
Cardioprotective Signaling Pathways in Obese Mice Submitted to Regular Exercise: Effect on Oxysterols

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
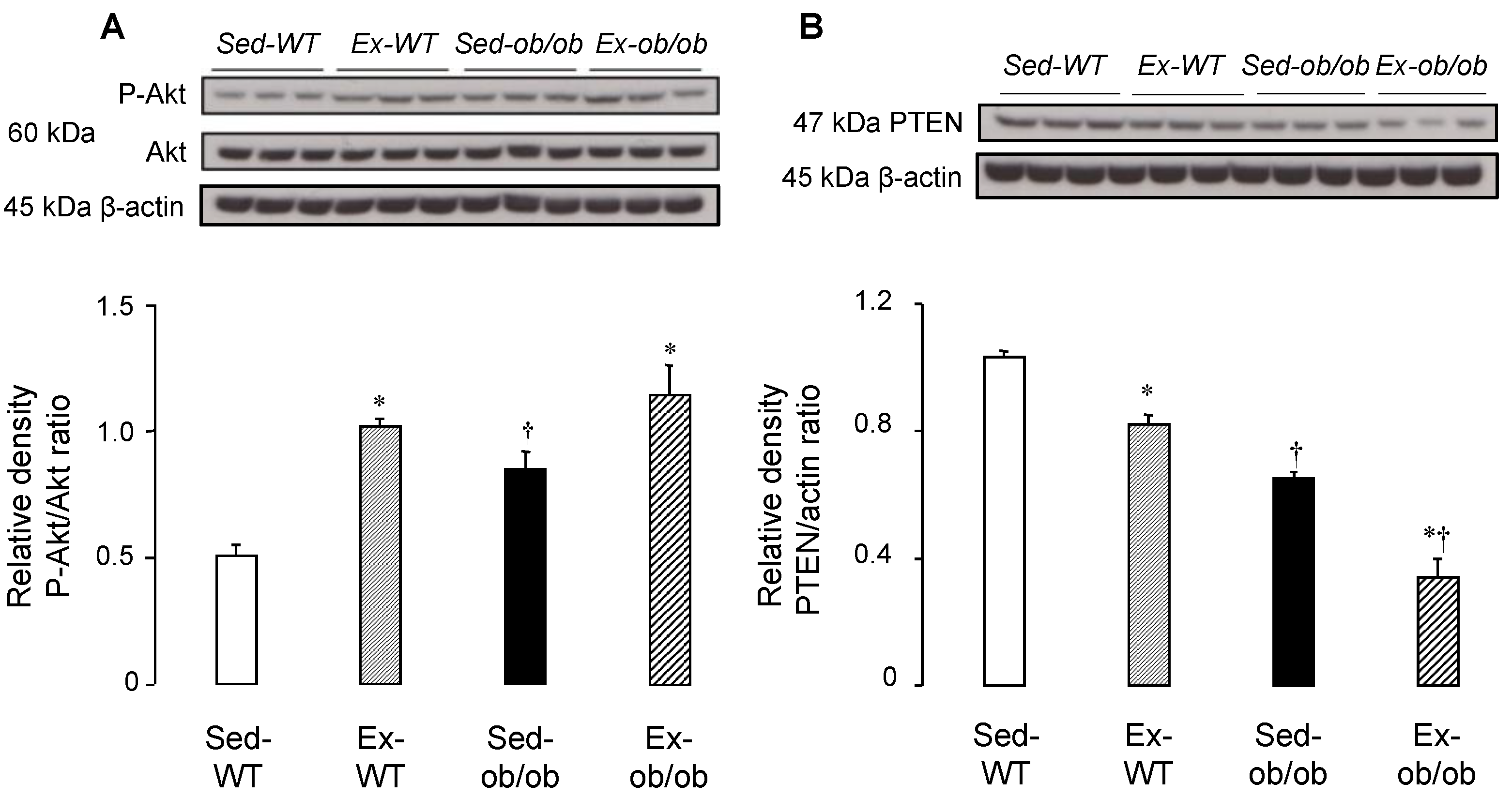
2.1. Effect of Regular Exercise on Signaling Pathways
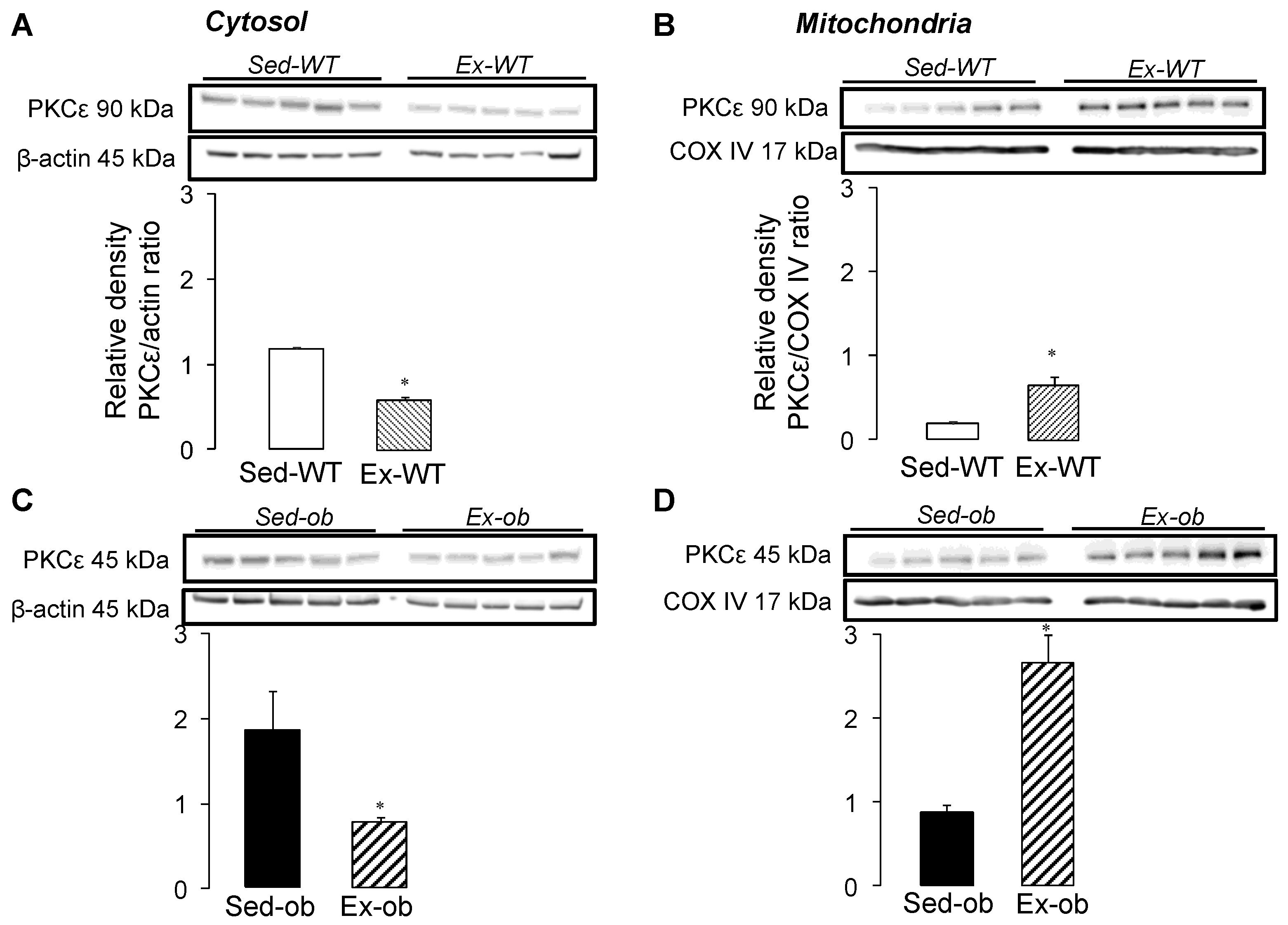
2.2. Translocation of PKCε Following Regular Exercise
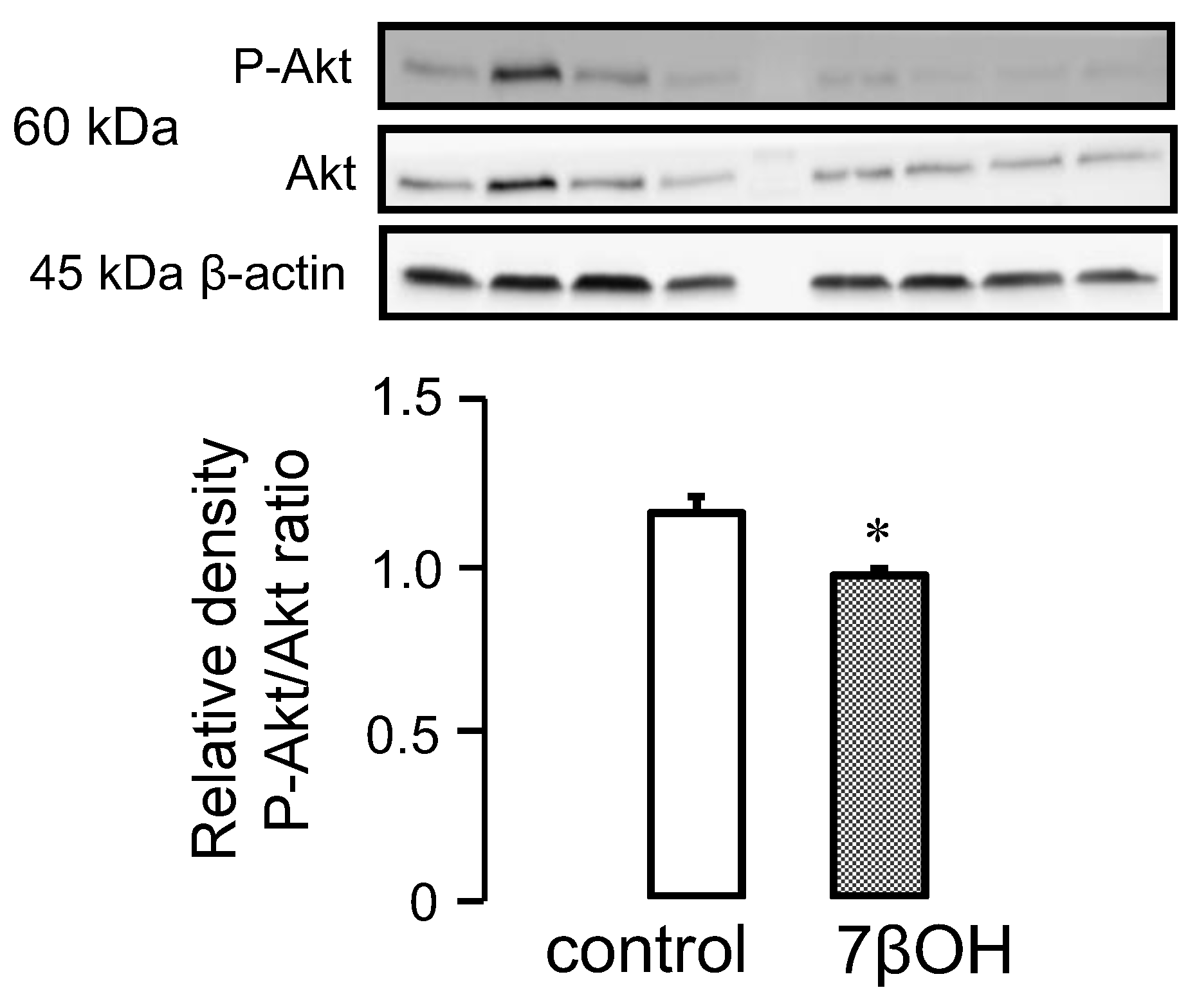
2.3. Effect of Oxysterols on Signaling Pathways
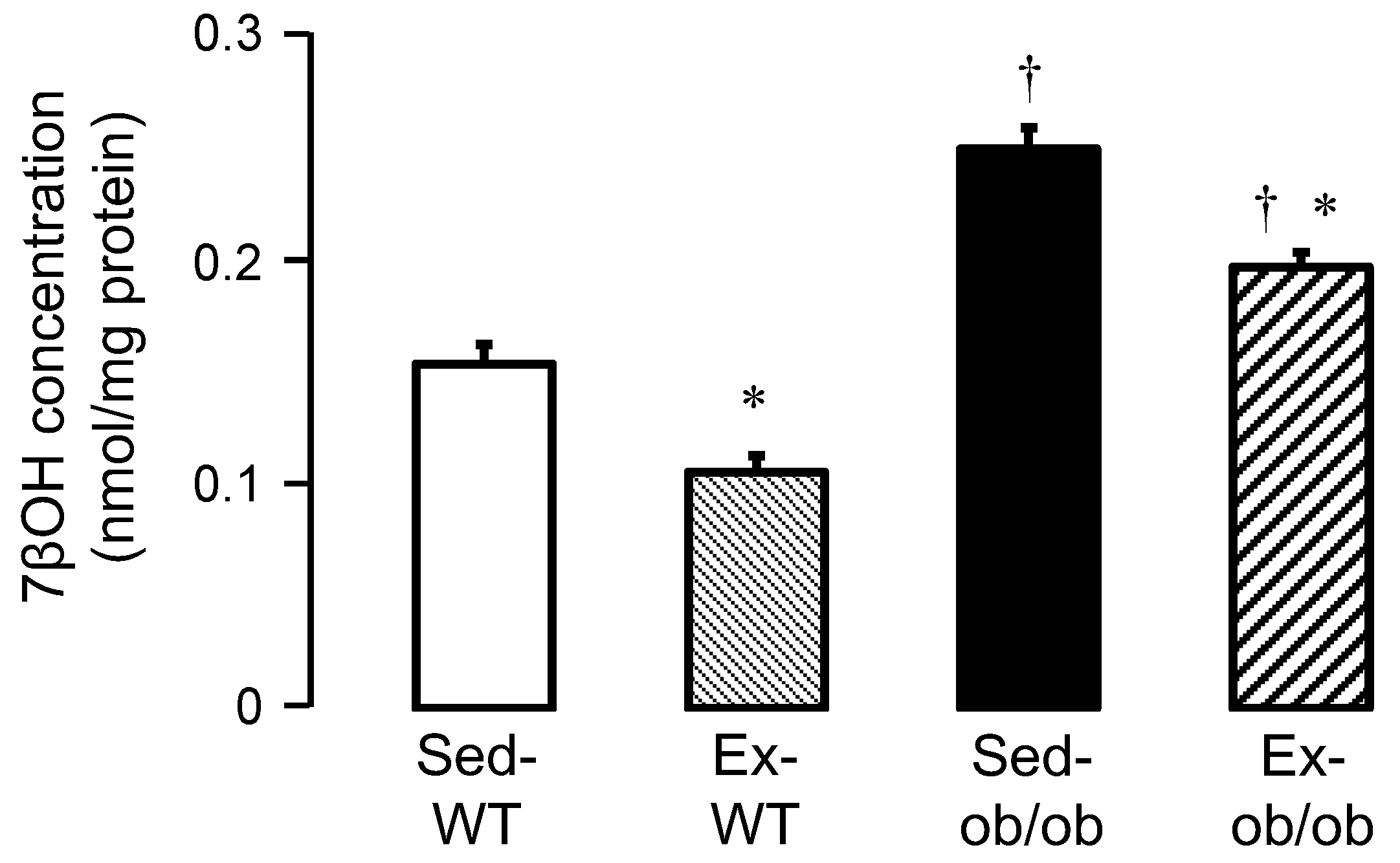
2.4. Effect of Exercise on Oxysterols
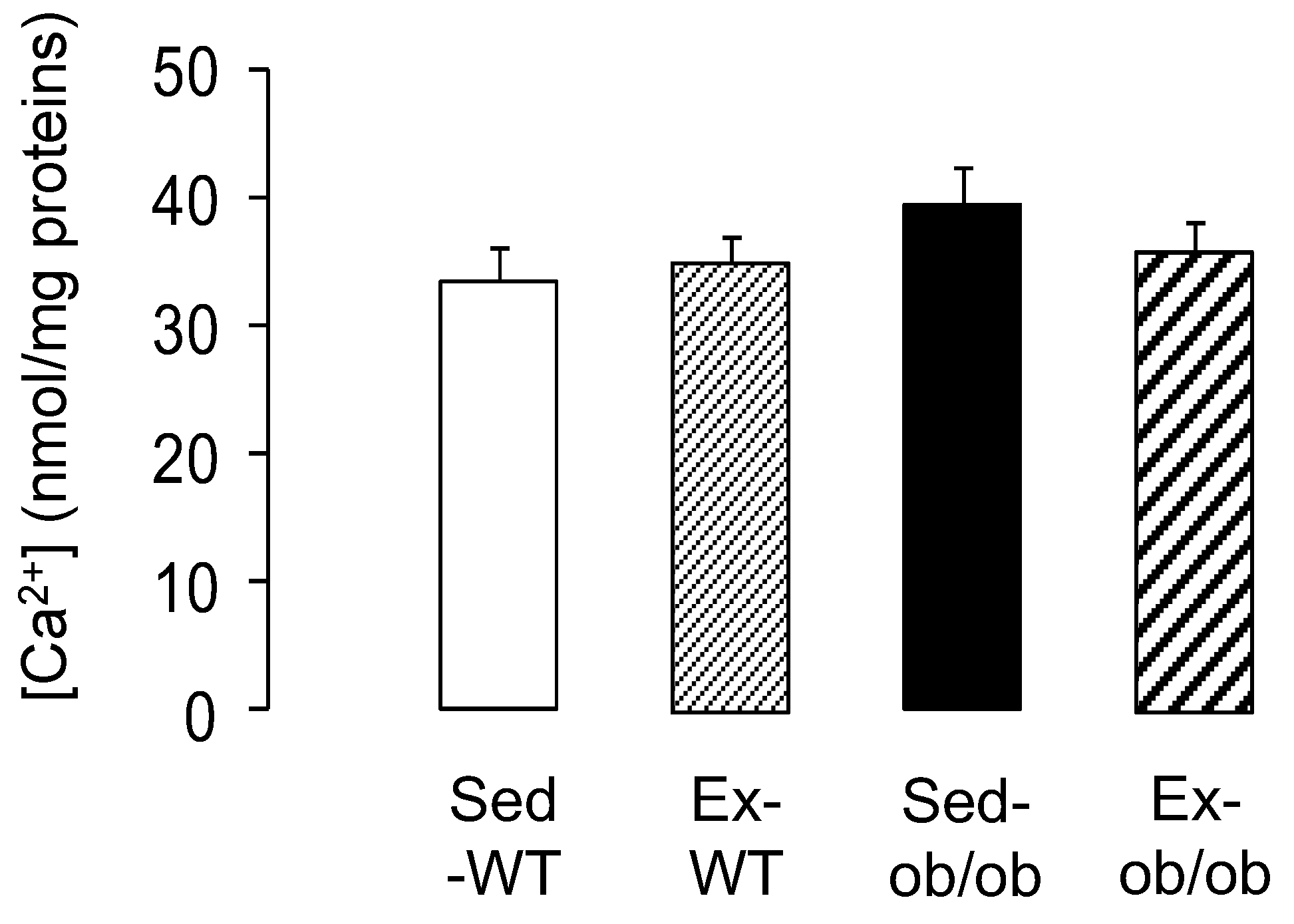
2.5. Effect of Exercice on mPTP Opening
2.6. Oxysterol Concentrations in Healthy Volunteers and Obese Patients
3. Discussion
4. Materials and Methods
4.1. Experimental Protocol in Mice
4.2. Healthy Volunteers and Obese Patients
4.3. Isolation of Adult Mouse Cardiomyocytes
4.4. Cytosol Preparation from Isolated Adult Mice Cardiomyocytes and Human PBMC
4.5. Preparation of Cytosols and Mitochondria from Fresh Ventricular Tissues
4.6. Evaluation of mPTP Opening
4.7. Western Blot Analysis
4.8. Oxysterol Assays
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heusch, G. Obesity—A risk factor or a RISK factor for myocardial infarction? Br. J. Pharmacol. 2006, 149, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Bouhidel, O.; Pons, S.; Souktani, R.; Zini, R.; Berdeaux, A.; Ghaleh, B. Myocardial ischemic postconditioning against ischemia-reperfusion is impaired in ob/ob mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1580–H1586. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D. New directions for protecting the heart against ischaemia–reperfusion injury: Targeting the Reperfusion Injury Salvage Kinase (RISK)-pathway. Cardiovasc. Res. 2004, 61, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D. Cross-talk between the survival kinases during early reperfusion: Its contribution to ischemic preconditioning. Cardiovasc. Res. 2004, 63, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D.J.; Yellon, D.M. Reperfusion injury salvage kinase signalling: Taking a RISK for cardioprotection. Heart Fail. Rev. 2007, 12, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D.J.; Tsang, A.; Mocanu, M.M.; Yellon, D.M. Ischemic preconditioning protects by activating prosurvival kinases at reperfusion. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H971–H976. [Google Scholar] [CrossRef] [PubMed]
- Ping, P.; Zhang, J.; Qiu, Y.; Tang, X.L.; Manchikalapudi, S.; Cao, X.; Bolli, R. Ischemic preconditioning induces selective translocation of protein kinase C isoforms ε and η in the heart of conscious rabbits without subcellular redistribution of total protein kinase C activity. Circ. Res. 1997, 81, 404–414. [Google Scholar] [CrossRef]
- Ytrehus, K.; Liu, Y.; Downey, J.M. Preconditioning protects ischemic rabbit heart by protein kinase C activation. Am. J. Physol. Heart Circ. Physiol. 1994, 266, H1145–H1152. [Google Scholar] [CrossRef]
- Liu, Y.; Ytrehus, K.; Downey, J.M. Evidence that translocation of protein kinase C is a key event during ischemic preconditioning of rabbit myocardium. J. Mol. Cell. Cardiol. 1994, 26, 661–668. [Google Scholar] [CrossRef]
