
Anti-Inflammatory Activity of Cajanin, an Isoflavonoid Derivative Isolated from Canavalia lineata Pods

 , and
, and
Abstract
:
1. Introduction
2. Results
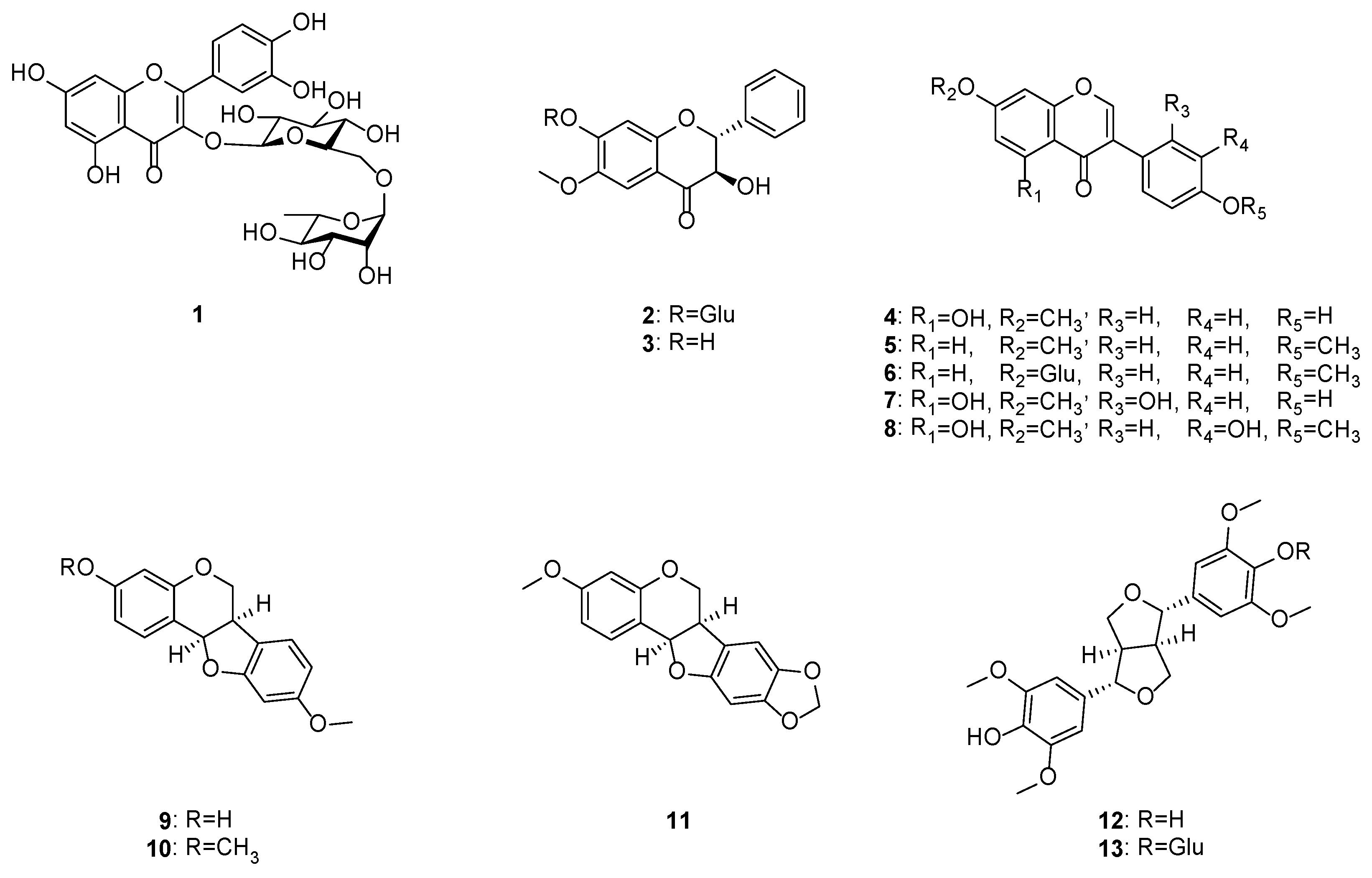
2.1. Isolation and Identification of Compounds 1–13 from C. lineata Pods
2.2. Elucidation of (2R,3R)-3-Hydroxy 7-O-β-d-Glucopyranoside-6-Methoxyflavanone (Compound 2)
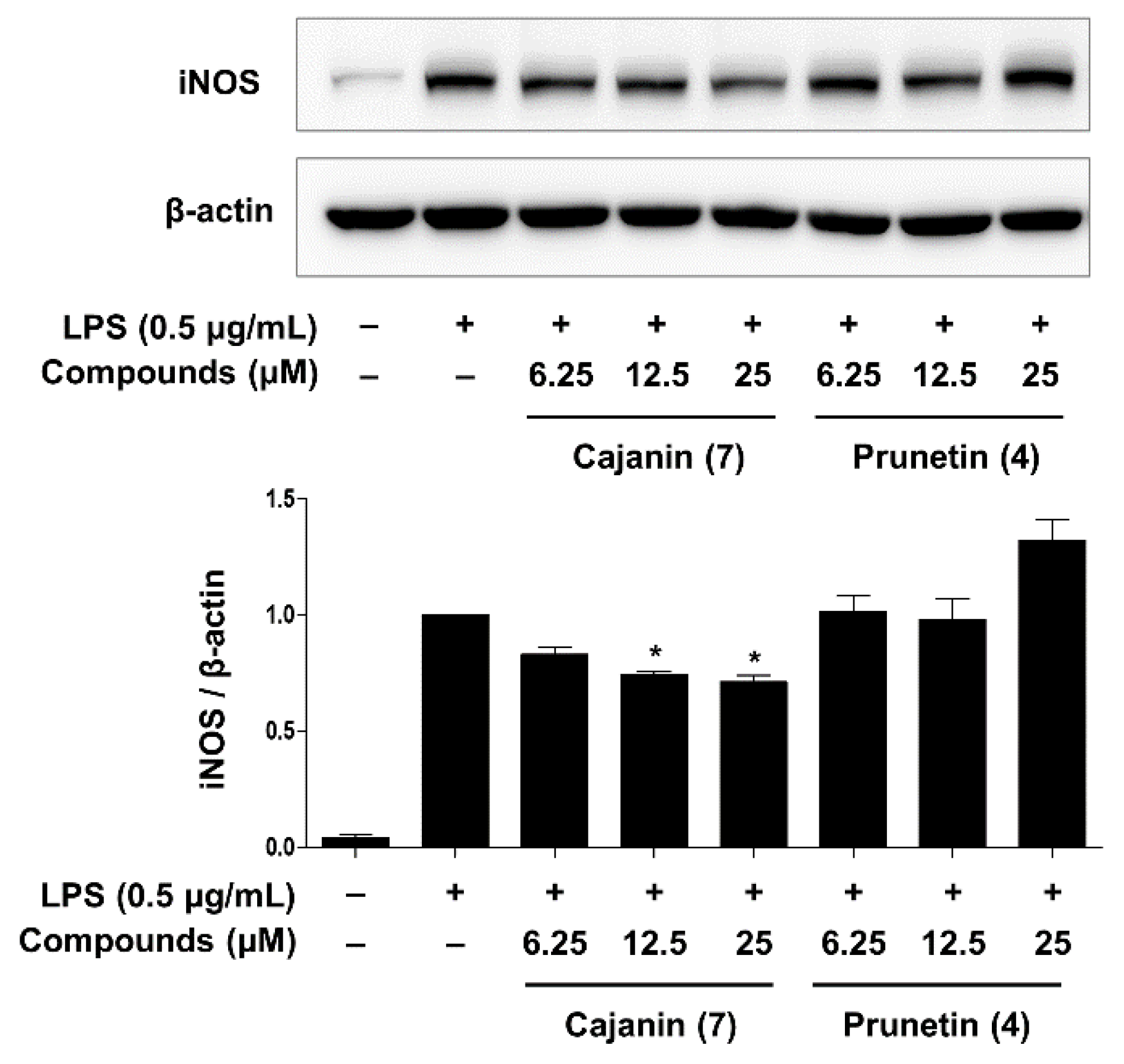
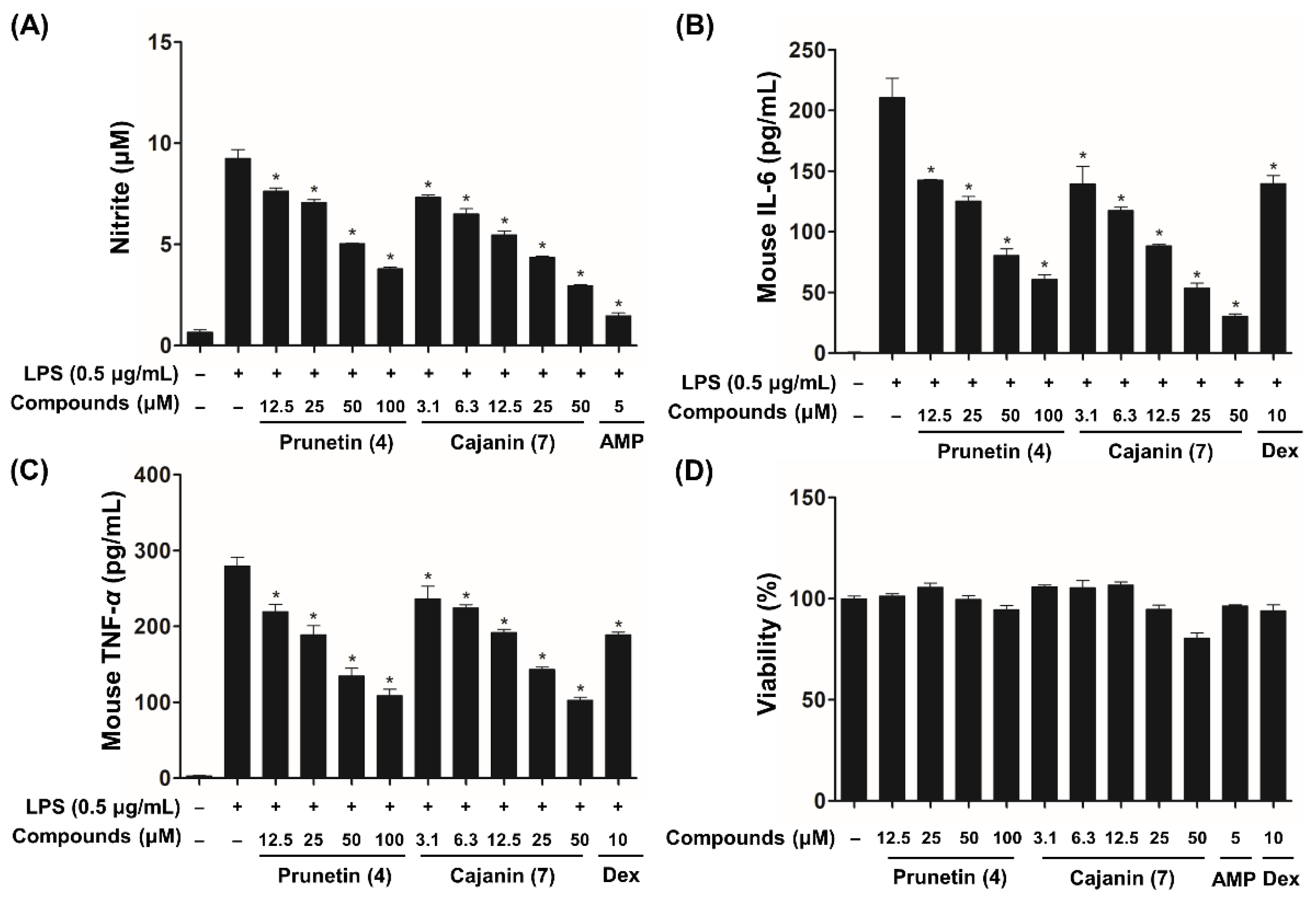
2.3. Inhibitory Effects of Compounds 4 and 7 on LPS-Enhanced Inflammatory Mediators
2.4. Anti-Inflammatory Effects of Compounds 4 and 7 through Inhibition of NF-κB Activation
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Materials
4.3. Extraction and Isolation
(2. R,3R)-3-Hydroxy 7-O-β-d-Glucopyranoside-6-Methoxyflavanone (2)
4.4. UPLC-QTOF-MS Analysis for Determining the Sugar Content
4.5. Cell Culture and Viability
4.6. Measurement of Nitric Oxide (NO) and Cytokine Production
4.7. Western Blot Analysis
4.8. Immunocytochemistry
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sridhar, K.R.; Seena, S. Nutritional and antinutritional significance of four unconventional legumes of the genus Canavalia—A comparative study. Food Chem. 2006, 99, 267–288. [Google Scholar] [CrossRef]
- Smartt, J. Grain Legumes; Cambridge University Press: Cambridge, MA, USA, 1990; pp. 301–309. [Google Scholar]
- Wang, T.L.; Domoney, C.; Hedley, C.L.; Casey, R.; Grusak, M.A. Can we improve the nutritional quality of legume seeds? Plant Physiol. 2003, 131, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.J.; Riu, K.Z.; Lee, S. Anti-melanogenesis effect of Canavalia lineata extract. J. Soc. Cosmet. Sci. Korean 2004, 30, 485–489. [Google Scholar]
