
Genome Editing of Golden SNP-Carrying Lycopene Epsilon-Cyclase (LcyE) Gene Using the CRSPR-Cas9/HDR and Geminiviral Replicon System in Rice

 ,
, 
 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
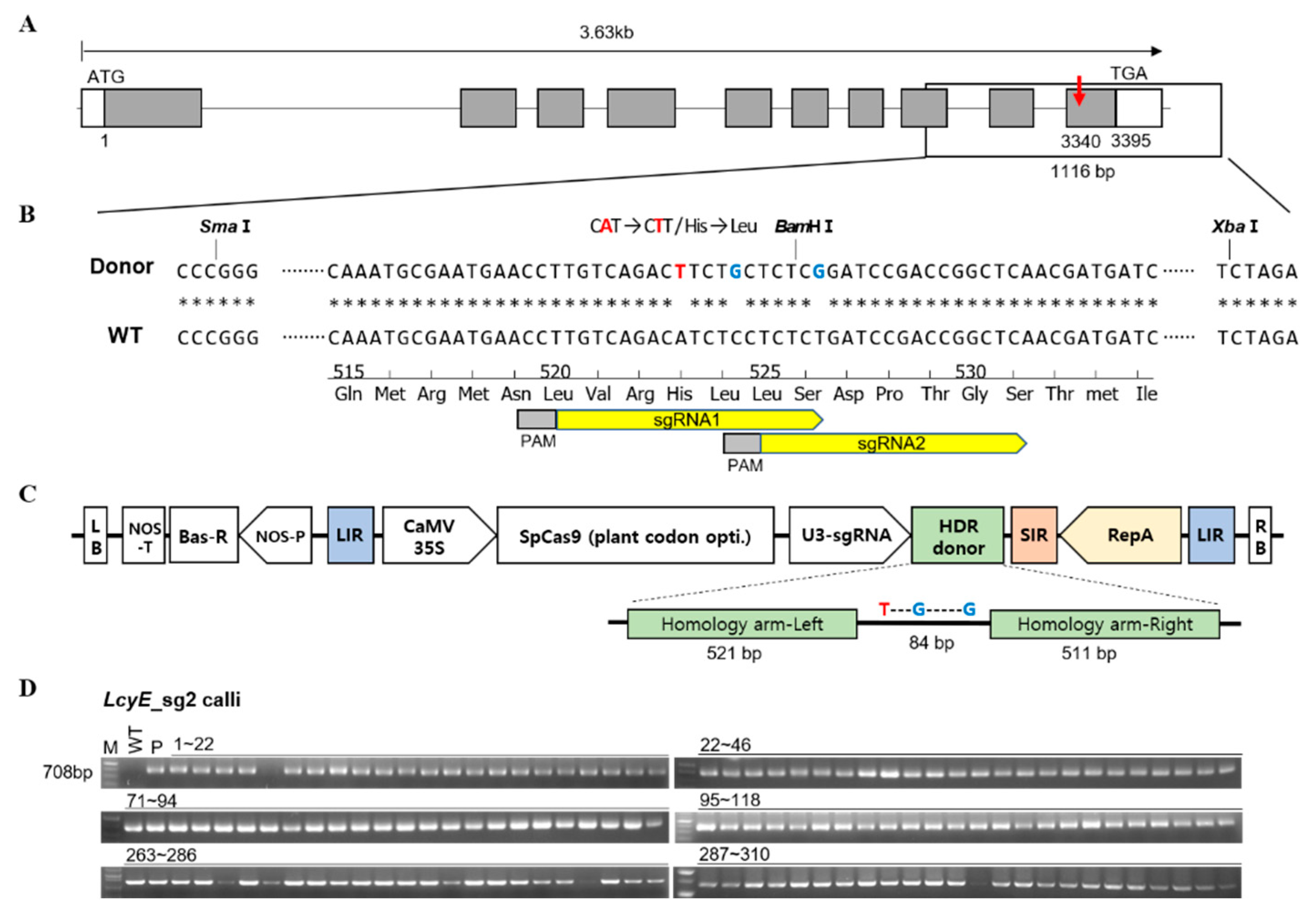
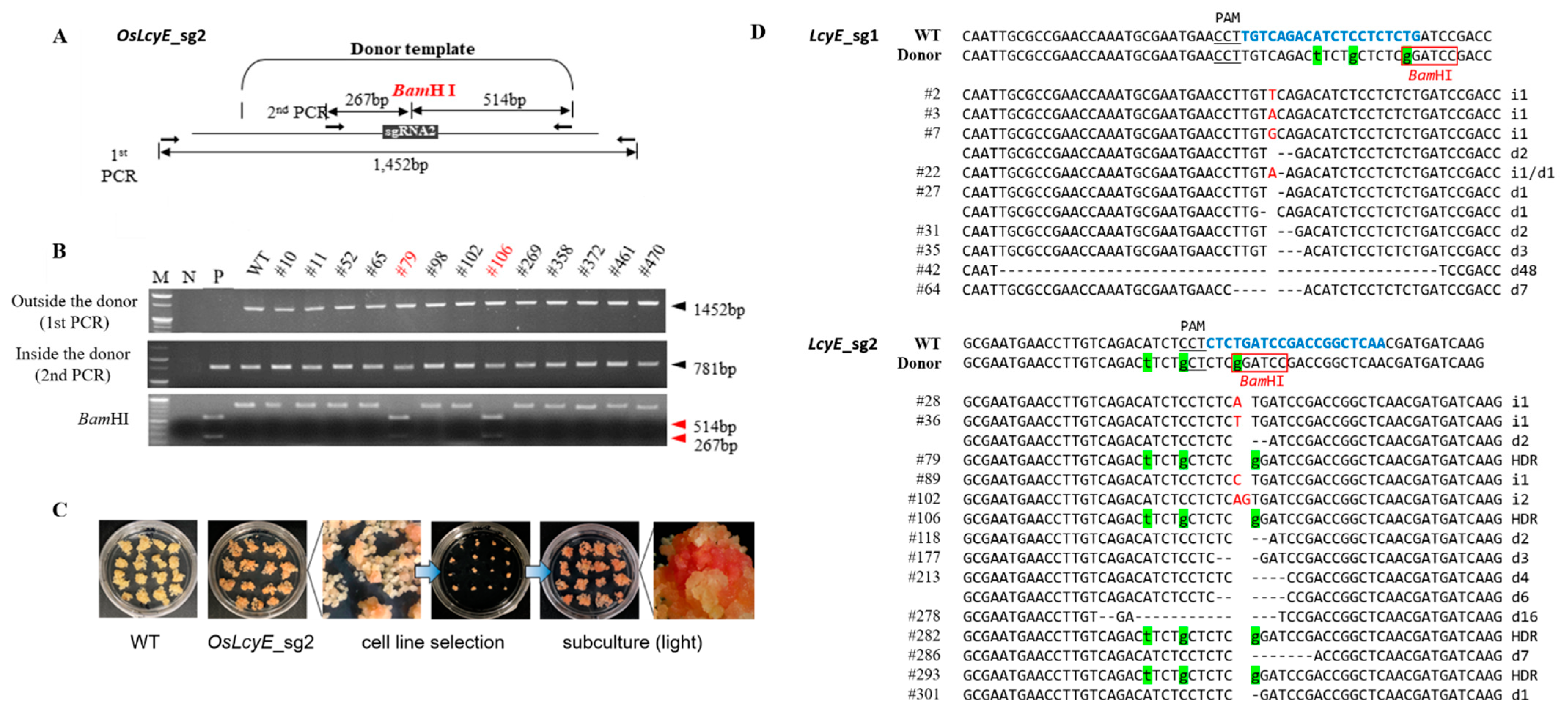
2.1. Efficient Allelic Replacement by the Geminiviral Replicon for Targeted Modifications
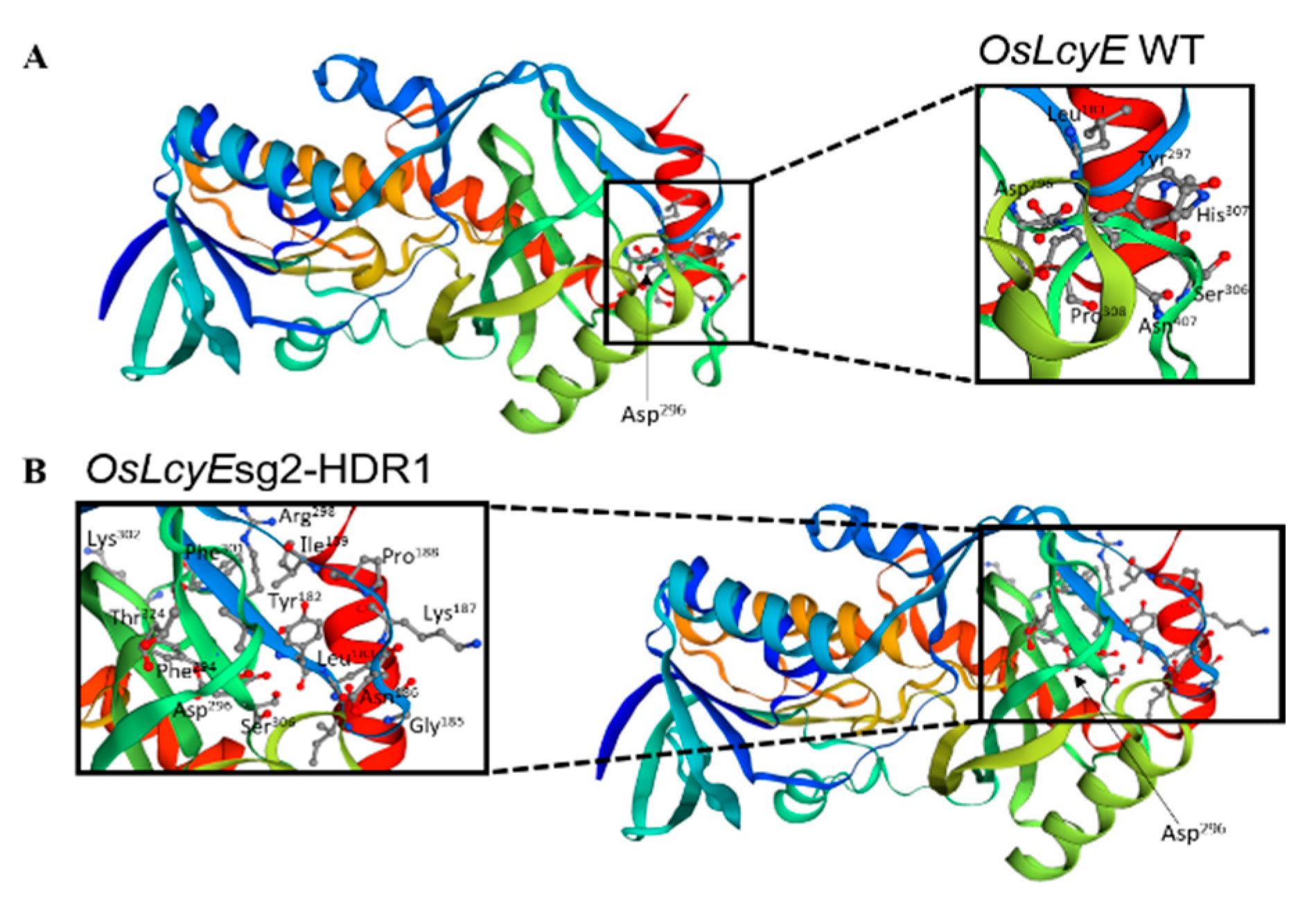
2.2. Protein Model According to the Golden SNP Replacement of OsLcyE
2.3. Carotenoid Analysis in the LcyEsg2-HDR1 and LcyEsg2-HDR2 Lines
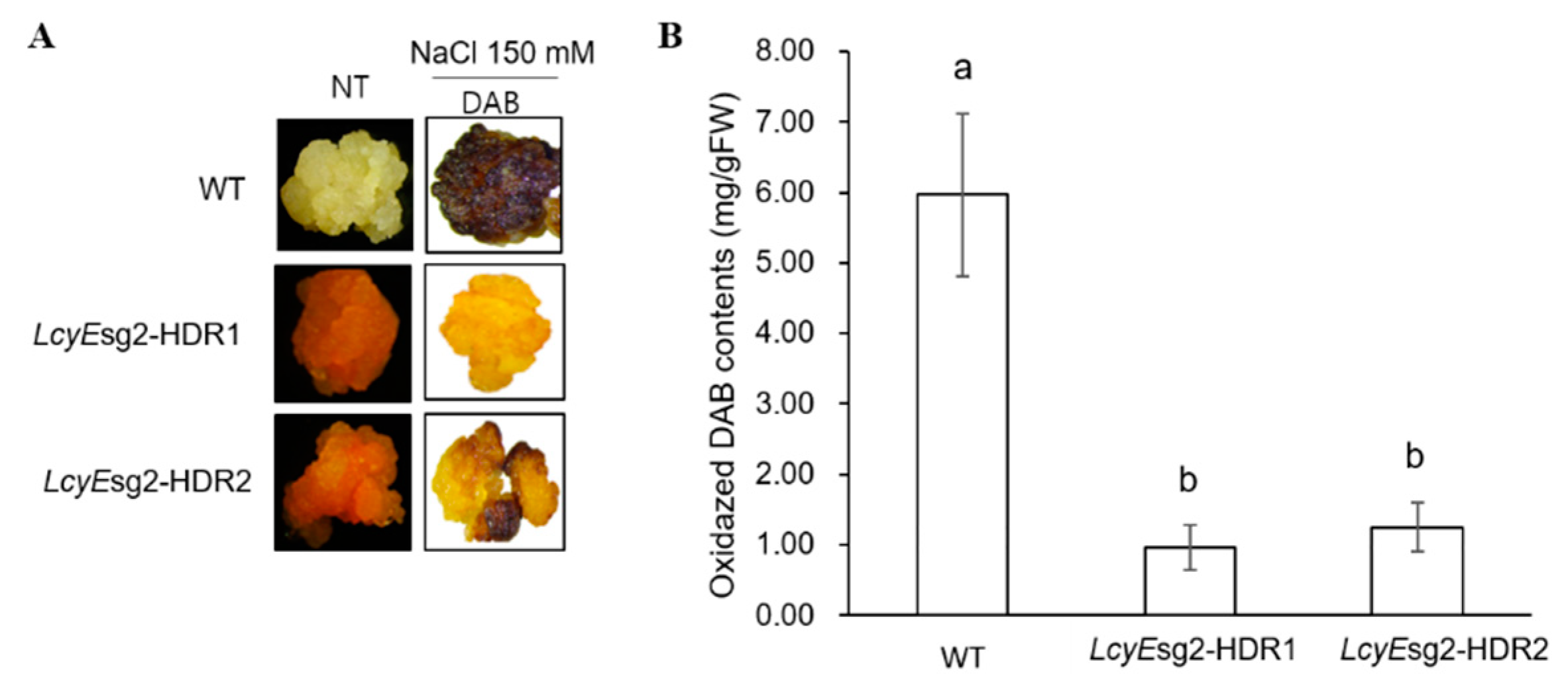
2.4. Reduced ROS Accumulation under Salt Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Construction of Gene-Editing Vectors and Agrobacterium-Mediated Transformation
4.2. Total DNA Extraction and Mutation Detection
4.3. Phenotypic Observations of the Golden SNP Replacement LcyE Mutant Calli
4.4. Homology Modeling and Protein Structure Analysis
4.5. Extraction of Pigments and Measurement of Absorption Spectra
4.6. Analysis of Carotenoid Contents
4.7. H2O2 Assays
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.-L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.L.; Zhang, Q.Y.; Zhu, Q.L.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.F.; Li, H.; Lin, Y.R.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lv, X.; Ye, L.; Zhou, P.; Yu, H. Construction of lycopene-overproducing Saccharomyces cerevisiae by combining directed evolution and metabolic engineering. Metab. Eng. 2015, 30, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free Arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Lee, G.J.; Bae, S.; Kang, K.K. Reduced ethylene production in tomato fruits upon CRSPR/Cas9-mediated leMADS-RIN mutagenesis. Hortic. Sci. Technol. 