- Kawamura, S.; Yoshida, K.I.; Miura, T.; Mizukami, Y.; Matsuzaki, M. Ischemic preconditioning translocates PKC-δ and -ε, which mediate functional protection in isolated rat heart. Am. J. Physiol. Heart Circ. Physiol. 1998, 275, H2266–H2271. [Google Scholar] [CrossRef]
- Tong, H.; Chen, W.; Steenbergen, C.; Murphy, E. Ischemic preconditioning activates phosphatidylinositol-3-kinase upstream of protein kinase C. Circ. Res. 2000, 87, 309–315. [Google Scholar] [CrossRef]
- Hausenloy, D.; Yellon, D. Survival kinases in ischemic preconditioning and postconditioning. Cardiovasc. Res. 2006, 70, 240–253. [Google Scholar] [CrossRef]
- Downey, J.M.; Davis, A.M.; Cohen, M.V. Signaling pathways in ischemic preconditioning. Heart Fail Rev. 2007, 12, 181–188. [Google Scholar] [CrossRef]
- Hausenloy, D.; Yellon, D. The mitochondrial permeability transition pore: Its fundamental role in mediating cell death during ischaemia and reperfusion. J. Mol. Cell. Cardiol. 2003, 35, 339–341. [Google Scholar] [CrossRef]
- Pons, S.; Martin, V.; Portal, L.; Zini, R.; Morin, D.; Berdeaux, A.; Ghaleh, B. Regular treadmill exercise restores cardioprotective signaling pathways in obese mice independently from improvement in associated co-morbidities. J. Mol. Cell. Cardiol. 2013, 54, 82–89. [Google Scholar] [CrossRef]
- Lordan, S.; Mackrill, J.J.; O’Brien, N.M. Oxysterols and mechanisms of apoptotic signaling: Implications in the pathology of degenerative diseases. J. Nutr. Biochem. 2009, 20, 321–336. [Google Scholar] [CrossRef]
