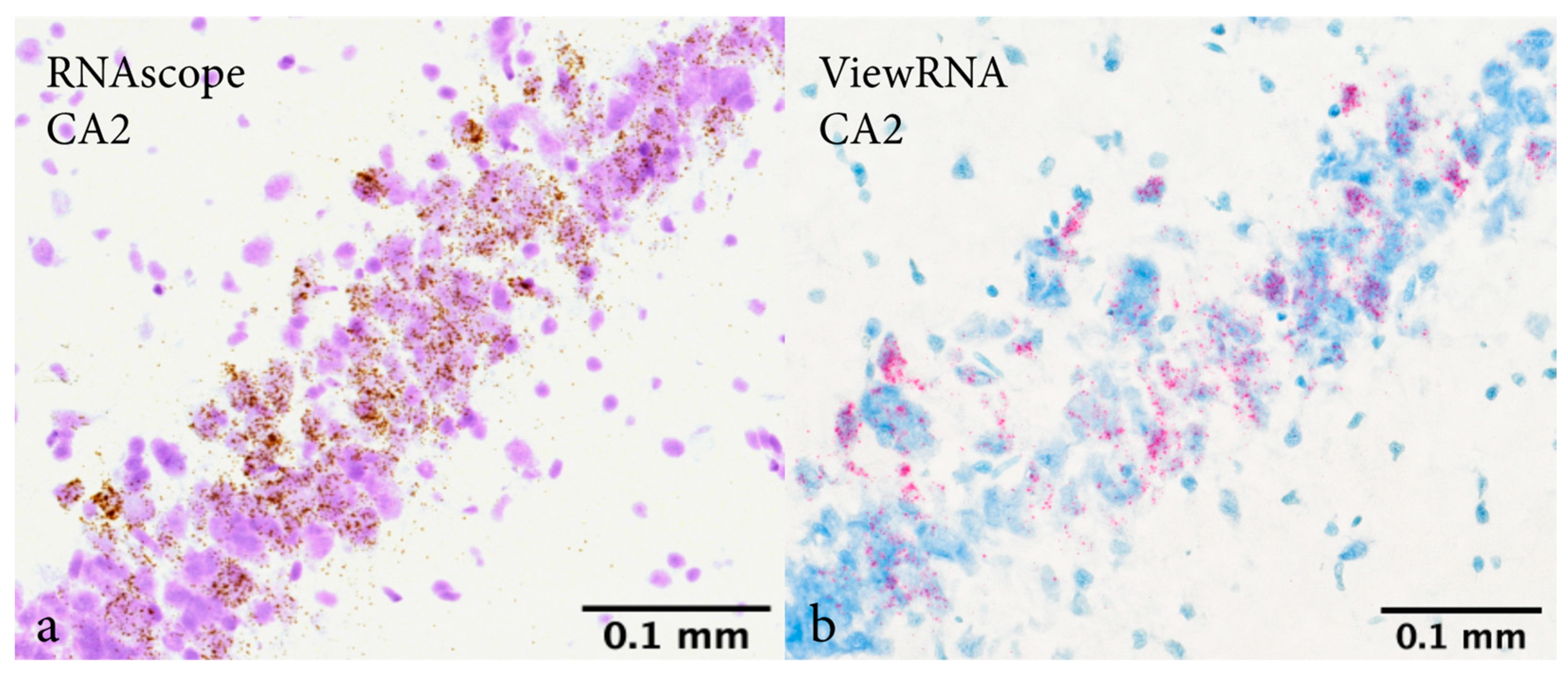
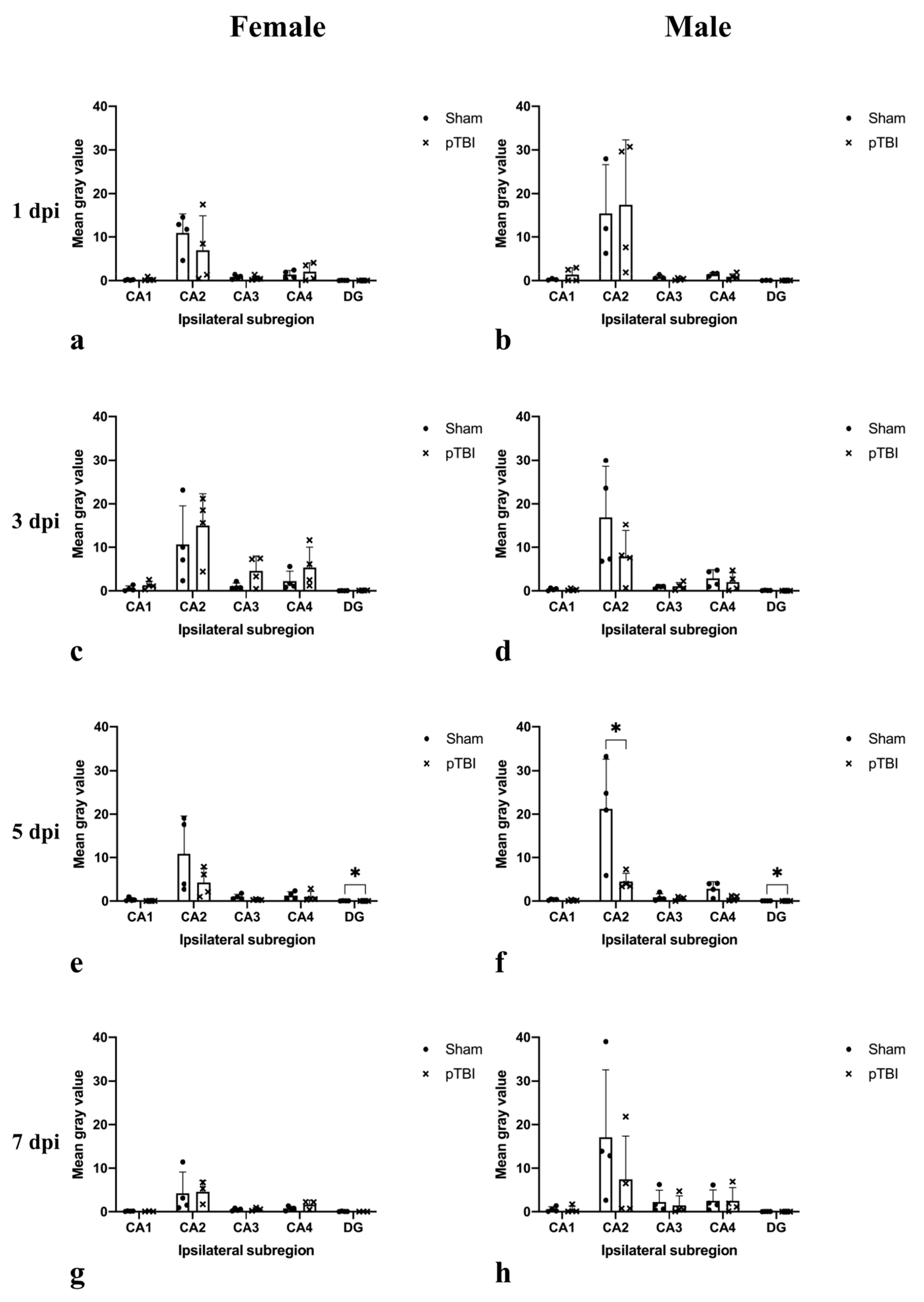
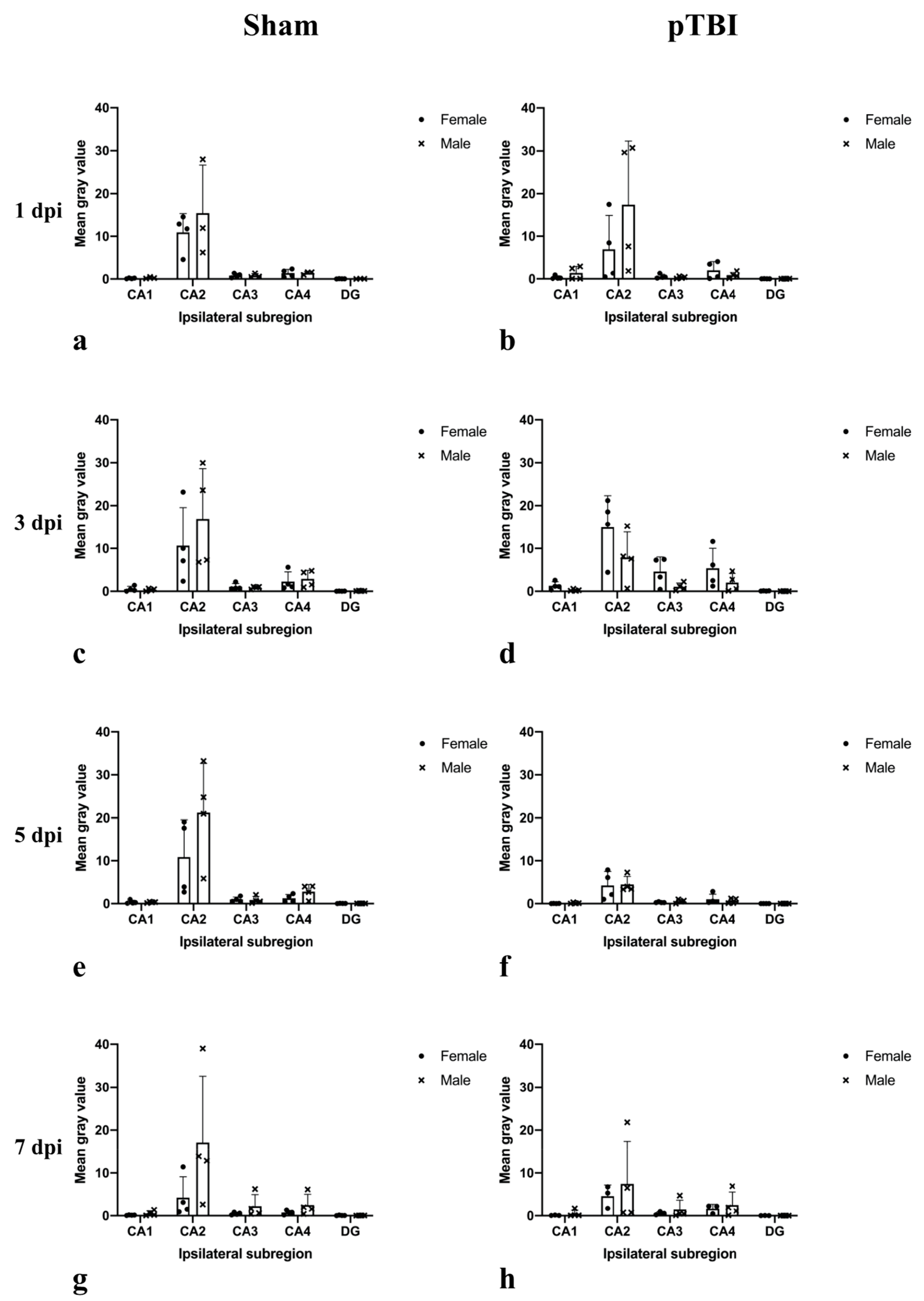
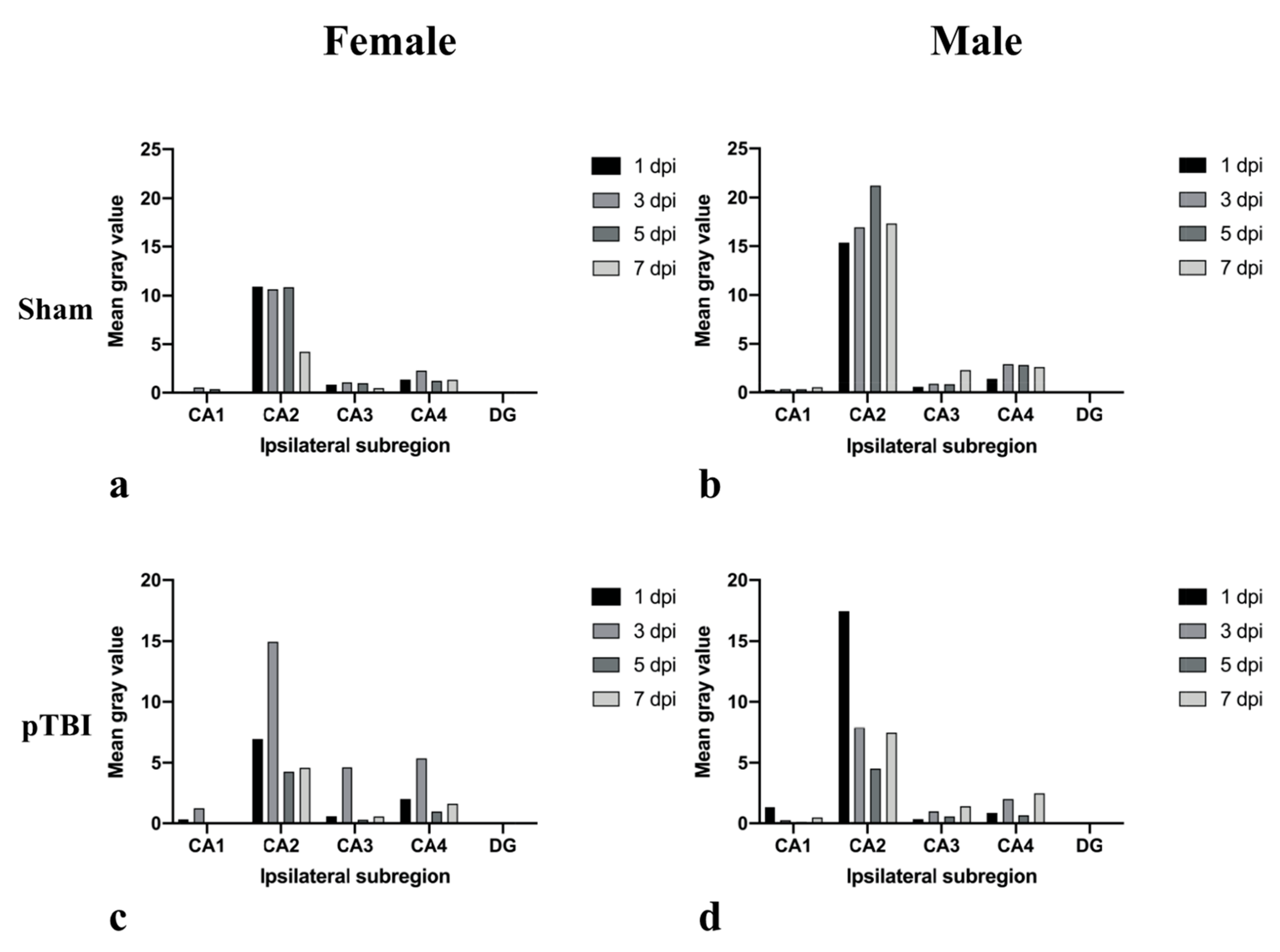
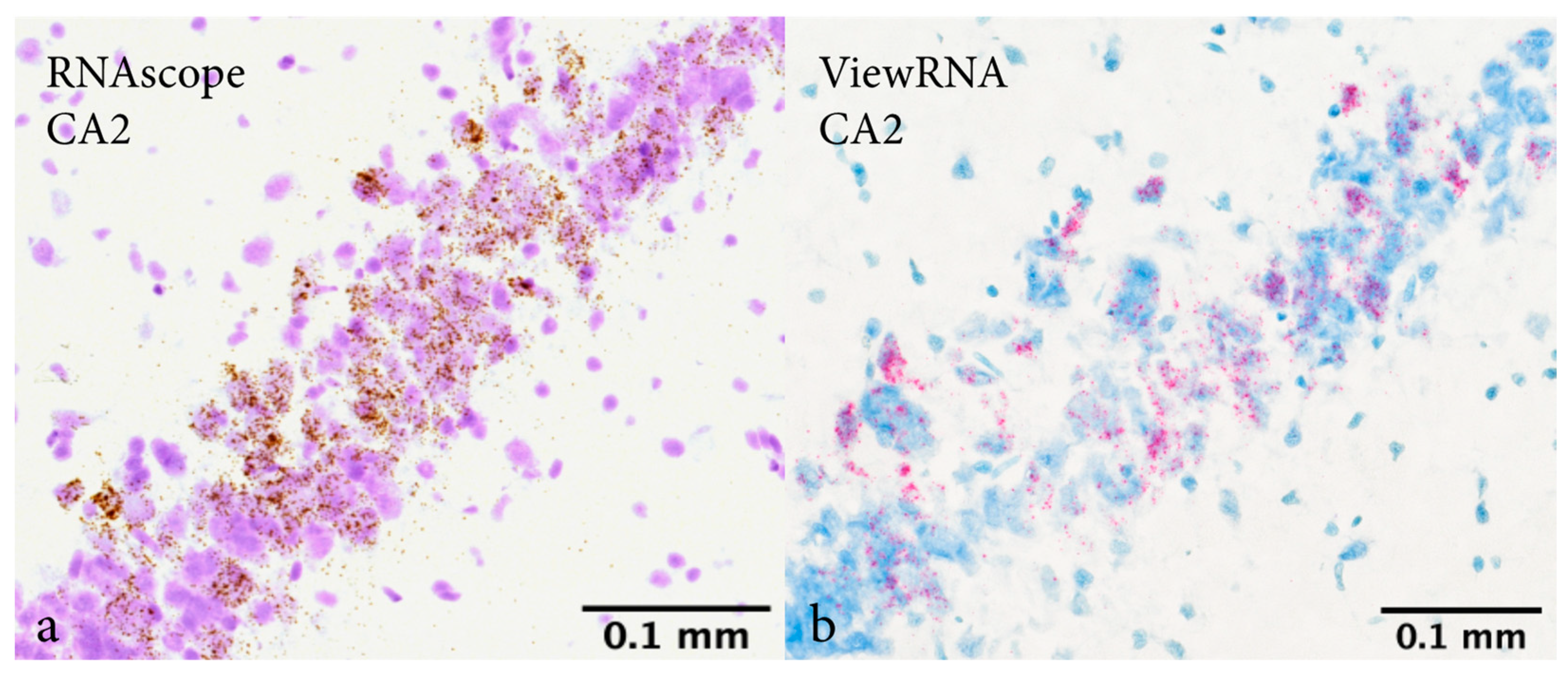
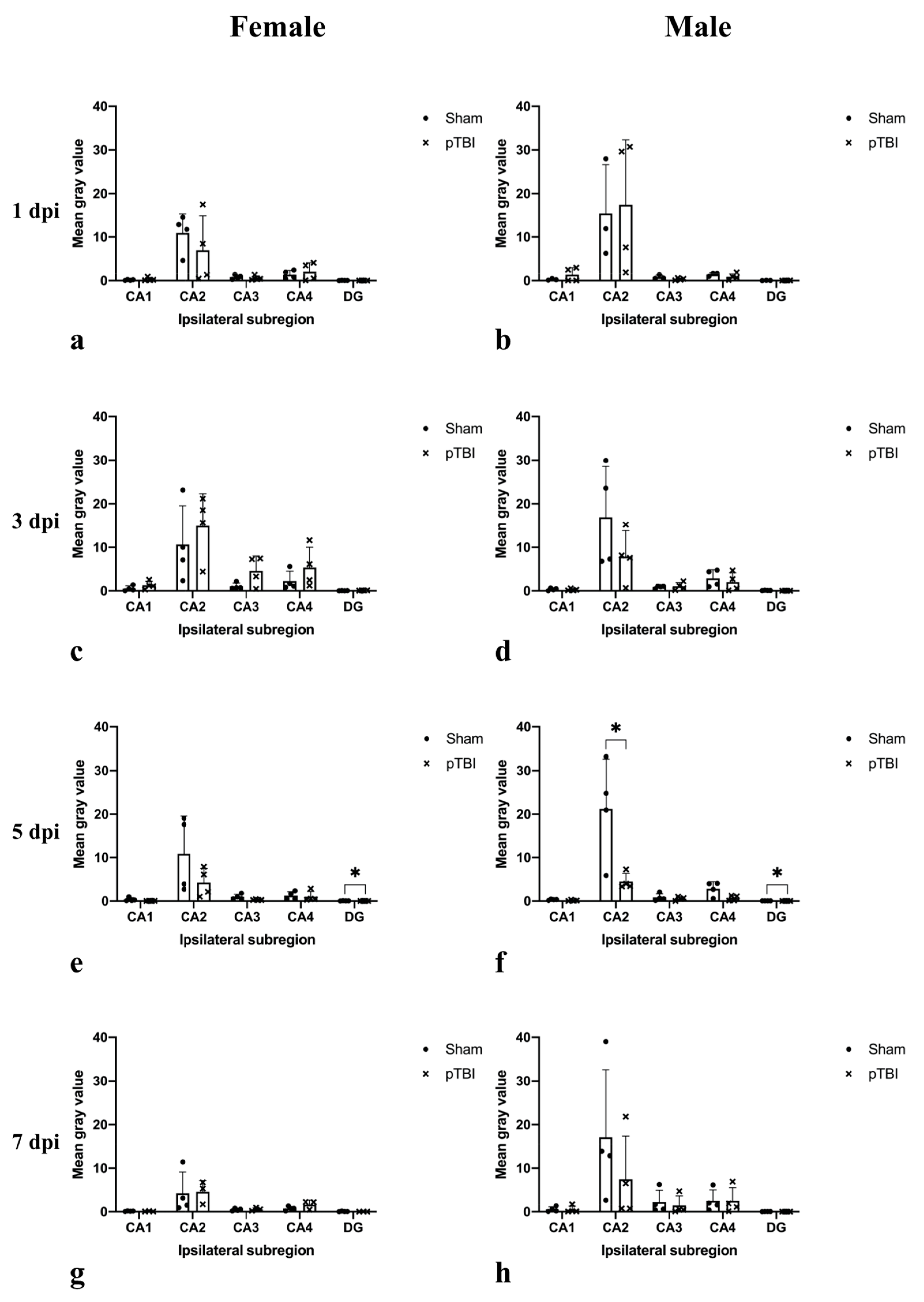
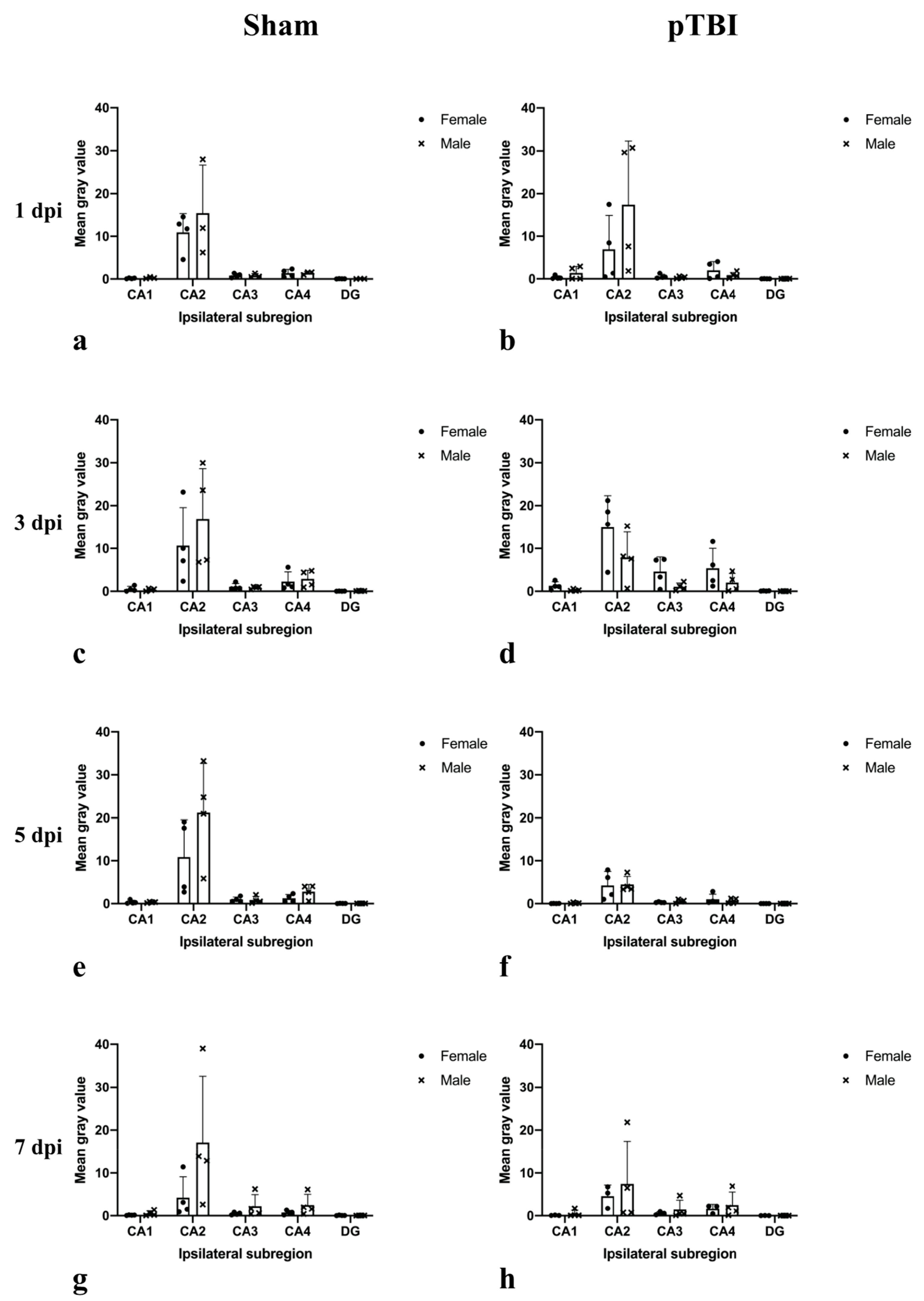
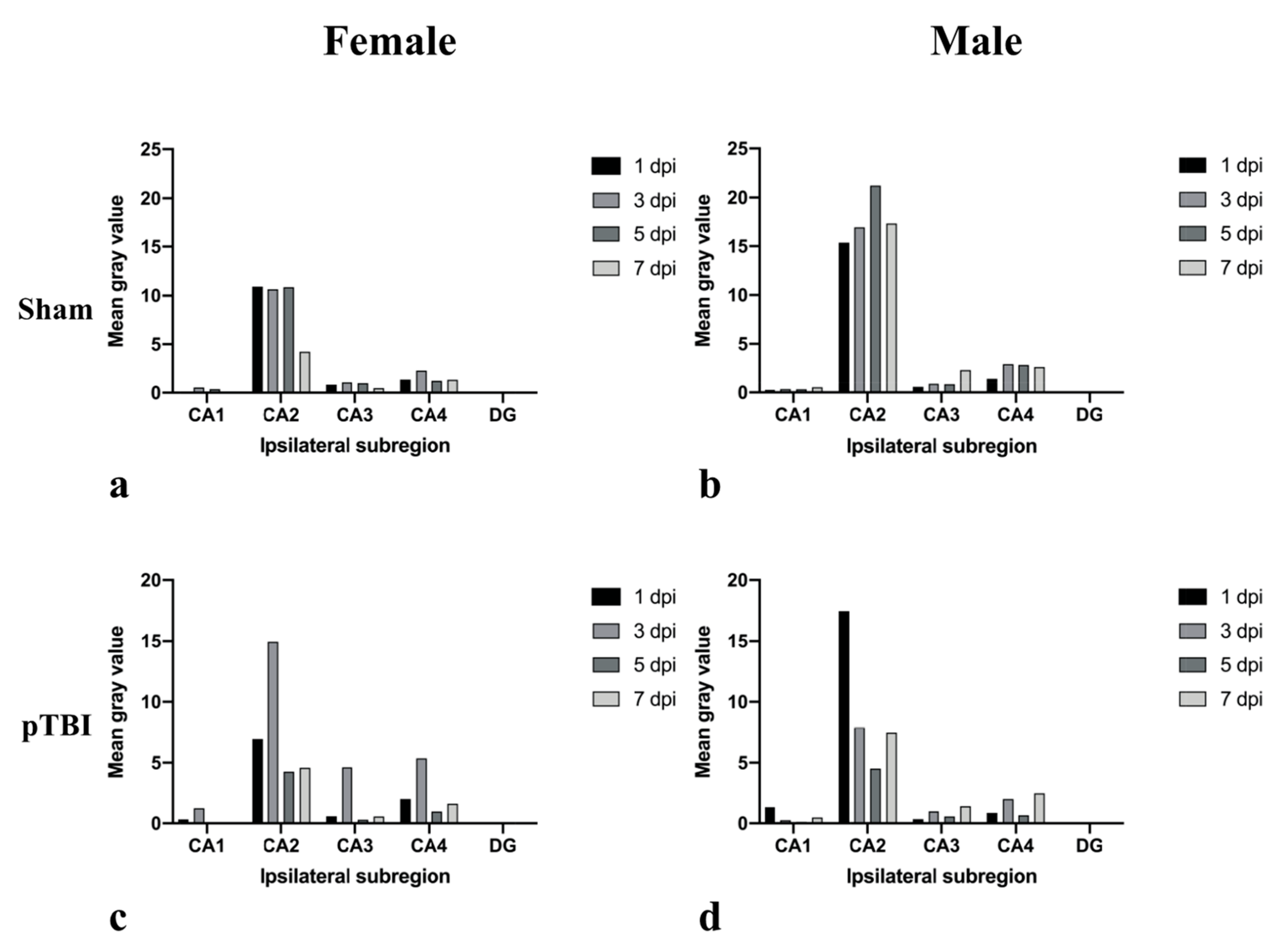
2.1. In Situ Hybridization
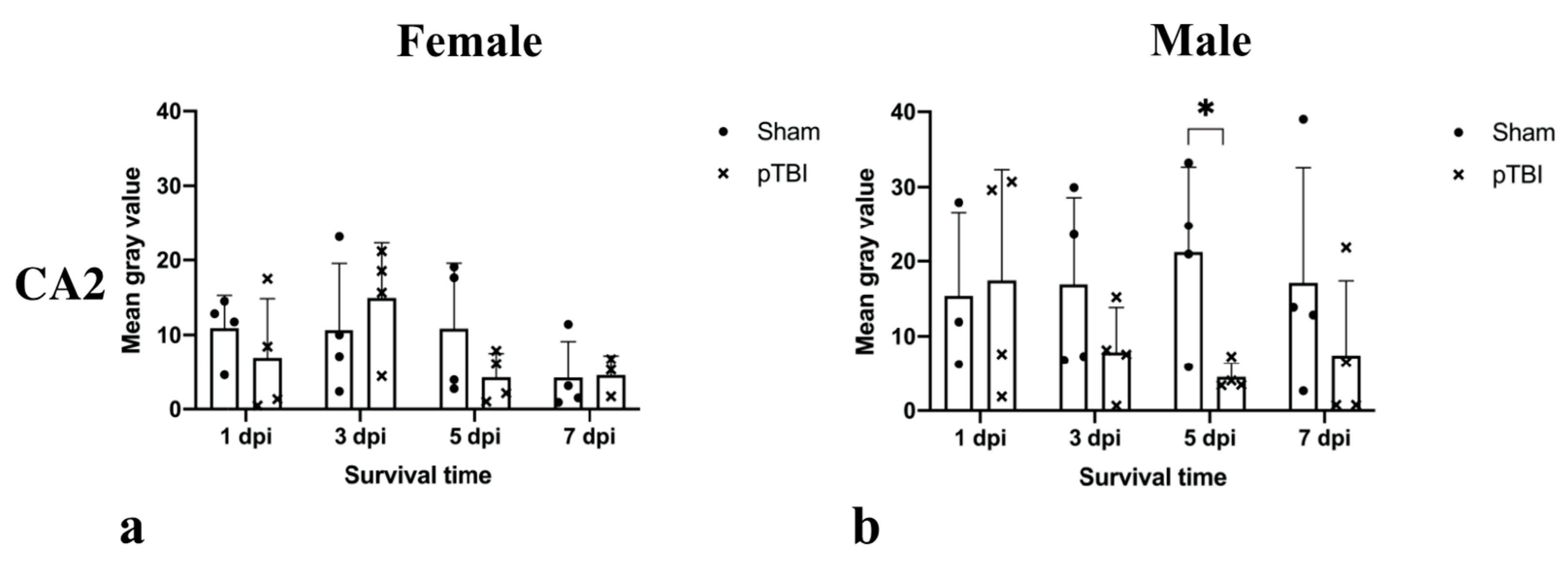
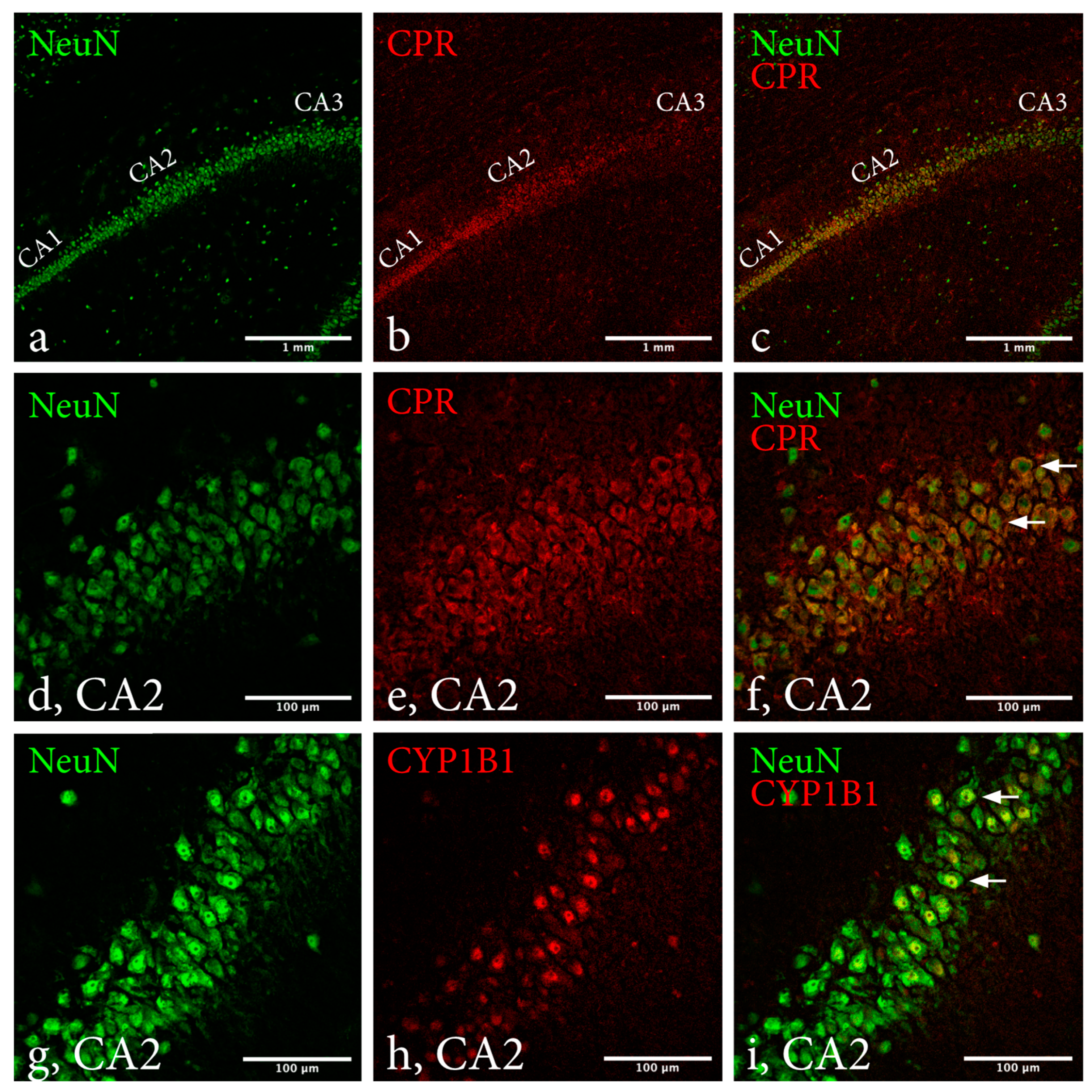
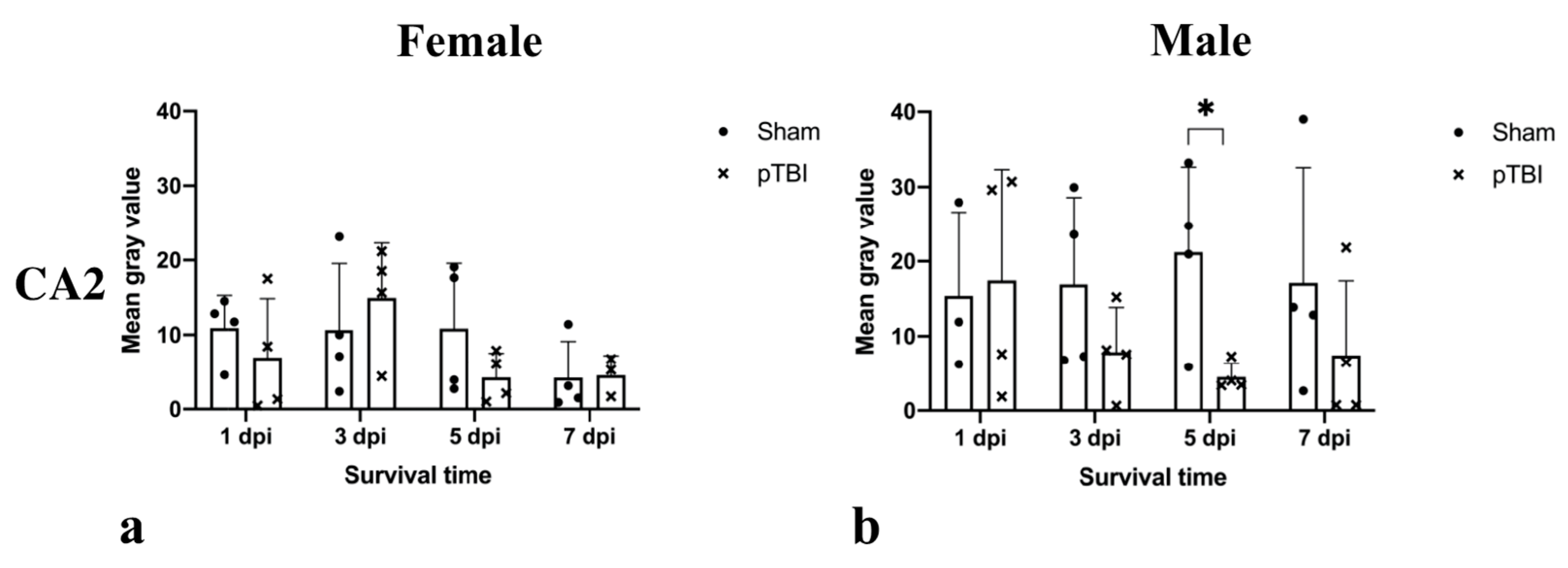
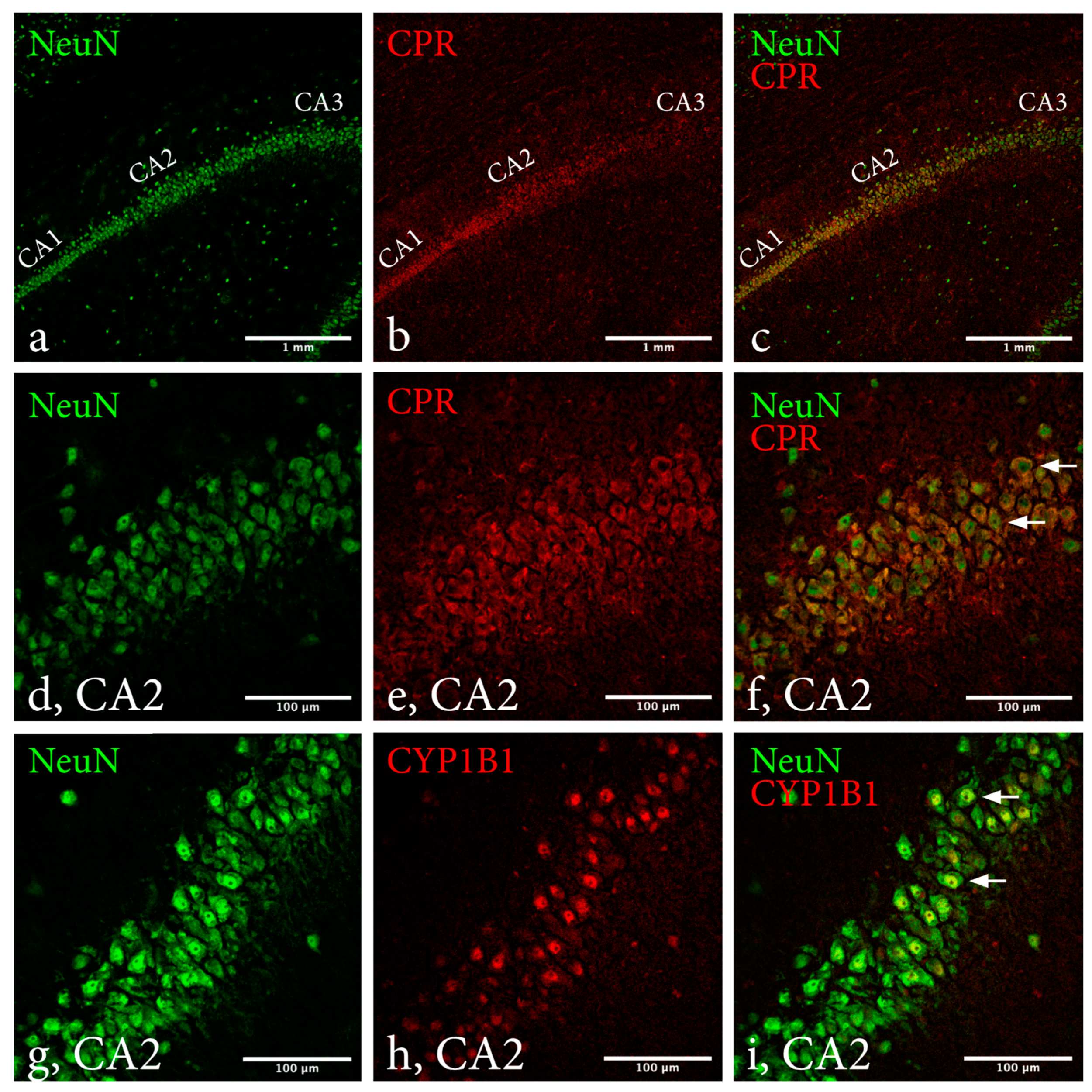
2.2. Immunohistochemistry
2.3. Estrus Cycle Staging
3. Discussion
4. Materials and Methods
4.1. Material
4.2. Statistics
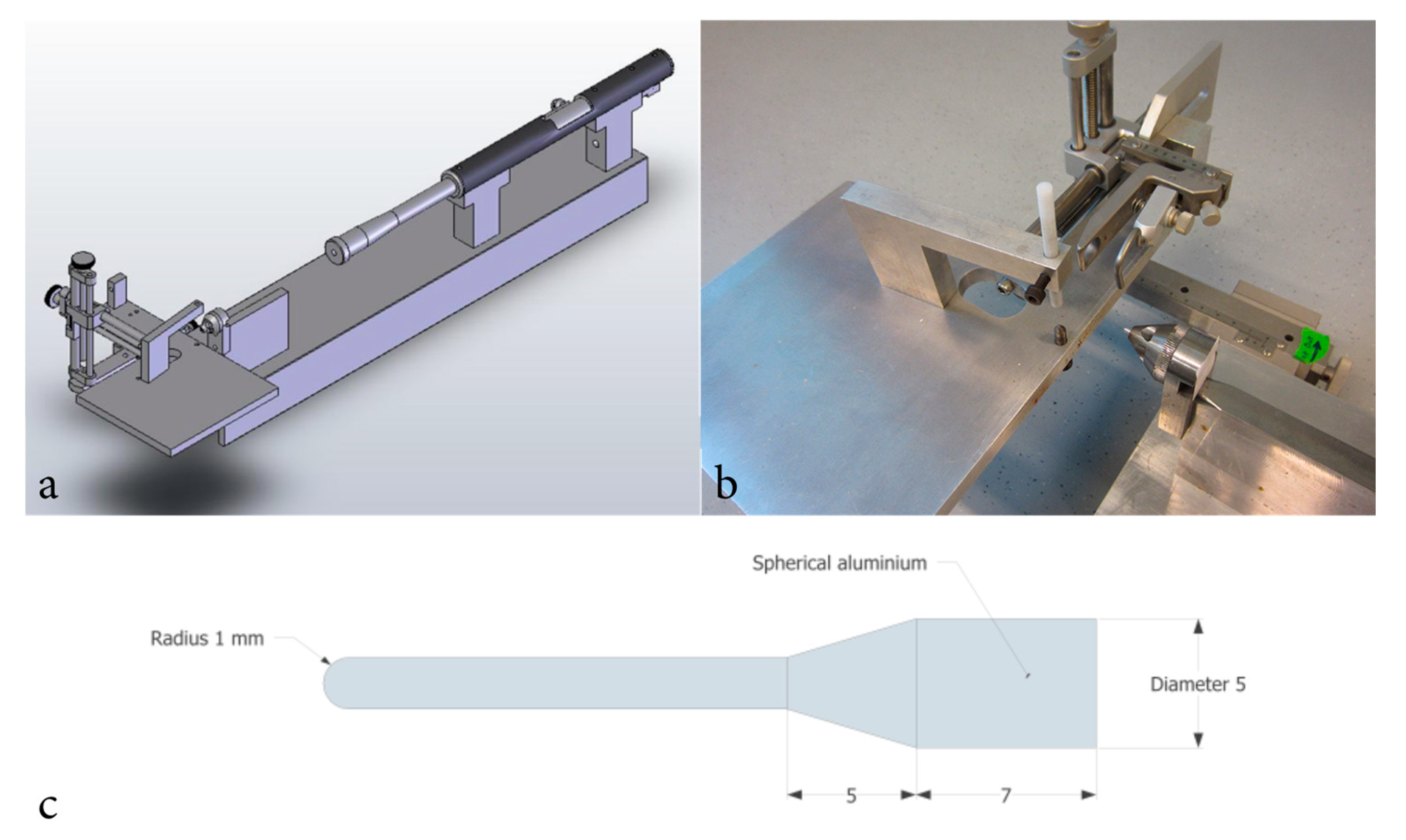
4.3. Surgery and Penetrating Trauma
4.4. Tissue Sectioning
4.5. In-Situ Hybridization
4.6. Estrus Cycle Determination
4.7. Immunohistochemistry
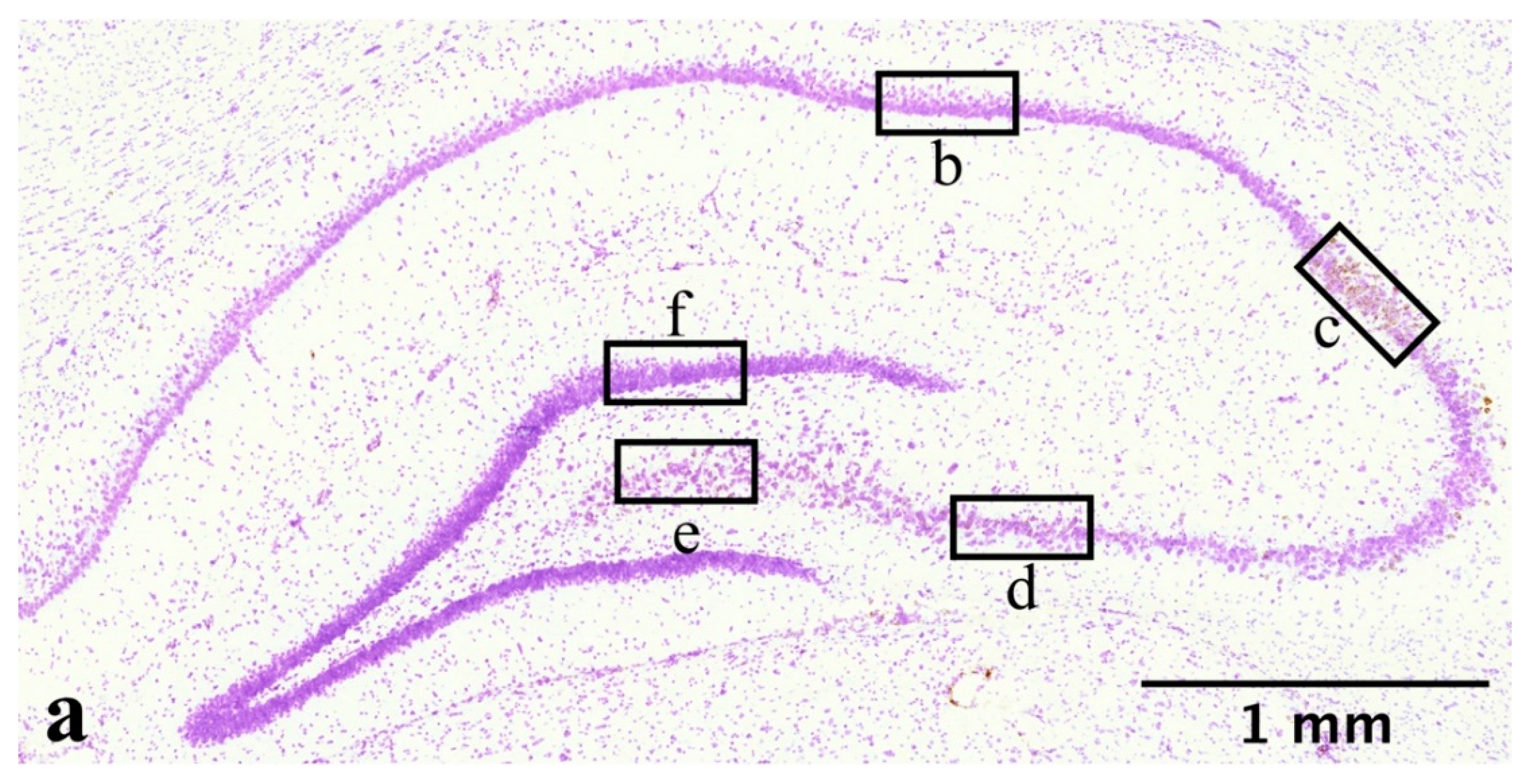
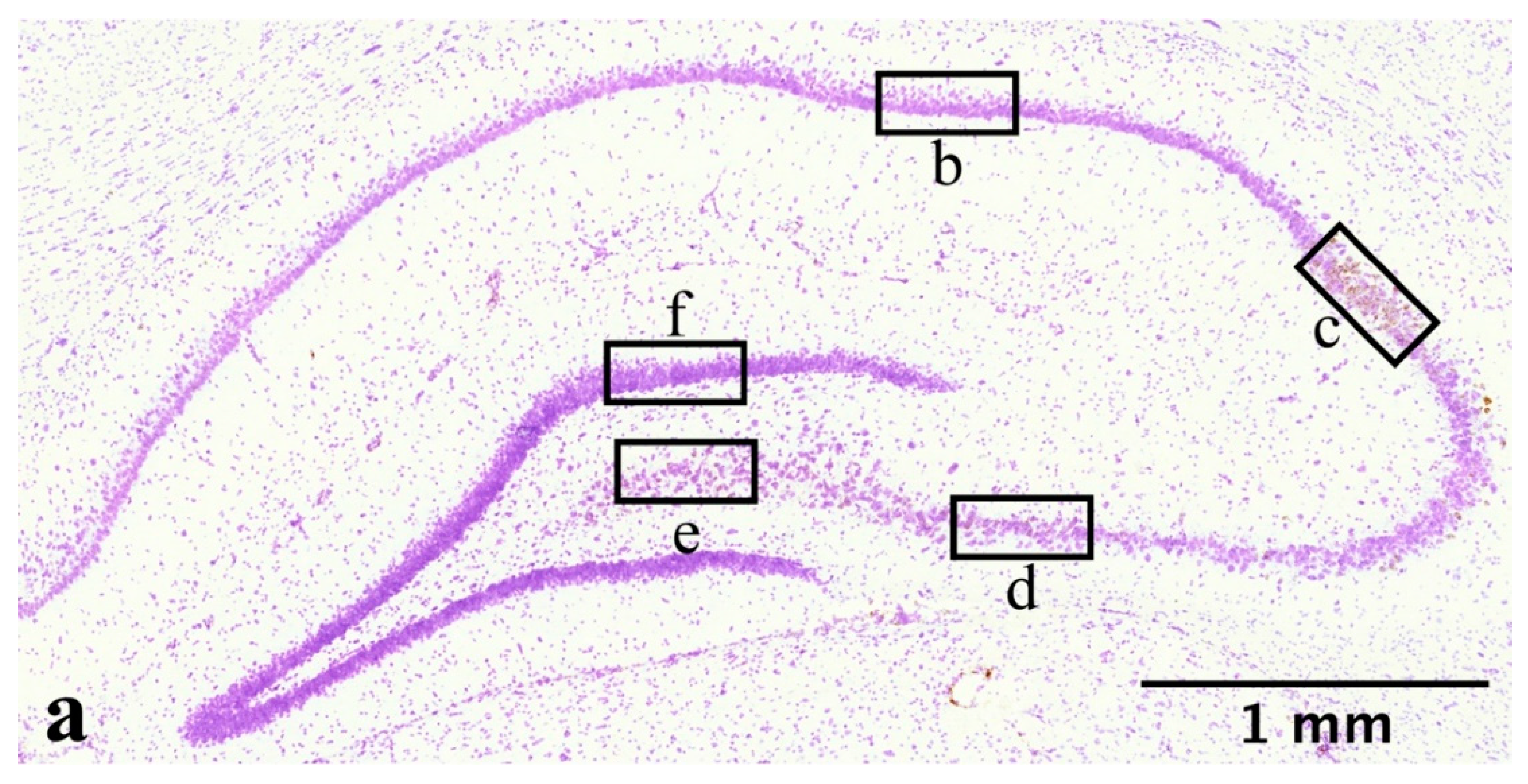
4.8. Image Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| BBB | blood-brain-barrier |
| CA | Cornu Ammonis |
| CPR | Cytochrome P450 reductase |
| CYP | Cytochrome P450 |
| CYP1B1 | Cytochrome P450 1B1 |
| DG | Dentate Gyrus |
| dpi | days post-injury |
| E2 | 17β-estradiol |
| HETEs | hydroxyeicosatetraenoic acids |
| IHC | immunohistochemistry |
| ISH | in situ hybridization |
| pTBI | penetrating traumatic brain injury |
| ROI | region of interest |
| TBI | traumatic brain injury |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CYP1A1 | CYP1A2 | CYP1B1 | CYP2A1 | CYP2A2 | CYP2E1 | CPR | |
|---|---|---|---|---|---|---|---|
| Hippocampus pTBI vs. Sham Fold change | 1.30 | 1.16 | 15.03 | 1.11 | 1.10 | 1.17 | 1.99 |
| Cortex pTBI vs. Sham Fold change | 1.08 | 1.04 | 4.65 | 1.08 | −1.18 | 1.08 | −1.63 |
References
- Sheriff, F.G.; Hinson, H.E. Pathophysiology and Clinical Management of Moderate and Severe Traumatic Brain Injury in the ICU. Semin. Neurol. 2015, 35, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Stein, D.G. Embracing failure: What the Phase III progesterone studies can teach about TBI clinical trials. Brain Inj. 2015, 29, 1259–1272. [Google Scholar] [CrossRef]
