
The Effects of Aging on the Intramuscular Connective Tissue

 ,
, 
 ,
,  ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. The Human Participants
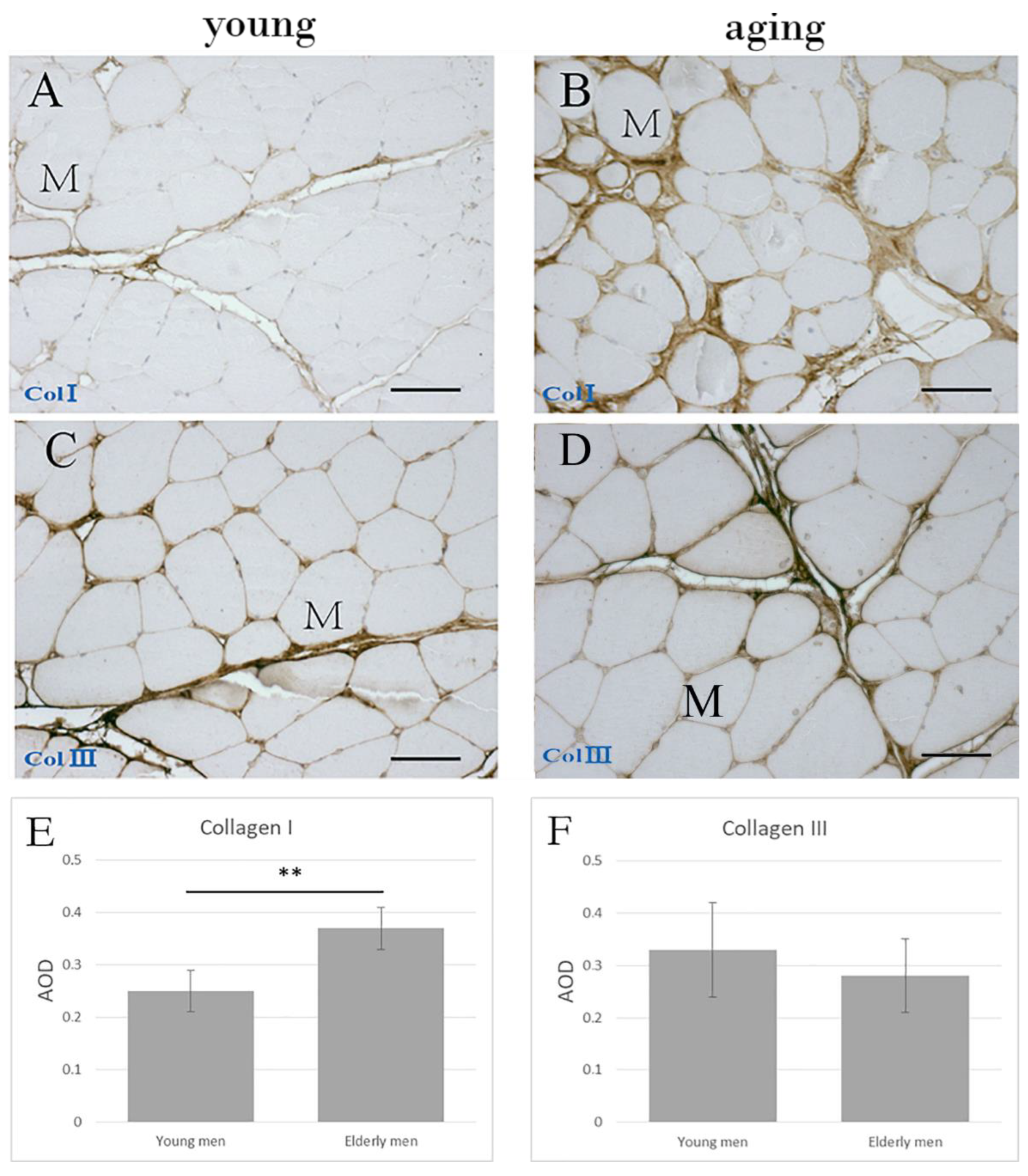
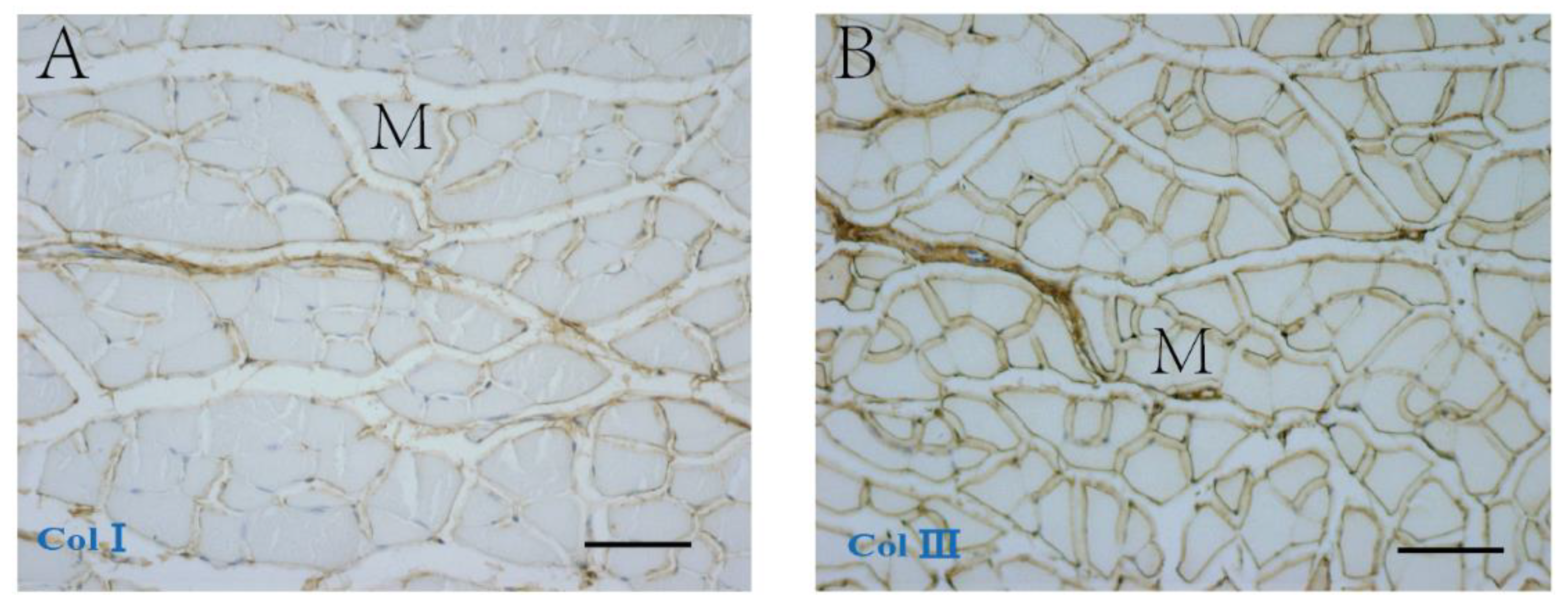
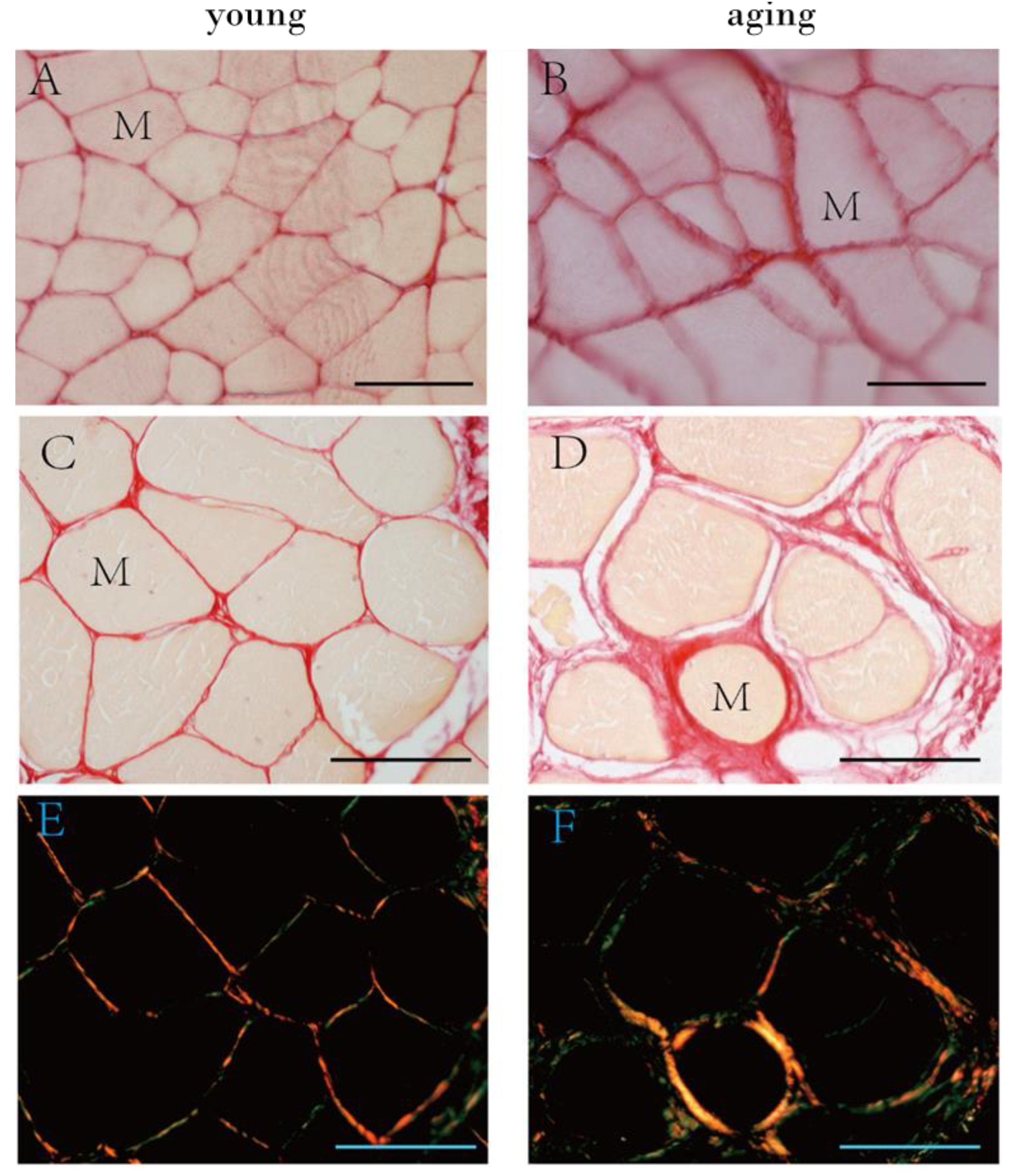
2.2. The Collagen Contents and Elastic Fibers of the ECM in the IMCT
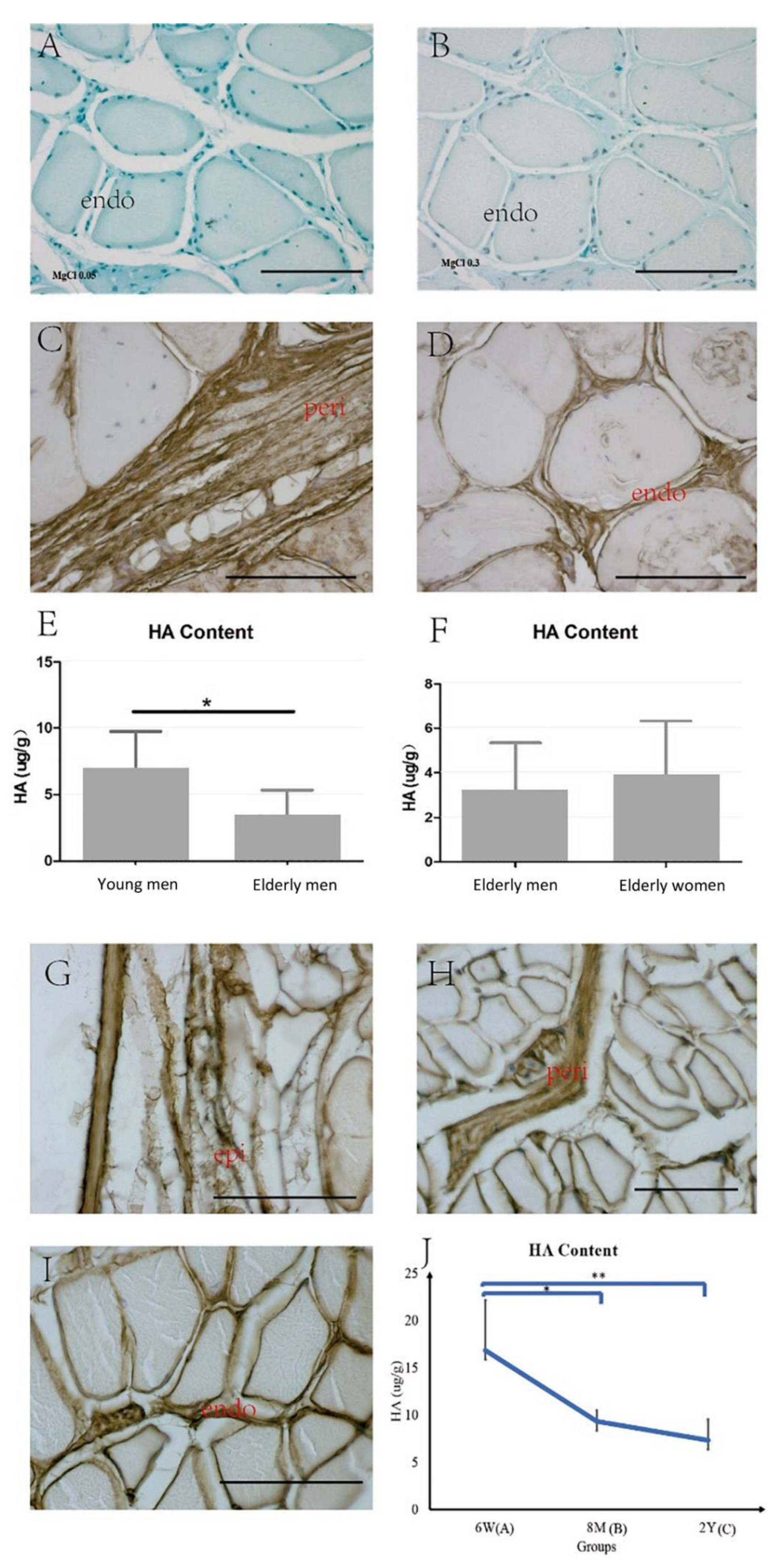
2.3. HA in the ECM
3. Discussion
3.1. Collagen and Elastic Fibers in the IMCT of Skeletal Muscle with Aging
3.2. HA in the IMCT of the Skeletal Muscle with Aging
3.3. The IMCT: A Clinical Viewpoint
3.4. Limitations and Further Research
4. Materials and Methods
4.1. Collection of Human Specimens
4.2. Collection of Animal Samples
4.3. Decalcification Protocol for the Mouse Hindlimbs
4.4. Histological Stainings and Morphological Analysis of the IMCT
4.5. Transmission Electron Microscopy (TEM) and Ultrastructural Analysis of the IMCT
4.6. Immunohistochemistry Staining: Analysis of Collagen Type I (COLI), Collagen Type III (COLIII) and Hyaluronic Acid Binding Protein (HABP)
4.7. Purple-Jelley HA Assay: Quantification of Hyaluronan in the IMCT
4.8. Image Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vervoort, D.; Buurke, T.J.W.; Vuillerme, N.; Hortobágyi, T.; den Otter, R.; Lamoth, C.J.C. Adaptive Control of Dynamic Balance across the Adult Lifespan. Med. Sci. Sports Exerc. 2020, 52, 2270–2277. [Google Scholar] [CrossRef] [PubMed]
- François, M.; Morice, A.H.; Blouin, J.; Montagne, G. Age-related decline in sensory processing for locomotion and interception. Neuroscience 2011, 172, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Kragstrup, T.W.; Kjaer, M.; Mackey, A. Structural, biochemical, cellular, and functional changes in skeletal muscle extracellular matrix with aging. Scand. J. Med. Sci. Sports 2011, 21, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.C.; Manini, T.M. Functional consequences of sarcopenia and dynapenia in the elderly. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 271. [Google Scholar] [CrossRef] [PubMed]
