
Panicle Apical Abortion 7 Regulates Panicle Development in Rice (Oryza sativa L.)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
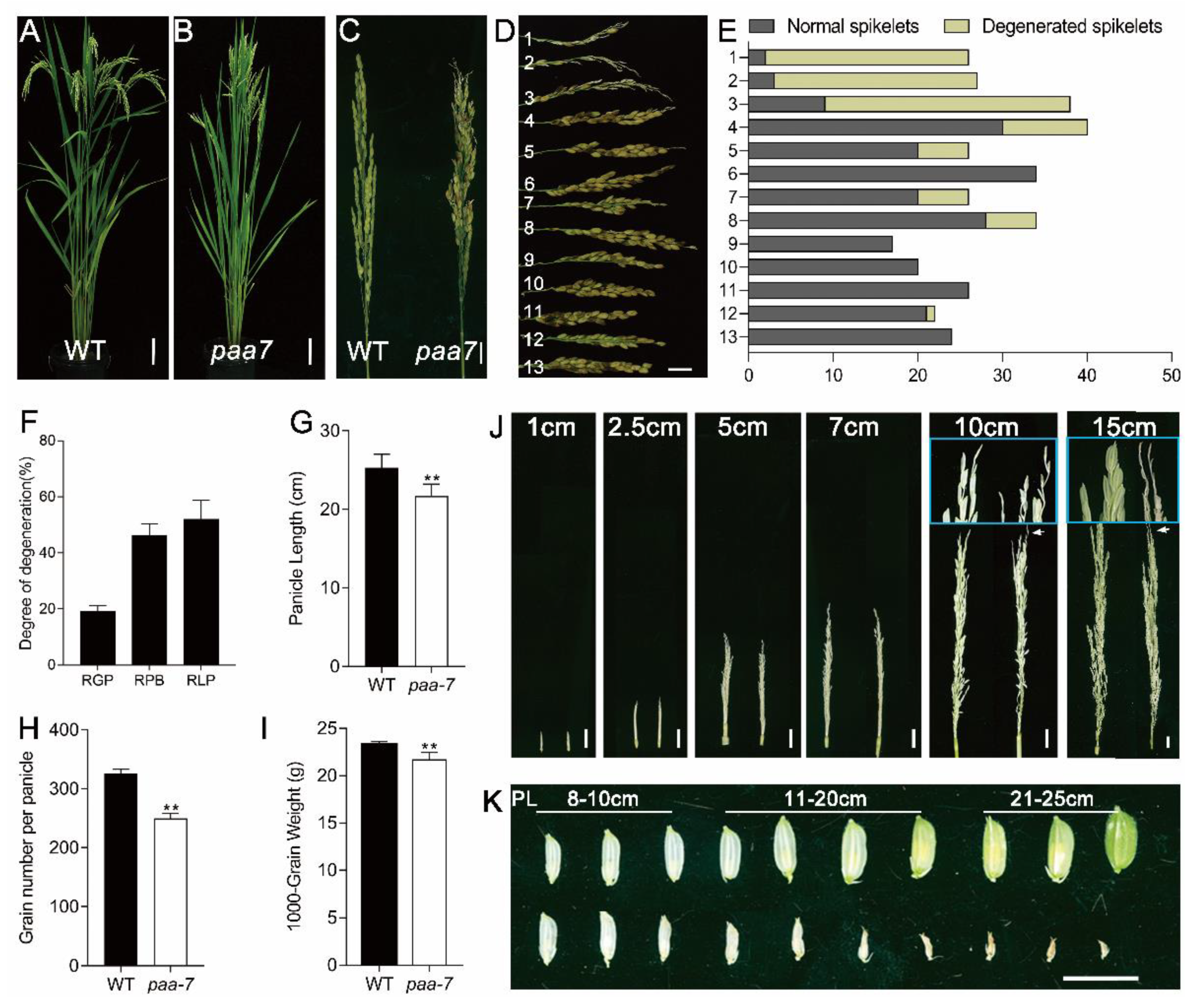
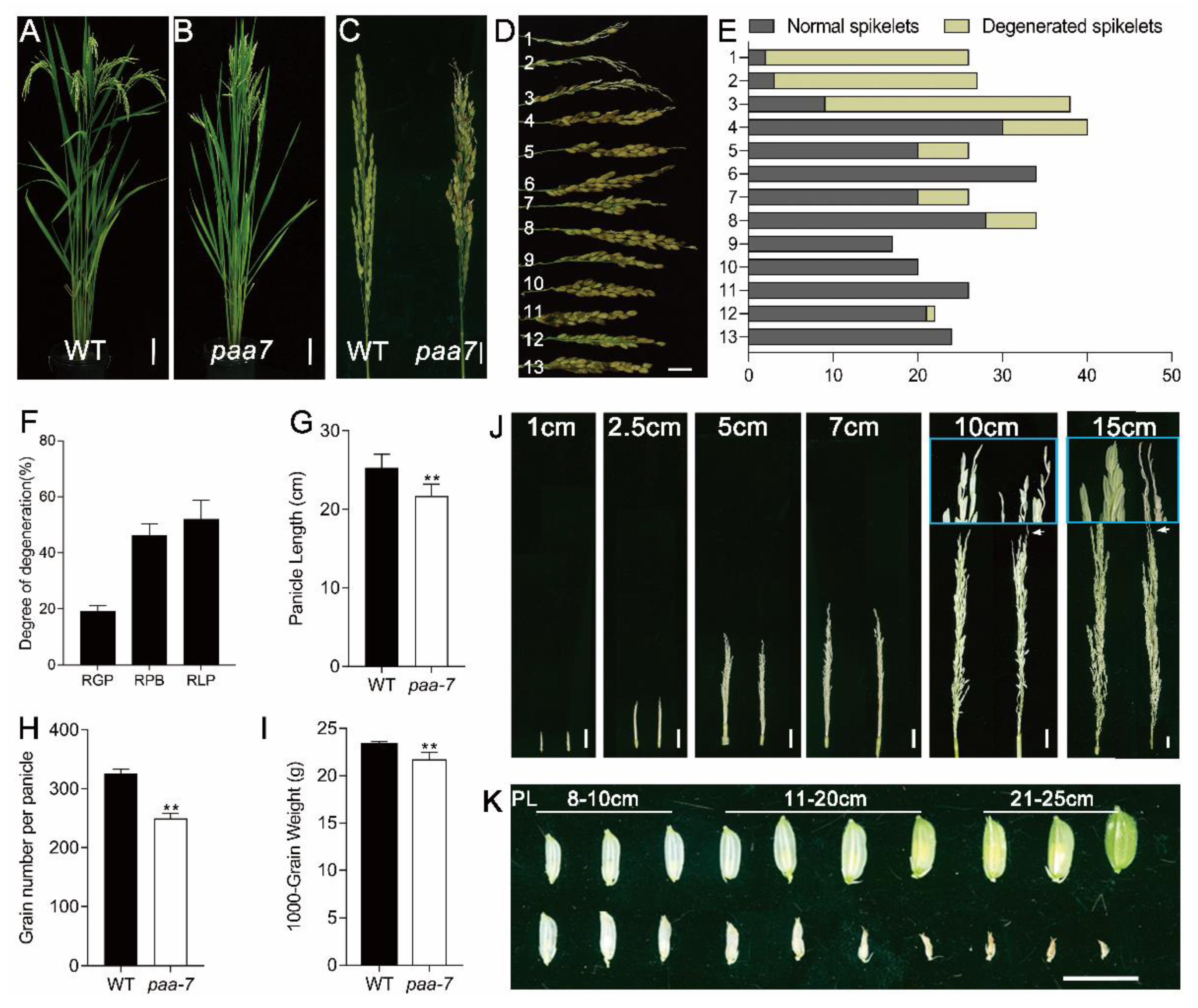
2.1. Characterization