
Blood RNA Sequencing Indicates Upregulated BATF2 and LY6E and Downregulated ISG15 and MT2A Expression in Children with Autism Spectrum Disorder
 , , ,
, , ,  ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
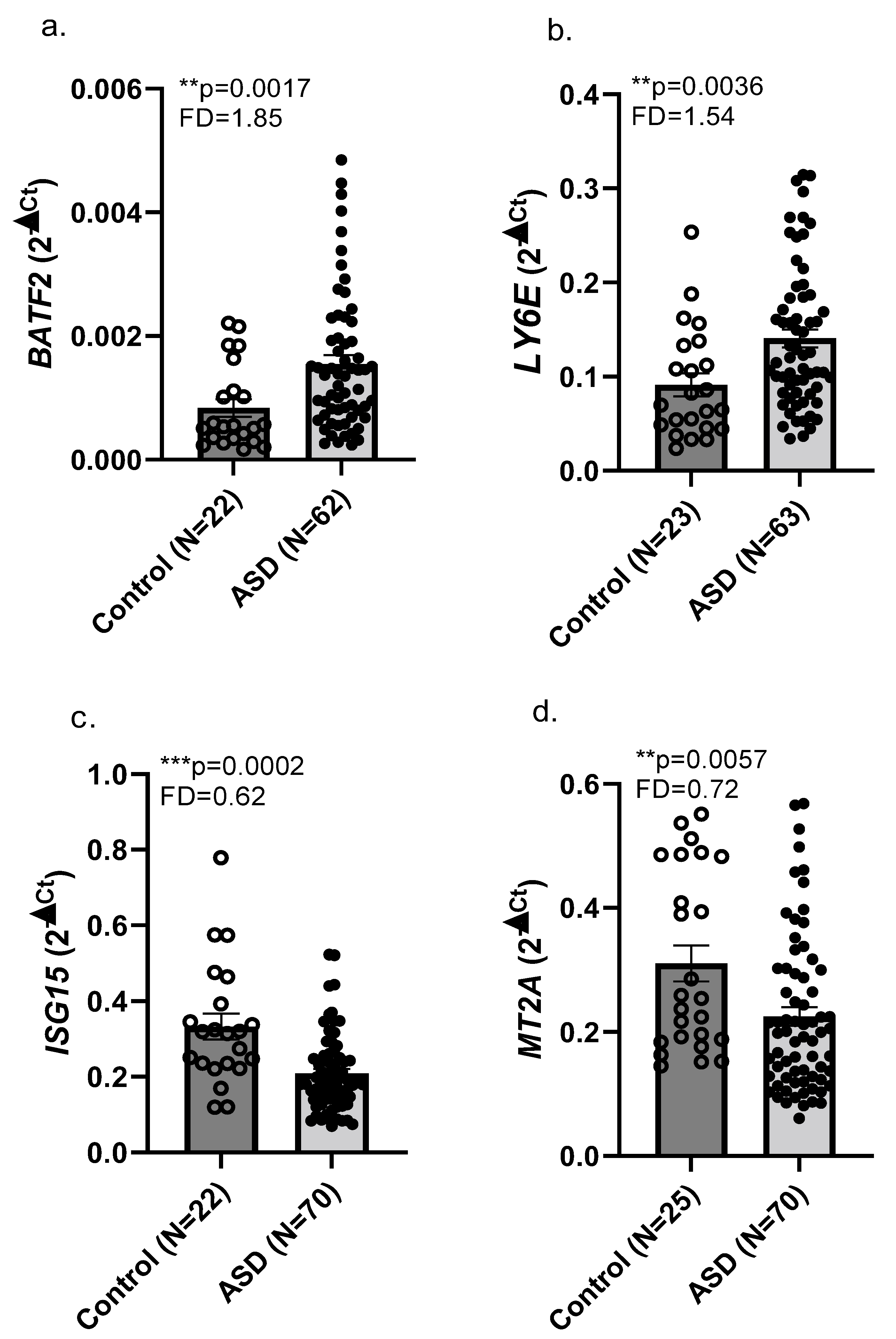
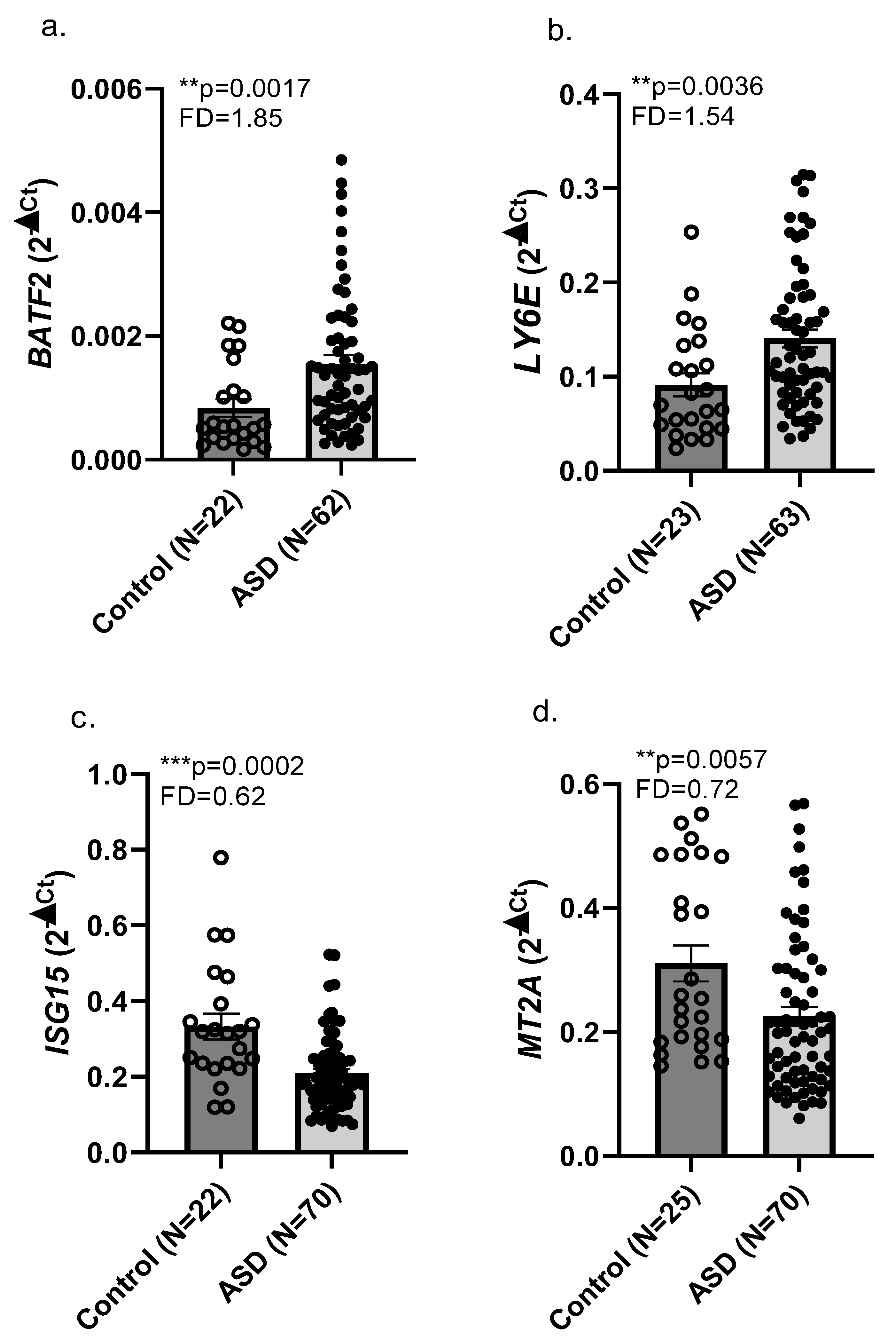
2.1. RNA Sequencing and Real-Time qPCR Validation
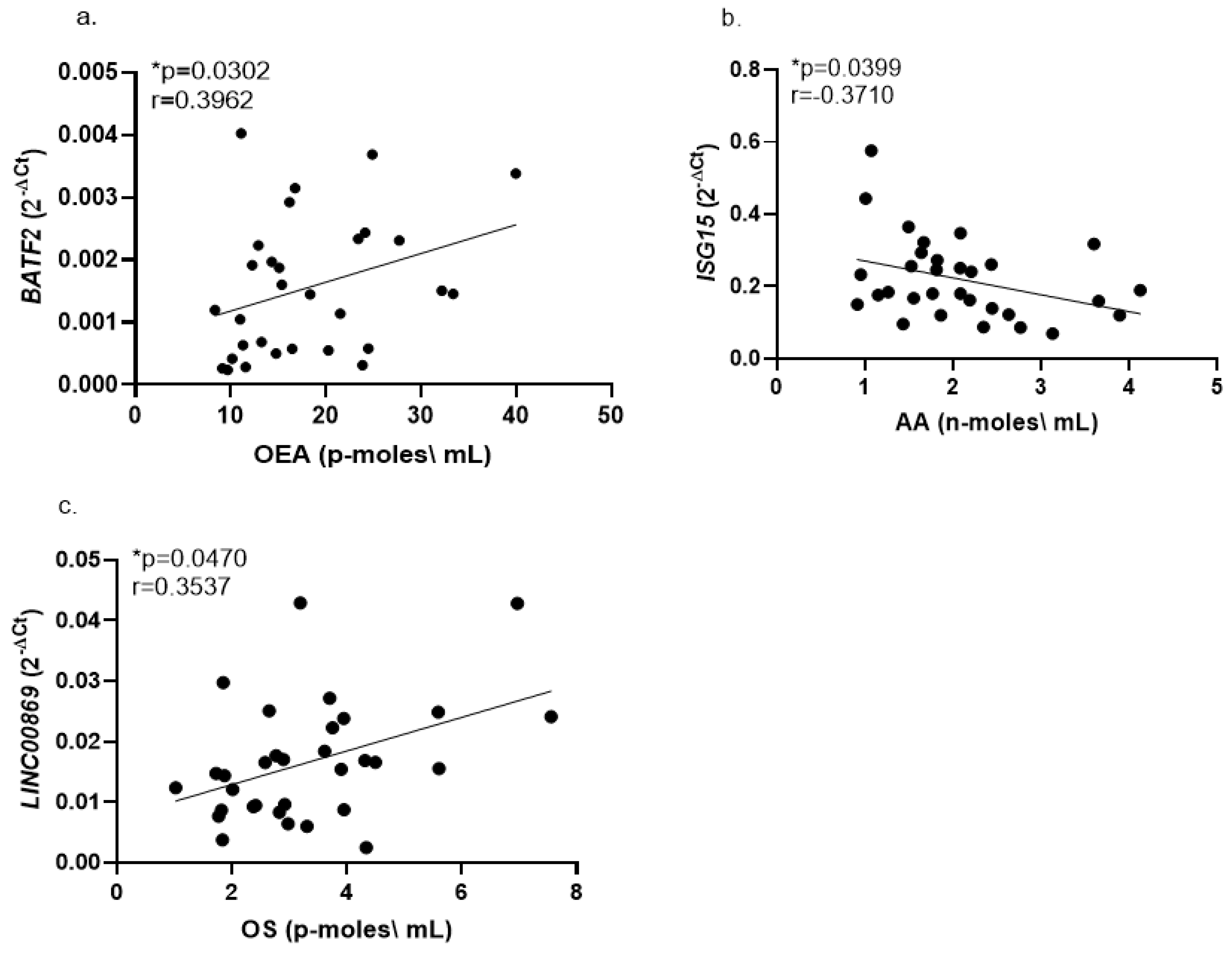
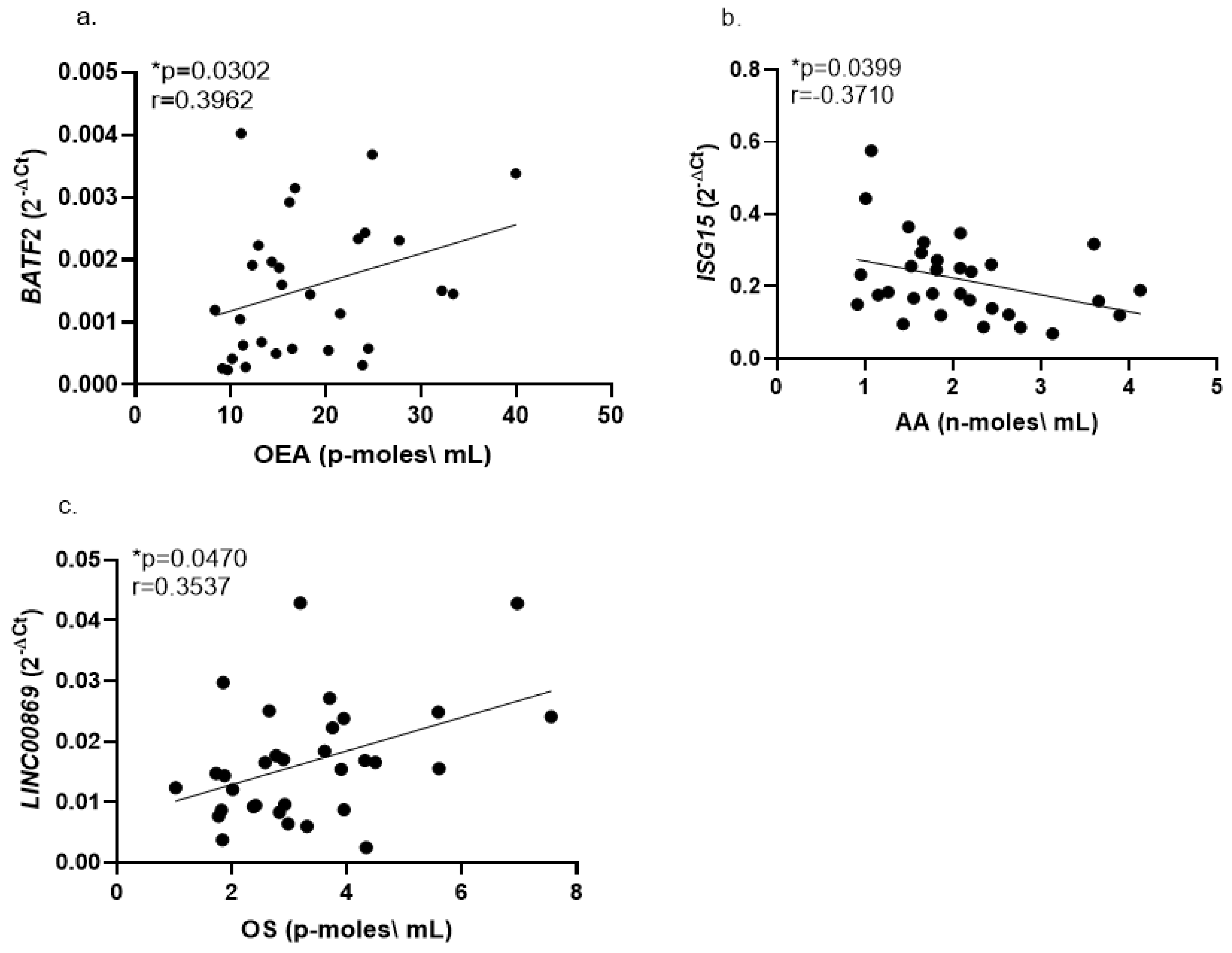
2.2. Correlations of Whole Blood Gene Expression Levels with Serum Endocannabinoids
3. Discussion
3.1. Upregulated BATF2 and LY6E and Downregulated ISG15 and MT2A Expression in Whole Blood of Children with ASD Compared with Neurotypical Children
3.2. Dysregulated ASD Genes and Cancer
3.3. Dysregulated ASD Genes and Immunity
3.4. Correlations of Whole Blood Gene Expression Levels with Serum Endocannabinoids
3.5. Study Limitations
4. Materials and Methods
4.1. Participants
4.2. Whole Blood Sample Collection
4.3. RNA Extraction
4.4. Library Preparation, RNA Sequencing, and Data Processing
4.5. Real-Time qPCR Validation
4.6. Correlations and Statistical Analysis
4.7. Behavioral Tests
4.8. Literature Survey of Transcriptomic Studies in Whole Blood from Individuals with ASD and Controls
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaaf, C.P.; Betancur, C.; Yuen, R.K.C.; Parr, J.R.; Skuse, D.H.; Gallagher, L.; Bernier, R.A.; Buchanan, J.A.; Buxbaum, J.; Chen, C.-A.; et al. A framework for an evidence-based gene list relevant to autism spectrum disorder. Nat. Rev. Genet. 2020, 21, 367–376. [Google Scholar] [CrossRef]
- Yoon, S.H.; Choi, J.; Lee, W.J.; Do, J.T. Genetic and Epigenetic Etiology Underlying Autism Spectrum Disorder. J. Clin. Med. 2020, 9, 966. [Google Scholar] [CrossRef]
- Frye, R.E.; Cakir, J.; Rose, S.; Palmer, R.F.; Austin, C.; Curtin, P. Physiological mediators of prenatal environmental influences in autism spectrum disorder. BioEssays 2021, 43, e2000307. [Google Scholar] [CrossRef]
- Rose, S.; Bennuri, S.C.; Wynne, R.; Melnyk, S.; James, S.J.; Frye, R.E. Mitochondrial and redox abnormalities in autism lymphoblastoid cells: A sibling control study. FASEB J. 2017, 31, 904–909. [Google Scholar] [CrossRef]
- Frye, R.E. Mitochondrial Dysfunction in Autism Spectrum Disorder: Unique Abnormalities and Targeted Treatments. Semin. Pediatr. Neurol. 2020, 35, 100829. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.; Machado, A.S.; von Doellinger, O.; Almeida, M.I.; Barbosa, M.A.; Coelho, R.; Santos, S.G. The Contribution of Inflammation to Autism Spectrum Disorders: Recent Clinical Evidence. Methods Mol. Biol. 2019, 2011, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Saghazadeh, A.; Ataeinia, B.; Keynejad, K.; Abdolalizadeh, A.; Hirbod-Mobarakeh, A.; Rezaei, N. A meta-analysis of pro-inflammatory cytokines in autism spectrum disorders: Effects of age, gender, and latitude. J. Psychiatr. Res. 2019, 115, 90–102. [Google Scholar] [CrossRef]