2018, 36, 396–405. [Google Scholar]
- Jung, Y.J.; Lee, H.J.; Bae, S.; Kim, J.H.; Kim, D.H.; Kim, H.K.; Nam, K.H.; Nogoy, F.M.; Cho, Y.-G.; Kang, K.K. Acquisition of seed dormancy breaking in rice (Oryza sativa L.) via CRISPR/Cas9-targeted mutagenesis of OsVP1 gene. Plant Biotechnol. Rep. 2019, 13, 511–520. [Google Scholar] [CrossRef]
- Jung, Y.J.; Kim, J.H.; Lee, H.J.; Kim, D.H.; Yu, J.; Bae, S.; Cho, Y.-G.; Kang, K.K. Generation and transcriptome profiling of Slr1-d7 and Slr1-d8 mutant lines with a new semi-dominant dwarf allele of SLR1 using the CRISPR/Cas9 system in rice. Int. J. Mol. Sci. 2020, 21, 5492. [Google Scholar] [CrossRef]
- Barrangou, R.; Doudna, J.A. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016, 34, 933–941. [Google Scholar] [CrossRef]
- Schaeffer, S.M.; Nakata, P.A. The expanding footprint of CRISPR/Cas9 in the plant sciences. Plant Cell Rep. 2016, 35, 1451–1468. [Google Scholar] [CrossRef]
- Pacher, M.; Puchta, H. From classical mutagenesis to nuclease-based breeding–directing natural DNA repair for a natural end-product. Plant J. 2017, 90, 819–833. [Google Scholar] [CrossRef]
- Shaked, H.; Melamed-Bessudo, C.; Levy, A.A. High-frequency gene targeting in Arabidopsis plants expressing the yeast RAD54 gene. Proc. Natl. Acad. Sci. USA 2005, 102, 12265–12269. [Google Scholar] [CrossRef] [PubMed]