- Risling, M.; Plantman, S.; Angeria, M.; Rostami, E.; Bellander, B.-M.; Kirkegaard, M.; Arborelius, U.; Davidsson, J. Mechanisms of blast induced brain injuries, experimental studies in rats. NeuroImage 2011, 54 (Suppl. 1), S89–S97. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhu, W.; Gonzalez, F.J. Potential role of CYP1B1 in the development and treatment of metabolic diseases. Pharmacol. Ther. 2017, 178, 18–30. [Google Scholar] [CrossRef]
- Palenski, T.L.; Gurel, Z.; Sorenson, C.M.; Hankenson, K.D.; Sheibani, N. Cyp1B1 expression promotes angiogenesis by suppressing NF-κB activity. Am. J. Physiol.-Cell Physiol. 2013, 305, C1170–C1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maayah, Z.H.; Althurwi, H.N.; El-Sherbeni, A.A.; Abdelhamid, G.; Siraki, A.G.; El-Kadi, A.O. The role of cytochrome P450 1B1 and its associated mid-chain hydroxyeicosatetraenoic acid metabolites in the development of cardiac hypertrophy induced by isoproterenol. Mol. Cell. Biochem. 2017, 429, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Farias, S.E.; Basselin, M.; Chang, L.; Heidenreich, K.A.; Rapoport, S.I.; Murphy, R.C. Formation of eicosanoids, E2/D2 isoprostanes, and docosanoids following decapitation-induced ischemia, measured in high-energy-microwaved rat brain. J. Lipid Res. 2008, 49, 1990–2000. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, Y.; Nakajima, M.; Yokoi, T. Cytochrome P450-mediated metabolism of estrogens and its regulation in human. Cancer Lett. 2005, 227, 115–124. [Google Scholar] [CrossRef]
- Day, N.L.; Floyd, C.L.; D’Alessandro, T.L.; Hubbard, W.J.; Chaudry, I.H. 17β-Estradiol Confers Protection after Traumatic Brain Injury in the Rat and Involves Activation of G Protein-Coupled Estrogen Receptor. J. Neurotrauma 2013, 30, 1531–1541. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Ma, K.; Jin, L.; Zhu, H.; Cao, R. 17β-estradiol rescues damages following traumatic brain injury from molecule to behavior in mice. J. Cell. Physiol. 2018, 233, 1712–1722. [Google Scholar] [CrossRef]