of the paa7 Mutant
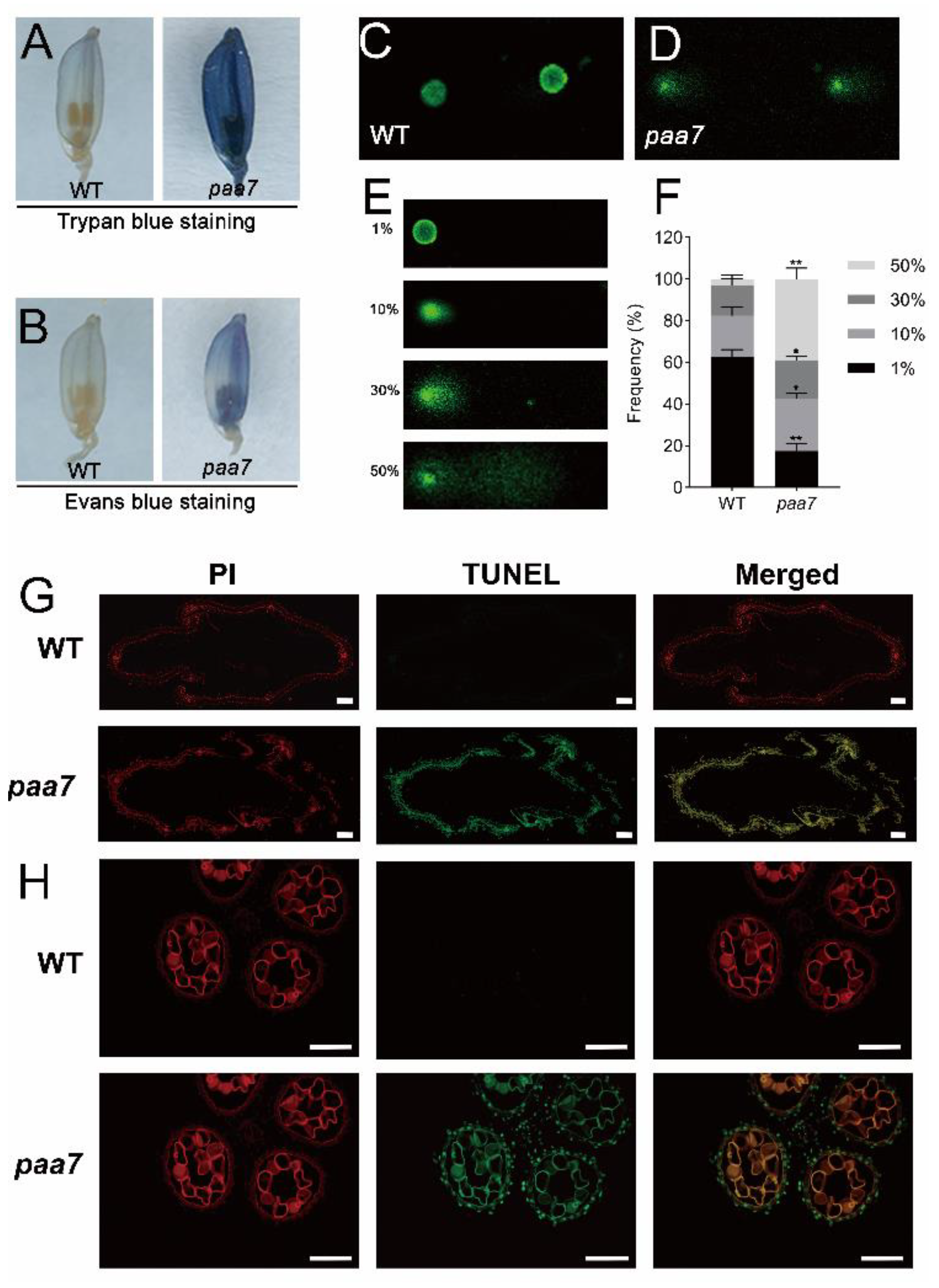