- Orr, R. Contribution of muscle weakness to postural instability in the elderly. A systematic review. Eur. J. Phys. Rehabil. Med. 2010, 46, 183–220. [Google Scholar] [PubMed]
- Purslow, P.P. The structure and role of intramuscular connective tissue in muscle function. Front. Physiol. 2020, 11, 495. [Google Scholar] [CrossRef]
- Blottner, D.; Huang, Y.; Trautmann, G.; Sun, L. The fascia: Continuum linking bone and myofascial bag for global and local body movement control on Earth and in Space. A scoping review. REACH 2019, 14, 100030. [Google Scholar] [CrossRef]
- Huijing, P.A. Epimuscular myofascial force transmission: A historical review and implications for new research. International Society of Biomechanics Muybridge Award Lecture, Taipei, 2007. J. Biomech. 2009, 42, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Kumka, M.; Bonar, J. Fascia: A morphological description and classification system based on a literature review. J. Can. Chiropr. Assoc. 2012, 56, 179. [Google Scholar]
- Helmbacher, F.; Stricker, S. Tissue cross talks governing limb muscle development and regeneration. Semin. Cell Dev. Biol. 2020, 104, 14–30. [Google Scholar] [CrossRef]
- Contreras, O.; Rossi, F.M.V.; Theret, M. Origins, potency, and heterogeneity of skeletal muscle fibro-adipogenic progenitors—Time for new definitions. Skelet. Muscle 2021, 11, 16. [Google Scholar] [CrossRef]
- Kjær, M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef]
- Fede, C.; Pirri, C.; Fan, C.; Petrelli, L.; Guidolin, D.; De Caro, R.; Stecco, C. A Closer Look at the Cellular and Molecular Components of the Deep/Muscular Fasciae. Int. J. Mol. Sci. 2021, 22, 1411. [Google Scholar] [CrossRef] [PubMed]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef]
