
Host Tau Genotype Specifically Designs and Regulates Tau Seeding and Spreading and Host Tau Transformation Following Intrahippocampal Injection of Identical Tau AD Inoculum

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. WT Mice, Mutant Wild-Type KO for Murine Tau (mtWT), and Tau Transgenic Mice Expressing Human Tau (hTau)
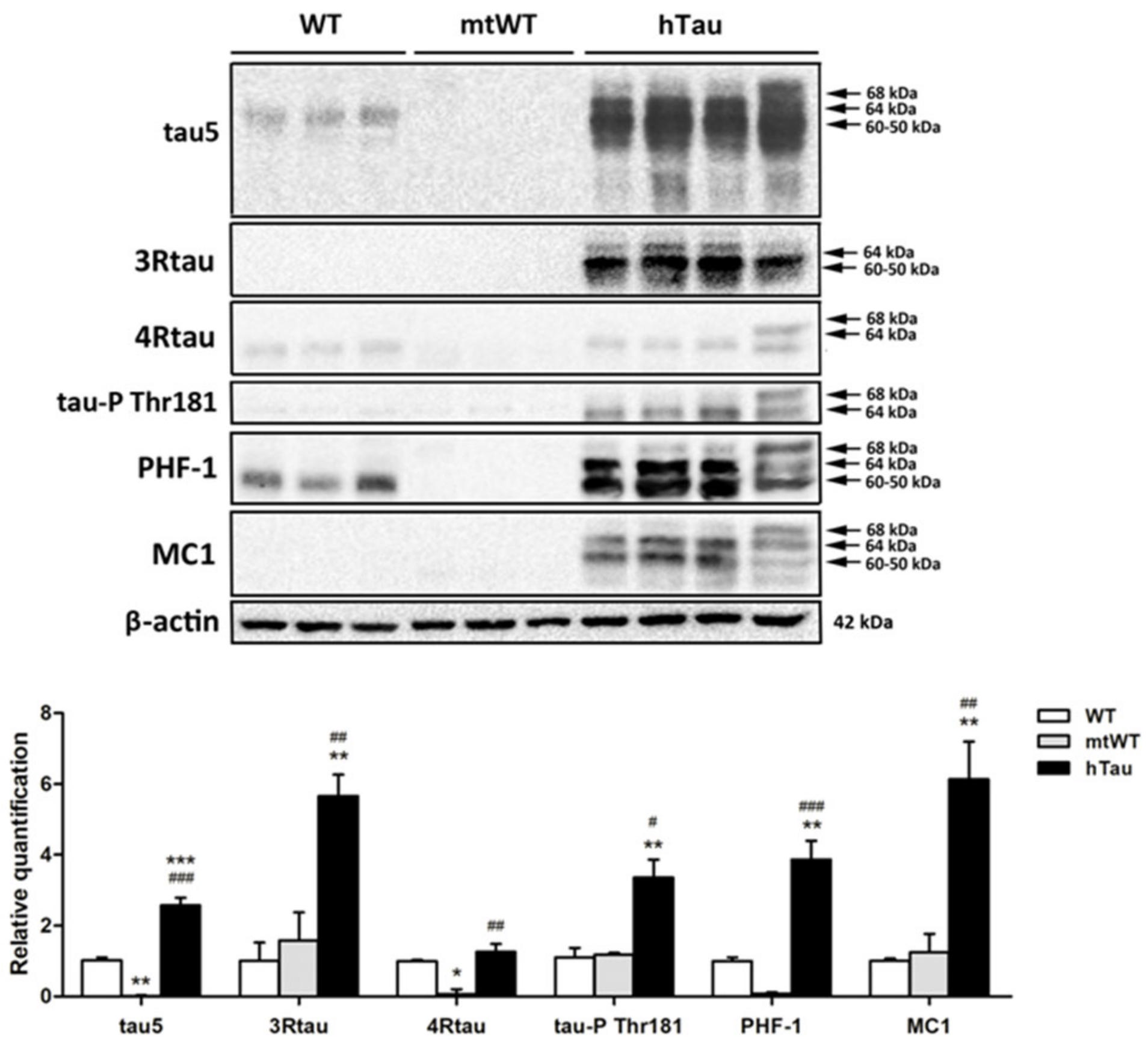
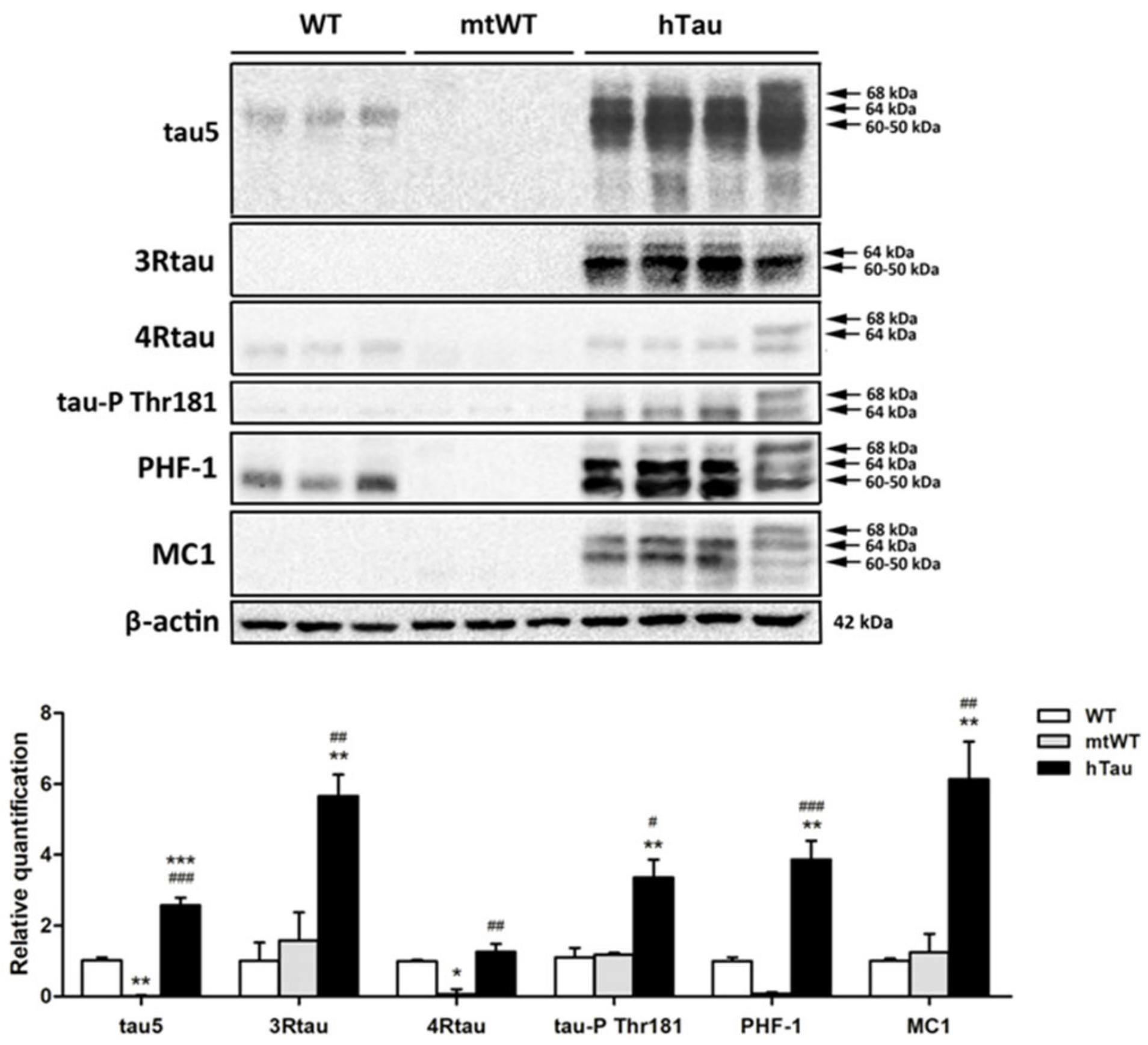
2.2. Western Blotting of Total Brain Homogenates in WT, hTau, and Mutant Wild-Type KO for Murine Tau (mtWt)
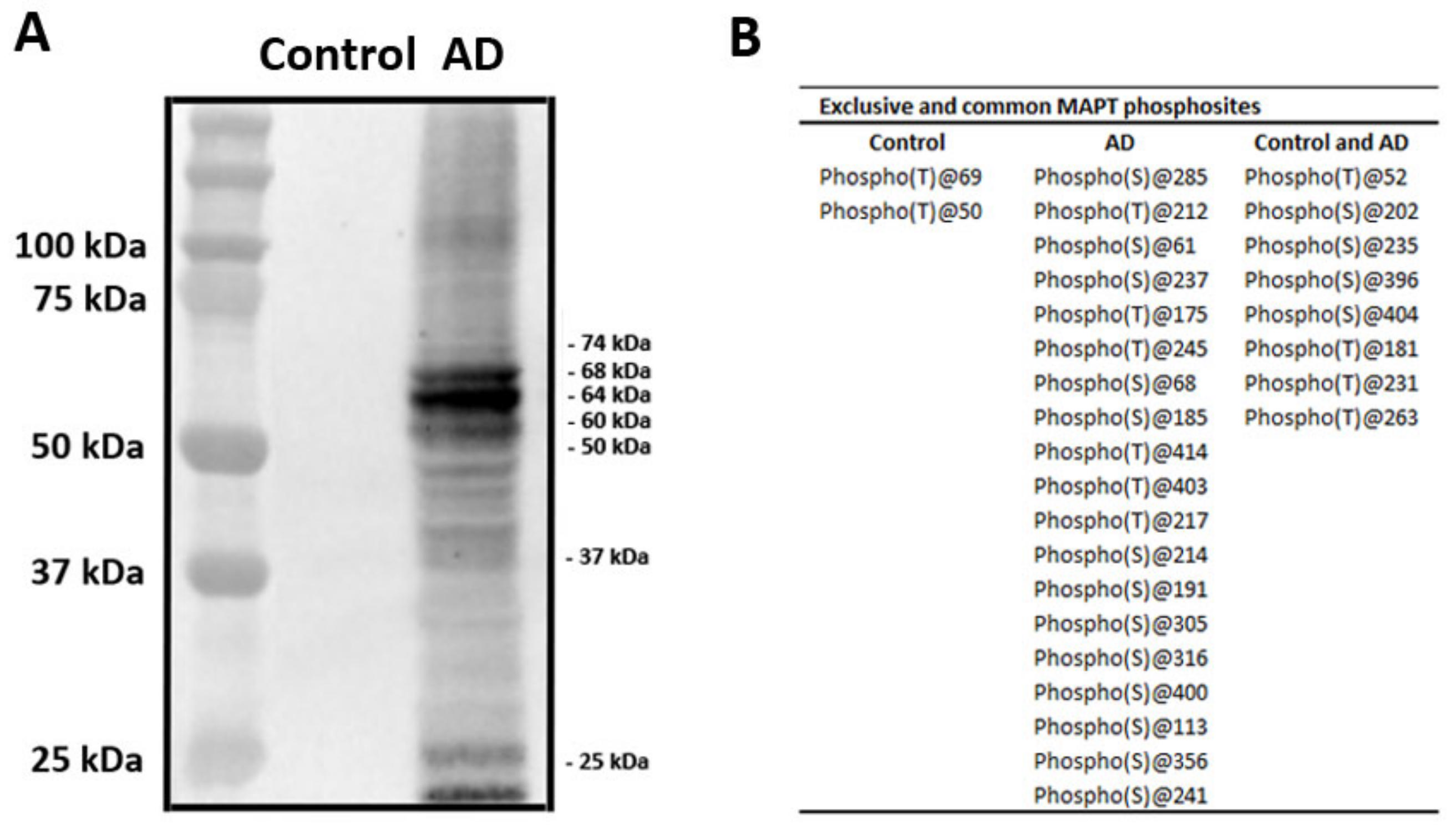
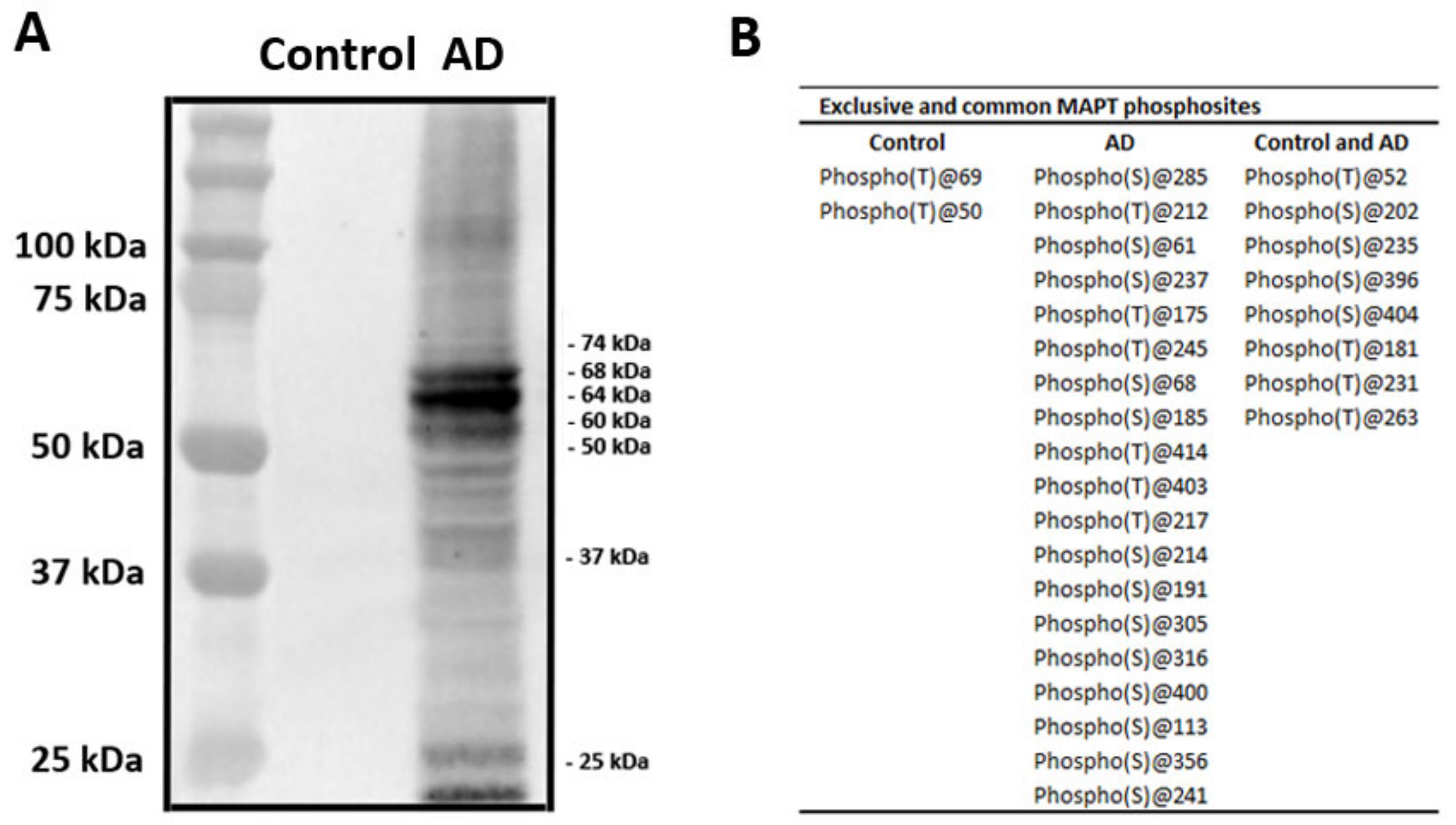
2.3. Characteristics of Sarkosyl-Insoluble Fractions Used for Inoculation
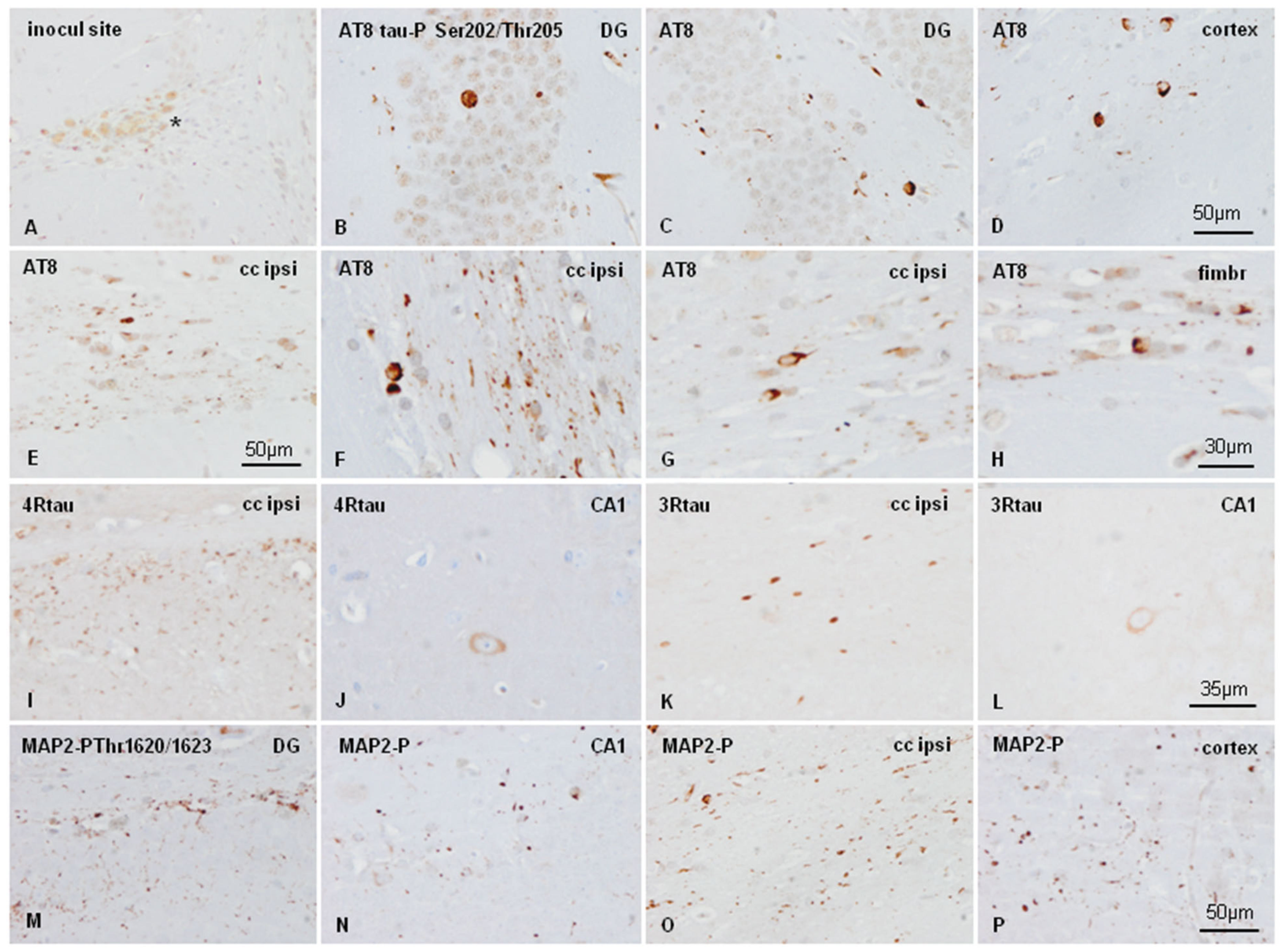
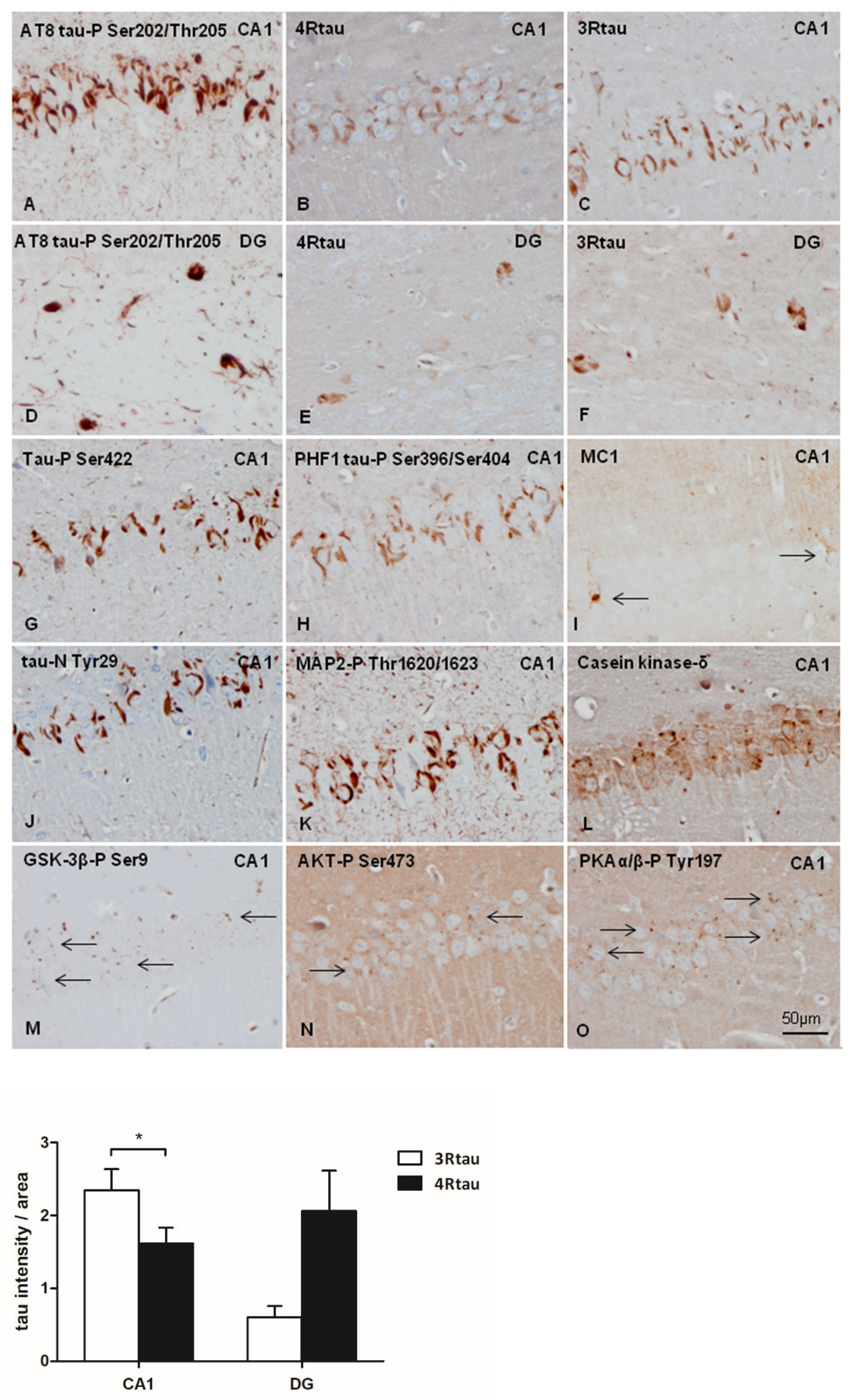
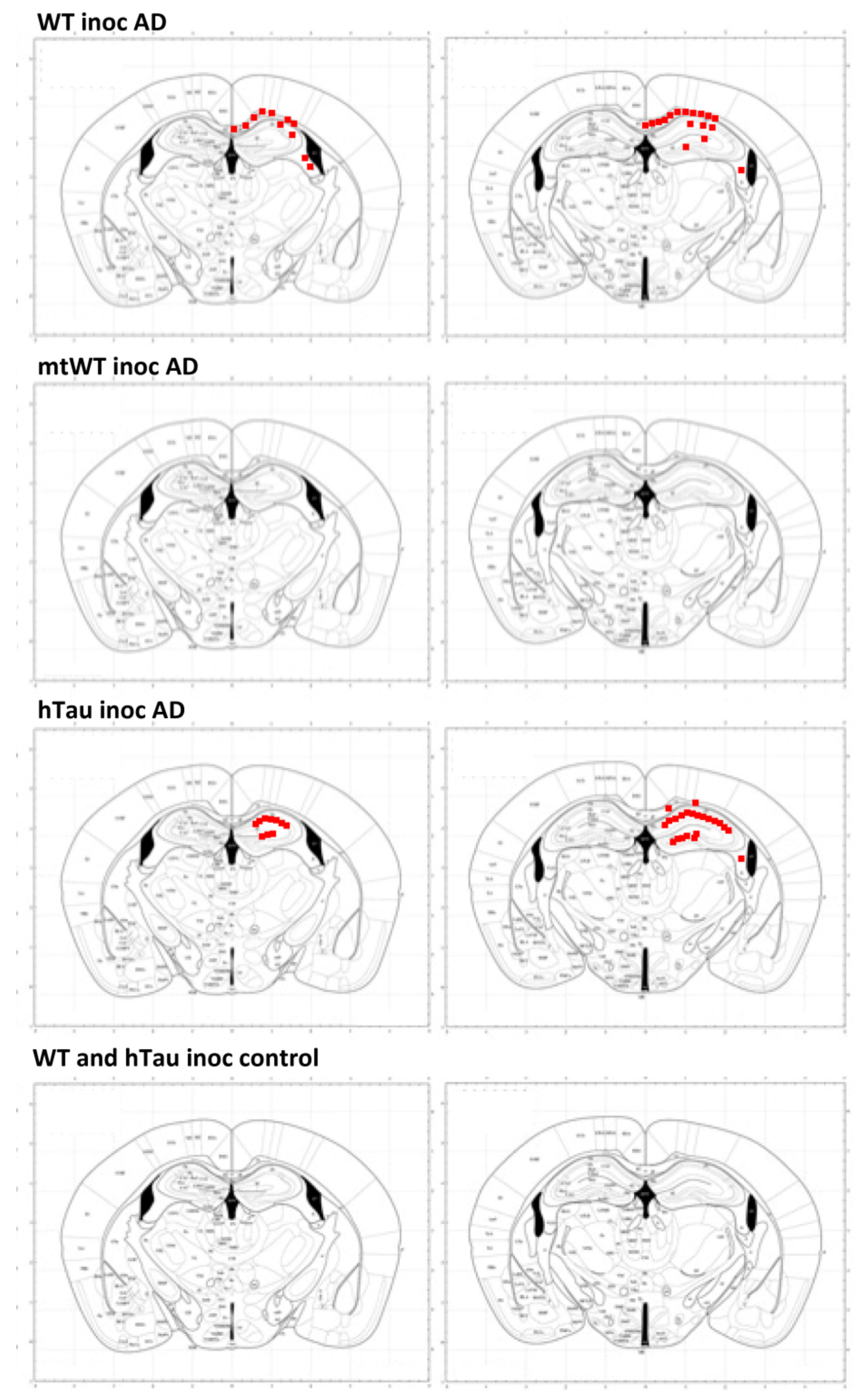
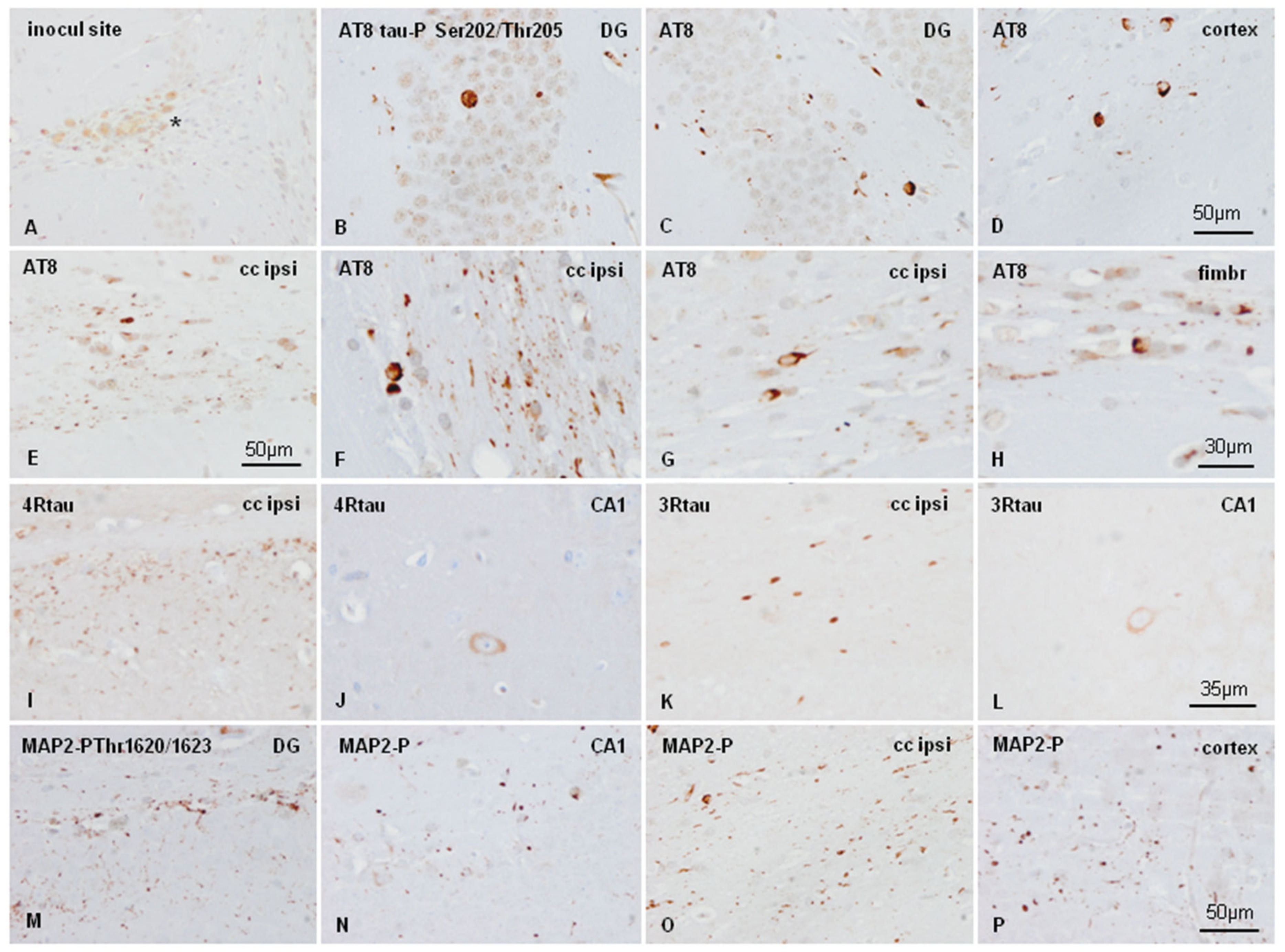
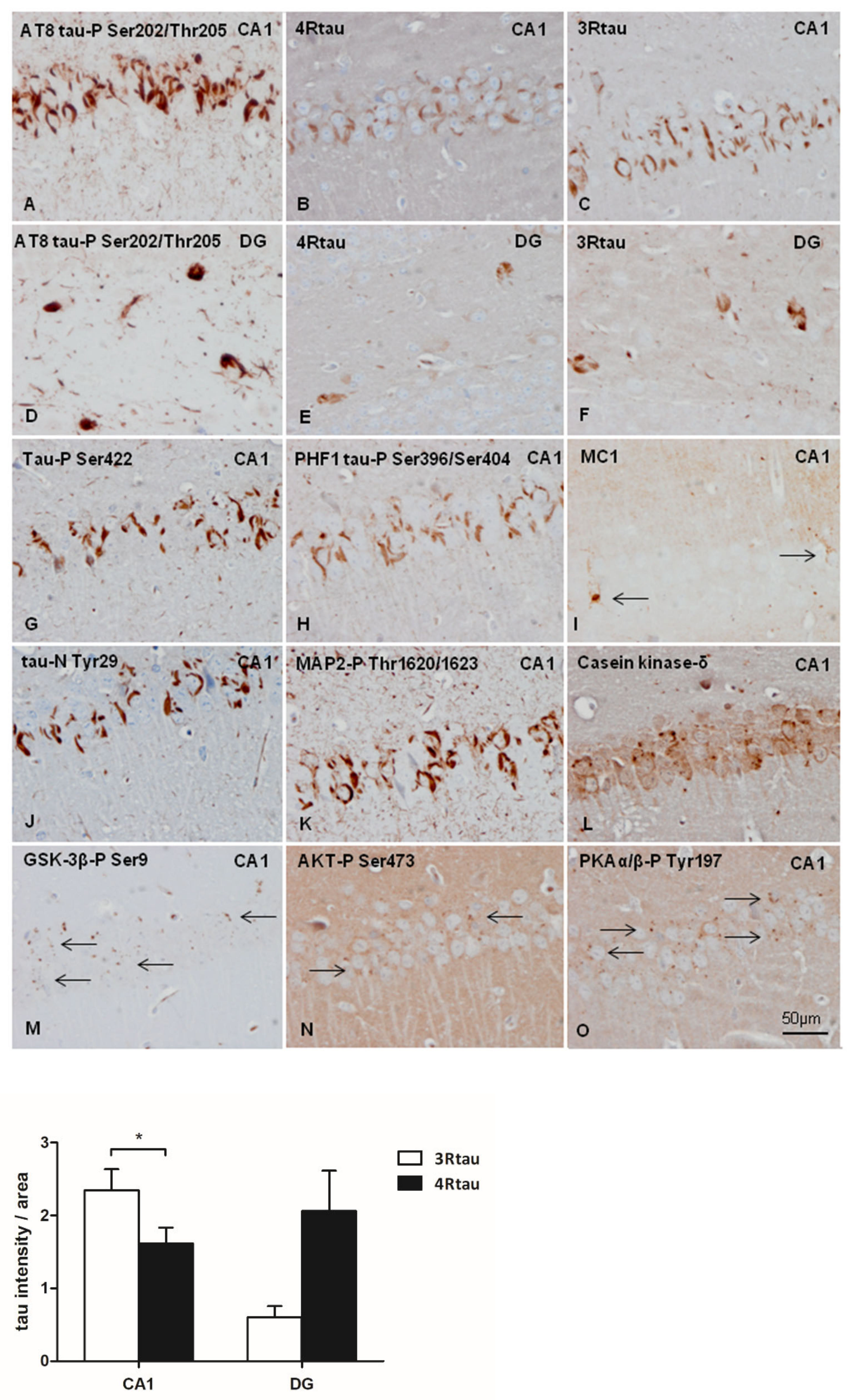
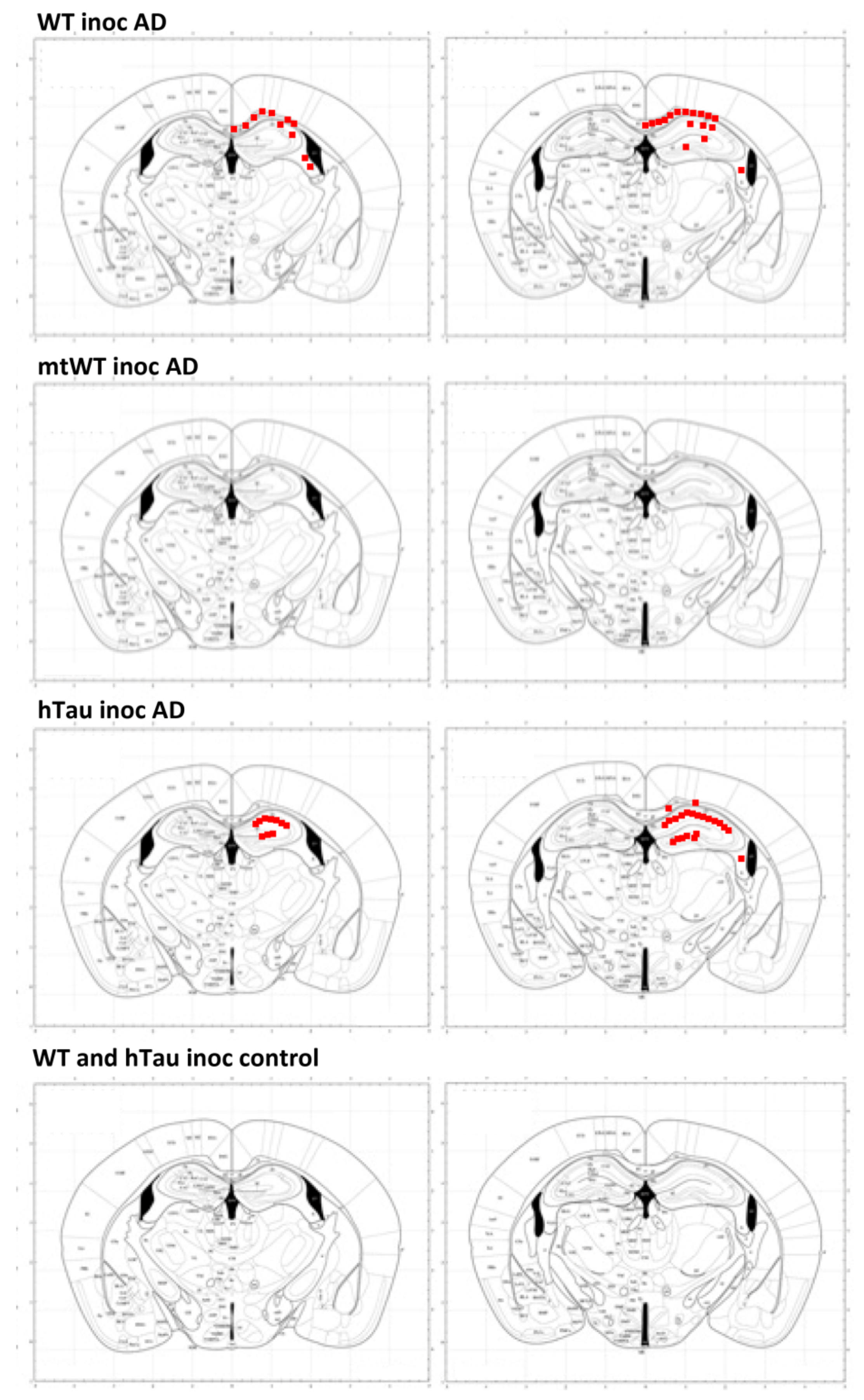
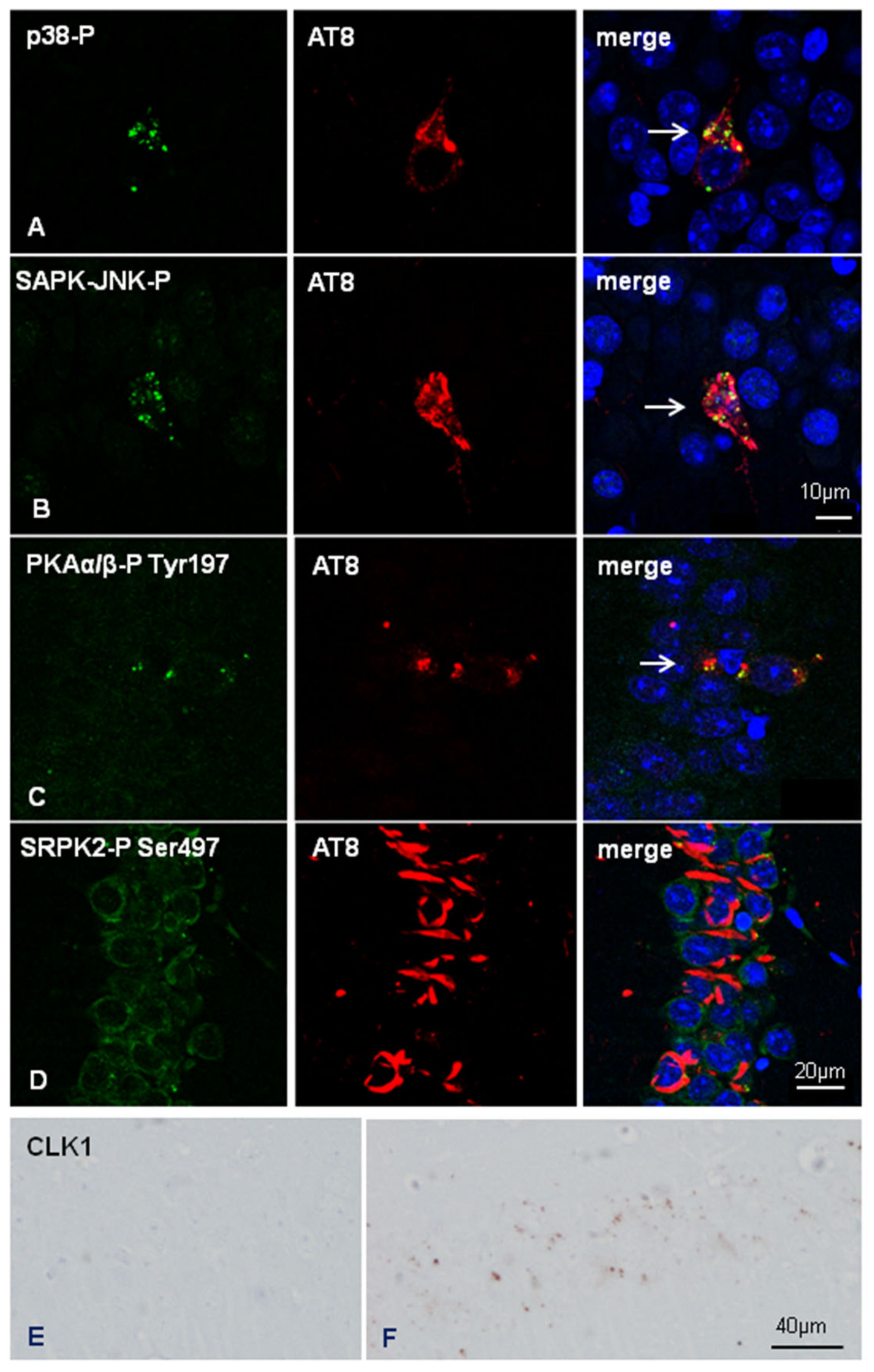
2.4. Inoculation of Mice with Sarkosyl-Insoluble Fractions from AD and Control; mtWT Mice, and Characteristics of Tau Seeding in WT and hTau Genotypes
3. Discussion
4. Materials and Methods
4.1. Brain Samples
4.2. Extraction of Sarkosyl-Insoluble Fractions and Western Blotting
4.3. Animals
4.4. (Phospho)Proteomics of Sarkosyl-Insoluble Fractions
4.5. Inoculation of Sarkosyl-Insoluble from Alzheimer Disease and Control Homogenates in the Hippocampus of WT, hTau, and mtWT Mice
4.6. Immunohistochemistry
4.7. Quantification of Abnormal Deposits Revealed by Immunohistochemistry
4.8. Double-Labeling Immunofluorescence and Confocal Microscopy
4.9. Gel Electrophoresis and Western Blotting
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AD: | Alzheimer’s disease |
| WT: | wild type mouse expressing murine 4Rtau |
| mtWT: | homozygous transgenic mouse knock-out for murine tau protein |
| hTau: | homozygous transgenic mouse knock-out for murine tau protein and heterozygous expressing human forms of 3Rtau and 4Rtau proteins |
| 3Rtau: | tau with three repeats |
| 4Rtau: | tau with four repeats |
| β-actin; CK1-δ | casein kinase δ |
| eIF2α | eukaryotic initiation factor 2α |
| GSK β: | glycogen synthase kinase 3β |
| IRE1: | inositol-requiring enzyme 1α |
| LAMP1: | lysosomal-associated membrane protein 3 |
| LC3: | microtubule-associated protein 1A/1B-light chain 3 |
| MAP2: | microtubule-associated protein 2 |
| NFL: | neurofilament light molecular weight |
| p38: | p38-kinase |
| PHF1: | anti-paired helical filament 1 |
| PKAα/β: | cAMP-dependent protein kinase A |
| SAPK/JNK: | stress-activated protein kinase/Jun amino-terminal kinase |