- Bu, H.J.; Lee, H.J.; Yoo, E.S.; Jung, D.S.; Riu, K.Z.; Lee, S. Antioxidant effects and inhibitory effect on NO synthesis by extracts of Canavalia lineata. Korean J. Pharmacogn. 2004, 35, 338–345. [Google Scholar]
- Meizlish, M.L.; Franklin, R.A.; Zhou, X.; Medzhitov, R. Tissue homeostasis and inflammation. Annu. Rev. Immunol. 2021, 39, 557–581. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The cytokines of asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Shaw, A.T.; Gravallese, E.M. Mediators of inflammation and bone remodeling in rheumatic disease. Semin. Cell. Dev. Biol. 2016, 49, 2–10. [Google Scholar] [CrossRef]
- Kopf, M.; Bachmann, M.F.; Marsland, B.J. Averting inflammation by targeting the cytokine environment. Nat. Rev. Drug Discov. 2010, 9, 703–718. [Google Scholar] [CrossRef]
- Hamidzadeh, K.; Christensen, S.M.; Dalby, E.; Chandrasekaran, P.; Mosser, D.M. Macrophages and the recovery from acute and chronic inflammation. Annu. Rev. Physiol. 2017, 79, 567–592. [Google Scholar] [CrossRef]
- Xiao, T.S. Innate immunity and inflammation. Cell Mol. Immunol. 2017, 14, 506–517. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-Year update. J. Physiol Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef]
- Lima, C.S.; Mottin, M.; de Assis, L.R.; Mesquita, N.C.M.R.; Sousa, B.K.P.; Coimbra, L.D.; Santos, K.B.; Zorn, K.M.; Guido, R.V.C.; Ekins, S.; et al. Flavonoids from Pterogyne nitens as Zika virus NS2B-NS3 protease inhibitors. Bioorg. Chem. 2021, 109, 104719. [Google Scholar] [CrossRef]