- Meyer, G.; Lieber, R.L. Muscle fibers bear a larger fraction of passive muscle tension in frogs compared with mice. J. Exp. Biol. 2018, 221, jeb182089. [Google Scholar] [CrossRef] [PubMed]
- Calve, S.; Simon, H.G. Biochemical and mechanical environment cooperatively regulate skeletal muscle regeneration. FASEB J. 2012, 26, 2538–2545. [Google Scholar] [CrossRef]
- Stearns-Reider, K.M.; D’Amore, A.; Beezhold, K.; Rothrauff, B.; Cavalli, L.; Wagner, W.R.; Vorp, D.A.; Tsamis, S.; Shinde, S.; Zhang, C.; et al. Aging of the skeletal muscle extracellular matrix drives a stem cell fibrogenic conversion. Aging Cell 2017, 16, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Theret, M.; Rossi, F.M.V.; Contreras, O. Evolving Roles of Muscle-Resident Fibro-Adipogenic Progenitors in Health, Regeneration, Neuromuscular Disorders, and Aging. Front. Physiol. 2021, 12, 673404. [Google Scholar] [CrossRef]
- Pavan, P.; Monti, E.; Bondí, M.; Fan, C.; Stecco, C.; Narici, M.; Reggiani, C.; Marcucci, L. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. Int. J. Mol. Sci. 2020, 21, 3992. [Google Scholar] [CrossRef] [PubMed]
- Lukjanenko, L.; Karaz, S.; Stuelsatz, P.; Gurriaran-Rodriguez, U.; Michaud, J.; Dammone, G.; Sizzano, F.; Mashinchian, O.; Ancel, S.; Migliavacca, E.; et al. Aging Disrupts Muscle Stem Cell Function by Impairing Matricellular WISP1 Secretion from Fibro-Adipogenic Progenitors. Cell Stem Cell 2019, 24, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Montes, G.S.; Junqueira, L.C. The use of the Picrosirius-polarization method for the study of the biopathology of collagen. Memórias Inst. Oswaldo Cruz 1991, 86, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.; Dorling, J. Differential staining of acid glycosaminoglycans (mucopolysaccharides) by alcian blue in salt solutions. Histochemie 