
HTRA1 Regulates Subclinical Inflammation and Activates Proangiogenic Response in the Retina and Choroid


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
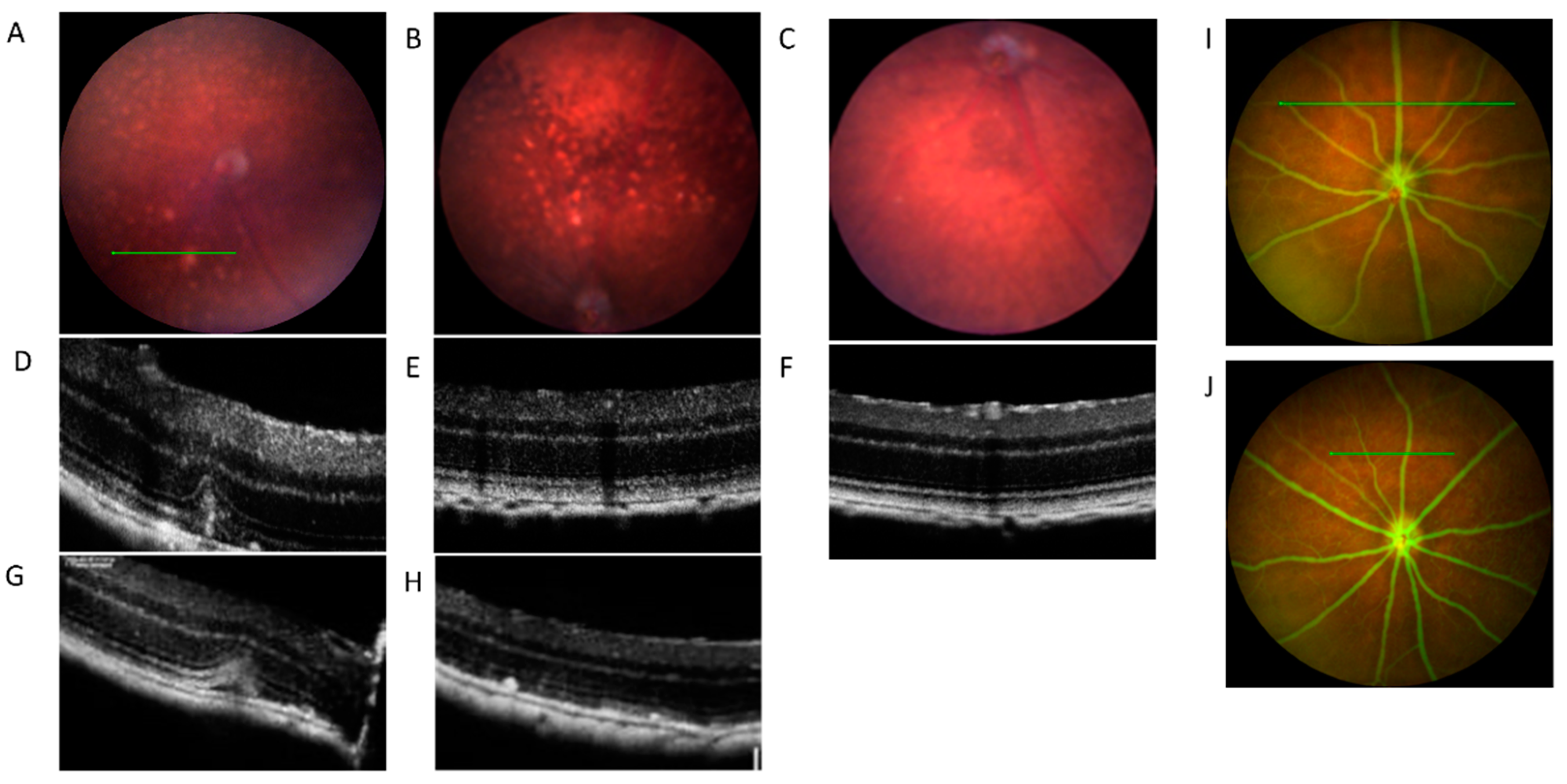
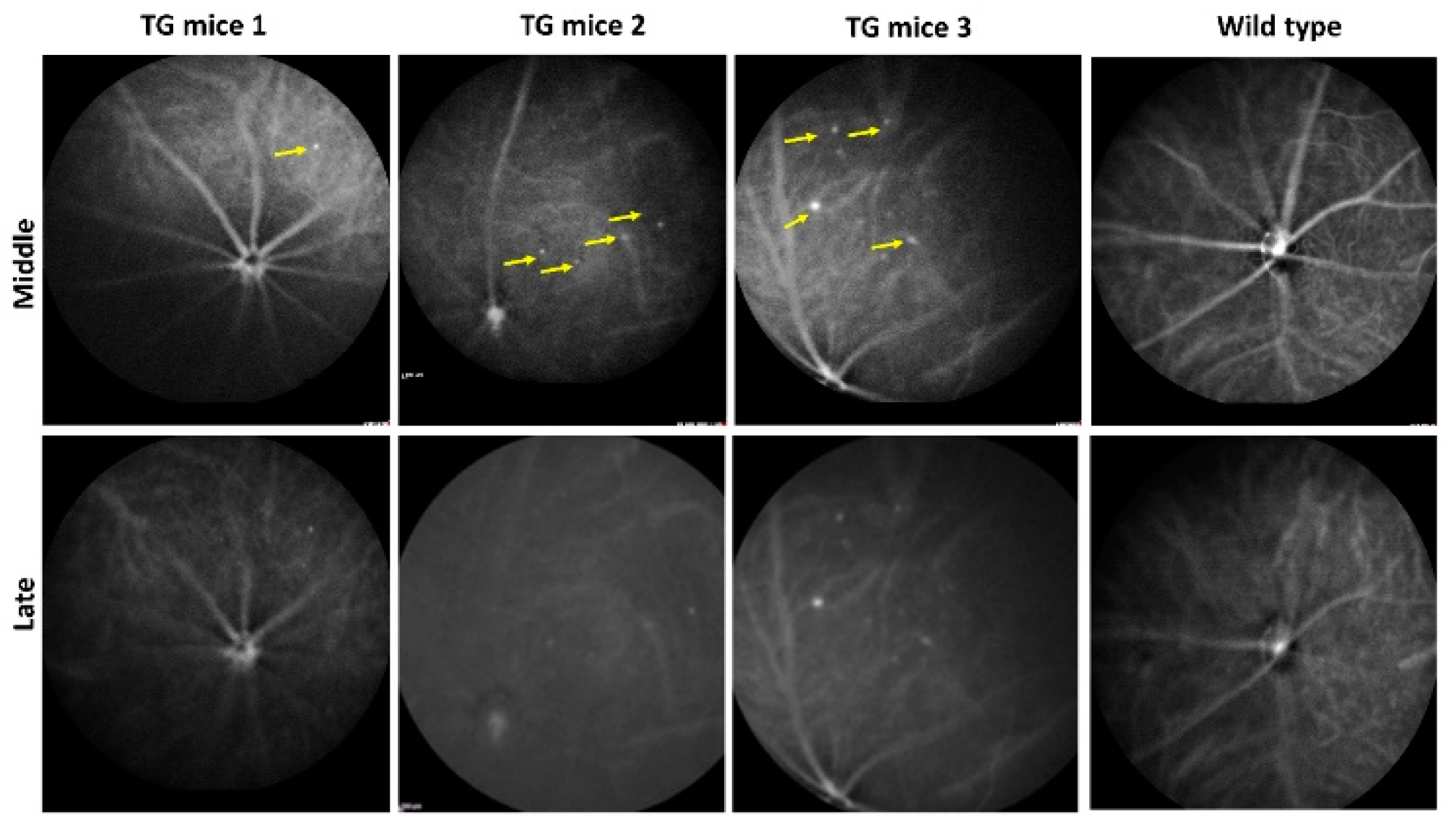
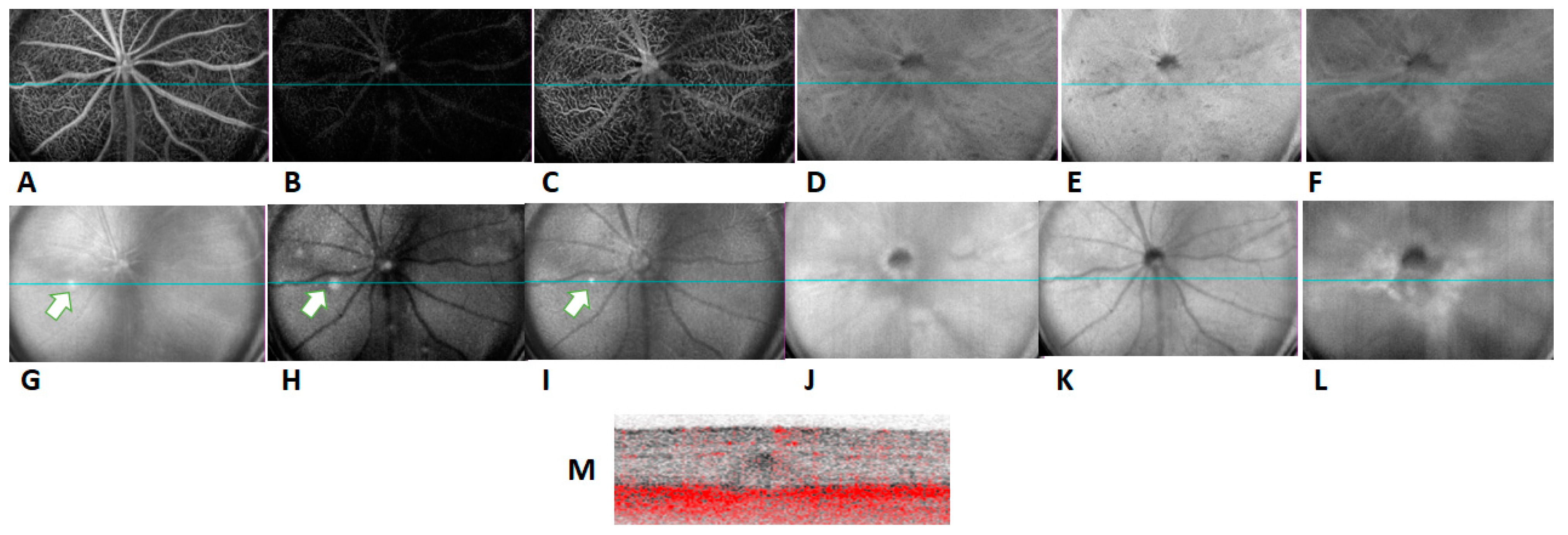
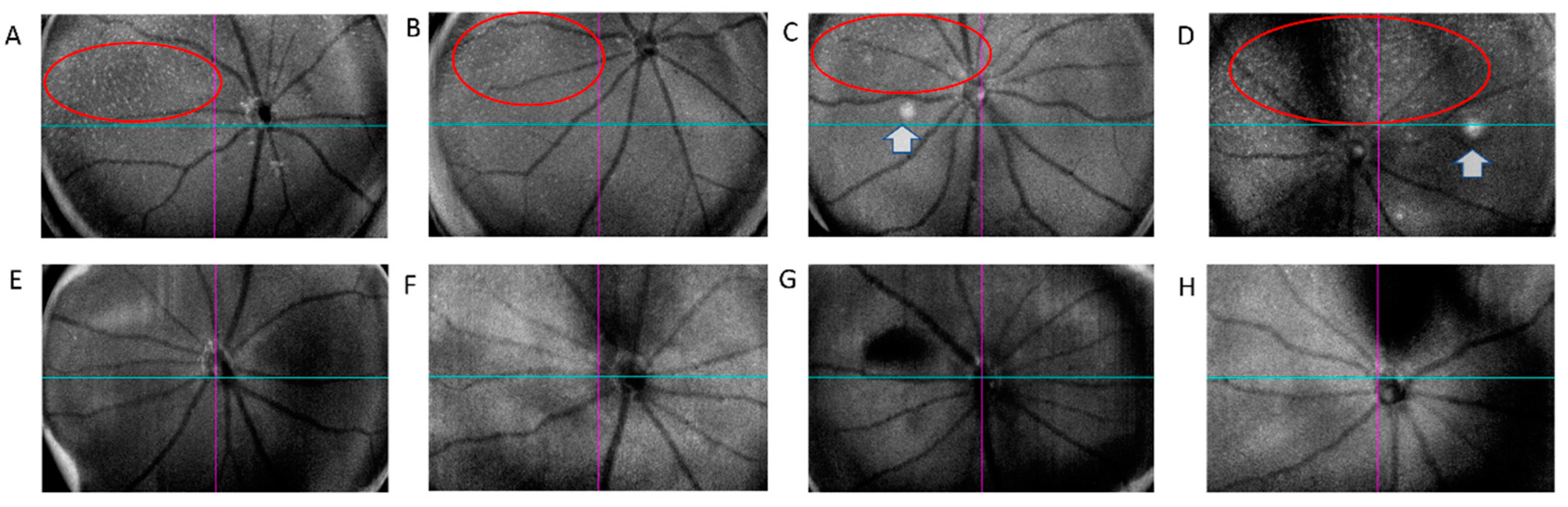
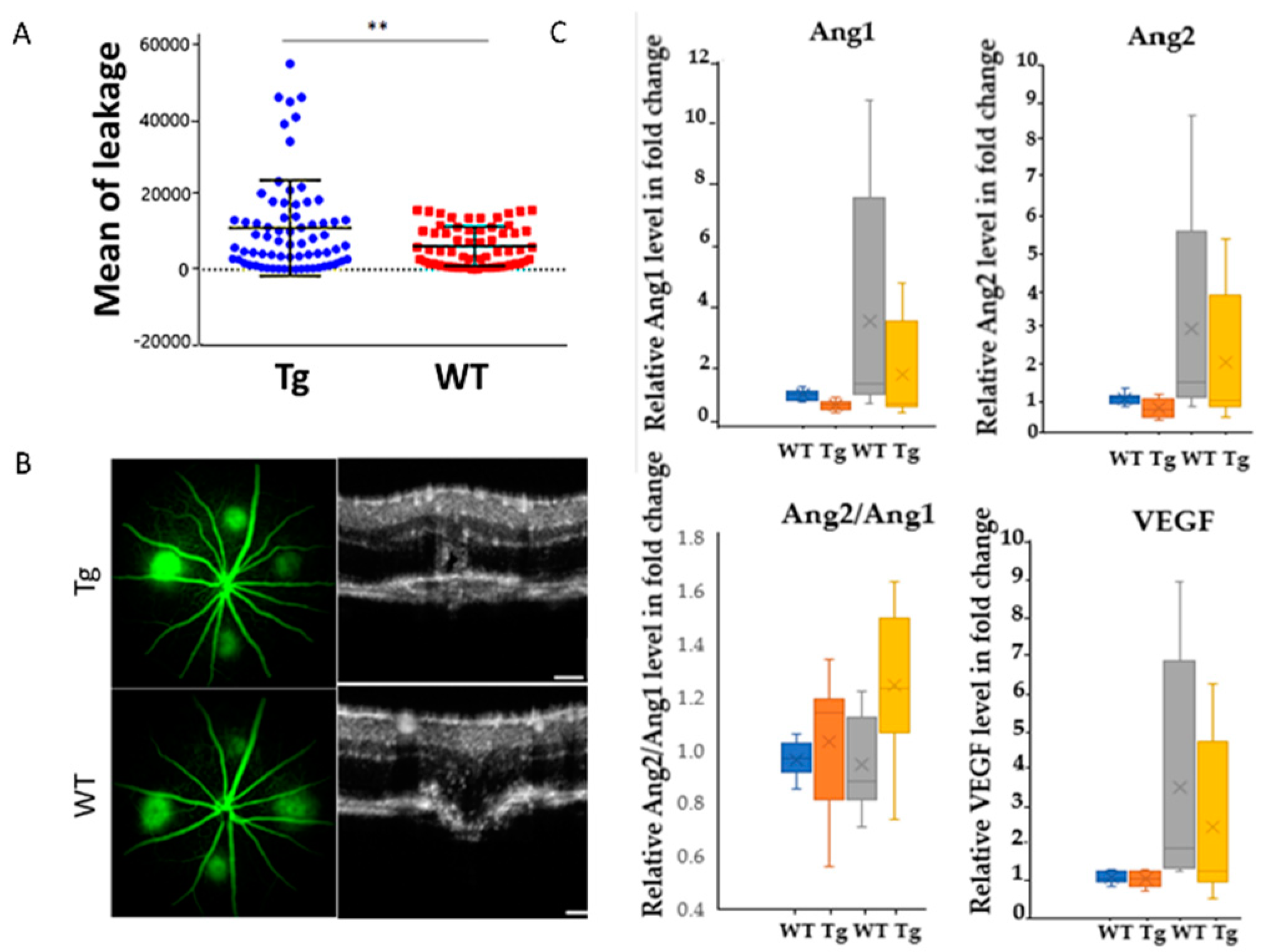
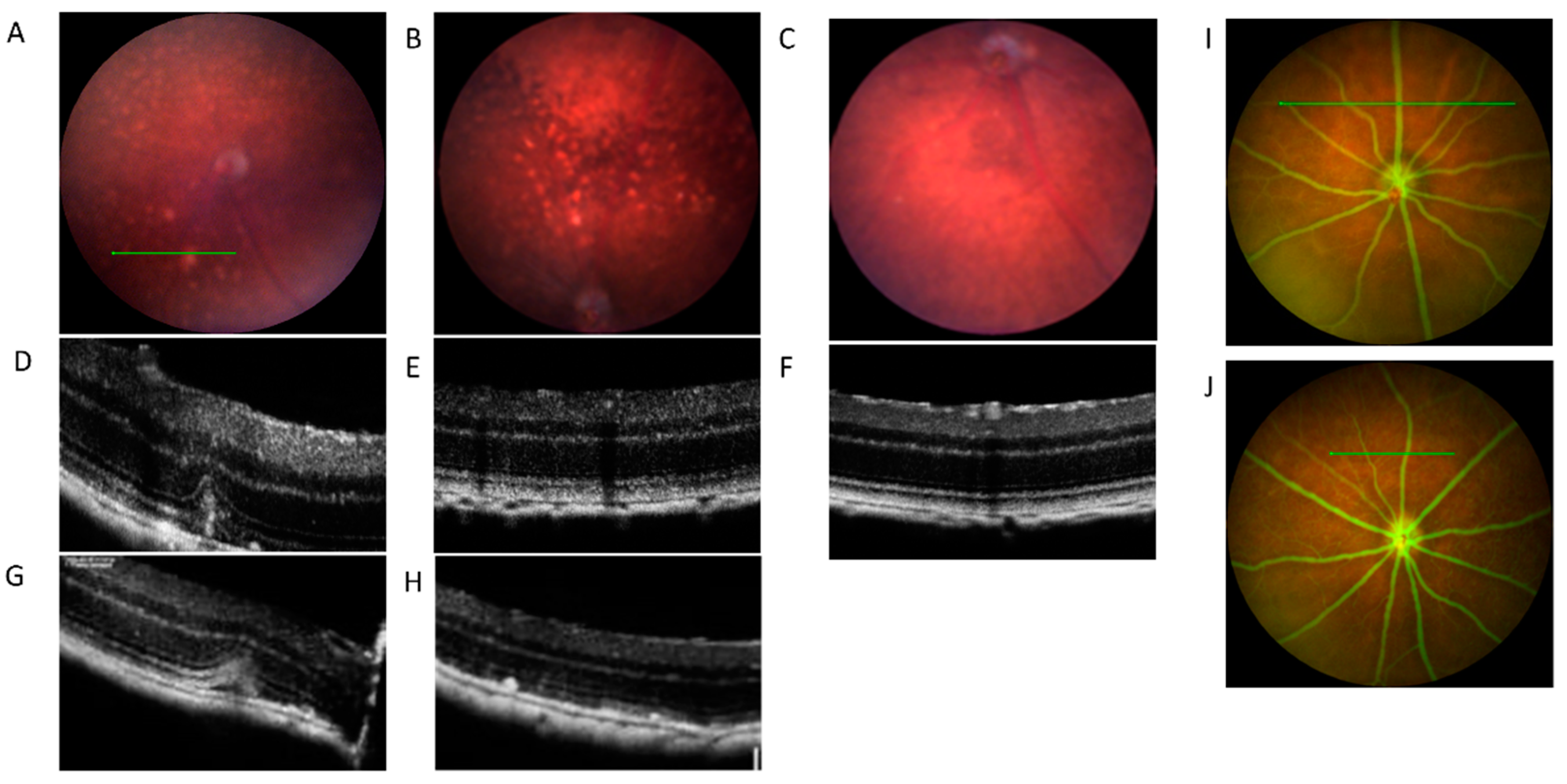
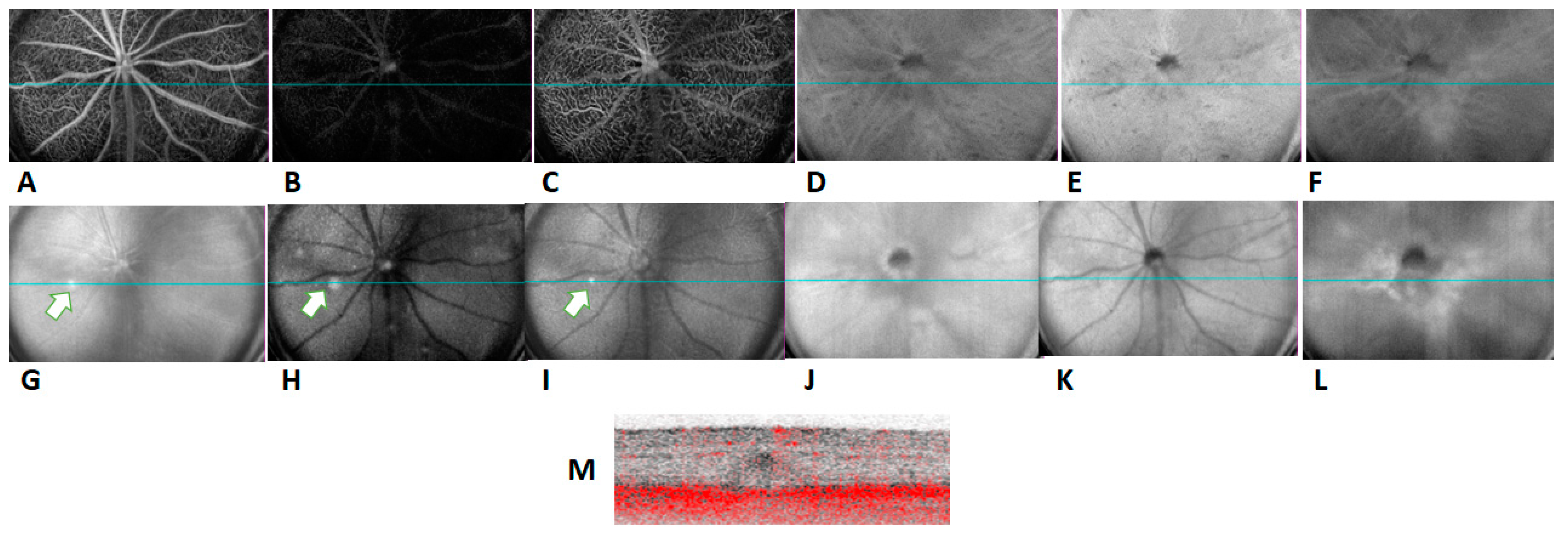
2.1. In Vivo Retinal Imaging of CAG-HtrA1 Tg Mice
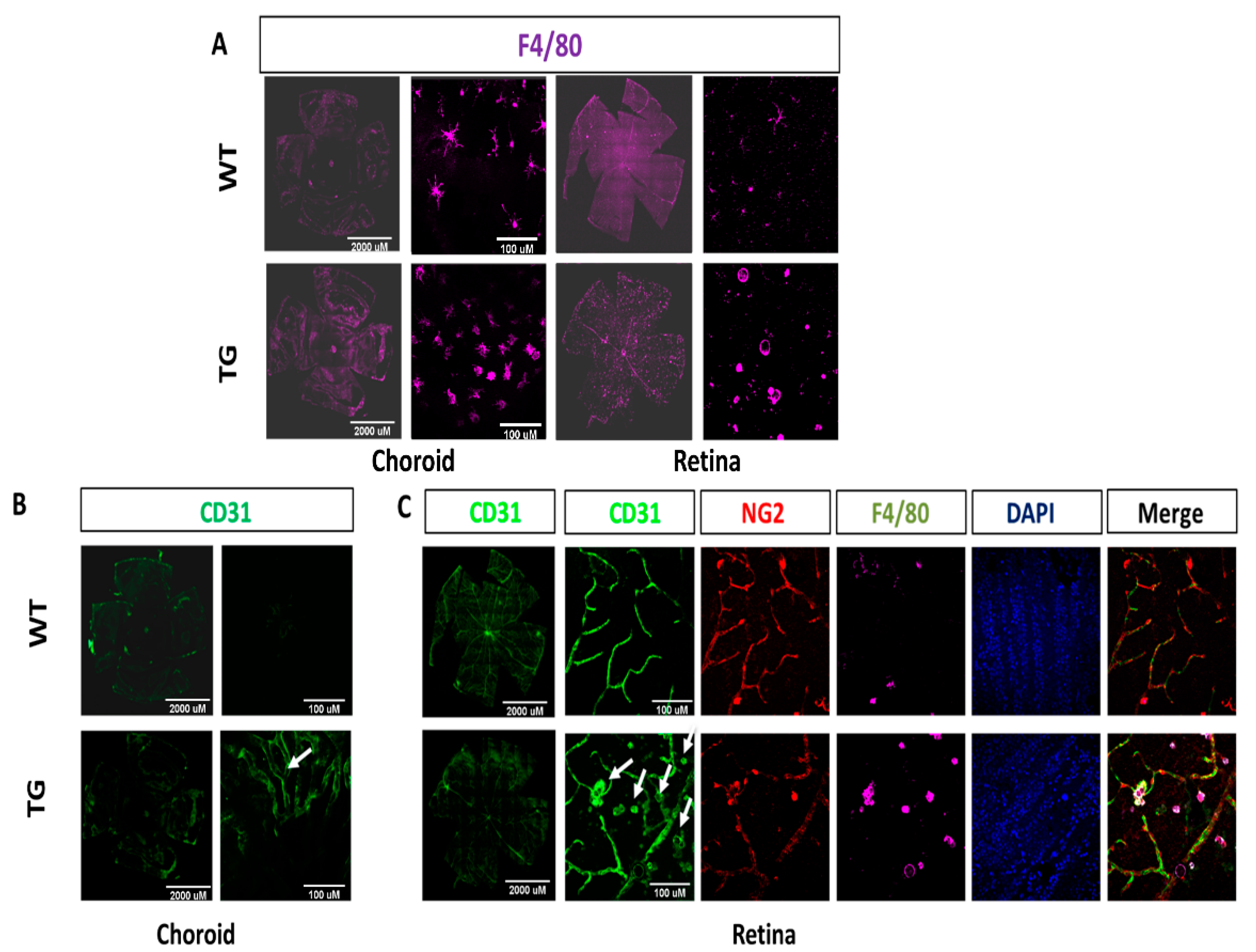
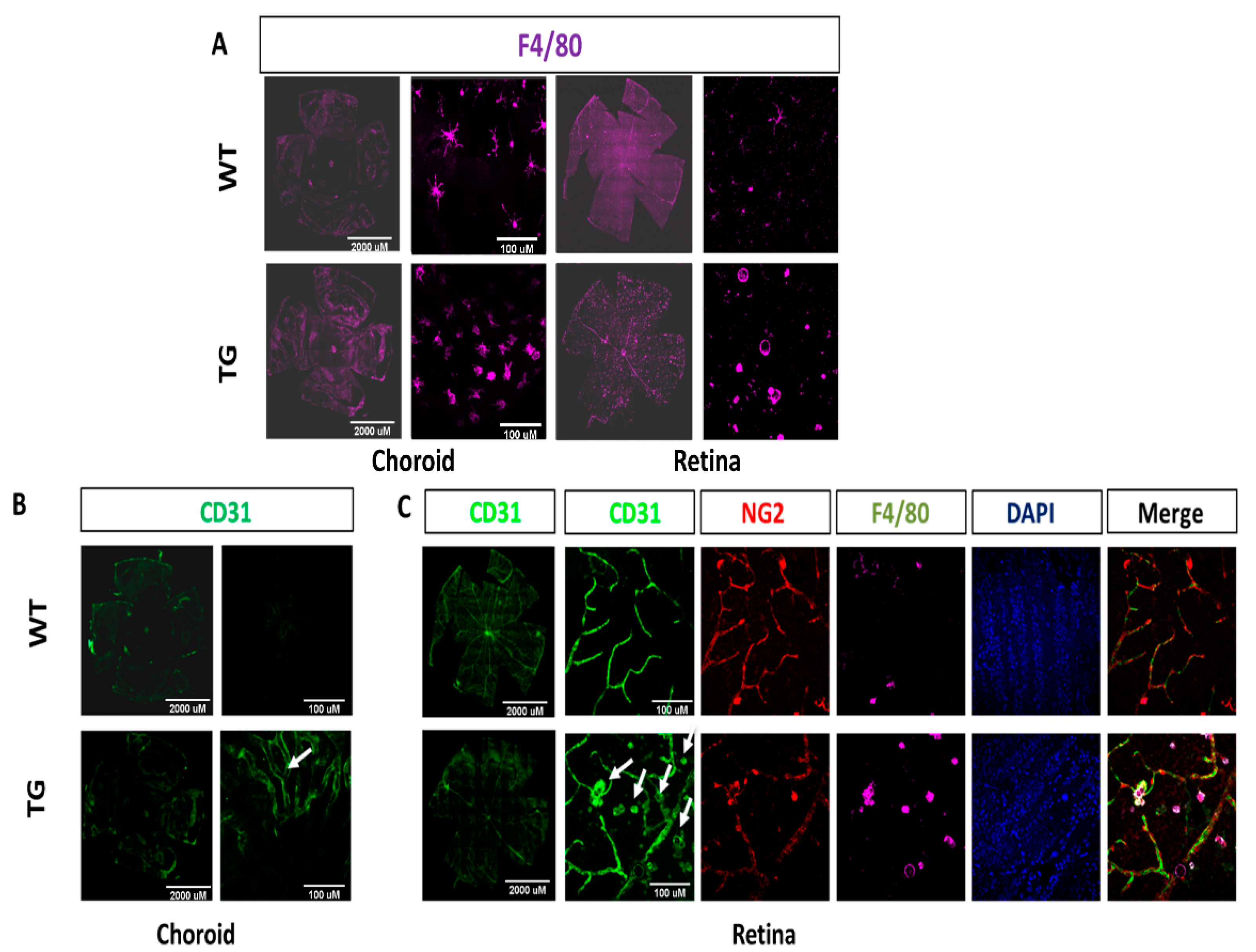
2.2. Increased Macrophage Infiltration and Activation in CAG-HtrA1 Tg Mice Retina
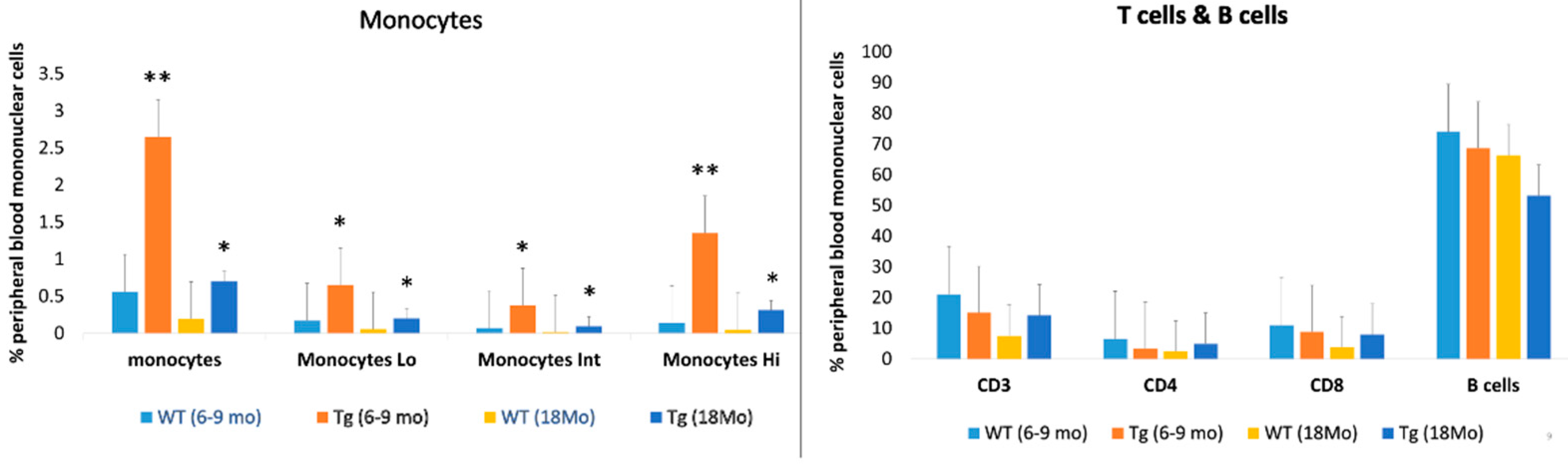
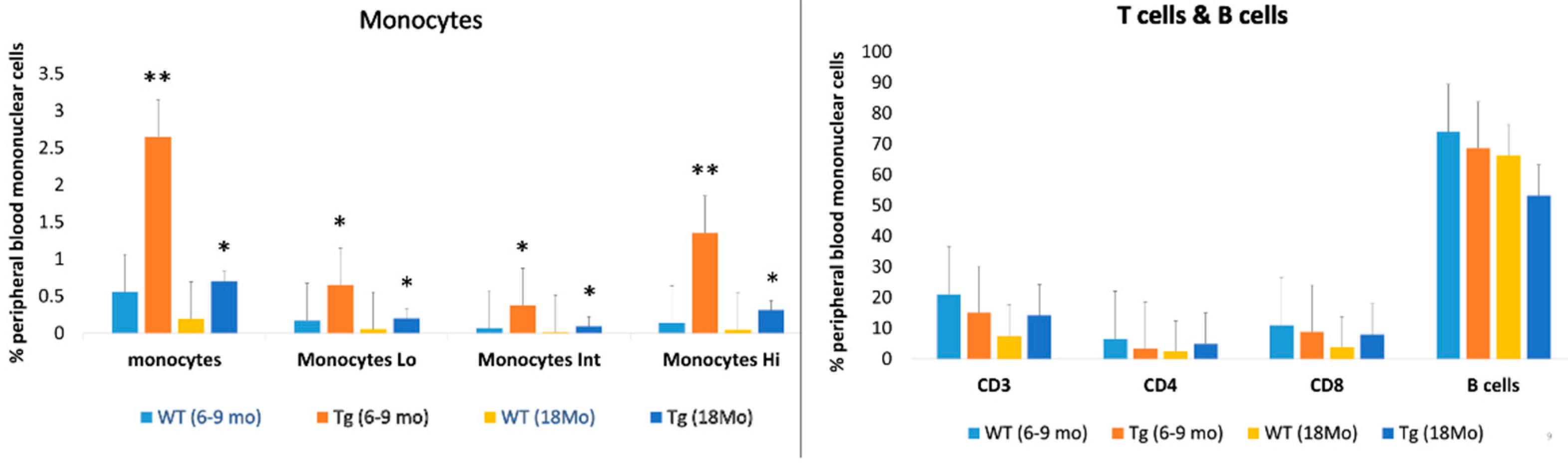
2.3. Increased Peripheral Blood Monocyte Subsets in CAG-HtrA1 Tg Mice
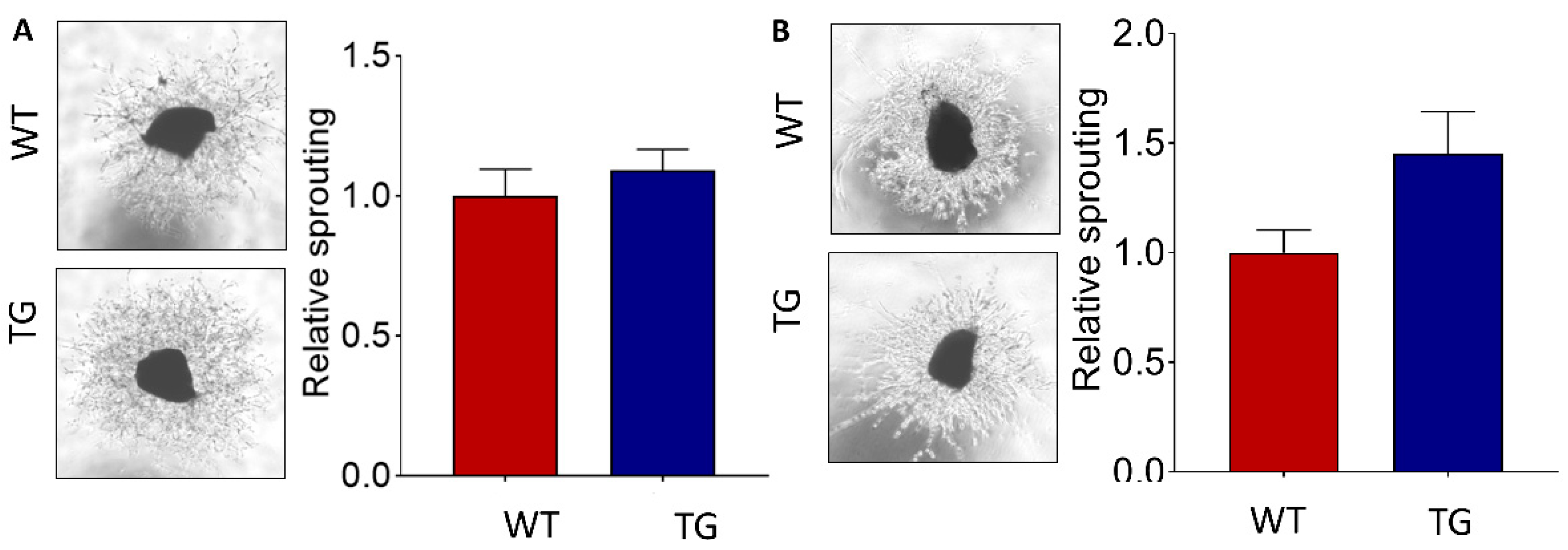
2.4. Increased Choroidal Angiogenic Potential of CAG-HtrA1 Tg Mice
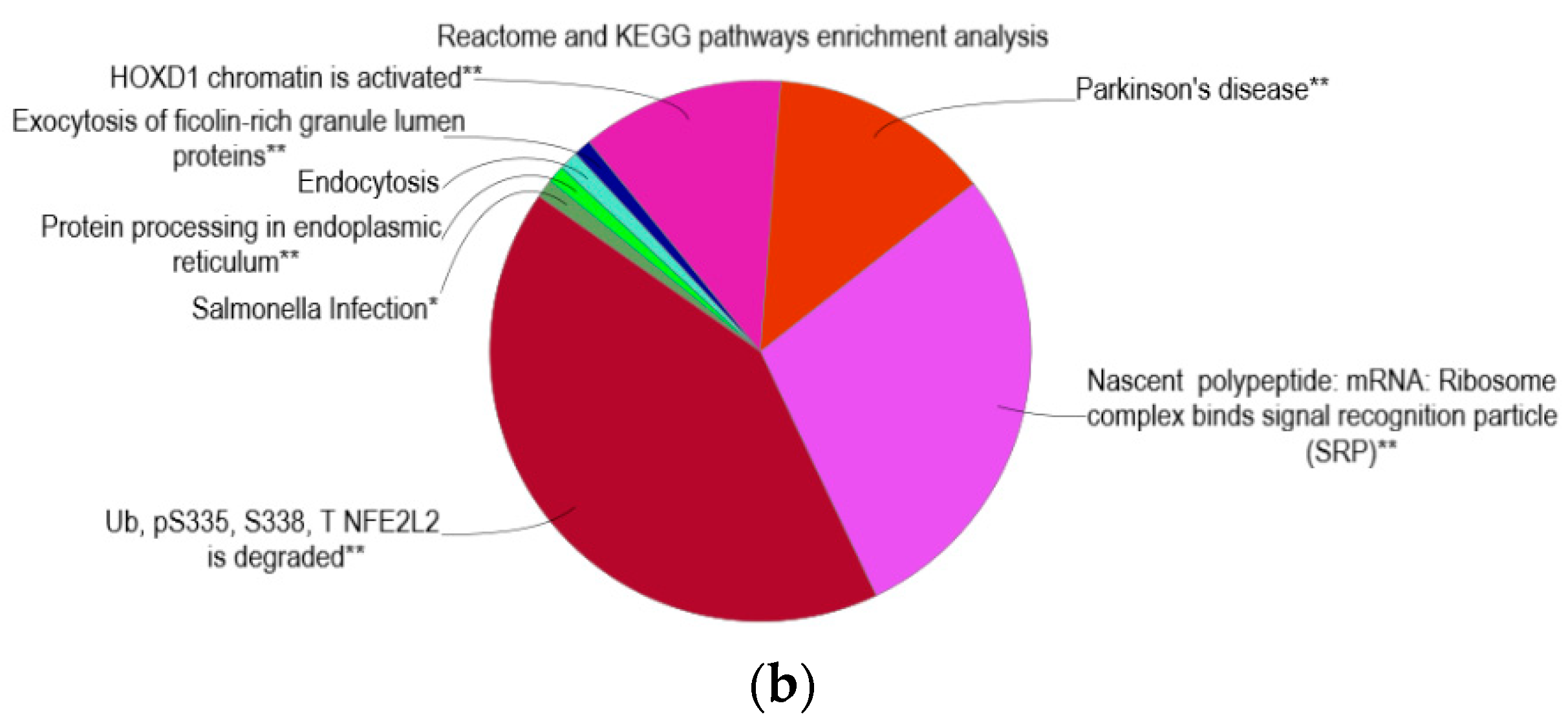
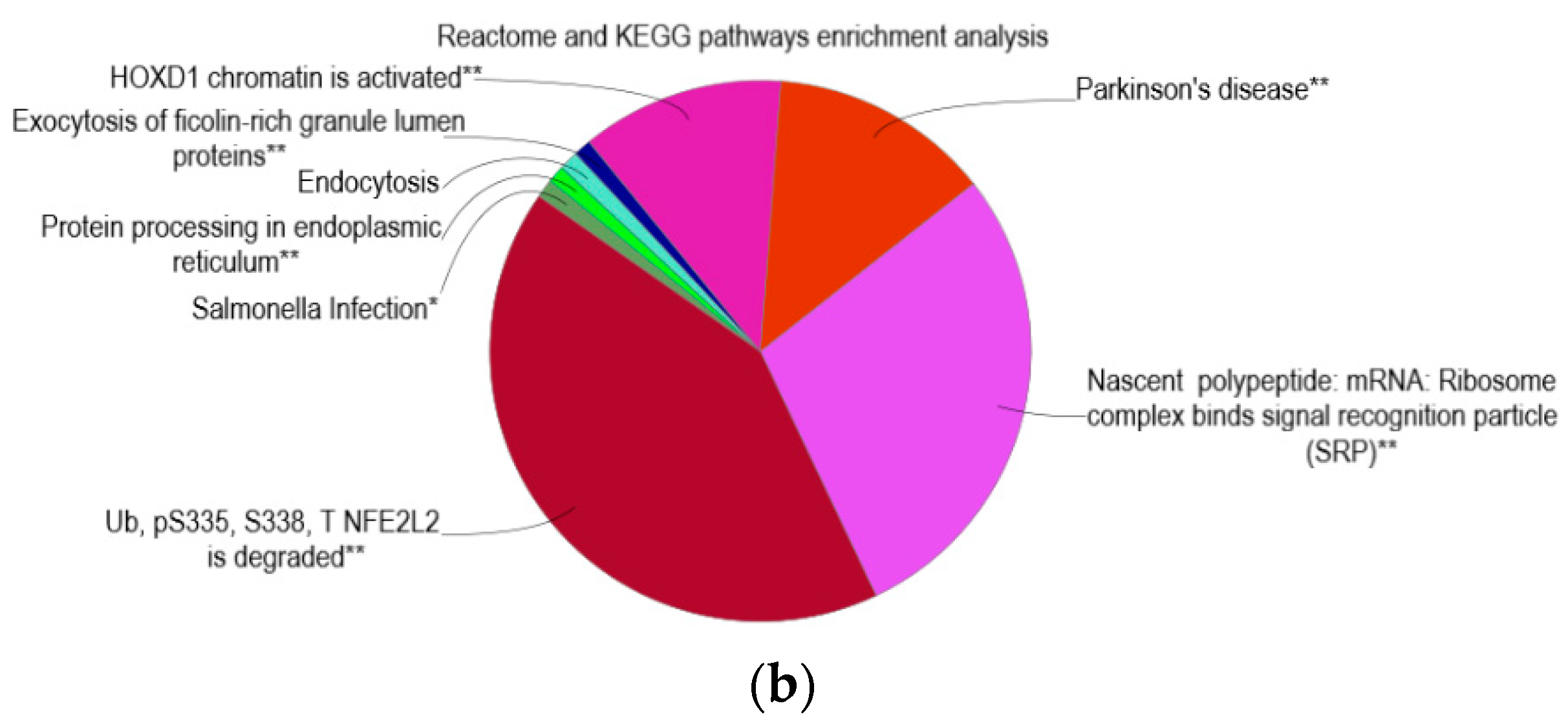
2.5. Reactome and KEGG Pathway Enrichment Analysis
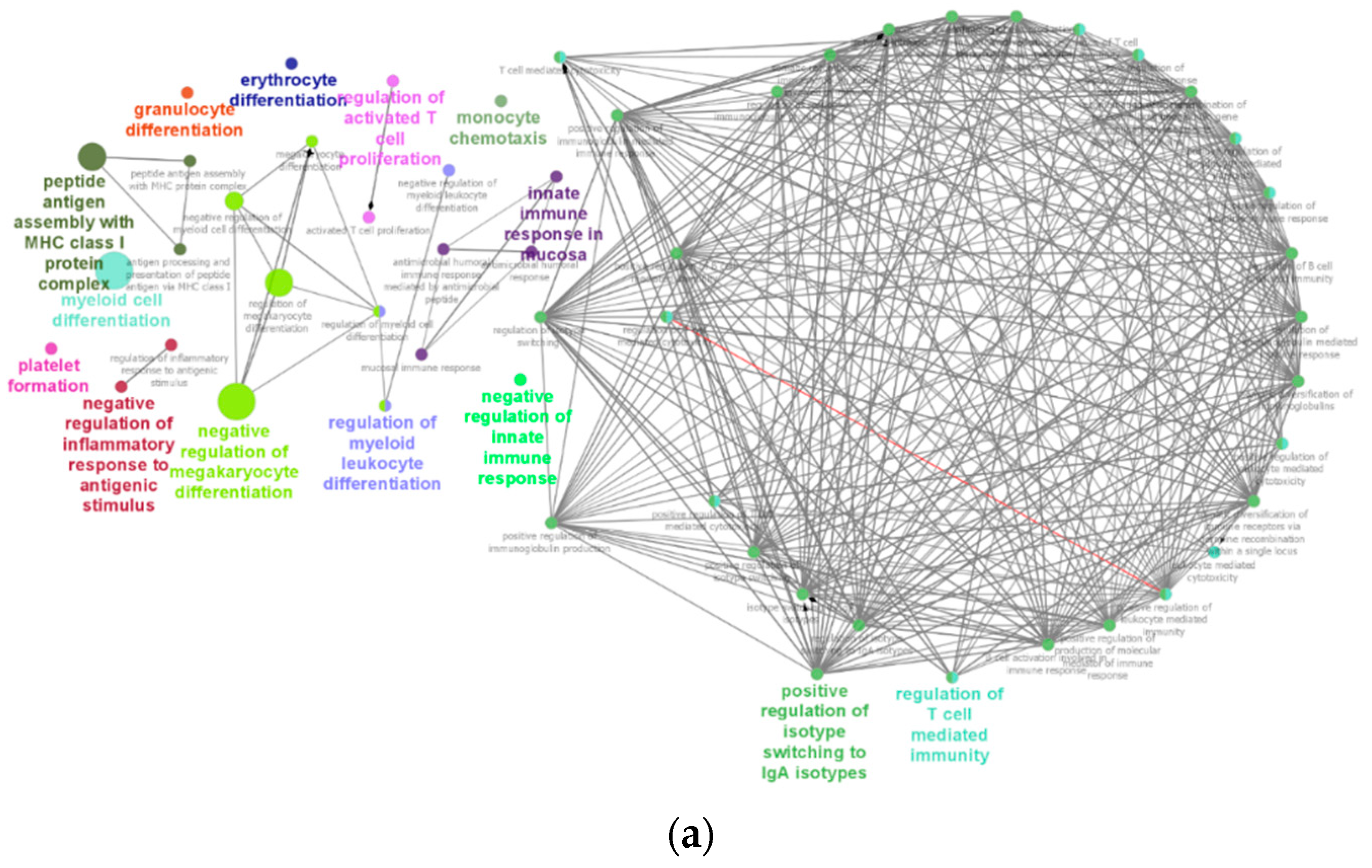
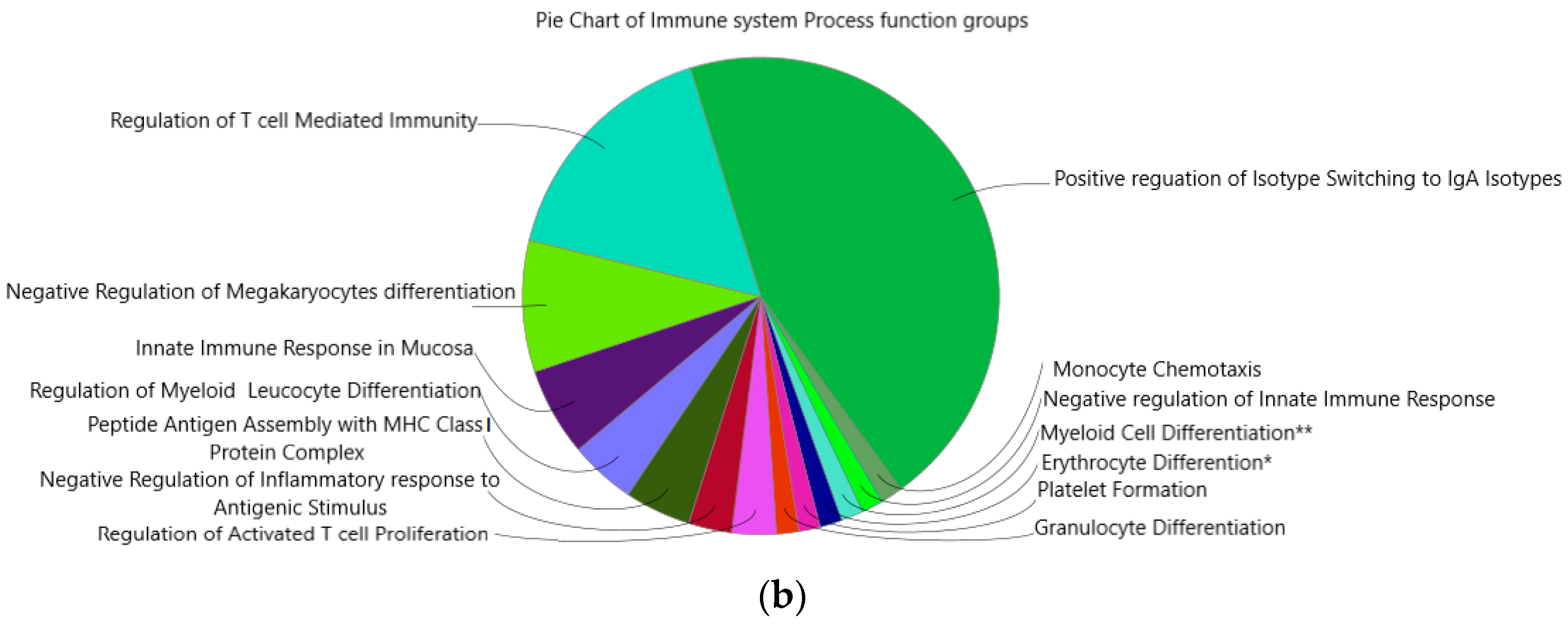
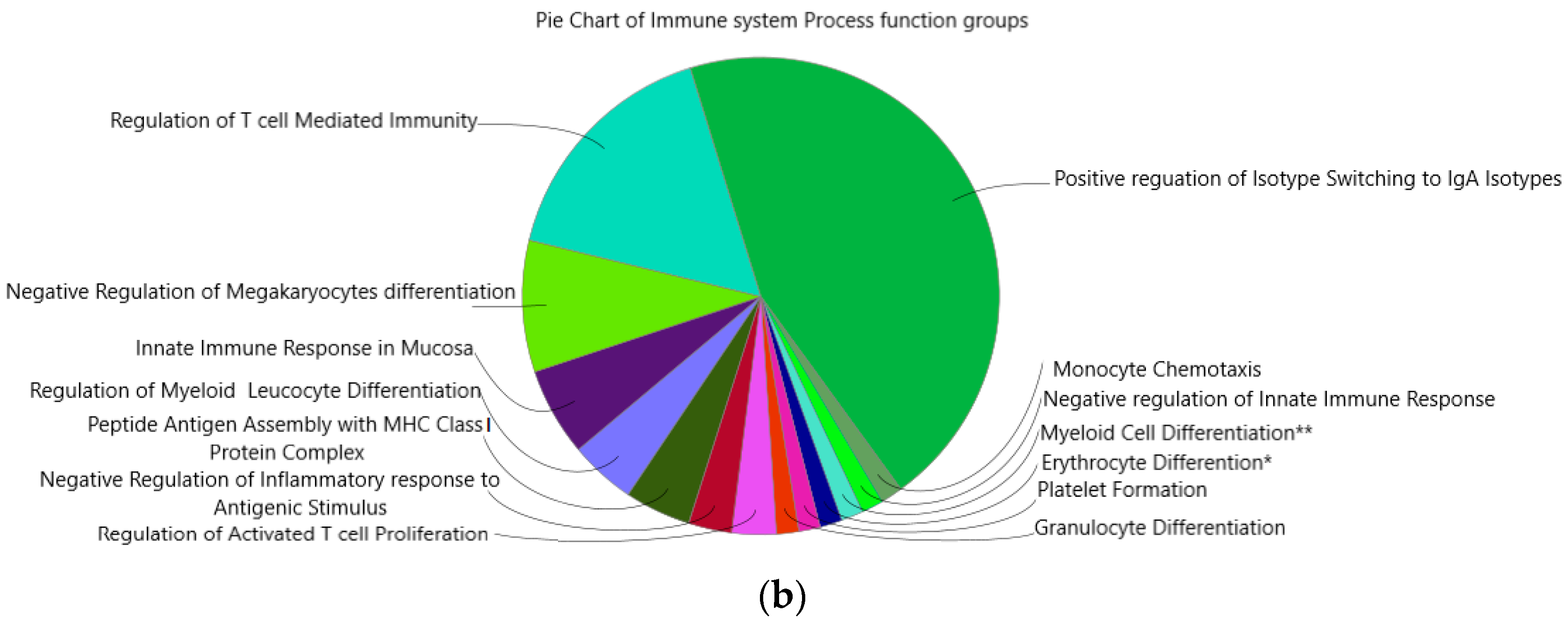
2.6. Immune System Network and GO-Terms Pathway Enrichment Analysis
3. Discussion
3.1. Key Findings in the Current Study
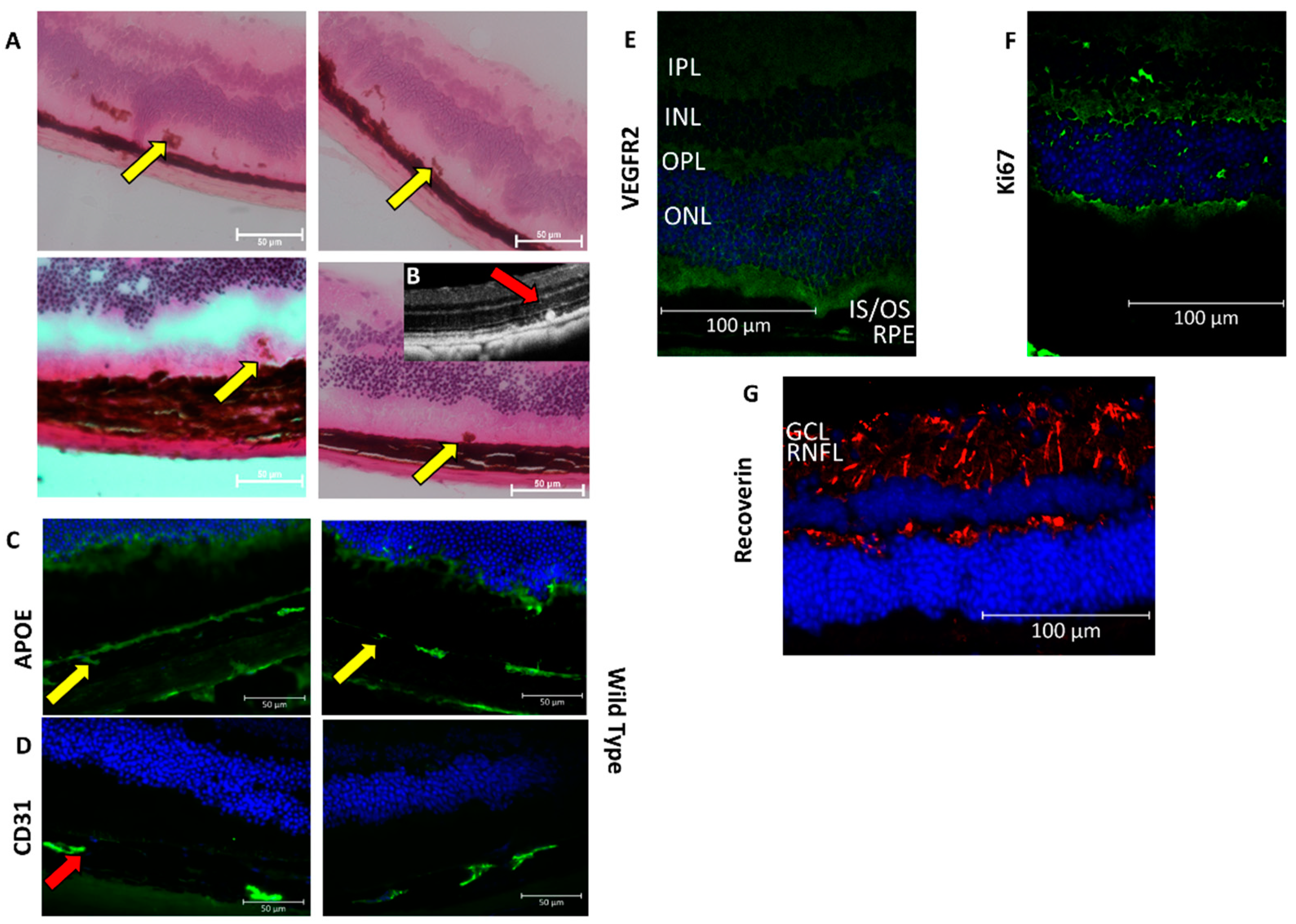
3.2. Type 1 Lesions in the CAG-HtrA1 Tg Mice Were Different from Those in Human PCV
3.3. Activated Macrophages in the Subretinal Lesions and Increased Subclinical Inflammation
3.4. HtrA1-Induced Subclinical Inflammation and Pro-Angiogenic Activities
3.5. Enrichment of Homeobox D1 and Parkinson’s Disease Pathways
3.6. Enrichment of IgA Isotype and Myeloid Cells Differentiation Pathways
3.7. Limitations
4. Materials and Methods
4.1. Animals
4.2. CFP, OCT and FFA
4.3. OCTA and ICGA Imaging
4.4. Histology and Immunohistochemistry
4.5. Flow Cytometric Analysis of Monocytes
4.6. Laser-Induced CNV
4.7. Choroidal Sprouting Assay
4.8. RNA Extraction, Sequencing, and Data Analyses
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef]