- Maurya, R.; Yadav, D.K.; Singh, G.; Bhargavan, B.; Narayana Murthy, P.S.; Sahai, M.; Singh, M.M. Osteogenic activity of constituents from Butea monosperma. Bioorg. Med. Chem. Lett. 2009, 19, 610–613. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, Y.; Liu, C.; Shi, C.; Zhang, Y. Determination and isolation of potential α-glucosidase and xanthine oxidase inhibitors from Trifolium pratense L. by ultrafiltration liquid chromatography and high-speed countercurrent chromatography. Med. Chem. Res. 2016, 25, 1020–1029. [Google Scholar] [CrossRef]
- Veitch, N.C.; Sutton, P.S.; Kite, G.C.; Ireland, H.E. Six new isoflavones and a 5-deoxyflavonol glycoside from the leaves of Ateleia herbert-smithii. J. Nat. Prod. 2003, 66, 210–216. [Google Scholar] [CrossRef]
- Fedoreyev, S.A.; Bulgakov, V.P.; Grishchenko, O.V.; Veselova, M.V.; Krivoschekova, O.E.; Kulesh, N.I.; Denisenko, V.A.; Tchernoded, G.K.; Zhuravlev, Y.N. Isoflavonoid composition of a callus culture of the relict tree Maackia amurensis Rupr. et Maxim. J. Agric. Food Chem. 2008, 56, 7023–7031. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Chang, M.; Choi, S.H.; Oh, S.R.; Wu, H.H.; Zhu, Y.; Gao, X.M.; Wang, X.; Zhang, B.; Lim, D.S.; et al. Estrogenic effects of phytoestrogens derived from Flemingia strobilifera in MCF-7 cells and immature rats. Arch. Pharm. Res. 2018, 41, 519–529. [Google Scholar] [CrossRef]
- Chiang, C.M.; Chang, Y.J.; Wu, J.Y.; Chang, T.S. Production and anti-Melanoma activity of methoxyisoflavones from the biotransformation of genistein by two recombinant Escherichia coli Strains. Molecules 2017, 22, 87. [Google Scholar] [CrossRef]