| SRC: | tyrosine-protein kinase SRC |
| Tau 5: | monoclonal recognizing total tau protein |
| Tau AT8 | monoclonal tau antibody phosphorylation sites Ser202/Thr305 |
| Tau C3: | tau truncated at aspartic acid 421 |
| Tau MC1: | conformation tau |
| Tau-N: | nitrated tau |
| Tau-P: | phosphorylated tau. The phosphorylation or nitration of the protein at the specific site is indicated in every case |
References
- Kovacs, G.G. Invited review: Neuropathology of tauopathies: Principles and practice. Neuropathol. Appl. Neurobiol. 2015, 41, 3–23. [Google Scholar] [CrossRef]
- Iqbal, K.; Liu, F.; Gong, C.X. Tau and neurodegenerative disease: The story so far. Nat. Rev. Neurol. 2016, 12, 15–27. [Google Scholar] [CrossRef]
- Arendt, T.; Stieler, J.T.; Holzer, M. Tau and tauopathies. Brain Res. Bull. 2016, 126, 238–292. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.; Dickson, D.W. Propagation of tau pathology: Hypotheses, discoveries, and yet unresolved questions from experimental and human brain studies. Acta Neuropathol. 2016, 131, 27–48. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Eisenberg, D.S.; Crowther, R.A. Propagation of tau aggregates and neurodegeneration. Annu. Rev. Neurosci. 2017, 40, 189–210. [Google Scholar] [CrossRef]
- Iba, M.; Guo, J.L.; McBride, J.D.; Zhang, B.; Trojanowski, J.Q.; Lee, V.M. Synthetic tau fibrils mediate transmission of neurofibrillary tangles in a transgenic mouse model of Alzheimer’s-like tauopathy. J. Neurosci. 2013, 33, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Peeraer, E.; Bottelbergs, A.; Van Kolen, K.; Stancu, I.C.; Vasconcelos, B.; Mahieu, M.; Duytschaever, H.; Ver Donck, L.; Torremans, A.; Sluydts, E.; et al. Intracerebral injection of preformed synthetic fibrils initiates widesprad tauopathy and neuronal loss in the brains of tau transgenic mice. Neurobiol. Dis. 2015, 73, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, Z.; Cooper, J.; Murray, T.K.; Garn, K.; McNaughton, E.; Clarke, H.; Parhizkar, S.; Ward, M.A.; Cavallini, A.; Jackson, S.; et al. A novel in vivo model of tau propagation with rapid and progressive neurofibrillary tangle pathology: The pattern of spread is determined by connectivity, not proximity. Acta Neuropathol. 2014, 127, 667–683. [Google Scholar] [CrossRef] [Green Version]
- Clavaguera, F.; Bolmont, T.; Crowther, R.A.; Abramowski, D.; Frank, S.; Probst, A.; Fraser, G.; Stalder, A.K.; Beibel, M.; Staufenbiel, M.; et al. Transmission and spreading of tauopathy in transgenic mouse brain. Nat. Cell Biol. 2009, 111, 909–913. [Google Scholar] [CrossRef]