- Even-Faitelson, L.; Samach, A.; Melamed-Bessudo, C.; Avivi-Ragolsky, N.; Levy, A.A. Localized egg-cell expression of effector proteins for targeted modification of the Arabidopsis genome. Plant J. 2011, 68, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.-I.; Abe, K.; Osakabe, K.; Endo, M.; Nishizawa-Yokoi, A.; Saika, H.; Shimada, H.; Toki, S. Overexpression of OsRecQl4 and/or OsExo1 enhances DSB-induced homologous recombination in rice. Plant Cell Physiol. 2012, 53, 2142–2152. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, B.; Chen, C.Y.; Yang, B. TALEN-mediated homologous recombination produces site-directed DNA base change and herbicide-resistant rice. J. Genet. Genom. 2016, 43, 297–305. [Google Scholar] [CrossRef]
- Wang, M.; Lu, Y.; Botella, J.R.; Mao, Y.; Hua, K.; Zhu, J.K. Gene targeting by homology-directed repair in rice using a geminivirus-based CRISPR/Cas9 system. Mol. Plant 2017, 10, 1007–1010. [Google Scholar] [CrossRef]
- Butler, N.M.; Baltes, N.J.; Voytas, D.F.; Douches, D.S. Geminivirus-mediated genome editing in potato (Solanum tuberosum L.) using sequence-specific nucleases. Front. Plant Sci. 2016, 7, 1045. [Google Scholar] [CrossRef]
- Gil-Humanes, J.; Wang, Y.; Liang, Z.; Shan, Q.; Ozuna, C.V.; Sánchez-León, S.; Baltes, N.J.; Starker, C.; Barro, F.; Gao, C.; et al. High-efficiency gene targeting in hexaploid wheat using DNA replicons and CRISPR/Cas9. Plant J. 2017, 89, 1251–1262. [Google Scholar] [CrossRef]