1965, 5, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, K.S.; Palmer, M.L.; van der Meulen, J.H.; Renoux, A.; Kostrominova, T.J.; Michele, D.E.; Faulkner, J.A. Lateral transmission of force is impaired in skeletal muscles of dystrophic mice and very old rats. J. Physiol. 2011, 589, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, J.S.; He, Q.; He, X.; Basava, R.R.; Hodgson, J.; Sinha, U.; Sinha, S. Microstructural analysis of skeletal muscle force generation during aging. Int. J. Numer. Methods Biomed. Eng. 2020, 36, e3295. [Google Scholar] [CrossRef]
- Haus, J.M.; Carrithers, J.A.; Trappe, S.W.; Trappe, T.A. Collagen, cross-linking, and advanced glycation end products in aging human skeletal muscle. J. Appl. Physiol. (1985) 2007, 103, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Karsdal, M.A.; Genovese, F.; Madsen, E.A.; Manon-Jensen, T.; Schuppan, D. Collagen and tissue turnover as a function of age: Implications for fibrosis. J. Hepatol. 2016, 64, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C. Functional Atlas of the Human Fascial System, 1st ed.; Elsevier Health Sciences: Edinburgh, UK, 2015. [Google Scholar]
- Parkin, J.D.; San Antonio, J.D.; Persikov, A.V.; Dagher, H.; Dalgleish, R.; Jensen, S.T.; Jeunemaitre, X.; Savige, J. The collαgen III fibril has a “flexi-rod” structure of flexible sequences interspersed with rigid bioactive domains including two with hemostatic roles. PLoS ONE 2017, 12, e0175582. [Google Scholar] [CrossRef]
- Rodrigues, C.; Rodrigues Junior, A.J. A comparative study of aging of the elastic fiber system of the diaphragm and the rectus abdominis muscles in rats. Braz. J. Med. Biol. Res. 2000, 33, 1449–1454. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Gao, J.; Pawlyk, B.; Starcher, B.; Spencer, J.A.; Yanagisawa, H.; Zuo, J.; Li, T. Elastic fiber homeostasis requires lysyl oxidase–like 1 protein. Nat. Genet. 2004, 36, 178–182. [Google Scholar] [CrossRef]