- Casanova, M.F.; Frye, R.E.; Gillberg, C.; Casanova, E.L. Editorial: Comorbidity and Autism Spectrum Disorder. Front. Psychiatry 2020, 11, 617395. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, E. Could the gut microbiome be linked to autism? Nature 2020, 577, S14–S15. [Google Scholar] [CrossRef]
- Bakulski, K.M.; Dou, J.F.; Feinberg, J.I.; Aung, M.T.; Ladd-Acosta, C.; Volk, H.E.; Newschaffer, C.J.; Croen, L.A.; Hertz-Picciotto, I.; Levy, S.E.; et al. Autism-Associated DNA Methylation at Birth From Multiple Tissues Is Enriched for Autism Genes in the Early Autism Risk Longitudinal Investigation. Front. Mol. Neurosci. 2021, 14, 775390. [Google Scholar] [CrossRef]
- Hannon, E.; iPSYCH-Broad ASD Group; Schendel, D.; Ladd-Acosta, C.; Grove, J.; Hansen, C.S.; Andrews, S.V.; Hougaard, D.M.; Bresnahan, M.; Mors, O.; et al. Elevated polygenic burden for autism is associated with differential DNA methylation at birth. Genome Med. 2018, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- McCarty, P.; Frye, R.E. Early Detection and Diagnosis of Autism Spectrum Disorder: Why Is It So Difficult? Semin. Pediatr. Neurol. 2020, 35, 100831. [Google Scholar] [CrossRef] [PubMed]
- Wallis, K.E. The Roadmap to Early and Equitable Autism Identification. Pediatrics 2021, 148, s21–s24. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.V.; Ellis, S.E.; Bakulski, K.M.; Sheppard, B.; Croen, L.A.; Hertz-Picciotto, I.; Newschaffer, C.J.; Feinberg, A.P.; Arking, D.E.; Ladd-Acosta, C.; et al. Cross-tissue integration of genetic and epigenetic data offers insight into autism spectrum disorder. Nat. Commun. 2017, 8, 1011. [Google Scholar] [CrossRef] [PubMed]
- Supekar, K.; Ryali, S.; Mistry, P.; Menon, V. Aberrant dynamics of cognitive control and motor circuits predict distinct restricted and repetitive behaviors in children with autism. Nat. Commun. 2021, 12, 3537. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-J.; Vogelstein, J.T.; Gozzi, A.; Bernhardt, B.C.; Yeo, B.T.; Milham, M.P.; Di Martino, A. Toward Neurosubtypes in Autism. Biol. Psychiatry 2020, 88, 111–128. [Google Scholar] [CrossRef]
- Gurwitz, D. Expression profiling: A cost-effective biomarker discovery tool for the personal genome era. Genome Med. 2013, 5, 41. [Google Scholar] [CrossRef]
- Aran, A.; Eylon, M.; Harel, M.; Polianski, L.; Nemirovski, A.; Tepper, S.; Schnapp, A.; Cassuto, H.; Wattad, N.; Tam, J. Lower circulating endocannabinoid levels in children with autism spectrum disorder. Mol. Autism 2019, 10, 2. [Google Scholar] [CrossRef]