- Hummel, A.W.; Chauhan, R.D.; Cermak, T.; Mutka, A.M.; Vijayaraghavan, A.; Boyher, A.; Starker, C.G.; Bart, R.; Voytas, D.F.; Taylor, N.J. Allele exchange at the EPSPS locus confers glyphosate tolerance in cassava. Plant Biotechnol. J. 2018, 16, 1275–1282. [Google Scholar] [CrossRef]
- Song, W.; Wei, F.; Gao, S.; Dong, C.; Hao, J.; Jin, L.; Li, F.; Wei, P.; Guo, J.; Wang, R. Functional characterization and comparison of lycopene epsilon-cyclase genes in Nicotiana tabacum. BMC Plant Biol. 2022, 22, 252. [Google Scholar] [CrossRef]
- Wakasa, K.; Kobayashi, M.; Kamada, H. Colony formation from protoplasts of nitrate reductase-deficient rice cell lines. J. Plant Physiol. 1984, 117, 223–231. [Google Scholar] [CrossRef]
- Ishihara, A.; Ohishi, K.; Yamada, T.; Shibata-Hatta, M.; Arai-Kichise, Y.; Watanabe, S.; Yoshikawa, H.; Wakasa, K. Biochemical and molecular characterization of orange-and tangerine-colored rice calli. Plant Biotechnol. 2015, 32, 193–203. [Google Scholar] [CrossRef]
- Dahan-Meir, T.; Filler-Hayut, S.; Melamed-Bessudo, C.; Bocobza, S.; Czosnek, H.; Aharoni, A.; Levy, A.A. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant J. 2018, 95, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.; Nekrasov, V.; Lippman, Z.B.; Van Eck, J. Efficient gene editing in tomato in the first generation using the clustered regularly interspaced short palindromic repeats/CRISPR-associated9 system. Plant Physiol. 2014, 166, 1292–1297. [Google Scholar] [CrossRef]
- Pan, C.; Ye, L.; Qin, L.; Liu, X.; He, Y.; Wang, J.; Chen, L.; Lu, G. CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations. Sci. Rep. 2016, 6, 24765. [Google Scholar] [CrossRef]