- Song, Y.; Pan, L.; Li, W.; Si, Y.; Zhou, D.; Zheng, C.; Hao, X.; Jia, X.; Jia, Y.; Shi, M.; et al. Natural neuro-inflammatory inhibitors from Caragana turfanensis. Bioorg. Med. Chem. Lett. 2017, 27, 4765–4769. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Curtis-Long, M.J.; Kim, J.H.; Jeong, S.H.; Yang, M.S.; Lee, K.W.; Lee, W.S.; Park, K.H. Pterocarpans and flavanones from Sophora flavescens displaying potent neuraminidase inhibition. Bioorg. Med. Chem. Lett. 2008, 18, 6046–6049. [Google Scholar] [CrossRef]
- Liu, L.; Zou, M.; Yin, Q.; Zhang, Z.; Zhang, X. Phenylpropanoids from Liparis nervosa and their in vitro antioxidant and α-glucosidase inhibitory activities. Med. Chem. Res. 2021, 30, 1005–1010. [Google Scholar] [CrossRef]
- Matos, F.D.A.; Gottlieb, O.R.; Andrade, C.S. Flavonoids from Dalbergia ecastophyllum. Phytochemistry 1975, 14, 825–826. [Google Scholar] [CrossRef]
- Wang, Y.H.; Avula, B.; Fu, X.; Wang, M.; Khan, I.A. Simultaneous Determination of the Absolute Configuration of Twelve Monosaccharide Enantiomers from Natural Products in a Single Injection by a UPLC-UV/MS Method. Planta Med. 2012, 78, 834–837. [Google Scholar] [CrossRef]
- Faraci, W.S.; Nagel, A.A.; Verdries, K.A.; Vincent, L.A.; Xu, H.; Nichols, L.E.; Labasi, J.M.; Salter, E.D.; Pettipher, E.R. 2-Amino-4-methylpyridine as a potent inhibitor of inducible NO synthase activity in vitro and in vivo. Br. J. Pharmacol. 1996, 119, 1101–1108. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Imam, F.; Al-Harbi, M.M.; Ansari, M.A.; Zoheir, K.M.; Korashy, H.M.; Sayed-Ahmed, M.M.; Attia, S.M.; Shabanah, O.A.; Ahmad, S.F. Dexamethasone attenuates LPS-induced acute lung injury through inhibition of NF-κB, COX-2, and pro-inflammatory mediators. Immunol. Investig. 2016, 45, 349–369. [Google Scholar] [CrossRef]