- Wang, J.; Hou, Y.; Zhang, L.; Liu, M.; Zhao, J.; Zhang, Z.; Ma, Y.; Hou, W. Estrogen Attenuates Traumatic Brain Injury by Inhibiting the Activation of Microglia and Astrocyte-Mediated Neuroinflammatory Responses. Mol. Neurobiol. 2021, 58, 1052–1061. [Google Scholar] [CrossRef]
- Guennoun, R. Progesterone in the Brain: Hormone, Neurosteroid and Neuroprotectant. Int. J. Mol. Sci. 2020, 21, 5271. [Google Scholar] [CrossRef] [PubMed]
- Silva-Alves, M.S.; Secolin, R.; Carvalho, B.S.; Yasuda, C.L.; Bilevicius, E.; Alvim, M.K.M.; Santos, R.O.; Maurer-Morelli, C.V.; Cendes, F.; Lopes-Cendes, I. A Prediction Algorithm for Drug Response in Patients with Mesial Temporal Lobe Epilepsy Based on Clinical and Genetic Information. PLoS ONE 2017, 12, e0169214. [Google Scholar] [CrossRef] [PubMed]
- Renic, M.; Klaus, J.A.; Omura, T.; Kawashima, N.; Onishi, M.; Miyata, N.; Koehler, R.C.; Harder, D.R.; Roman, R.J. Effect of 20-HETE Inhibition on Infarct Volume and Cerebral Blood Flow after Transient Middle Cerebral Artery Occlusion. J. Cereb. Blood Flow Metab. 2009, 29, 629–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Weng, Y.; Zhang, Q.Y.; Cui, H.; Behr, M.; Wu, L.; Yang, W.; Zhang, L.; Ding, X. Liver-specific deletion of the NADPH-cytochrome P450 reductase gene: Impact on plasma cholesterol homeostasis and the function and regulation of microsomal cytochrome P450 and heme oxygenase. J. Biol. Chem. 2003, 278, 25895–25901. [Google Scholar] [CrossRef] [Green Version]
- Henderson, C.J.; Pass, G.J.; Wolf, C.R. The hepatic cytochrome P450 reductase null mouse as a tool to identify a successful candidate entity. Toxicol. Lett. 2006, 162, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Günther, M.; Plantman, S.; Davidsson, J.; Angéria, M.; Mathiesen, T.; Risling, M. COX-2 regulation and TUNEL-positive cell death differ between genders in the secondary inflammatory response following experimental penetrating focal brain injury in rats. Acta Neurochir. 