- Yannuzzi, L.A.; Ciardella, A.; Spaide, R.F.; Rabb, M.; Freund, K.B.; Orlock, D.A. The Expanding Clinical Spectrum of Idiopathic Polypoidal Choroidal Vasculopathy. Arch. Ophthalmol. 1997, 115, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Engelbert, M.; Iida, T.; Freund, K.B.; Yannuzzi, L.A. Polypoidal Choroidal Vasculopathy: A Review. Surv. Ophthalmol. 2010, 55, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.H.; Schubert, C.; Ferrara, D.C.; Merriam, J.E.; Imamura, Y.; Freund, K.B.; Spaide, R.F.; Yannuzzi, L.A.; Allikmets, R. Three Major Loci Involved in Age-Related Macular Degeneration Are Also Associated with Polypoidal Choroidal Vasculopathy. Ophthalmology 2010, 117, 1567–1570. [Google Scholar] [CrossRef]
- Lee, K.Y.; Vithana, E.N.; Mathur, R.; Yong, V.H.; Yeo, I.Y.; Thalamuthu, A.; Lee, M.-W.; Koh, A.H.; Lim, M.C.; How, A.C.; et al. Association Analysis of CFH, C2, BF, and HTRA1 Gene Polymorphisms in Chinese Patients with Polypoidal Choroidal Vasculopathy. Investig. Opthalmol. Vis. Sci. 2008, 49, 2613–2619. [Google Scholar] [CrossRef] [PubMed]
- DeWan, A.; Liu, M.; Hartman, S.; Zhang, S.S.-M.; Liu, D.T.L.; Zhao, C.; Tam, P.O.S.; Chan, W.M.; Lam, D.S.C.; Snyder, M.; et al. HTRA1 Promoter Polymorphism in Wet Age-Related Macular Degeneration. Science 2006, 314, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Kumar, S.; Zhang, N.; Tong, Z.; Yang, J.-H.; Watt, C.; Anderson, J.; Amrita; Fillerup, H.; McCloskey, M.; et al. Increased expression of multifunctional serine protease, HTRA1, in retinal pigment epithelium induces polypoidal choroidal vasculopathy in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 14578–14583. [Google Scholar] [CrossRef]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef]
- Pan, Y.; Iejima, D.; Nakayama, M.; Suga, A.; Noda, T.; Kaur, I.; Das, T.; Chakrabarti, S.; Guymer, R.H.; DeAngelis, M.M.; et al. Binding of Gtf2i-β/δ transcription factors to the ARMS2 gene leads to increased circulating HTRA1 in AMD patients and in vitro. J. Biol. Chem. 2021, 296, 100456. [Google Scholar] [CrossRef]
- Kumar, S.; Berriochoa, Z.; Ambati, B.K.; Fu, Y. Angiographic Features of Transgenic Mice With Increased Expression of Human Serine Protease HTRA1 in Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2014, 55, 3842–3850. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Nakashizuka, H.; Jones, A.; Lambert, A.; Zhao, X.; Shen, M.; Parker, M.; Wang, S.; Berriochoa, Z.; Fnu, A.; et al. Proteolytic Degradation and Inflammation Play Critical Roles in Polypoidal Choroidal Vasculopathy. Am. J. Pathol. 2017, 187, 2841–2857. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Berriochoa, Z.; Jones, A.D.; Fu, Y. Detecting Abnormalities in Choroidal Vasculature in a Mouse Model of Age-related Macular Degeneration by Time-course Indocyanine Green Angiography. J. Vis. Exp. 2014, 84, 51061. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Iejima, D.; Akahori, M.; Kamei, J.; Goto, A.; Iwata, T. Overexpression of HtrA1 and Exposure to Mainstream Cigarette Smoke Leads to Choroidal Neovascularization and Subretinal Deposits in Aged Mice. Investig. Opthalmol. Vis. Sci. 2014, 55, 6514–6523. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, Y.; Foo, V.H.X.; Yoshida, A. Asian age-related macular degeneration: From basic science research perspective. Eye 2018, 33, 34–49. [Google Scholar] [CrossRef]
- Meyer, J.H.; Larsen, P.P.; Strack, C.; Harmening, W.M.; Krohne, T.U.; Holz, F.G.; Schmitz-Valckenberg, S. Optical coherence tomography angiography (OCT-A) in an animal model of laser-induced choroidal neovascularization. Exp. Eye Res. 2019, 184, 162–171. [Google Scholar] [CrossRef]
- Agrawal, R.; Tun, S.B.B.; Balne, P.K.; Zhu, H.-Y.; Khandelwal, N.; A Barathi, V. Fluorescein Labeled Leukocytes for in vivo Imaging of Retinal Vascular Inflammation and Infiltrating Leukocytes in Laser-Induced Choroidal Neovascularization Model. Ocul. Immunol. Inflamm. 2018, 28, 7–13. [Google Scholar] [CrossRef]
- Lu, Z.; Lin, V.; May, A.; Che, B.; Xiao, X.; Shaw, D.H.; Su, F.; Wang, Z.; Du, H.; Shaw, P.X. HTRA1 synergizes with oxidized phospholipids in promoting inflammation and macrophage infiltration essential for ocular VEGF expression. PLoS ONE 2019, 14, e0216808. [Google Scholar] [CrossRef]
- Yannuzzi, L.A. Indocyanine Green Angiography: A Perspective on Use in the Clinical Setting. Am. J. Ophthalmol. 2011, 151, 745–751.e1. [Google Scholar] [CrossRef]
- Kim, S.-J.; Kim, J.-S.; Papadopoulos, J.; Kim, S.W.; Maya, M.; Zhang, F.; He, J.; Fan, D.; Langley, R.; Fidler, I.J. Circulating Monocytes Expressing CD31: Implications for Acute and Chronic Angiogenesis. Am. J. Pathol. 2009, 174, 1972–1980. [Google Scholar] [CrossRef]
- Shao, Z.; Friedlander, M.; Hurst, C.G.; Cui, Z.; Pei, D.T.; Evans, L.P.; Juan, A.; Tahiri, H.; Duhamel, F.; Chen, J.; et al. Choroid Sprouting Assay: An Ex Vivo Model of Microvascular Angiogenesis. PLoS ONE 2013, 8, e69552. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.T.; Vajzovic, L. Analyzing Optical Coherence Tomography Angiography. In Handbook of Pediatric Retinal OCT and the Eye-Brain Connection; Elsevier: Amsterdam, The Netherlands, 2019; pp. 41–44. [Google Scholar] [CrossRef]