- Horiuchi, F.; Yoshino, Y.; Kumon, H.; Hosokawa, R.; Nakachi, K.; Kawabe, K.; Iga, J.-I.; Ueno, S.-I. Identification of aberrant innate and adaptive immunity based on changes in global gene expression in the blood of adults with autism spectrum disorder. J. Neuroinflamm. 2021, 18, 102. [Google Scholar] [CrossRef]
- Lee, S.C.; Quinn, T.P.; Lai, J.; Kong, S.W.; Hertz-Picciotto, I.; Glatt, S.J.; Crowley, T.M.; Venkatesh, S.; Nguyen, T. Solving for X: Evidence for sex-specific autism biomarkers across multiple transcriptomic studies. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2019, 180, 377–389. [Google Scholar] [CrossRef]
- Walker, S.J.; Beavers, D.P.; Fortunato, J.; Krigsman, A. A Putative Blood-Based Biomarker for Autism Spectrum Disorder-Associated Ileocolitis. Sci. Rep. 2016, 6, 35820. [Google Scholar] [CrossRef] [PubMed]
- Saffari, A.; Arno, M.; Nasser, E.; Ronald, A.; Wong, C.C.Y.; Schalkwyk, L.C.; Mill, J.; Dudbridge, F.; Meaburn, E.L. RNA sequencing of identical twins discordant for autism reveals blood-based signatures implicating immune and transcriptional dysregulation. Mol. Autism 2019, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Filosi, M.; Kam-Thong, T.; Essioux, L.; Muglia, P.; Trabetti, E.; Spooren, W.; Müller-Myshok, B.; Alibrio, G.; Anchisi, L.; Andruccioli, M.; et al. Transcriptome signatures from discordant sibling pairs reveal changes in peripheral blood immune cell composition in Autism Spectrum Disorder. Transl. Psychiatry 2020, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Gregg, J.P.; Lit, L.; Baron, C.A.; Hertz-Picciotto, I.; Walker, W.; Davis, R.A.; Croen, L.A.; Ozonoff, S.; Hansen, R.; Pessah, I.N.; et al. Gene expression changes in children with autism. Genomics 2008, 91, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.W.; Collins, C.D.; Shimizu-Motohashi, Y.; Holm, I.; Campbell, M.G.; Lee, I.-H.; Brewster, S.J.; Hanson, E.; Harris, H.K.; Lowe, K.R.; et al. Characteristics and Predictive Value of Blood Transcriptome Signature in Males with Autism Spectrum Disorders. PLoS ONE 2012, 7, e49475. [Google Scholar] [CrossRef]
- Glatt, S.J.; Tsuang, M.T.; Winn, M.; Chandler, S.D.; Collins, M.; Lopez, L.; Weinfeld, M.; Carter, C.; Schork, N.; Pierce, K.; et al. Blood-Based Gene Expression Signatures of Infants and Toddlers with Autism. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 934–944.e2. [Google Scholar] [CrossRef]
- Kanemaru, H.; Yamane, F.; Fukushima, K.; Matsuki, T.; Kawasaki, T.; Ebina, I.; Kuniyoshi, K.; Tanaka, H.; Maruyama, K.; Maeda, K.; et al. Antitumor effect of Batf2 through IL-12 p40 up-regulation in tumor-associated macrophages. Proc. Natl. Acad. Sci. USA 2017, 114, E7331–E7340. [Google Scholar] [CrossRef]