2015, 157, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Roof, R.L.; Duvdevani, R.; Stein, D.G. Gender influences outcome of brain injury: Progesterone plays a protective role. Brain Res. 1993, 607, 333–336. [Google Scholar] [CrossRef]
- Jones, N.C.; Constantin, D.; Prior, M.J.W.; Morris, P.G.; Marsden, C.A.; Murphy, S. The neuroprotective effect of progesterone after traumatic brain injury in male mice is independent of both the inflammatory response and growth factor expression. Eur. J. Neurosci. 2005, 21, 1547–1554. [Google Scholar] [CrossRef]
- Ottochian, M.; Salim, A.; Berry, C.; Chan, L.S.; Wilson, M.T.; Margulies, D.R. Severe traumatic brain injury: Is there a gender difference in mortality? Am. J. Surg. 2009, 197, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Czosnyka, M.; Radolovich, D.; Balestreri, M.; Lavinio, A.; Hutchinson, P.; Timofeev, I.; Smielewski, P.; Pickard, J.D. Gender-related differences in intracranial hypertension and outcome after traumatic brain injury. In Acta Neurochirurgica Supplements; Springer: Vienna, Austria, 2008; Volume 102, pp. 25–28. [Google Scholar] [CrossRef]
- Davis, D.P.; Douglas, D.J.; Smith, W.; Sise, M.J.; Vilke, G.M.; Holbrook, T.L.; Kennedy, F.; Eastman, A.B.; Velky, T.; Hoyt, D.B. Traumatic Brain Injury Outcomes in Pre- and Post- Menopausal Females Versus Age-Matched Males. J. Neurotrauma 2006, 23, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.; Ley, E.J.; Tillou, A.; Cryer, G.; Margulies, D.R.; Salim, A. The Effect of Gender on Patients with Moderate to Severe Head Injuries. J. Trauma 2009, 67, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.; Engelhardt, M. Anatomy of the Hippocampal Formation. Front. Neurol. Neurosci. 