- Clavaguera, F.; Akatsu, H.; Fraser, G.; Crowther, R.A.; Frank, S.; Hench, J.; Probst, A.; Winkler, D.T.; Reichwald, J.; Staufenbiel, M.; et al. Brain homogenates from human tauopathies induce tau inclusions in mouse brain. Proc. Natl. Acad. Sci. USA 2013, 110, 9535–9540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavaguera, F.; Hench, J.; Goedert, M.; Tolnay, M. Invited review: Prion-like transmission and spreading of tau pathology. Neuropathol. Appl. Neurobiol. 2015, 41, 47–58. [Google Scholar] [CrossRef]
- Boluda, S.; Iba, M.; Zhang, B.; Raible, K.M.; Lee, V.M.; Trojanowski, J.Q. Differential induction and spread of tau pathology in young PS19 tau transgenic mice following intracerebral injections of pathological tau from Alzheimer’s disease or corticobasal degeneration brains. Acta Neuropathol. 2015, 129, 221–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audouard, E.; Houben, S.; Masaracchia, C.; Yilmaz, Z.; Suain, V.; Authelet, M.; De Decker, R.; Buée, L.; Boom, A.; Leroy, K.; et al. High-molecular weight paired helical filaments from Alzheimer brain induces seeding of wild-type mouse tau into argyrophilic 4R tau pathology in vivo. Am. J. Pathol. 2016, 186, 2709–2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.L.; Narasimhan, S.; Changolkar, L.; He, Z.; Stieber, A.; Zhang, B.; Gathagan, R.J.; Iba, M.; McBride, J.D.; Trojanowski, J.Q.; et al. Unique pathological tau conformers from Alzheimer’s brains transmit tau pathology in nontransgenic mice. J. Exp. Med. 2016, 213, 2635–2654. [Google Scholar] [CrossRef]
- Narashima, S.; Guo, J.L.; Changolkar, L.; Stieber, A.; McBride, J.D.; Silva, L.V.; He, Z.; Zhang, B.; Gathagan, R.J.; Trojanowski, J.Q.; et al. Pathological tau strains from human brains recapitulate the diversity of tauopathies in non-transgenic mouse brain. J. Neurosci. 2017, 37, 11406–11423. [Google Scholar] [CrossRef]
- Weitzman, S.A.; Narasimhan, S.; He, Z.; Changolkar, L.; McBride, J.D.; Zhang, B.; Schellenberg, G.D.; Trojanowski, J.Q.; Lee, V.M.Y. Insoluble tau from human FTDP-17 cases exhibit unique transmission properties in vivo. J. Neuropathol. Exp. Neurol. 2020, 79, 941–949. [Google Scholar] [CrossRef]
- Goedert, M. Tau proteinopathies and the prion concept. Prog. Mol. Biol. Transl. Sci. 2020, 175, 239–259. [Google Scholar] [CrossRef]
- Goedert, M.; Spillantini, M.G. Propagation of tau aggregates. Mol. Brain. 2017, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudner, A.; Colin, M.; Dujardin, S.; Medina, M.; Dewachter, I.; Naini, S.M.A.; Mandelkow, E.M.; Mandelkow, E.; Luc Buée, L.; Michel Goedert, M.; et al. What is the evidence that tau pathology spreads through prion-like propagation? Acta Neuropathol. Commun. 2017, 5, 99. [Google Scholar] [CrossRef]