- Xie, J.-W.; Huang, X.-B.; Chen, Q.-Y.; Ma, Y.-B.; Zhao, Y.-J.; Liu, L.-C.; Wang, J.-B.; Lin, J.-X.; Lu, J.; Cao, L.-L.; et al. m6A modification-mediated BATF2 acts as a tumor suppressor in gastric cancer through inhibition of ERK signaling. Mol. Cancer 2020, 19, 114. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Y.; Dai, L.; Shi, G.; Deng, J.; Luo, Q.; Xie, Q.; Cheng, L.; Li, C.; Lin, Y.; et al. BATF2 prevents glioblastoma multiforme progression by inhibiting recruitment of myeloid-derived suppressor cells. Oncogene 2021, 40, 1516–1530. [Google Scholar] [CrossRef]
- Han, T.; Wang, Z.; Yang, Y.; Shu, T.; Li, W.; Liu, D.; Li, P.; Qi, R.; Ren, Y.; Li, L.; et al. The tumor-suppressive role of BATF2 in esophageal squamous cell carcinoma. Oncol. Rep. 2015, 34, 1353–1360. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Jia, Y.; Pan, P.; Zhang, X.; Jia, Y.; Zhu, P.; Chen, X.; Jiao, Y.; Kang, G.; Zhang, L.; et al. α5-nAChR associated with Ly6E modulates cell migration via TGF-β1/Smad signaling in non-small cell lung cancer. Carcinogenesis 2022, 43, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, W.; Wang, C.; Wang, M.; Hui, W.; Lu, J.; Gao, F. Multidimension Analysis of the Prognostic Value, Immune Regulatory Function, and ceRNA Network of LY6E in Individuals with Colorectal Cancer. J. Immunol. Res. 2022, 2022, 5164265. [Google Scholar] [CrossRef]
- Tolaney, S.M.; Do, K.T.; Eder, J.P.; LoRusso, P.M.; Weekes, C.D.; Chandarlaplaty, S.; Chang, C.-W.; Chen, S.-C.; Nazzal, D.; Schuth, E.; et al. A Phase I Study of DLYE5953A, an Anti-LY6E Antibody Covalently Linked to Monomethyl Auristatin E, in Patients with Refractory Solid Tumors. Clin. Cancer Res. 2020, 26, 5588–5597. [Google Scholar] [CrossRef] [PubMed]
- Yip, V.; Figueroa, I.; Latifi, B.; Masih, S.; Ng, C.; Leipold, D.; Kamath, A.; Shen, B.-Q. Anti-Lymphocyte Antigen 6 Complex, Locus E-Seco-Cyclopropabenzindol-4-One-Dimer Antibody-Drug Conjugate That Forms Adduct with α1-Microglobulin Demonstrates Slower Systemic Antibody Clearance and Reduced Tumor Distribution in Animals. Drug Metab. Dispos. 2020, 48, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Chuh, J.D.C.; Go, M.; Chen, Y.; Guo, J.; Rafidi, H.; Mandikian, D.; Sun, Y.; Lin, Z.; Schneider, K.; Zhang, P.; et al. Preclinical optimization of Ly6E-targeted ADCs for increased durability and efficacy of anti-tumor response. mAbs 2021, 13, 1862452. [Google Scholar] [CrossRef]
- Han, H.G.; Moon, H.W.; Jeon, Y.J. ISG15 in cancer: Beyond ubiquitin-like protein. Cancer Lett. 2018, 438, 52–62. [Google Scholar] [CrossRef]
- Alcalá, S.; Sancho, P.; Martinelli, P.; Navarro, D.; Pedrero, C.; Martín-Hijano, L.; Valle, S.; Earl, J.; Rodríguez-Serrano, M.; Ruiz-Cañas, L.; et al. ISG15 and ISGylation is required for pancreatic cancer stem cell mitophagy and metabolic plasticity. Nat. Commun. 2020, 11, 2682. [Google Scholar] [CrossRef]