2014, 34, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Cembrowski, M.S.; Spruston, N. Heterogeneity within classical cell types is the rule: Lessons from hippocampal pyramidal neurons. Nat. Rev. Neurosci. 2019, 20, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Dudek, S.M.; Alexander, G.; Farris, S. Rediscovering area CA2: Unique properties and functions. Nat. Rev. Neurosci. 2016, 17, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Davidsson, J.; Risling, M. Characterization of pressure distribution in penetrating traumatic brain injuries. Front. Neurol. 2015, 6, 51. [Google Scholar] [CrossRef] [Green Version]
- Plantman, S.; Ng, K.C.; Lu, J.; Davidsson, J.; Risling, M. Characterization of a Novel Rat Model of Penetrating Traumatic Brain Injury. J. Neurotrauma 2012, 29, 1219–1232. [Google Scholar] [CrossRef]
- Davidsson, J.; Risling, M. Experimental Models for Neurotrauma Research. In Injury Models of the Central Nervous System: Methods and Protocols; Kobeissy, F.H., Dixon, C.E., Hayes, R.L., Mondello, S., Eds.; Springer: New York, NY, USA, 2016; pp. 267–288. [Google Scholar]
- Yu, X.; Wu, J.; Hu, M.; Wu, J.; Zhu, Q.; Yang, Z.; Xie, X.; Feng, Y.-Q.; Yue, J. Glutamate affects the CYP1B1- and CYP2U1-mediated hydroxylation of arachidonic acid metabolism via astrocytic mGlu5 receptor. Int. J. Biochem. Cell Biol. 2019, 110, 111–121. [Google Scholar] [CrossRef]
- Arevalo, M.-A.; Azcoitia, I.; Garcia-Segura, L.M. The neuroprotective actions of oestradiol and oestrogen receptors. Nat. Rev. Neurosci. 2015, 16, 17–29. [Google Scholar] [CrossRef]
- Paterno, R.; Folweiler, K.A.; Cohen, A.S. Pathophysiology and Treatment of Memory Dysfunction After Traumatic Brain Injury. Curr. Neurol. Neurosci. Rep. 2017, 17, 52. [Google Scholar] [CrossRef]
- Schmitt, S.; Dichter, M.A. Electrophysiologic recordings in traumatic brain injury. Handb. Clin. Neurol. 2015, 127, 319–339. [Google Scholar]