- Vaquer-Alicea, J.; Diamond, M.I.; Joachimiak, L.A. Tau strains shape disease. Acta Neuropathol. 2021, 142, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Tseng, J.H.; Ajit, A.; Tabassum, Z.; Patel, N.; Tian, X.; Chen, Y.; Prevatte, A.W.; Ling, K.; Rigo, F.; Meeker, R.B.; et al. Tau seeds are subject to aberrant modifications resulting in distinct signatures. Cell Rep. 2021, 35, 109037. [Google Scholar] [CrossRef]
- Ferrer, I.; García, M.A.; González, I.L.; Lucena, D.D.; Villalonga, A.R.; Carmona, M.; Llorens, F.; Garcia-Esparcia, P.; Martinez-Maldonado, A.; Mendez, M.F.; et al. Aging-related tau astrogliopathy (ARTAG): Not only tau phosphorylation in astrocytes. Brain Pathol. 2018, 28, 965–985. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, I.; García, M.A.; Carmona, M.; Andrés-Benito, P.; Torrejón-Escribano, B.; Garcia-Esparcia, P.; del Rio, J.A. Involvement of oligodendrocytes in tau seeding and spreading in tauopathies. Front. Aging Neurosci. 2019, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, I.; Andrés-Benito, P.; Sala-Jarque, J.; Gil, V.; Del Rio, J.A. Capacity for seeding and spreading of argyrophilic grain disease in a wild-type murine model; comparisons with primary age-related tauopathy. Front. Mol. Neurosci. 2020, 13, 101. [Google Scholar] [CrossRef]
- Ferrer, I.; Andrés-Benito, P.; Zelaya, M.V.; Aguirre, M.E.E.; Carmona, M.; Ausín, K.; Lachén-Montes, M.; Fernández-Irigoyen, J.; Santamaría, E.; Del Rio, J.A. Familial globular glial tauopathy linked to MAPT mutations: Molecular neuropathology and seeding capacity of a prototypical mixed neuronal and glial tauopathy. Acta Neuropathol. 2020, 139, 735–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, I.; Zelaya, M.V.; García, M.A.; Carmona, M.; López-González, I.; Andrés-Benito, P.; López-González, I.; Andrés-Benito, P.; Lidón, L.; Gavín, R.; et al. Relevance of host tau in tau seeding and spreading in tauopathies. Brain Pathol. 2020, 30, 298–318. [Google Scholar] [CrossRef]
- Andorfer, C.; Kress, Y.; Espinoza, M.; de Silva, R.; Tucker, K.L.; Barde, Y.A.; Duff, K.; Davies, P. Hyperphosphorylation and aggregation of tau in mice expressing normal human tau isoforms. J. Neurochem. 2003, 86, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Andorfer, C.; Acker, C.M.; Kress, Y.; Hof, P.R.; Duff, K.; Davies, P. Cell-cycle reentry and cell death in transgenic mice expressing nonmutant human tau isoforms. J. Neurosci. 2005, 25, 5446–5454. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- LoPresti, P.; Szuchet, S.; Papasozomenos, S.C.; Zinkowski, R.P.; Binder, L.I. Functional implications for the microtubule-associated protein tau: Localization in oligodendrocytes. Proc. Natl. Acad. Sci. USA 1995, 92, 10369–10373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorath, M.; Stahnke, T.; Mronga, T.; Goldbaum, O.; Richter-Landsberg, C. Developmental changes of tau protein and mRNA in cultured rat brain oligodendrocytes. Glia 2001, 36, 89–101. [Google Scholar] [CrossRef]