- Kariri, Y.A.; Alsaleem, M.; Joseph, C.; Alsaeed, S.; Aljohani, A.; Shiino, S.; Mohammed, O.J.; Toss, M.S.; Green, A.R.; Rakha, E.A. The prognostic significance of interferon-stimulated gene 15 (ISG15) in invasive breast cancer. Breast Cancer Res. Treat. 2021, 185, 293–305. [Google Scholar] [CrossRef]
- Shin, C.-H.; Lee, M.-G.; Han, J.; Jeong, S.-I.; Ryu, B.-K.; Chi, S.-G. Identification of XAF1–MT2A mutual antagonism as a molecular switch in cell-fate decisions under stressful conditions. Proc. Natl. Acad. Sci. USA 2017, 114, 5683–5688. [Google Scholar] [CrossRef]
- Mangelinck, A.; Da Costa, M.E.M.; Stefanovska, B.; Bawa, O.; Polrot, M.; Gaspar, N.; Fromigué, O. MT2A is an early predictive biomarker of response to chemotherapy and a potential therapeutic target in osteosarcoma. Sci. Rep. 2019, 9, 12301. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Du, T.; Wu, W.; Chen, X.; Lai, Y.; Zhu, D.; Wang, Q.; Ma, X.; Lin, C.; Li, Z.; et al. Decreased expression of serine protease inhibitor family G1 (SERPING1) in prostate cancer can help distinguish high-risk prostate cancer and predicts malignant progression. Urol. Oncol. Semin. Orig. Investig. 2018, 36, 366.e1–366.e9. [Google Scholar] [CrossRef] [PubMed]
- Sbattella, M.; Zanichelli, A.; Ghia, P.; Gattei, V.; Suffritti, C.; Teatini, T.; Cicardi, M.; Castelli, R. Splenic marginal zone lymphomas in acquired C1-inhibitor deficiency: Clinical and molecular characterization. Med. Oncol. 2018, 35, 118. [Google Scholar] [CrossRef] [PubMed]
- Darbro, B.W.; Singh, R.; Zimmerman, M.B.; Mahajan, V.; Bassuk, A.G. Autism Linked to Increased Oncogene Mutations but Decreased Cancer Rate. PLoS ONE 2016, 11, e0149041. [Google Scholar] [CrossRef] [PubMed]
- Bishop-Fitzpatrick, L.; Movaghar, A.; Greenberg, J.S.; Page, D.; Dawalt, L.S.; Brilliant, M.H.; Mailick, M.R. Using machine learning to identify patterns of lifetime health problems in decedents with autism spectrum disorder. Autism Res. 2018, 11, 1120–1128. [Google Scholar] [CrossRef]
- Chiang, H.-L.; Liu, C.-J.; Hu, Y.-W.; Chen, S.-C.; Hu, L.-Y.; Shen, C.-C.; Yeh, C.-M.; Chen, T.-J.; Gau, S.S.-F. Risk of Cancer in Children, Adolescents, and Young Adults with Autistic Disorder. J. Pediatr. 2015, 166, 418–423.e1. [Google Scholar] [CrossRef]
- Hirvikoski, T.; Mittendorfer-Rutz, E.; Boman, M.; Larsson, H.; Lichtenstein, P.; Bölte, S. Premature mortality in autism spectrum disorder. Br. J. Psychiatry 2016, 208, 232–238. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Frye, R.E. A review of research trends in physiological abnormalities in autism spectrum disorders: Immune dysregulation, inflammation, oxidative stress, mitochondrial dysfunction and environmental toxicant exposures. Mol. Psychiatry 2012, 17, 389–401. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat. Rev. Neurosci. 2015, 16, 469–486. [Google Scholar] [CrossRef]