- Leng, Y.; Abdullah, A.; Wendt, M.K.; Calve, S. Hyaluronic acid, CD44 and RHAMM regulate myoblast behavior during embryogenesis. Matrix Biol. 2019, 78, 236–254. [Google Scholar] [CrossRef]
- Wang, W.; Pan, H.; Murray, K.; Jefferson, B.S.; Li, Y. Matrix metalloproteinase-1 promotes muscle cell migration and differentiation. Am. J. Pathol. 2009, 174, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Necas, J.; Bartosikova, L.; Brauner, P.; Kolar, J. Hyaluronic acid (hyaluronan): A review. Vet. Med. 2008, 53, 397–411. [Google Scholar] [CrossRef]
- Meyer, L.J.; Stern, R. Age-dependent changes of hyaluronan in human skin. J. Investig. Dermatol. 1994, 102, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Legouffe, R.; Jeanneton, O.; Gaudin, M.; Tomezyk, A.; Gerstenberg, A.; Dumas, M.; Heusèle, C.; Bonnel, D.; Stauber, J.; Schnebert, S. Hyaluronic acid detection and relative quantification by mass spectrometry imaging in human skin tissues. Anal. Bioanal. Chem. 2022; Epub ahead of print. [Google Scholar]
- Boyer, K.A.; Johnson, R.T.; Banks, J.J.; Jewell, C.; Hafer, J.F. Systematic review and meta-analysis of gait mechanics in young and older adults. Exp. Gerontol. 2017, 95, 63–70. [Google Scholar] [CrossRef]
- Depalle, B.; Duarte, A.G.; Fiedler, I.A.K.; Pujo-Menjouet, L.; Buehler, M.J.; Berteau, J.P. The different distribution of enzymatic collagen cross-links found in adult and children bone result in different mechanical behavior of collagen. Bone 2018, 110, 107–114. [Google Scholar] [CrossRef]
- Pirri, C.; Petrelli, L.; Pérez-Bellmunt, A.; Ortiz-Miguel, S.; Fede, C.; De Caro, R.; Miguel-Pérez, M.; Stecco, C. Fetal Fascial Reinforcement Development: From “a White Tablet” to a Sculpted Precise Organization by Movement. Biology 2022, 11, 735. [Google Scholar] [CrossRef]