- Musman, J.; Pons, S.; Barau, C.; Caccia, C.; Leoni, V.; Berdeaux, A.; Ghaleh, B.; Morin, D. Regular treadmill exercise inhibits mitochondrial accumulation of cholesterol and oxysterols during myocardial ischemia-reperfusion in wild-type and ob/ob mice. Free Radic. Biol. Med. 2016, 101, 317–324. [Google Scholar] [CrossRef]
- Lordan, S.; O’Neill, C.; O’Brien, N.M. Effects of apigenin, lycopene and astaxanthin on 7β-hydroxycholesterol-induced apoptosis and Akt phosphorylation in U937 cells. Br. J. Nutr. 2008, 100, 287–296. [Google Scholar] [CrossRef]
- Clarion, L.; Schindler, M.; de Weille, J.; Lolmède, K.; Laroche-Clary, A.; Uro-Coste, E.; Robert, J.; Mersel, M.; Bakalara, N. 7β-Hydroxycholesterol-induced energy stress leads to sequential opposing signaling responses and to death of c6 glioblastoma cells. Biochem. Pharmacol. 2012, 83, 37–46. [Google Scholar] [CrossRef]
- Boonsong, T.; Norton, L.; Chokkalingam, K.; Jewell, K.; Macdonald, I.; Bennett, A.; Tsintzas, K. Effect of exercise and insulin on SREBP-1c expression in human skeletal muscle: Potential roles for the ERK1/2 and Akt signalling pathways. Biochem. Soc. Trans. 2007, 35, 1310–1311. [Google Scholar] [CrossRef]
- Sakamoto, K.; Aschenbach, W.G.; Hirshman, M.F.; Goodyear, L.J. Akt signaling in skeletal muscle: Regulation by exercise and passive stretch. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1081–E1088. [Google Scholar] [CrossRef]
- Cao, S.; Li, B.; Yi, X.; Chang, B.; Zhu, B.; Lian, Z.; Zhang, Z.; Zhao, G.; Liu, H.; Zhang, H.; et al. Effects of exercise on AMPK signaling and downstream components to PI3K in rat with type 2 diabetes. PLoS ONE 2012, 7, e51709. [Google Scholar] [CrossRef]