- Roy, S.; Guler, R.; Parihar, S.P.; Schmeier, S.; Kaczkowski, B.; Nishimura, H.; Shin, J.W.; Negishi, Y.; Ozturk, M.; Hurdayal, R.; et al. Batf2/Irf1 Induces Inflammatory Responses in Classically Activated Macrophages, Lipopolysaccharides, and Mycobacterial Infection. J. Immunol. 2015, 194, 6035–6044. [Google Scholar] [CrossRef]
- Altmeyer, A.; Staruch, M.J.; Cofano, F.; Landolfo, S.; Dumont, F.J. Multiple cytokine interactions regulate Ly-6E antigen expression: Cooperative Ly-6E induction by IFNs, TNF, and IL-1 in a T cell lymphoma and in its induction-deficient variants. Cell. Immunol. 1991, 138, 94–107. [Google Scholar] [CrossRef]
- Fan, G.; Liu, J.; Zhang, Y.; Guan, X. LINC00473 exacerbates osteoarthritis development by promoting chondrocyte apoptosis and proinflammatory cytokine production through the miR-424-5p/LY6E axis. Exp. Ther. Med. 2021, 22, 1247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bogunovic, D.; Payelle-Brogard, B.; Francois-Newton, V.; Speer, S.D.; Yuan, C.; Volpi, S.; Li, Z.; Sanal, O.; Mansouri, D.; et al. Human intracellular ISG15 prevents interferon-α/β over-amplification and auto-inflammation. Nature 2015, 517, 89–93. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, P.F.; Van Weyenbergh, J.; Delgobo, M.; Patricio, D.D.O.; Ferguson, B.J.; Guabiraba, R.; Dierckx, T.; Menezes, S.M.; Báfica, A.; Mansur, D.S. ISG15-Induced IL-10 Is a Novel Anti-Inflammatory Myeloid Axis Disrupted during Active Tuberculosis. J. Immunol. 2018, 200, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Junior, E.A.D.S.; Medeiros, W.M.B.; Torro, N.; de Sousa, J.M.M.; de Almeida, I.B.C.M.; da Costa, F.B.; Pontes, K.M.; Nunes, E.L.G.; da Rosa, M.D.; de Albuquerque, K.L.G.D. Cannabis and cannabinoid use in autism spectrum disorder: A systematic review. Trends Psychiatry Psychother. 2021. [Google Scholar] [CrossRef]
- Zamberletti, E.; Gabaglio, M.; Parolaro, D. The Endocannabinoid System and Autism Spectrum Disorders: Insights from Animal Models. Int. J. Mol. Sci. 2017, 18, 1916. [Google Scholar] [CrossRef]
- Smith, D.R.; Stanley, C.M.; Foss, T.; Boles, R.G.; McKernan, K. Rare genetic variants in the endocannabinoid system genes CNR1 and DAGLA are associated with neurological phenotypes in humans. PLoS ONE 2017, 12, e0187926. [Google Scholar] [CrossRef]
- Hillard, C.J. Circulating Endocannabinoids: From Whence Do They Come and Where are They Going? Neuropsychopharmacology 2018, 43, 155–172. [Google Scholar] [CrossRef]