- Filler Hayut, S.; Melamed Bessudo, C.; Levy, A.A. Targeted recombination between homologous chromosomes for precise breeding in tomato. Nat. Commun. 2017, 8, 15605. [Google Scholar] [CrossRef]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 232. [Google Scholar] [CrossRef]
- Baltes, N.J.; Gil-Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA replicons for plant genome engineering. Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.-H.; Wang, B.; Li, N.; Tang, Y.; Yang, S.; Yang, T.; Xu, J.; Guo, C.; Yan, P.; Wang, Q.; et al. CRISPR/Cas9-induced targeted mutagenesis and gene replacement to generate long-shelf life tomato lines. Sci. Rep. 2017, 7, 11874. [Google Scholar] [CrossRef]
- Begemann, M.B.; Gray, B.N.; January, E.; Gordon, G.C.; He, Y.; Liu, H.; Wu, X.; Brutnell, T.P.; Mockler, T.C.; Oufattole, M. Precise insertion and guided editing of higher plant genomes using Cpf1 CRISPR nucleases. Sci. Rep. 2017, 7, 11606. [Google Scholar] [CrossRef]
- Li, G.; Zhang, X.; Zhong, C.; Mo, J.; Quan, R.; Yang, J.; Liu, D.; Li, Z.; Yang, H.; Wu, Z. Small molecules enhance CRISPR/Cas9-mediated homology-directed genome editing in primary cells. Sci. Rep. 2017, 7, 8943. [Google Scholar] [CrossRef] [PubMed]
- De Pater, S.; Klemann, B.J.; Hooykaas, P.J. True gene-targeting events by CRISPR/Cas-induced DSB repair of the PPO locus with an ectopically integrated repair template. Sci. Rep. 2018, 8, 3338. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachâal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 72. [Google Scholar] [CrossRef]