- Moncada, S. Nitric oxide in the vasculature: Physiology and pathophysiology. Ann. N. Y. Acad. Sci. 1997, 811, 60–69. [Google Scholar] [CrossRef]
- Laroux, F.S.; Pavlick, K.P.; Hines, I.N.; Kawachi, S.; Harada, H.; Bharwani, S.; Hoffman, J.M.; Grisham, M.B. Role of nitric oxide in inflammation. Acta. Physiol. Scand. 2001, 173, 113–118. [Google Scholar] [CrossRef]
- Wullaert, A.; Bonnet, M.C.; Pasparakis, M. NF-κB in the regulation of epithelial homeostasis and inflammation. Cell Res. 2011, 21, 146–158. [Google Scholar] [CrossRef]
- Hwang, I.D.; Kim, S.G.; Kwon, Y.M. Canavanine metabolism in tissue cultures of Canavalia lineata. Plant Cell Tissue Organ. Cult. 1996, 45, 17–23. [Google Scholar] [CrossRef]
- Park, K.S.; Kwon, Y.M. The analysis of canavanine content in leaves, roots, and xylem exudate of Canavalia lineata. Korean J. Bot. 1990, 33, 119–126. [Google Scholar]
- Dirsch, V.M.; Stuppner, H.; Vollmar, A.M. The Griess assay: Suitable for a bio-guided fractionation of anti-inflammatory plant extracts? Planta Med. 1998, 64, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Yang, G.; Ham, I.; Choi, H.Y. Anti-inflammatory effect of prunetin via the suppression of NF-κB pathway. Food Chem. Toxicol. 2013, 58, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi-Tago, M.; Ohsawa, K.; Ishikawa, T.; Nakamura, F.; Ueda, F.; Narukawa, Y.; Kiuchi, F.; Tamura, H.; Tago, K.; Kasahara, T. Inhibitory effects of flavonoids extracted from Nepalese propolis on the LPS signaling pathway. Int. Immunopharmacol. 