- Caplash, S.; Kodati, S.; Cheng, S.K.; Akanda, M.; Vitale, S.; Thompson, I.; Gangaputra, S.; Sen, H.N. Repeatability of Optical Coherence Tomography Angiography in Uveitic Eyes. Transl. Vis. Sci. Technol. 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Stanzel, T.P.; Devarajan, K.; Lwin, N.C.; Yam, G.H.; Schmetterer, L.; Mehta, J.S.; Ang, M. Comparison of Optical Coherence Tomography Angiography to Indocyanine Green Angiography and Slit Lamp Photography for Corneal Vascularization in an Animal Model. Sci. Rep. 2018, 8, 11493. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.M.G.; Lee, W.K.; Koizumi, H.; Dansingani, K.; Lai, T.Y.Y.; Freund, K.B. Pachychoroid disease. Eye 2019, 33, 14–33. [Google Scholar] [CrossRef]
- Matsumoto, H.; Mukai, R.; Morimoto, M.; Tokui, S.; Kishi, S.; Akiyama, H. Clinical characteristics of pachydrusen in central serous chorioretinopathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 257, 1127–1132. [Google Scholar] [CrossRef]
- Tsujikawa, A.; Ojima, Y.; Yamashiro, K.; Ooto, S.; Tamura, H.; Nakagawa, S.; Yoshimura, N. Punctate hyperfluorescent spots associated with central serous chorioretinopathy as seen on indocyanine green angiography. Retina 2010, 30, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.; Datta, S.; Wang, L.; Pegany, R.; Cano, M.; Handa, J.T. The impact of lipids, lipid oxidation, and inflammation on AMD, and the potential role of miRNAs on lipid metabolism in the RPE. Exp. Eye Res. 2018, 181, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.S.; on behalf of the EVEREST Study Group; Ngo, W.K.; Lim, L.W.; Tan, N.W.; Lim, T.H. EVEREST study report 3: Diagnostic challenges of polypoidal choroidal vasculopathy. Lessons learnt from screening failures in the EVEREST study. Graefe’s Arch. Clin. Exp. Ophthalmol. 2016, 254, 1923–1930. [Google Scholar] [CrossRef]
- Mishima, H.; Kondo, K. Extrusion of lysosomal bodies from apical mouse retinal pigment epithelium. Albr. Graefes Arch. Klin. Exp. Ophthalmol. 1981, 216, 209–217. [Google Scholar] [CrossRef]
- Xu, H.; Chen, M.; Manivannan, A.; Lois, N.; Forrester, J.V. Age-dependent accumulation of lipofuscin in perivascular and subretinal microglia in experimental mice. Aging Cell 2008, 7, 58–68. [Google Scholar] [CrossRef]