- Wolf, J.A.; Koch, P.F. Disruption of Network Synchrony and Cognitive Dysfunction After Traumatic Brain Injury. Front. Syst. Neurosci. 2016, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Taxier, L.R.; Gross, K.S.; Frick, K.M. Oestradiol as a neuromodulator of learning and memory. Nat. Rev. Neurosci. 2020, 21, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Mannella, P.; Simoncini, T.; Genazzani, A.R. Estrogens and progestins: Molecular effects on brain cells. Horm. Mol. Biol. Clin. Investig. 2010, 4, 609–613. [Google Scholar] [CrossRef]
- Kato, A.; Hojo, Y.; Higo, S.; Komatsuzaki, Y.; Murakami, G.; Yoshino, H.; Uebayashi, M.; Kawato, S. Female hippocampal estrogens have a significant correlation with cyclic fluctuation of hippocampal spines. Front. Neural Circuits 2013, 7, 149. [Google Scholar] [CrossRef] [Green Version]
- Doran, S.J.; Ritzel, R.M.; Glaser, E.P.; Henry, R.; Faden, A.I.; Loane, D. Sex Differences in Acute Neuroinflammation after Experimental Traumatic Brain Injury Are Mediated by Infiltrating Myeloid Cells. J. Neurotrauma 2018, 36, 1040–1053. [Google Scholar] [CrossRef] [PubMed]
- Villapol, S.; Loane, D.J.; Burns, M.P. Sexual dimorphism in the inflammatory response to traumatic brain injury. Glia 2017, 65, 1423–1438. [Google Scholar] [CrossRef]
- Drozdzik, A.; Dziedziejko, V.; Kurzawski, M. IL-1 and TNF-α regulation of aryl hydrocarbon receptor (AhR) expression in HSY human salivary cells. Arch. Oral Biol. 2014, 59, 434–439. [Google Scholar] [CrossRef]
- Bhattacharyya, K.K.; Brake, P.B.; Eltom, S.E.; Otto, S.A.; Jefcoate, C.R. Identification of a rat adrenal cytochrome P450 active in polycyclic hydrocarbon metabolism as rat CYP1B1. Demonstration of a unique tissue-specific pattern of hormonal and aryl hydrocarbon receptor-linked regulation. J. Biol. Chem. 1995, 270, 11595–11602. [Google Scholar] [CrossRef] [Green Version]
- Christou, M.; Savas, U.; Schroeder, S.; Shen, X.; Thompson, T.; Gould, M.N.; Jefcoate, C.R. Cytochromes CYP1A1 and CYP1B1 in the rat mammary gland: Cell-specific expression and regulation by polycyclic aromatic hydrocarbons and hormones. Mol. Cell. Endocrinol. 1995, 115, 41–50. [Google Scholar] [CrossRef]