- Fede, C.; Pirri, C.; Fan, C.; Albertin, G.; Porzionato, A.; Macchi, V.; De Caro, R.; Stecco, C. Sensitivity of the fasciae to sex hormone levels: Modulation of collagen-I, collagen-III and fibrillin production. PLoS ONE 2019, 14, e0223195. [Google Scholar] [CrossRef]
- Doessing, S.; Kjaer, M. Growth hormone and connective tissue in exercise. Scand. J. Med. Sci. Sports 2005, 15, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.A.; Manolagas, S.C. Effects of sex steroids on bones and muscles: Similarities, parallels, and putative interactions in health and disease. Bone 2015, 80, 67–78. [Google Scholar] [CrossRef]
- Doessing, S.; Heinemeier, K.M.; Holm, L.; Mackey, A.L.; Schjerling, P.; Rennie, M.; Smith, K.; Reitelseder, S.; Kappelgaard, A.M.; Rasmussen, M.H.; et al. Growth hormone stimulates the collagen synthesis in human tendon and skeletal muscle without affecting myofibrillar protein synthesis. J. Physiol. 2010, 588, 341–351. [Google Scholar] [CrossRef]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix–What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Rezuş, E.; Burlui, A.; Cardoneanu, A.; Rezuş, C.; Codreanu, C.; Pârvu, M.; Rusu Zota, G.; Tamba, B.I. Inactivity and Skeletal Muscle Metabolism: A Vicious Cycle in Old Age. Int. J. Mol. Sci. 2020, 21, 592. [Google Scholar] [CrossRef]
- Fan, C.; Pirri, C.; Fede, C.; Guidolin, D.; Biz, C.; Petrelli, L.; Porzionato, A.; Macchi, V.; De Caro, R.; Stecco, C. Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles. J. Clin. Med. 2021, 11, 86. [Google Scholar] [CrossRef]
- Wood, L.K.; Kayupov, E.; Gumucio, J.P.; Mendias, C.L.; Claflin, D.R.; Brooks, S.V. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. J. Appl. Physiol. 2014, 117, 363–369. [Google Scholar] [CrossRef]
- Stecco, C.; Pirri, C.; Fede, C.; Yucesoy, C.A.; De Caro, R.; Stecco, A. Fascial or Muscle Stretching? A Narrative Review. Appl. Sci. 2021, 11, 307. [Google Scholar] [CrossRef]