- Fenton, R.A.; Dickson, E.W.; Dobson, J.G. Inhibition of phosphatase activity enhances preconditioning and limits cell death in the ischemic/reperfused aged rat heart. Life Sci. 2005, 77, 3375–3388. [Google Scholar] [CrossRef]
- Przyklenk, K.; Maynard, M.; Darling, C.E.; Whittaker, P. Aging mouse hearts are refractory to infarct size reduction with post-conditioning. J. Am. Coll. Cardiol. 2008, 51, 1393–1398. [Google Scholar] [CrossRef]
- Weinbrenner, C.; Baines, C.P.; Liu, G.S.; Armstrong, S.C.; Ganote, C.E.; Walsh, A.H.; Honkanen, R.E.; Cohen, M.V.; Downey, J.M. Fostriecin, an inhibitor of protein phosphatase 2A, limits myocardial infarct size even when administered after onset of ischemia. Circulation 1998, 98, 899–905. [Google Scholar] [CrossRef]
- Armstrong, S.; Gao, W.; Lane, J.; Ganote, C. Protein phosphatase inhibitors calyculin A and fostriecin protect rabbit cardiomyocytes in late ischemia. J. Mol. Cell. Cardiol. 1998, 30, 61–73. [Google Scholar] [CrossRef]
- Sghaier, R.; Zarrouk, A.; Nury, T.; Badreddine, I.; O’Brien, N.; Mackrill, J.J.; Vejux, A.; Samadi, M.; Nasser, B.; Caccia, C.; et al. Biotin attenuation of oxidative stress, mitochondrial dysfunction, lipid metabolism alteration and 7β-hydroxycholesterol-induced cell death in 158N murine oligodendrocytes. Free Radic. Res. 2019, 53, 535–561. [Google Scholar] [CrossRef]
- Ghzaiel, I.; Zarrouk, A.; Essadek, S.; Martine, L.; Hammouda, S.; Yammine, A.; Ksila, M.; Nury, T.; Meddeb, W.; Tahri Joutey, M.; et al. Protective effects of milk thistle (Sylibum marianum) seed oil and α-tocopherol against 7β-hydroxycholesterol-induced peroxisomal alterations in murine C2C12 myoblasts: Nutritional insights associated with the concept of pexotherapy. Steroids 2022, 183, 109032. [Google Scholar] [CrossRef]