- Dell’Aquila, M.; Maiese, A.; De Matteis, A.; Viola, R.V.; Arcangeli, M.; La Russa, R.; Fineschi, V. Traumatic brain injury: Estimate of the age of the injury based on neuroinflammation, endothelial activation markers and adhesion molecules. Histol. Histopathol. 2021, 36, 795–806. [Google Scholar]
- Ziegler, N.; Awwad, K.; Fisslthaler, B.; Reis, M.; Devraj, K.; Corada, M.; Minardi, S.P.; Dejana, E.; Plate, K.H.; Fleming, I.; et al. β-Catenin Is Required for Endothelial Cyp1b1 Regulation Influencing Metabolic Barrier Function. J. Neurosci. 2016, 36, 8921–8935. [Google Scholar] [CrossRef]
- Roland, J.J.; Stewart, A.L.; Janke, K.L.; Gielow, M.R.; Kostek, J.A.; Savage, L.M.; Servatius, R.; Pang, K.C. Medial Septum-Diagonal Band of Broca (MSDB) GABAergic Regulation of Hippocampal Acetylcholine Efflux Is Dependent on Cognitive Demands. J. Neurosci. 2014, 34, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Ghadiri, T.; Vakilzadeh, G.; Hajali, V.; Khodagholi, F. Progesterone modulates post-traumatic epileptogenesis through regulation of BDNF-TrkB signaling and cell survival-related pathways in the rat hippocampus. Neurosci. Lett. 2019, 709, 134384. [Google Scholar] [CrossRef]
- Hummel, R.; Ulbrich, S.; Appel, D.; Li, S.; Hirnet, T.; Zander, S.; Bobkiewicz, W.; Gölz, C.; Schäfer, M.K. Administration of all-trans retinoic acid after experimental traumatic brain injury is brain protective. Br. J. Pharmacol. 2020, 177, 5208–5223. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Kelly, K.K.; Rumian, N.L.; Siegenthaler, J.A. Retinoic Acid Is Required for Neural Stem and Progenitor Cell Proliferation in the Adult Hippocampus. Stem Cell Rep. 2018, 10, 1705–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y.; et al. RNAscope: A novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Player, A.N.; Shen, L.P.; Kenny, D.; Antao, V.P.; Kolberg, J.A. Single-copy gene detection using branched DNA (bDNA) in situ hybridization. J. Histochem. Cytochem. 