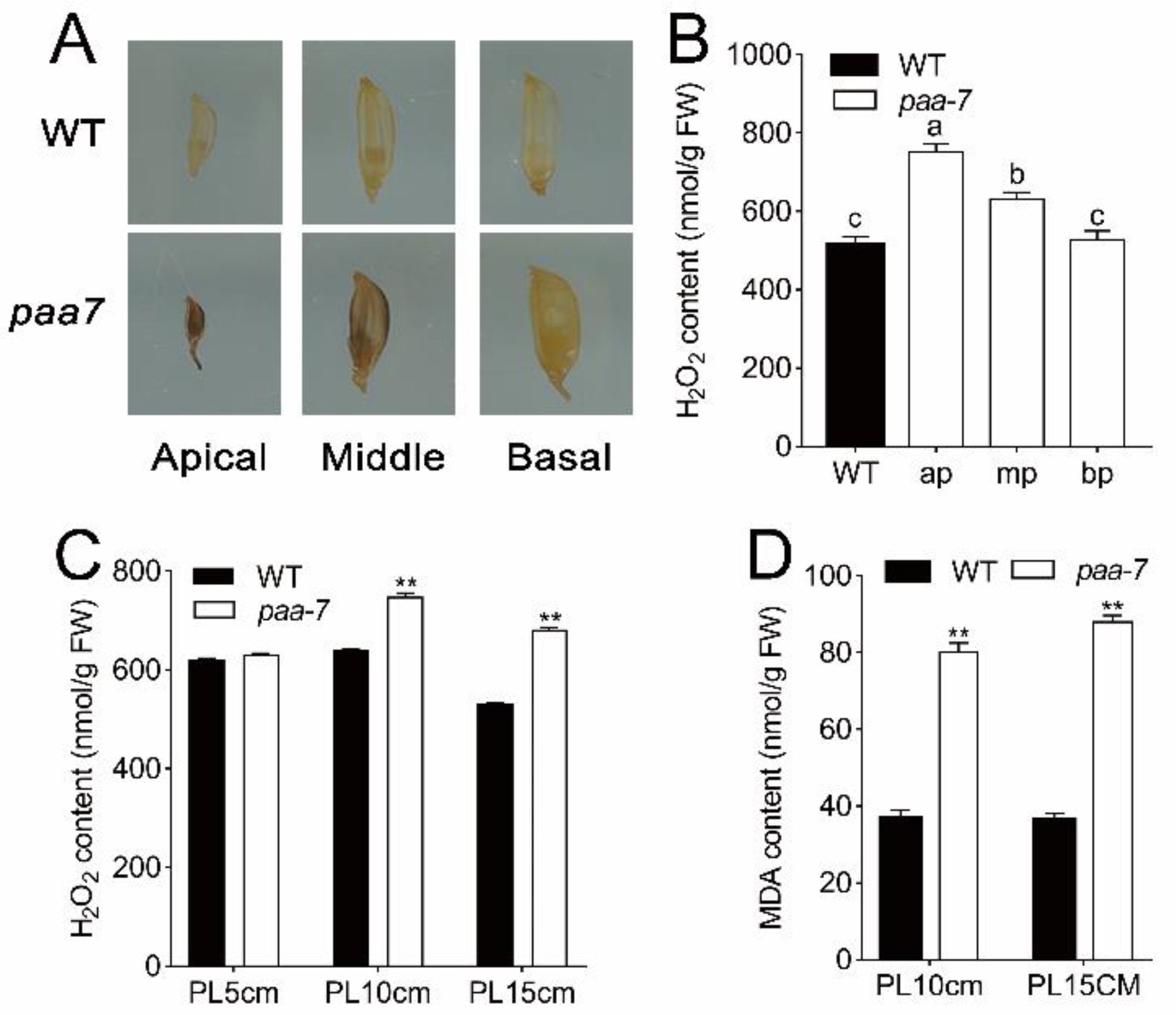
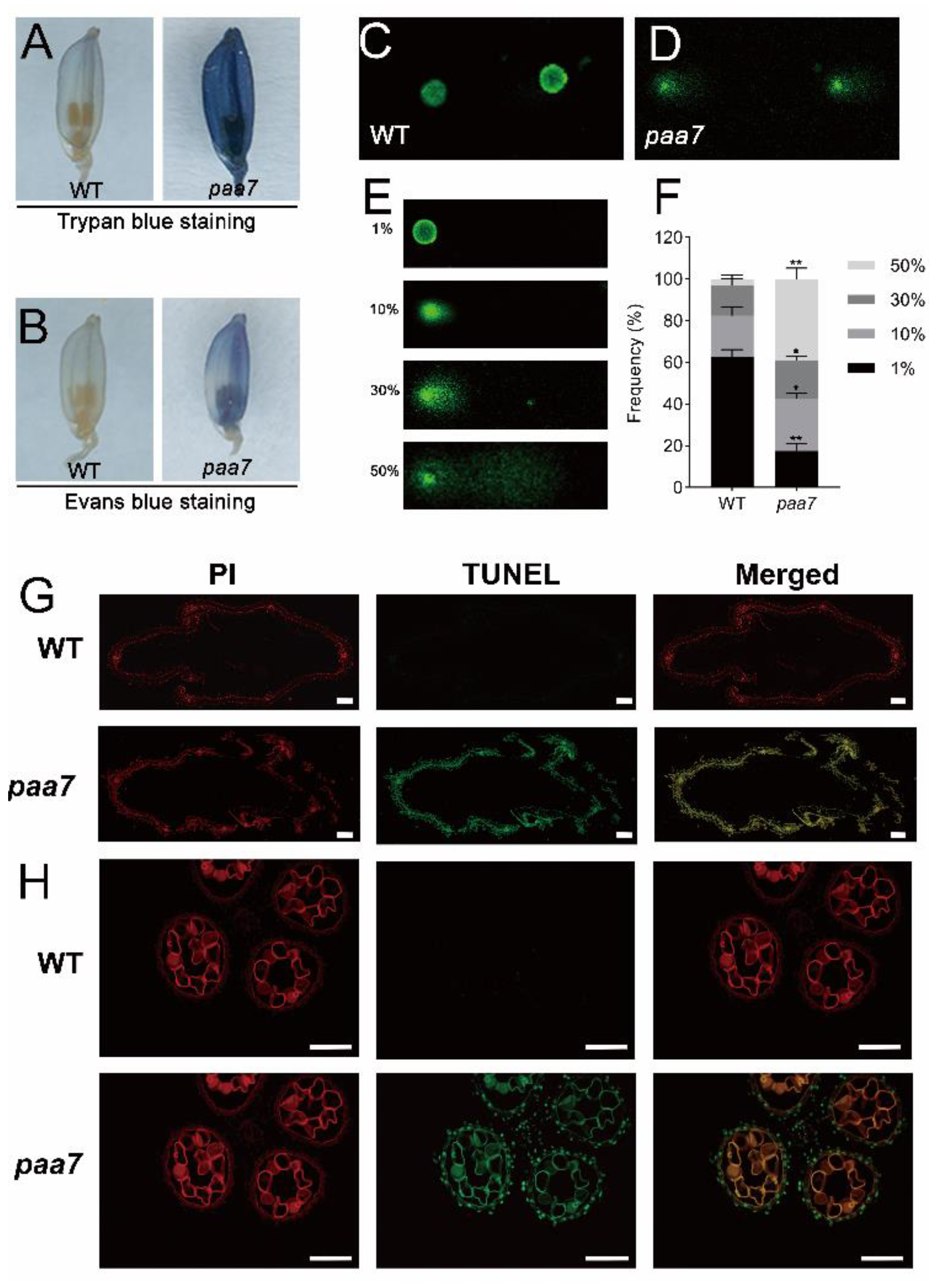