- Tamiya, S.; Liu, L.; Kaplan, H.J. Epithelial-Mesenchymal Transition and Proliferation of Retinal Pigment Epithelial Cells Initiated upon Loss of Cell-Cell Contact. Investig. Opthalmol. Vis. Sci. 2010, 51, 2755–2763. [Google Scholar] [CrossRef]
- Curcio, C.A.; Zanzottera, E.C.; Ach, T.; Balaratnasingam, C.; Freund, K.B. Activated Retinal Pigment Epithelium, an Optical Coherence Tomography Biomarker for Progression in Age-Related Macular Degeneration. Investig. Opthalmol. Vis. Sci. 2017, 58, BIO211–BIO226. [Google Scholar] [CrossRef]
- Nassisi, M.; Lei, J.; Abdelfattah, N.; Karamat, A.; Balasubramanian, S.; Fan, W.; Uji, A.; Marion, K.M.; Baker, K.; Huang, X.; et al. OCT Risk Factors for Development of Late Age-Related Macular Degeneration in the Fellow Eyes of Patients Enrolled in the HARBOR Study. Ophthalmology 2019, 126, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Curcio, C.A.; Johnson, M.; Huang, J.-D.; Rudolf, M. Aging, age-related macular degeneration, and the response-to-retention of apolipoprotein B-containing lipoproteins. Prog. Retin. Eye Res. 2009, 28, 393–422. [Google Scholar] [CrossRef] [PubMed]
- Hageman, G.S. An Integrated Hypothesis That Considers Drusen as Biomarkers of Immune-Mediated Processes at the RPE-Bruch’s Membrane Interface in Aging and Age-Related Macular Degeneration. Prog. Retin. Eye Res. 2001, 20, 705–732. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, F.; Yan, A.; Xia, X. Overexpression of HTRA1 increases the proliferation and migration of retinal pigment epithelium. Adv. Clin. Exp. Med. 2021, 30, 859–864. [Google Scholar] [CrossRef]
- Combadiere, C.; Feumi, C.; Raoul, W.; Keller, N.; Rodero, M.; Pézard, A.; Lavalette, S.; Houssier, M.; Jonet, L.; Picard, E.; et al. CX3CR1-dependent subretinal microglia cell accumulation is associated with cardinal features of age-related macular degeneration. J. Clin. Investig. 2007, 117, 2920–2928. [Google Scholar] [CrossRef]
- Luhmann, U.F.O.; Robbie, S.; Munro, P.M.G.; Barker, S.E.; Duran, Y.; Luong, V.; Fitzke, F.W.; Bainbridge, J.W.B.; Ali, R.R.; MacLaren, R.E. The Drusenlike Phenotype in Aging Ccl2-Knockout Mice Is Caused by an Accelerated Accumulation of Swollen Autofluorescent Subretinal Macrophages. Investig. Opthalmol. Vis. Sci. 2009, 50, 5934–5943. [Google Scholar] [CrossRef]