2001, 49, 603–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, F.R. The Female Rat Reproductive Cycle: A Practical Histological Guide to Staging. Toxicol. Pathol. 2008, 36, 375–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Intervention | Survival Time | Number of ISH | Number of IHC |
|---|---|---|---|
| Sham-surgery | 24 h | Male = 3, Female = 4 | Male = 3, Female = 4 |
| pTBI | 24 h | Male = 4, Female = 4 | Male = 4, Female = 4 |
| Sham-surgery | 72 h | Male = 4, Female = 4 | Male = 4, Female = 4 |
| pTBI | 72 h | Male = 4, Female = 4 | Male = 4, Female = 4 |
| Sham-surgery | 120 h | Male = 4, Female = 4 | Male = 4, Female = 4 |
| pTBI | 120 h | Male = 4, Female = 4 | Male = 4, Female = 4 |
| Sham-surgery | 168 h | Male = 4, Female = 4 | Male = 4, Female = 4 |
| pTBI | 168 h | Male = 4, Female = 3 | Male = 4, Female = 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lidin, E.; Sköld, M.K.; Angéria, M.; Davidsson, J.; Risling, M. Hippocampal Expression of Cytochrome P450 1B1 in Penetrating Traumatic Brain Injury. Int. J. Mol. Sci. 2022, 23, 722. https://doi.org/10.3390/ijms23020722
Lidin E, Sköld MK, Angéria M, Davidsson J, Risling M. Hippocampal Expression of Cytochrome P450 1B1 in Penetrating Traumatic Brain Injury. International Journal of Molecular Sciences. 2022; 23(2):722. https://doi.org/10.3390/ijms23020722
Chicago/Turabian StyleLidin, Erik, Mattias K. Sköld, Maria Angéria, Johan Davidsson, and Mårten Risling. 2022. "Hippocampal Expression of Cytochrome P450 1B1 in Penetrating Traumatic Brain Injury" International Journal of Molecular Sciences 23, no. 2: 722. https://doi.org/10.3390/ijms23020722
APA StyleLidin, E., Sköld, M. K., Angéria, M., Davidsson, J., & Risling, M. (2022). Hippocampal Expression of Cytochrome P450 1B1 in Penetrating Traumatic Brain Injury. International Journal of Molecular Sciences, 23(2), 722. https://doi.org/10.3390/ijms23020722