- Lerner, R.; Post, J.M.; Loch, S.; Lutz, B.; Bindila, L. Targeting brain and peripheral plasticity of the lipidome in acute kainic acid-induced epileptic seizures in mice via quantitative mass spectrometry. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 255–267. [Google Scholar] [CrossRef]
- Pietropaolo, S.; Bellocchio, L.; Bouzón-Arnáiz, I.; Yee, B.K. The role of the endocannabinoid system in autism spectrum disorders: Evidence from mouse studies. Prog. Mol. Biol. Transl. Sci. 2020, 173, 183–208. [Google Scholar] [CrossRef]
- Fyke, W.; Premoli, M.; Alzate, V.E.; López-Moreno, J.A.; Lemaire-Mayo, V.; Crusio, W.E.; Marsicano, G.; Wöhr, M.; Pietropaolo, S. Communication and social interaction in the cannabinoid-type 1 receptor null mouse: Implications for autism spectrum disorder. Autism Res. 2021, 14, 1854–1872. [Google Scholar] [CrossRef]
- Ueda, N.; Yamanaka, K.; Yamamoto, S. Purification and Characterization of an Acid Amidase Selective for N-Palmitoylethanolamine, a Putative Endogenous Anti-inflammatory Substance. J. Biol. Chem. 2001, 276, 35552–35557. [Google Scholar] [CrossRef] [PubMed]
- Citraro, R.; Russo, E.; Scicchitano, F.; van Rijn, C.M.; Cosco, D.; Avagliano, C.; Russo, R.; D’Agostino, G.; Petrosino, S.; Guida, F.; et al. Antiepileptic action of N-palmitoylethanolamine through CB1 and PPAR-α receptor activation in a genetic model of absence epilepsy. Neuropharmacology 2013, 69, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, L.D.C.; Pacini, A.; Corti, F.; Boccella, S.; Luongo, L.; Esposito, E.; Cuzzocrea, S.; Maione, S.; Calignano, A.; Ghelardini, C. Antineuropathic Profile of N-Palmitoylethanolamine in a Rat Model of Oxaliplatin-Induced Neurotoxicity. PLoS ONE 2015, 10, e0128080. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Israeli Cohort | U.S. Cohort | |||
|---|---|---|---|---|
| Neurotypical controls | Children with ASD | Neurotypical controls | Children with ASD | |
| N | 21 | 36 | 5 | 37 |
| Age (years) | 16.3 ± 2.7 | 13.2 ± 4.8 | 8.9 ± 5.4 | 7.4 ± 3.7 |
| % Male | 75% | 83% | 76% | 84% |
| Behavioral scores | ||||
| VABS | N/A | 46 ± 11 | N/A | N/A |
| SRS * | N/A | 78 ± 10 | N/A | 78 ± 13 |
| CBCL | N/A | 55 ± 11 | N/A | N/A |
| ABC * | N/A | N/A | 21 ± 29 | 60 ± 22 |
| Gene Name | Gene ID | Mean Reads | FD | p-Value | Padj |
|---|---|---|---|---|---|
| SERPING1 | ENSG00000149131 | 494.495 | 3.4962 | 3.73 × 10−7 | 0.0072 |
| EFHC2 | ENSG00000183690 | 93.312 | 0.6086 | 2.81 × 10−6 | 0.0261 |
| BATF2 | ENSG00000168062 | 127.616 | 3.4360 | 4.06 × 10−6 | 0.0261 |
| CDC20 | ENSG00000117399 | 66.744 | 3.1642 | 6.06 × 10−6 | 0.0292 |
| FCGR1A | ENSG00000150337 | 416.310 | 2.6044 | 1.37 × 10−5 | 0.0379 |
| MT2A | ENSG00000125148 | 407.810 | 1.8252 | 1.35 × 10−5 | 0.0379 |
| ISG15 | ENSG00000187608 | 1262.341 | 3.1128 | 1.36 × 10−5 | 0.0379 |
| FBXO6 | ENSG00000116663 | 365.625 | 1.5121 | 1.89 × 10−5 | 0.0416 |