- Park, J.; Bae, S.; Kim, J.S. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef]
- Park, J.; Lim, K.; Kim, J.S.; Bae, S. Cas-analyzer: An online tool for assessing genome editing results using NGS data. Bioinformatics 2017, 33, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Kim, H.Y.; Lee, C.H.; Kim, Y.U.; Back, K.W.; Bae, J.M.; Ahn, M.J. Variation in carotenoid composition in carrots during storage and cooking. PNF 2009, 14, 240–245. [Google Scholar] [CrossRef][Green Version]
- Kim, H.S.; Ji, C.Y.; Lee, C.J.; Kim, S.E.; Park, S.C.; Kwak, S.S. Orange: A target gene for regulating carotenoid homeostasis and increasing plant tolerance to environmental stress in marginal lands. J. Exp. Bot. 2018, 69, 3393–3400. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Region | No. of Examined Calli | DNA Repair Pathway | No. of Edited Calli (%) | Mutant Type | No. of Mutation |
|---|---|---|---|---|---|
| LcyE-sg1 | 83 | NHEJ | 48 (57.8%) | Homo | 8 |
| Bi-allelic | 13 | ||||
| Hetero | 27 | ||||
| HDR | 0 (0%) | - | - | ||
| LcyE-sg2 | 304 | NHEJ | 169 (54.5%) | Homo | 24 |
| Bi-allelic | 46 | ||||
| Hetero | 93 | ||||
| HDR | 4 (1.32%) | Homo | 4 |
| Lines | Lycopene | Lutein | 13Z-β-Carotene | α-Carotene | β-Carotene | 9Z-β-Carotene | Others | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| WT | 1.71 | 0.22 | 2.31 | 0.71 | 0.29 | 0.04 | 0.67 | 0.13 | 0.38 | 0.18 | 0.42 | 0.08 | 9.33 | 1.43 | 15.11 | 2.79 |
| LcyEsg2-HDR1 | 205.1 | 36.0 | 2.87 | 0.10 | 0.23 | 0.01 | 1.08 | 0.41 | 0.09 | 0.01 | 0.44 | 0.02 | 21.11 | 1.21 | 230.92 | 37.76 |