- Melling, C.W.J.; Thorp, D.B.; Milne, K.J.; Noble, E.G. Myocardial Hsp70 phosphorylation and PKC-mediated cardioprotection following exercise. Cell Stress Chaperones 2009, 14, 141–150. [Google Scholar] [CrossRef]
- Carson, L.D.; Korzick, D.H. Dose-dependent effects of acute exercise on PKC levels in rat heart: Is PKC the heart’s prophylactic? Signal transduction and exercise-induced cardioprotection. Acta Physiol. Scand. 2003, 178, 97–106. [Google Scholar] [CrossRef]
- Korge, P.; Honda, H.M.; Weiss, J.N. Protection of cardiac mitochondria by diazoxide and protein kinase C: Implications for ischemic preconditioning. Proc. Natl. Acad. Sci. USA 2002, 99, 3312–3317. [Google Scholar] [CrossRef] [PubMed]
- Baines, C.P.; Song, C.X.; Zheng, Y.T.; Wang, G.W.; Zhang, J.; Wang, O.L.; Guo, Y.; Bolli, R.; Cardwell, E.M.; Ping, P. Protein Kinase Cε interacts with and inhibits the permeability transition pore in cardiac mitochondria. Circ. Res. 2003, 92, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Khazaei, M.; Moien-Afshari, F.; Ang, L.S.; Granville, D.J.; Verchere, C.B.; Dunn, S.R.; Mc Cue, P.; Mizisin, A.; Sharma, K.; et al. Moderate exercise attenuates caspase-3 activity, oxidative stress, and inhibits progression of diabetic renal disease in db/db mice. Am. J. Physiol. Ren. Physiol. 2009, 296, F700–F708. [Google Scholar] [CrossRef] [PubMed]