- Joussen, A.M.; Ricci, F.; Paris, L.P.; Korn, C.; Quezada-Ruiz, C.; Zarbin, M. Angiopoietin/Tie2 signalling and its role in retinal and choroidal vascular diseases: A review of preclinical data. Eye 2021, 35, 1305–1316. [Google Scholar] [CrossRef]
- Owen, L.A.; Shirer, K.; Collazo, S.A.; Szczotka, K.; Baker, S.; Wood, B.; Carroll, L.; Haaland, B.; Iwata, T.; Katikaneni, L.D.; et al. The Serine Protease HTRA-1 Is a Biomarker for ROP and Mediates Retinal Neovascularization. Front. Mol. Neurosci. 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, T.; Wang, Y.; Xie, M.; Ji, X.; Luo, X.; Huang, W.; Xia, L. Homeobox Genes in Cancers: From Carcinogenesis to Recent Therapeutic Intervention. Front. Oncol. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- McGinnis, W.; Levine, M.S.; Hafen, E.; Kuroiwa, A.; Gehring, W.J. A conserved DNA sequence in homoeotic genes of the Drosophila Antennapedia and bithorax complexes. Nature 1984, 308, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Choi, H.-J.; Kim, J.; Kim, M.; Rho, S.-S.; Hwang, D.; Kim, Y.-M.; Kwon, Y.-G. Homeobox D1 regulates angiogenic functions of endothelial cells via integrin β1 expression. Biochem. Biophys. Res. Commun. 2011, 408, 186–192. [Google Scholar] [CrossRef]
- Toshner, M.; Dunmore, B.J.; McKinney, E.F.; Southwood, M.; Caruso, P.; Upton, P.D.; Waters, J.P.; Ormiston, M.L.; Skepper, J.N.; Nash, G.; et al. Transcript analysis reveals a specific HOX signature associated with positional identity of human endothelial cells. PLoS ONE 2014, 9, e91334. [Google Scholar] [CrossRef]
- Hansen, S.L.; Myers, C.A.; Charboneau, A.; Young, D.M.; Boudreau, N. HoxD3 Accelerates Wound Healing in Diabetic Mice. Am. J. Pathol. 2003, 163, 2421–2431. [Google Scholar] [CrossRef]
- Boudreau, N.; Andrews, C.; Srebrow, A.; Ravanpay, A.; Cheresh, D.A. Induction of the Angiogenic Phenotype by Hox D3. J. Cell Biol. 1997, 139, 257–264. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, B.; Arderiu, G.; Hashimoto, T.; Young, W.L.; Boudreau, N.; Yang, G.-Y. Retroviral Delivery of Homeobox D3 Gene Induces Cerebral Angiogenesis in Mice. J. Cereb. Blood Flow Metab. 2004, 24, 1280–1287. [Google Scholar] [CrossRef]
- Barcia, C.; Bautista, V.; Sanchez-Bahillo, A.; Fernandez-Villalba, E.; Faucheux, B.; Poza, M.P.Y.; Barreiro, A.F.; Hirsch, E.; Herrero, M.-T. Changes in vascularization in substantia nigra pars compacta of monkeys rendered parkinsonian. J. Neural Transm. 2005, 112, 1237–1248. [Google Scholar] [CrossRef]