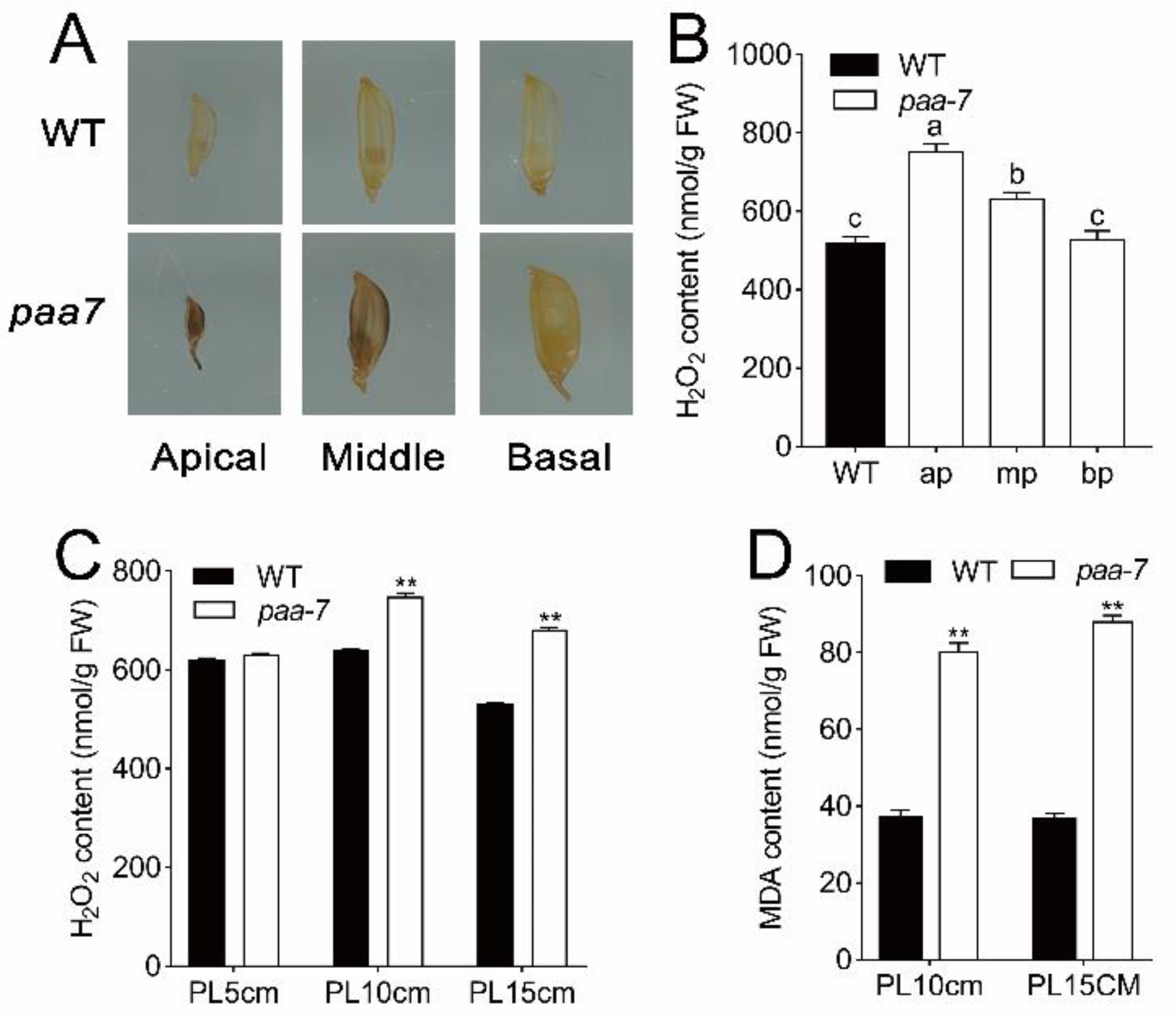
2.2. Enhanced Cell Death in paa7 Mutant