- Starnes, J.W.; Barnes, B.D.; Olsen, M.E. Exercise training decreases rat heart mitochondria free radical generation but does not prevent Ca2+-induced dysfunction. J. Appl. Physiol. 2007, 102, 1793–1798. [Google Scholar] [CrossRef] [PubMed]
- Marcil, M.; Bourduas, K.; Ascah, A.; Burelle, Y. Exercise training induces respiratory substrate-specific decrease in Ca2+-induced permeability transition pore opening in heart mitochondria. Am. J. Physiol. Heart Circ Physiol. 2006, 290, H1549–H1557. [Google Scholar] [CrossRef] [PubMed]
- Ziedén, B.; Kaminskas, A.; Kristenson, M.; Kucinskienê, Z.; Vessby, B.; Olsson, A.G.; Diczfalusy, U. Increased plasma 7β-hydroxycholesterol concentrations in a population with a high risk for cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Tremblay-Franco, M.; Zerbinati, C.; Pacelli, A.; Palmaccio, G.; Lubrano, C.; Ducheix, S.; Guillou, H.; Iuliano, L. Effect of obesity and metabolic syndrome on plasma oxysterols and fatty acids in human. Steroids 2015, 99, 287–292. [Google Scholar] [CrossRef]
- Massett, M.P.; Berk, B.C. Strain-dependent differences in responses to exercise training in inbred and hybrid mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1006–R1013. [Google Scholar] [CrossRef]
- Leoni, V.; Caccia, C. Oxysterols as biomarkers in neurodegenerative diseases. Chem. Phys. Lipids 2011, 164, 515–524. [Google Scholar] [CrossRef]
- Dzeletovic, S.; Breuer, O.; Lund, E.; Diczfalusy, U. Determination of cholesterol oxidation products in human plasma by isotope dilution-mass spectrometry. Anal. Biochem. 1995, 225, 73–80. [Google Scholar] [CrossRef]