| LcyEsg2-HDR2 | 217.2 | 41.2 | 2.93 | 0.81 | 0.31 | 0.01 | 1.34 | 0.81 | 0.14 | 0.01 | 0.39 | 0.01 | 24.01 | 1.80 | 246.32 | 44.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Yu, J.; Kim, H.K.; Kim, J.Y.; Kim, M.-S.; Cho, Y.-G.; Bae, S.; Kang, K.K.; Jung, Y.J. Genome Editing of Golden SNP-Carrying Lycopene Epsilon-Cyclase (LcyE) Gene Using the CRSPR-Cas9/HDR and Geminiviral Replicon System in Rice. Int. J. Mol. Sci. 2022, 23, 10383. https://doi.org/10.3390/ijms231810383
Kim JH, Yu J, Kim HK, Kim JY, Kim M-S, Cho Y-G, Bae S, Kang KK, Jung YJ. Genome Editing of Golden SNP-Carrying Lycopene Epsilon-Cyclase (LcyE) Gene Using the CRSPR-Cas9/HDR and Geminiviral Replicon System in Rice. International Journal of Molecular Sciences. 2022; 23(18):10383. https://doi.org/10.3390/ijms231810383
Chicago/Turabian StyleKim, Jong Hee, Jihyeon Yu, Hee Kyoung Kim, Jin Young Kim, Me-Sun Kim, Yong-Gu Cho, Sangsu Bae, Kwon Kyoo Kang, and Yu Jin Jung. 2022. "Genome Editing of Golden SNP-Carrying Lycopene Epsilon-Cyclase (LcyE) Gene Using the CRSPR-Cas9/HDR and Geminiviral Replicon System in Rice" International Journal of Molecular Sciences 23, no. 18: 10383. https://doi.org/10.3390/ijms231810383
APA StyleKim, J. H., Yu, J., Kim, H. K., Kim, J. Y., Kim, M.-S., Cho, Y.-G., Bae, S., Kang, K. K., & Jung, Y. J. (2022). Genome Editing of Golden SNP-Carrying Lycopene Epsilon-Cyclase (LcyE) Gene Using the CRSPR-Cas9/HDR and Geminiviral Replicon System in Rice. International Journal of Molecular Sciences, 23(18), 10383. https://doi.org/10.3390/ijms231810383