| LINC00869 | ENSG00000277147 | 306.080 | 1.4388 | 1.94 × 10−5 | 0.0416 |
| LY6E | ENSG00000160932 | 3225.314 | 1.8098 | 2.49 × 10−5 | 0.0481 |
| Study | Dysregulated Genes |
|---|---|
| a. RNA-seq studies | |
| Walker et al., 2016 [21] | CYP2S1, TNFRSF12A, IL1RN, TNFAIP3, SIGLECP3 |
| Saffari et al., 2019 [22] | IGHG4, PRR13P5, DEPDC1B, ZNF501, EVI2A, SNORD15B |
| Filosiet al., 2020 [23] | NMUR1, HMGB3, PTPRN2 |
| Horiuchi et al., 2021 [19] | ETV7, BATF2, SERPING1, CD274, FCGR1A, WARS, CASP5, PI3, RPSAP58, GZMK, KLRB1, GZMA, DUSP2, GNLY, A2M-AS1, TOMM7, LINC00612, SNORD3A, FCGR1B, TMEM176B |
| b. Meta-analysis of blood-based microarray data sets | |
| Gregg et al., 2007 [24] | PAM, SPON2, IL2RB, PRF1, GZMB, CX3CR1, SH2D1B/EAT |
| Kong et al., 2012 [25] | CREBZF, HNRNPA2B1, KIDINS220, LBR, MED23, ZDHHC17, ZMAT1, ZNF12, SULF2, RBBP6, SPATA13, TMEM30A |
| Glatt et al., 2012 [26] | FCGR1A, ANKRD22, GBP5, L-RAP, FCGR1B, GBP1, LGALS2, ANXA3, LILRA3, ADM, PLSCR1, NAMPT, HLA-H, STX11, PTER, BC050625, SLC22A4, IL1RN, BTN3A2, S100A12, SLC26A8, GNG10, DQ893795, USP10, PARP9, CST7, GCH1 |
| Lee et al., 2019 [20] | TTF2, UTY, KCNJ8, NCS1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voinsky, I.; Zoabi, Y.; Shomron, N.; Harel, M.; Cassuto, H.; Tam, J.; Rose, S.; Scheck, A.C.; Karim, M.A.; Frye, R.E.; et al. Blood RNA Sequencing Indicates Upregulated BATF2 and LY6E and Downregulated ISG15 and MT2A Expression in Children with Autism Spectrum Disorder. Int. J. Mol. Sci. 2022, 23, 9843. https://doi.org/10.3390/ijms23179843
Voinsky I, Zoabi Y, Shomron N, Harel M, Cassuto H, Tam J, Rose S, Scheck AC, Karim MA, Frye RE, et al. Blood RNA Sequencing Indicates Upregulated BATF2 and LY6E and Downregulated ISG15 and MT2A Expression in Children with Autism Spectrum Disorder. International Journal of Molecular Sciences. 2022; 23(17):9843. https://doi.org/10.3390/ijms23179843
Chicago/Turabian StyleVoinsky, Irena, Yazeed Zoabi, Noam Shomron, Moria Harel, Hanoch Cassuto, Joseph Tam, Shannon Rose, Adrienne C. Scheck, Mohammad A. Karim, Richard E. Frye, and et al. 2022. "Blood RNA Sequencing Indicates Upregulated BATF2 and LY6E and Downregulated ISG15 and MT2A Expression in Children with Autism Spectrum Disorder" International Journal of Molecular Sciences 23, no. 17: 9843. https://doi.org/10.3390/ijms23179843
APA StyleVoinsky, I., Zoabi, Y., Shomron, N., Harel, M., Cassuto, H., Tam, J., Rose, S., Scheck, A. C., Karim, M. A., Frye, R. E., Aran, A., & Gurwitz, D. (2022). Blood RNA Sequencing Indicates Upregulated BATF2 and LY6E and Downregulated ISG15 and MT2A Expression in Children with Autism Spectrum Disorder. International Journal of Molecular Sciences, 23(17), 9843. https://doi.org/10.3390/ijms23179843