- Paradis, S.; Leoni, V.; Caccia, C.; Berdeaux, A.; Morin, D. Cardioprotection by the TSPO ligand 4′-chlorodiazepam is associated with inhibition of mitochondrial accumulation of cholesterol at reperfusion. Cardiovasc. Res. 2013, 98, 420–427. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barau, C.; Leick, S.; Caccia, C.; Portal, L.; Leoni, V.; Le Corvoisier, P.; Morin, D.; Ghaleh, B.; Pons, S. Cardioprotective Signaling Pathways in Obese Mice Submitted to Regular Exercise: Effect on Oxysterols. Int. J. Mol. Sci. 2022, 23, 10840. https://doi.org/10.3390/ijms231810840
Barau C, Leick S, Caccia C, Portal L, Leoni V, Le Corvoisier P, Morin D, Ghaleh B, Pons S. Cardioprotective Signaling Pathways in Obese Mice Submitted to Regular Exercise: Effect on Oxysterols. International Journal of Molecular Sciences. 2022; 23(18):10840. https://doi.org/10.3390/ijms231810840
Chicago/Turabian StyleBarau, Caroline, Shirin Leick, Claudio Caccia, Lolita Portal, Valerio Leoni, Philippe Le Corvoisier, Didier Morin, Bijan Ghaleh, and Sandrine Pons. 2022. "Cardioprotective Signaling Pathways in Obese Mice Submitted to Regular Exercise: Effect on Oxysterols" International Journal of Molecular Sciences 23, no. 18: 10840. https://doi.org/10.3390/ijms231810840
APA StyleBarau, C., Leick, S., Caccia, C., Portal, L., Leoni, V., Le Corvoisier, P., Morin, D., Ghaleh, B., & Pons, S. (2022). Cardioprotective Signaling Pathways in Obese Mice Submitted to Regular Exercise: Effect on Oxysterols. International Journal of Molecular Sciences, 23(18), 10840. https://doi.org/10.3390/ijms231810840