- Reeves, N.D.; Narici, M.V.; Maganaris, C.N. Myotendinous plasticity to ageing and resistance exercise in humans. Exp. Physiol. 2006, 91, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Witvrouw, E.; Mahieu, N.; Roosen, P.; McNair, P. The role of stretching in tendon injuries. Br. J. Sports Med. 2007, 41, 224–226. [Google Scholar] [CrossRef]
- Cowman, M.K. Hyaluronan and hyaluronan fragments. In Advances in Carbohydrate Chemistry and Biochemistry; Elsevier: Edinburgh, UK, 2017; Volume 74, pp. 1–59. [Google Scholar]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Stecco, C.; Fede, C.; Macchi, V.; Porzionato, A.; Petrelli, L.; Biz, C.; De Caro, R. The fasciacytes: A new cell devoted to fascial gliding regulation. Clin. Anat. 2018, 31, 667–676. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Y Men (n = 10) | E Men (n = 12) | E Women (n = 16) | Y vs. E Men p-Value | E Men vs. Women p-Value |
|---|---|---|---|---|---|
| Age (y) | 37.5 ±9.5 | 79.0 ±12.4 | 80.56 ± 11.4 | <0.001 *** | 0.931 |
| Sex | M | M | F | NA | NA |
| Muscle | 6VL, 4RF | 8VL, 4RF | 9VL, 7RF | NA | NA |
| Height (cm) | 177.0 ± 6.4 | 176.8 ± 5.0 | 165.1 ± 3.7 | 0.919 | <0.001 *** |
| Weight (kg) | 75.0 ± 6.9 | 77.3 ± 9.3 | 63.9 ± 9.6 | 0.519 | 0.001 *** |
| BMI (kg/m2) | 23.9 ± 1.5 | 24.8 ± 3.1 | 23.5 ± 4.0 | 0.429 | 0.624 |
| Characteristic | Y Men (n = 10) | E Men (n = 12) | E Women (n = 16) | Y vs. E Men p-Value | E Men vs. Women p-Value |
|---|---|---|---|---|---|
| collagen content (% area) (10× magnification) | 5.99 ± 1.34 | 10.0 2 ± 3.69 | 9.90 ± 1.50 | 0.001 *** | 0.927 |
| collagen content (% area) (20× magnification) | 2.95 ± 1.59 | 6.87 ± 1.79 | 6.58 ± 1.12 | 0.001 *** | 0.995 |
| Fat infiltration | 0 | 9 (75%) | 12 (75%) | NA | NA |
| Muscle atrophy | 0 | 10 (83%) | 13 (81.25%) | NA | NA |