- Hoozemans, J.J.; van Haastert, E.S.; Nijholt, D.A.; Rozemuller, A.J.; Eikelenboom, P.; Scheper, W. The unfolded protein response is activated in pretangle neurons in Alzheimer’s disease hippocampus. Am. J. Pathol. 2009, 174, 1241–1251. [Google Scholar] [CrossRef] [Green Version]
- Wiersma, V.I.; van Ziel, A.M.; Vazquez-Sanchez, S.; Nölle, A.; Berenjeno-Correa, E.; Bonaterra-Pastra, A.; Clavaguera, F.; Tolnay, M.; Musters, R.J.P.; van Weering, J.R.T.; et al. Granulovacuolar degeneration bodies are neuron-selective lysosomal structures induced by intracellular tau pathology. Acta Neuropathol. 2019, 138, 943–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, K.E.; Mrak, R.E.; Kuret, J. Granulovacuolar degeneration (GVD) bodies of Alzheimer’s disease (AD) resemble late-stage autophagic organelles. Neuropathol. Appl. Neurobiol. 2011, 37, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Köhler, C. Granulovacuolar degeneration: A neurodegenerative change that accompanies tau pathology. Acta Neuropathol. 2016, 132, 339–359. [Google Scholar] [CrossRef] [PubMed]
- González, M.V.; Vallés-Saiz, L.; Hernández, I.H.; Avila, J.; Hernández, F.; Pérez-Alvarez, M.J. Focal cerebral ischemia induces changes in oligodendrocytic tau isoforms in the damaged area. Glia 2020, 68, 2471–2485. [Google Scholar] [CrossRef] [PubMed]
- Janke, C.; Beck, M.; Stahl, T.; Holzer, M.; Brauer, K.; Bigl, V.; Arendt, T. Phylogenetic diversity of the expression of the microtubule-associated protein tau: Implications for neurodegenerative disorders. Brain Res. Mol. Brain Res. 1999, 68, 119–128. [Google Scholar] [CrossRef]
- Hefti, M.M.; Farrell, K.; Kim, S.; Bowles, K.R.; Fowkes, M.E.; Raj, T.; Crary, J.F. High-resolution temporal and regional mapping of MAPT expression and splicing in human brain development. PLoS ONE 2018, 13, e0195771. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Liu, F. Review·Regulation of alternative splicing of tau exon 10. Neurosci. Bull. 2014, 30, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Duncan, P.I.; Stojdl, D.F.; Marius, R.M.; Scheit, K.H.; Bell, J.C. The Clk2 and Clk3 dual-specificity protein kinases regulate the intranuclear distribution of SR proteins and influence premRNA splicing. Exp. Cell. Res. 1998, 241, 300–308. [Google Scholar] [CrossRef]