2016, 40, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Ryu, H.W.; Lee, S.U.; Kim, M.G.; Kwon, O.K.; Kim, M.O.; Oh, T.K.; Lee, J.K.; Kim, T.Y.; Lee, S.W.; et al. Pistacia weinmannifolia ameliorates cigarette smoke and lipopolysaccharide-induced pulmonary inflammation by inhibiting interleukin-8 production and NF-κB activation. Int. J. Mol. Med. 2019, 44, 949–959. [Google Scholar] [CrossRef]
- Kwon, O.K.; Lee, J.W.; Xuezhen, X.; Harmalkar, D.S.; Song, J.G.; Park, J.W.; Hwang, D.; Min, J.H.; Kim, J.H.; Han, H.K.; et al. DK-1108 exerts anti-inflammatory activity against phorbol 12-myristate 13-acetate-induced inflammation and protective effect against OVA-induced allergic asthma. Biomed. Pharmacother. 2020, 132, 110950. [Google Scholar] [CrossRef]
- Park, J.W.; Ryu, H.W.; Ahn, H.I.; Min, J.H.; Kim, S.M.; Kim, M.G.; Kwon, O.K.; Hwang, D.; Kim, S.Y.; Choi, S.; et al. The Anti-Inflammatory Effect of Trichilia martiana C. DC. in the lipopolysaccharide-stimulated inflammatory response in macrophages and airway epithelial cells and in LPS-challenged mice. J. Microbiol. Biotechnol. 2020, 30, 1614–1625. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 2 a | HMBC | |
|---|---|---|---|
| δH (J in Hz) | δC | H→C | |
| 2 | 5.20, d (11.9) | 83.9 | 3, 4, 1′, 2′, 6′ |
| 3 | 4.59, d (11.9) | 72.6 | 2, 4, 1′ |
| 4 | - | 192.6 | - |
| 5 | 7.21, s | 107.2 | 4, 6, 7, 9, 10 |
| 6 | - | 144.5 | - |
| 7 | - | 153.6 | - |
| 8 | 6.77, s | 103.3 | 4, 6, 7, 9, 10 |
| 9 | - | 156.6 | - |
| 10 | - | 112.3 | - |
| 1′ | - | 137.5 | - |
| 2′, 6′ | 7.55, d (7.0) | 128.1 | 2, 2′, 4′, 6′ |
| 3′, 5′ | 7.42, t (7.0) b | 128.2 | 1′, 3′, 5′ |