- Bradaric, B.D.; Patel, A.; Schneider, J.A.; Carvey, P.M.; Hendey, B. Evidence for angiogenesis in Parkinson’s disease, incidental Lewy body disease, and progressive supranuclear palsy. J. Neural Transm. 2011, 119, 59–71. [Google Scholar] [CrossRef]
- Faucheux, B.A.; Agid, Y.; Hirsch, E.; Bonnet, A.-M. Blood vessels change in the mesencephalon of patients with Parkinson’s disease. Lancet 1999, 353, 981–982. [Google Scholar] [CrossRef]
- Yu, H.; Yuan, L.; Yang, Y.; Ma, S.; Peng, L.; Wang, Y.; Zhang, C.; Li, T. Increased serum IgA concentration and plasmablast frequency in patients with age-related macular degeneration. Immunobiology 2016, 221, 650–656. [Google Scholar] [CrossRef]
- Hansen, I.S.; Baeten, D.L.P.; Den Dunnen, J. The inflammatory function of human IgA. Cell. Mol. Life Sci. 2019, 76, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Mestas, J.; Hughes, C.C.W. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [PubMed]
- Bruhns, P.; Jönsson, F. Mouse and human FcR effector functions. Immunol. Rev. 2015, 268, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A. The regulation of IgA class switching. Nat. Rev. Immunol. 2008, 8, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, H.; Minato, N. Myeloid cells. Int. J. Biochem. Cell Biol. 2004, 36, 1374–1379. [Google Scholar] [CrossRef]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef]
- Vogt, S.D.; Curcio, C.A.; Wang, L.; Li, C.-M.; McGwin, G., Jr.; Medeiros, N.E.; Philp, N.J.; Kimble, J.A.; Read, R.W. Retinal pigment epithelial expression of complement regulator CD46 is altered early in the course of geographic atrophy. Exp. Eye Res. 2011, 93, 413–423. [Google Scholar] [CrossRef]
- Sica, A.; Erreni, M.; Allavena, P.; Porta, C. Macrophage polarization in pathology. Exp. 2015, 72, 4111–4126. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Tan, B.; Barathi, V.A.; Lin, E.; Ho, C.; Gan, A.; Yao, X.; Chan, A.; Wong, D.W.; Chua, J.; Tan, G.S.; et al. Longitudinal Structural and Microvascular Observation in RCS Rat Eyes Using Optical Coherence Tomography Angiography. Investig. Opthalmol. Vis. Sci. 2020, 61, 54. [Google Scholar] [CrossRef]