- Mermoud, J.E.; Cohen, P.T.; Lamond, A.I. Regulation of mammalian spliceosome assembly by a protein phosphorylation mechanism. EMBO J. 1994, 13, 5679–5688. [Google Scholar] [CrossRef]
- Stojdl, D.F.; Bell, J.C. SR protein kinases: The splice of life. Biochem. Cell Biol. 1999, 77, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, F.; Perez, M.; Lucas, J.J.; Mata, A.M.; Bhat, R.; Avila, J. Glycogen synthase kinase-3 plays a crucial role in tau exon 10 splicing and intranuclear distribution of SC35: Implications for Alzheimer’s disease. J. Biol. Chem. 2004, 279, 3801–3806. [Google Scholar] [CrossRef] [Green Version]
- Kvissel, A.K.; Orstavik, S.; Eikvar, S.; Brede, G.; Jahnsen, T.; Collas, P.; Akusjärvi, G.; Skålhegg, B.S. Involvement of the catalytic subunit of protein kinase A and of HA95 in pre-mRNA splicing. Exp. Cell. Res. 2007, 313, 2795–2809. [Google Scholar] [CrossRef]
- Patel, N.A.; Kaneko, S.; Apostolatos, H.S.; Bae, S.S.; Watson, J.E.; Davidowitz, K.; Chappell, D.S.; Birnbaum, M.J.; Cheng, J.Q.; Cooper, D.R. Molecular and genetic studies imply Akt-mediated signaling promotes protein kinase CbetaII alternative splicing via phosphorylation of serine/arginine-rich splicing factor SRp40. J. Biol. Chem. 2005, 280, 14302–14309. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Qian, W.; Yin, X.; Iqbal, K.; Grundke-Iqbal, I.; Gu, X.; Ding, F.; Gong, C.X.; Liu, F. Cyclic AMP-dependent protein kinase regulates the alternative splicing of tau exon 10: A mechanism involved in tau pathology of Alzheimer’s disease. J. Biol. Chem. 2011, 286, 14639–14648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujardin, S.; Commins, C.; Lathuiliere, A.; Beerepoot, P.; Fernandes, A.R.; Kamath, T.V.; De Los Santos, M.B.; Klickstein, N.; Corjuc, D.L.; Corjuc, B.T.; et al. Tau molecular diversity contributes to clinical heterogeneity in Alzheimer’s disease. Nat. Med. 2020, 26, 1256–1263. [Google Scholar] [CrossRef]
- Li, L.; Shi, R.; Gu, J.; Tung, Y.C.; Zhou, Y.; Zhou, D.; Wu, R.; Chu, D.; Jin, N.; Deng, K.; et al. Alzheimer’s disease brain contains tau fractions with differential prion-like activities. Acta Neuropathol. Commun. 2021, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, I. Brain banking. In Encyclopedia of the Neurological Sciences, 2nd ed.; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 1, pp. 467–473. [Google Scholar]
- Thingholm, T.E.; Jensen, O.N.; Robinson, P.J.; Larsen, M.R. SIMAC (sequential elution from IMAC), a phosphoproteomics strategy for the rapid separation of monophosphorylated from multiply phosphorylated peptides. Mol. Cell. Proteom. 2008, 7, 661–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | WT | hTau | mtTau | Inoc AD WT | inoc AD Tau | Inoc C hTau |