| 4′ | 7.39, t (7.0) b | 128.6 | 2′, 6′ |
| 1″ | 5.05, d (7.7) | 99.4 | 7, 5″ |
| 2″ | 3.25 m b | 73.0 | 1″, 2″, 3″ |
| 3″ | 3.25 m b | 76.6 | 1″, 2″, 3″ |
| 4″ | 3.37, m b | 77.0 | 5″, 6″ |
| 5″ | 3.14, t (9.1) | 69.4 | 3″, 4″, 6″ |
| 6″ | 3.63, dd (11.9, 9.1) | 60.5 | 4″, 5″ |
| 3.40, dd (11.9, 5.6) | |||
| OCH3-6 | 3.79, s | 55.9 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.-J.; Kwon, O.-K.; Hwang, D.; Goo, S.H.; Kim, D.-Y.; Kim, M.H.; Kim, S.-Y.; Jang, H.-J.; Oh, S.-R. Anti-Inflammatory Activity of Cajanin, an Isoflavonoid Derivative Isolated from Canavalia lineata Pods. Int. J. Mol. Sci. 2022, 23, 9492. https://doi.org/10.3390/ijms23169492
Hong S-J, Kwon O-K, Hwang D, Goo SH, Kim D-Y, Kim MH, Kim S-Y, Jang H-J, Oh S-R. Anti-Inflammatory Activity of Cajanin, an Isoflavonoid Derivative Isolated from Canavalia lineata Pods. International Journal of Molecular Sciences. 2022; 23(16):9492. https://doi.org/10.3390/ijms23169492
Chicago/Turabian StyleHong, Su-Jin, Ok-Kyoung Kwon, Daseul Hwang, Su Hyun Goo, Doo-Young Kim, Min Ha Kim, Soo-Young Kim, Hyun-Jae Jang, and Sei-Ryang Oh. 2022. "Anti-Inflammatory Activity of Cajanin, an Isoflavonoid Derivative Isolated from Canavalia lineata Pods" International Journal of Molecular Sciences 23, no. 16: 9492. https://doi.org/10.3390/ijms23169492
APA StyleHong, S.-J., Kwon, O.-K., Hwang, D., Goo, S. H., Kim, D.-Y., Kim, M. H., Kim, S.-Y., Jang, H.-J., & Oh, S.-R. (2022). Anti-Inflammatory Activity of Cajanin, an Isoflavonoid Derivative Isolated from Canavalia lineata Pods. International Journal of Molecular Sciences, 23(16), 9492. https://doi.org/10.3390/ijms23169492