- Tan, X.; Fujiu, K.; Manabe, I.; Nishida, J.; Yamagishi, R.; Nagai, R.; Yanagi, Y. Choroidal neovascularization is inhibited via an intraocular decrease of inflammatory cells in mice lacking complement component C3. Sci. Rep. 2015, 5, 15702. [Google Scholar] [CrossRef] [PubMed]
- Fujiu, K.; Manabe, I.; Nagai, R. Renal collecting duct epithelial cells regulate inflammation in tubulointerstitial damage in mice. J. Clin. Investig. 2011, 121, 3425–3441. [Google Scholar] [CrossRef] [PubMed]
- Tobe, T.; Ortega, S.; Luna, J.D.; Ozaki, H.; Okamoto, N.; Derevjanik, N.L.; Vinores, S.A.; Basilico, C.; Campochiaro, P.A. Targeted Disruption of the FGF2 Gene Does Not Prevent Choroidal Neovascularization in a Murine Model. Am. J. Pathol. 1998, 153, 1641–1646. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Changes | HtrA1 Tg | WT |
|---|---|---|
| Lesion specific | ||
| Type 1—Large white deposits |
|
|
| Type 2- multiple small white-yellowish foci |
|
|
| Global | ||
| ICGA |
|
|
| OCTA |
|
|
| IHC |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahamed, W.; Yu, R.M.C.; Pan, Y.; Iwata, T.; Barathi, V.A.; Wey, Y.S.; Tun, S.B.B.; Qiu, B.; Tan, A.; Wang, X.; et al. HTRA1 Regulates Subclinical Inflammation and Activates Proangiogenic Response in the Retina and Choroid. Int. J. Mol. Sci. 2022, 23, 10206. https://doi.org/10.3390/ijms231810206
Ahamed W, Yu RMC, Pan Y, Iwata T, Barathi VA, Wey YS, Tun SBB, Qiu B, Tan A, Wang X, et al. HTRA1 Regulates Subclinical Inflammation and Activates Proangiogenic Response in the Retina and Choroid. International Journal of Molecular Sciences. 2022; 23(18):10206. https://doi.org/10.3390/ijms231810206
Chicago/Turabian StyleAhamed, Waseem, Richard Ming Chuan Yu, Yang Pan, Takeshi Iwata, Veluchamy Amutha Barathi, Yeo Sia Wey, Sai Bo Bo Tun, Beiying Qiu, Alison Tan, Xiaomeng Wang, and et al. 2022. "HTRA1 Regulates Subclinical Inflammation and Activates Proangiogenic Response in the Retina and Choroid" International Journal of Molecular Sciences 23, no. 18: 10206. https://doi.org/10.3390/ijms231810206
APA StyleAhamed, W., Yu, R. M. C., Pan, Y., Iwata, T., Barathi, V. A., Wey, Y. S., Tun, S. B. B., Qiu, B., Tan, A., Wang, X., Cheung, C. M. G., Wong, T. Y., & Yanagi, Y. (2022). HTRA1 Regulates Subclinical Inflammation and Activates Proangiogenic Response in the Retina and Choroid. International Journal of Molecular Sciences, 23(18), 10206. https://doi.org/10.3390/ijms231810206