|---|---|---|---|---|---|---|
| 3Rtau | - | diffuse | - | + | ++ (nft) | diffuse |
| 4Rtau | diffuse | diffuse | - | + | +++ (nft) | diffuse |
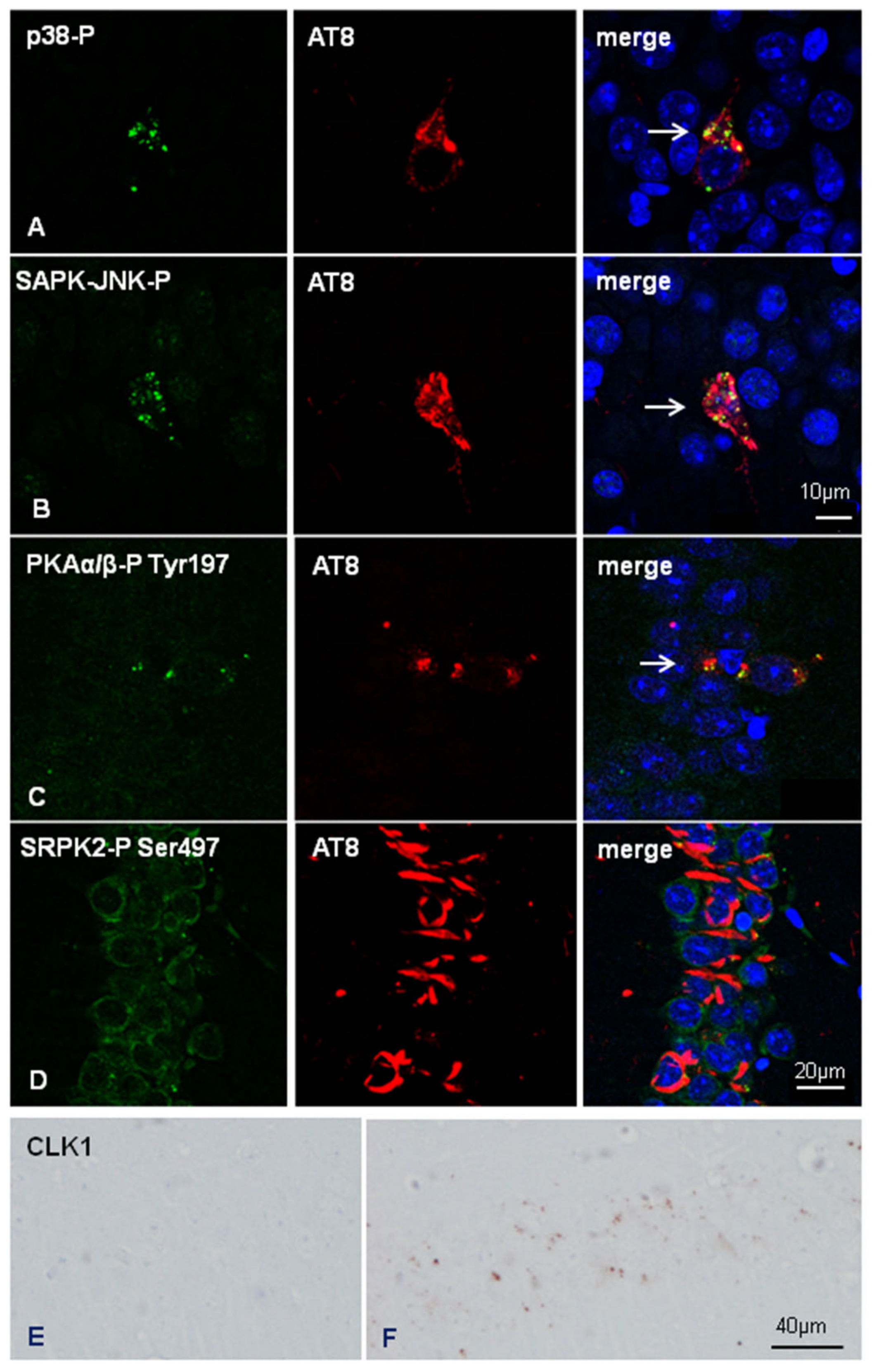
| AKT-P Ser473 | - | - | - | - | ++ (gr) | - |
| CLK1 | - | - | - | - | ++ (gr) | - |
| CK1-δ | - | - | - | - | ++ (gr) | - |
| eIF2α-P Ser51 | - | - | - | - | - | - |
| GSK-3β-P Ser9 | - | - | - | - | ++ (gr) | - |
| IRE-P Ser274 | - | - | - | - | - | - |
| LAMP1 | - | - | - | - | - | - |
| LC3 | - | - | - | - | - | - |
| MAP2-P Thr1620/1623 | - | - | - | + (ptg) | +++ (nft) | - |
| NFL-P Ser473 | - | - | - | - | - | - |
| p38-P Thr180/Tyr182 | - | - | - | + (gr) | ++ (gr) | - |
| PHF1, tau-P Ser396/Ser404 | - | +/− (gl) | - | - | ++ (nft) | +/− (gl) |
| PKAα/β-P Tyr197 | - | - | - | - | ++ (gr) | - |
| SAPK/JNK-P-Thr183/Thr185 | - | - | - | + (gr) | ++ (gr) | - |
| SRC-P Tyr416 | - | - | - | - | - | - |
| SRPK1 | - | - | - | - | - | - |
| SRPK2-P Ser497 | - | - | - | - | - | - |
| tau AT8-P Ser202/Thr205 | nuclei | - | - | + (ptg) | +++ (nft) | - |
| tau C-3 | - | - | - | - | - | - |
| tau MC1 | - | +/− (gl) | - | + (ptg) | +++ (nft) | +/− (gl) |
| tau-N Tyr29 | - | - | - | - | ++ (nft) | - |
| tau-P Ser422 | - | - | - | + (ptg) | +++ nft) | - |
| Ubiquitin | - | - | - | - | - | - |
| Antibody | Supplier | Reference | Host | WB Dil | IHQ Dil |
|---|---|---|---|---|---|
| 3Rtau | Upstate (Syracuse, NY, USA) | 05-803 | Ms | 1/1000 | 1/800 |
| 4Rtau | Millipore (Barcelona, Spain) | 05-804 | Ms | 1/1000 | 1/50 |
| AKT1-P Ser473 | Abcam (Cambridge, UK) | Ab18206 | Rb | - | 1:100 |
| β-actin | Sigma (Barcelona, Spain) | A5316 | Ms | 1/30,000 | - |
| CLK1 (75-100 aa) | LS Bio (Derio, Spain) | LS-C382760 | Ms | - | 1/100 |
| CK1-δ | Abcam (Cambridge, UK) | ab85320 | Ms | - | 1/500 |
| eIF2α-P Ser51 | Thermo Scientific (Barcelona, Spain) | MA5-15133 | Rb | - | 1/100 |
| GFAP | Dako (Barcelona, Spain) | Z0334 | Rb | - | 1/400 |
| GSK-3β-P Ser9 | Cell signaling (Danvers, MA, USA) | 9336 | Rb | - | 1/100 |
| Iba1 | Wako (Richmond, VA, USA) | 019-19741 | Rb | - | 1/1000 |
| IRE-P Ser274 | Abcam (Cambridge, UK) | ab48187 | Rb | - | 1/100 |
| LAMP1 | Santa Cruz (CA, USA) | Sc5570 | Rb | - | 1/10 |
| LC3 | Cell Signaling (Danvers, MA, USA) | 2775 | Rb | - | 1/100 |
| MAP2-P Thr1620/1623 | Cell Signaling (Danvers, MA, USA) | 4544 | Rb | - | 1/1000 |
| NFL-P Ser473 | Millipore (Burlington, MA, USA) | MABN2431 | Ms | - | 1/100 |
| p38-P Thr180/Tyr182 | Cell Signaling (Danvers, MA, USA) | 9211 | Rb | - | 1/100 |