| Variable | Y Men (n = 10) | E Men (n = 12) | p-Value |
|---|---|---|---|
| Elastic fiber (% area) | 7.99 ± 1.65 | 3.50 ± 1.66 | 0.001 *** |
| AOD of COLI | 0.25 ± 0.04 | 0.37 ± 0.04 | 0.001 *** |
| AOD of COLIII | 0.33 ± 0.09 | 0.28 ± 0.07 | 0.293 |
| Characteristic | Group A (n = 5) | Group B (n = 5) | Group C (n = 5) | A vs. B p-Value | A vs. C p-Value | B vs. C p-Value |
|---|---|---|---|---|---|---|
| # Collagen content (% area) | 2.05 ± 0.44 | 3.26 ± 0.86 | 7.97 ± 0.80 | 0.071 | <0.001 *** | <0.001 *** |
| # AOD of COLI | 0.17 ± 0.01 | 0.22 ± 0.02 | 0.30 ± 0.03 | 0.013 * | 0.002 ** | <0.001 *** |
| $ AOD of COLIII | 0.23 ± 0.08 | 0.23 ± 0.04 | 0.22 ± 0.05 | 0.994 | 0.897 | 0.982 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fede, C.; Fan, C.; Pirri, C.; Petrelli, L.; Biz, C.; Porzionato, A.; Macchi, V.; De Caro, R.; Stecco, C. The Effects of Aging on the Intramuscular Connective Tissue. Int. J. Mol. Sci. 2022, 23, 11061. https://doi.org/10.3390/ijms231911061
Fede C, Fan C, Pirri C, Petrelli L, Biz C, Porzionato A, Macchi V, De Caro R, Stecco C. The Effects of Aging on the Intramuscular Connective Tissue. International Journal of Molecular Sciences. 2022; 23(19):11061. https://doi.org/10.3390/ijms231911061
Chicago/Turabian StyleFede, Caterina, Chenglei Fan, Carmelo Pirri, Lucia Petrelli, Carlo Biz, Andrea Porzionato, Veronica Macchi, Raffaele De Caro, and Carla Stecco. 2022. "The Effects of Aging on the Intramuscular Connective Tissue" International Journal of Molecular Sciences 23, no. 19: 11061. https://doi.org/10.3390/ijms231911061
APA StyleFede, C., Fan, C., Pirri, C., Petrelli, L., Biz, C., Porzionato, A., Macchi, V., De Caro, R., & Stecco, C. (2022). The Effects of Aging on the Intramuscular Connective Tissue. International Journal of Molecular Sciences, 23(19), 11061. https://doi.org/10.3390/ijms231911061