| PHF1, tau-P Ser396/Ser404 | Dr. Peter Davies | - | Ms | - | 1/500 |
| PKAα/β-P Tyr197 | Invitrogen (Carlsbad, CA, USA) | 44988 | Rb | 1/100 | |
| SAPK/JNK-P-Thr183/Thr185 | Cell Signaling (Danvers, MA, USA) | 9251 | Rb | - | 1/25 |
| SRC-P Tyr416 | Millipore (Barcelona, Spain) | 05-677 | Ms | - | 1/100 |
| SRPK1 | Abcam (Cambridge, UK) | ab90527 | Rb | - | 1/100 |
| SRPK2-P Ser497 | Affbiotech (Bionova, Spain) | 3632 | Rb | - | 1/100 |
| tau 5 | Thermo Scientific (Barcelona, Spain) | MA5-12808 | Ms | 1/500 | - |
| tau AT8-P Ser202/Thr205 | Innogenetics (Barcelona, Spain) | 90206 | Ms | - | 1/50 |
| tau C-3 | Millipore (Burlington, MA, USA) | 36-017 | Ms | - | 1/100 |
| tau MC1 | Dr. Peter Davies | - | Ms | - | 1/20 |
| tau-N Tyr29 | Millipore (Burlington, MA, USA) | MAB2244 | Ms | - | 1/200 |
| tau-P Ser422 | Thermo Scientific (Barcelona, Spain) | 44764 | Rb | - | 1/50 |
| tau-P Thr181 | Thermo Scientific (Barcelona, Spain) | PA1-14413 | Rb | 1/1000 | - |
| Ubiquitin | Dako (Barcelona, Spain) | Z0458 | Ms | - | 1/250 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrés-Benito, P.; Carmona, M.; Jordán, M.; Fernández-Irigoyen, J.; Santamaría, E.; del Rio, J.A.; Ferrer, I. Host Tau Genotype Specifically Designs and Regulates Tau Seeding and Spreading and Host Tau Transformation Following Intrahippocampal Injection of Identical Tau AD Inoculum. Int. J. Mol. Sci. 2022, 23, 718. https://doi.org/10.3390/ijms23020718
Andrés-Benito P, Carmona M, Jordán M, Fernández-Irigoyen J, Santamaría E, del Rio JA, Ferrer I. Host Tau Genotype Specifically Designs and Regulates Tau Seeding and Spreading and Host Tau Transformation Following Intrahippocampal Injection of Identical Tau AD Inoculum. International Journal of Molecular Sciences. 2022; 23(2):718. https://doi.org/10.3390/ijms23020718
Chicago/Turabian StyleAndrés-Benito, Pol, Margarita Carmona, Mónica Jordán, Joaquín Fernández-Irigoyen, Enrique Santamaría, José Antoni del Rio, and Isidro Ferrer. 2022. "Host Tau Genotype Specifically Designs and Regulates Tau Seeding and Spreading and Host Tau Transformation Following Intrahippocampal Injection of Identical Tau AD Inoculum" International Journal of Molecular Sciences 23, no. 2: 718. https://doi.org/10.3390/ijms23020718
APA StyleAndrés-Benito, P., Carmona, M., Jordán, M., Fernández-Irigoyen, J., Santamaría, E., del Rio, J. A., & Ferrer, I. (2022). Host Tau Genotype Specifically Designs and Regulates Tau Seeding and Spreading and Host Tau Transformation Following Intrahippocampal Injection of Identical Tau AD Inoculum. International Journal of Molecular Sciences, 23(2), 718. https://doi.org/10.3390/ijms23020718