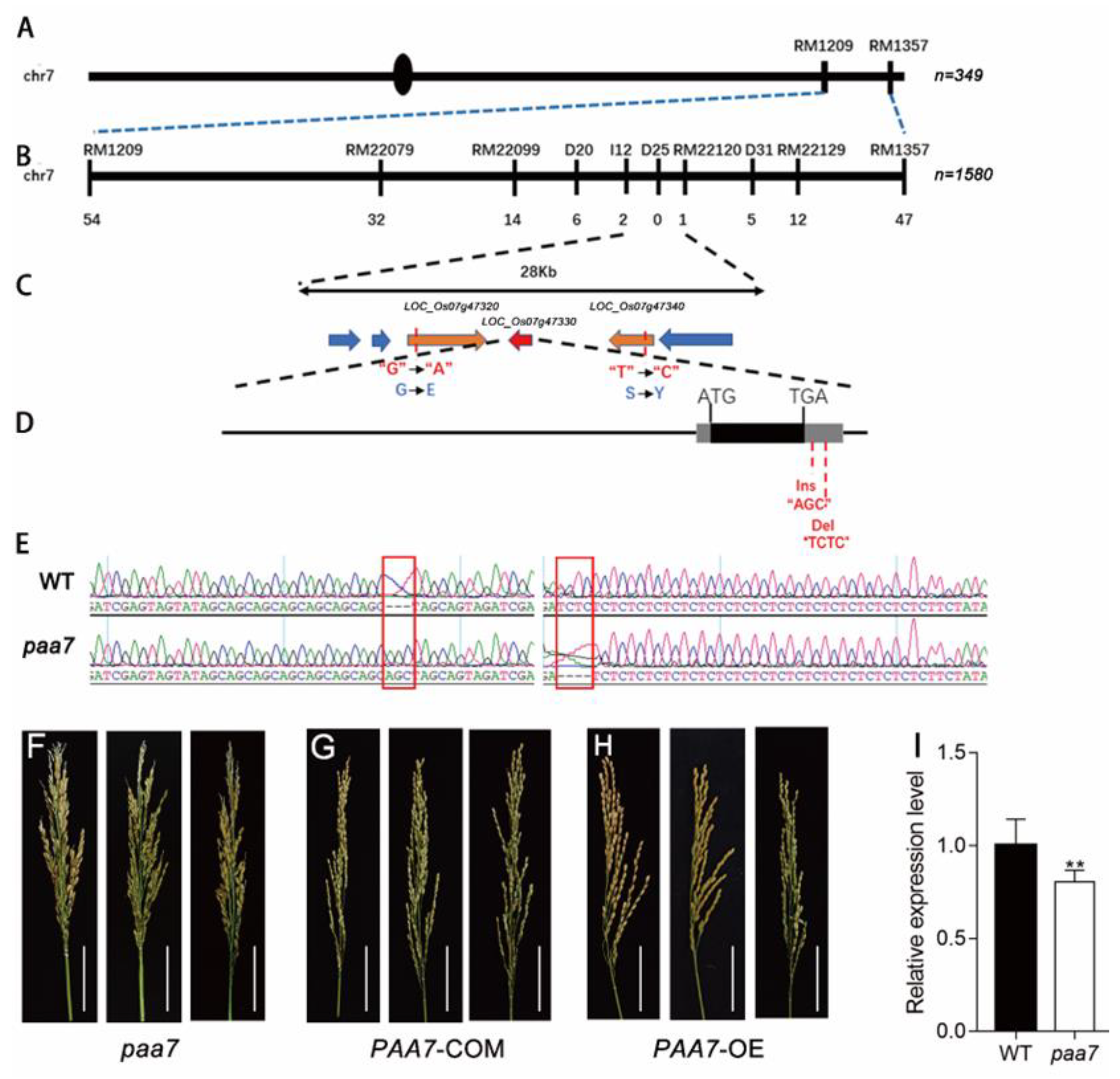
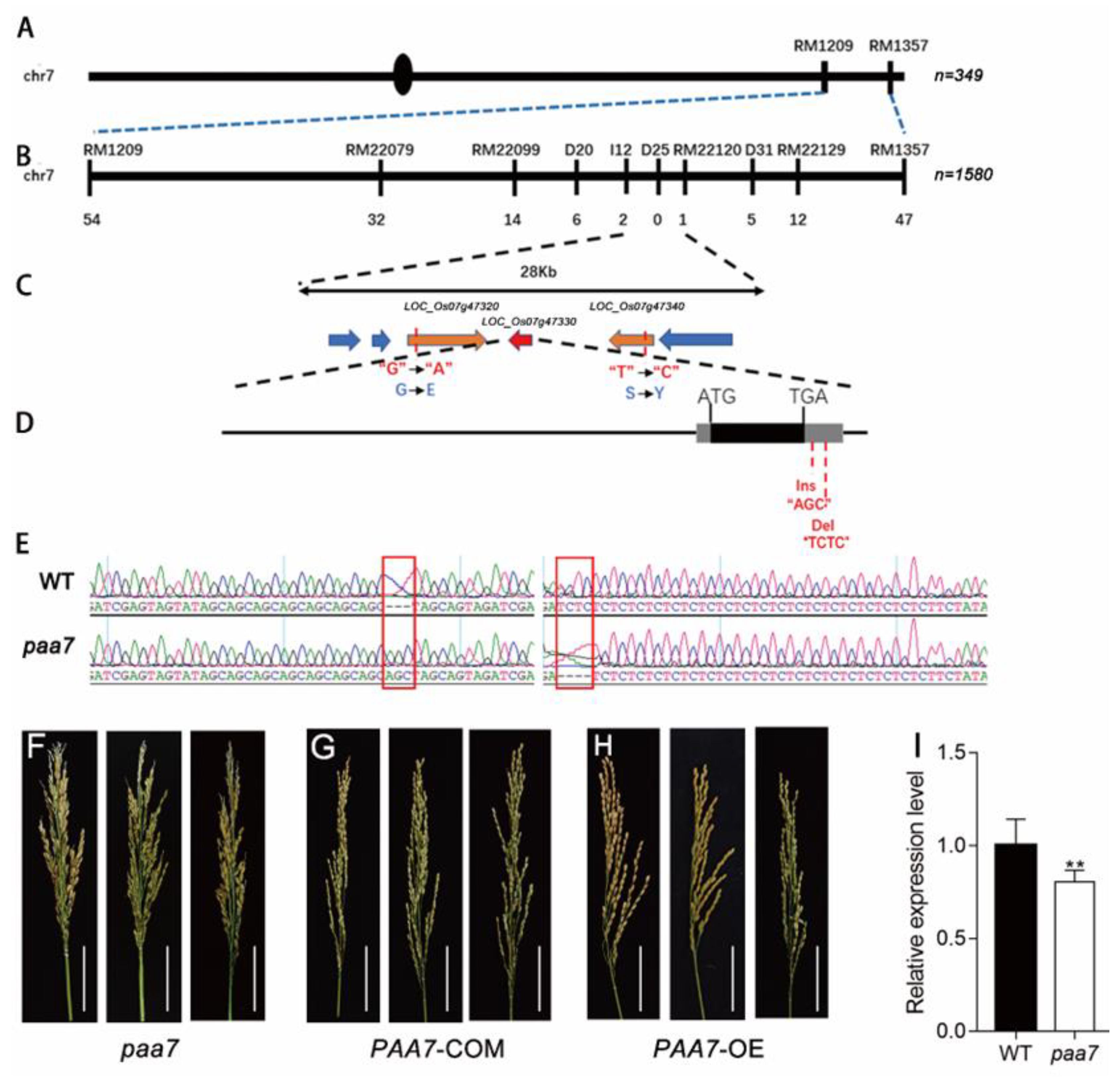
2.3. Mutations in 3′-UTR of PAA7 Responsible for Panicle Apical Abortion
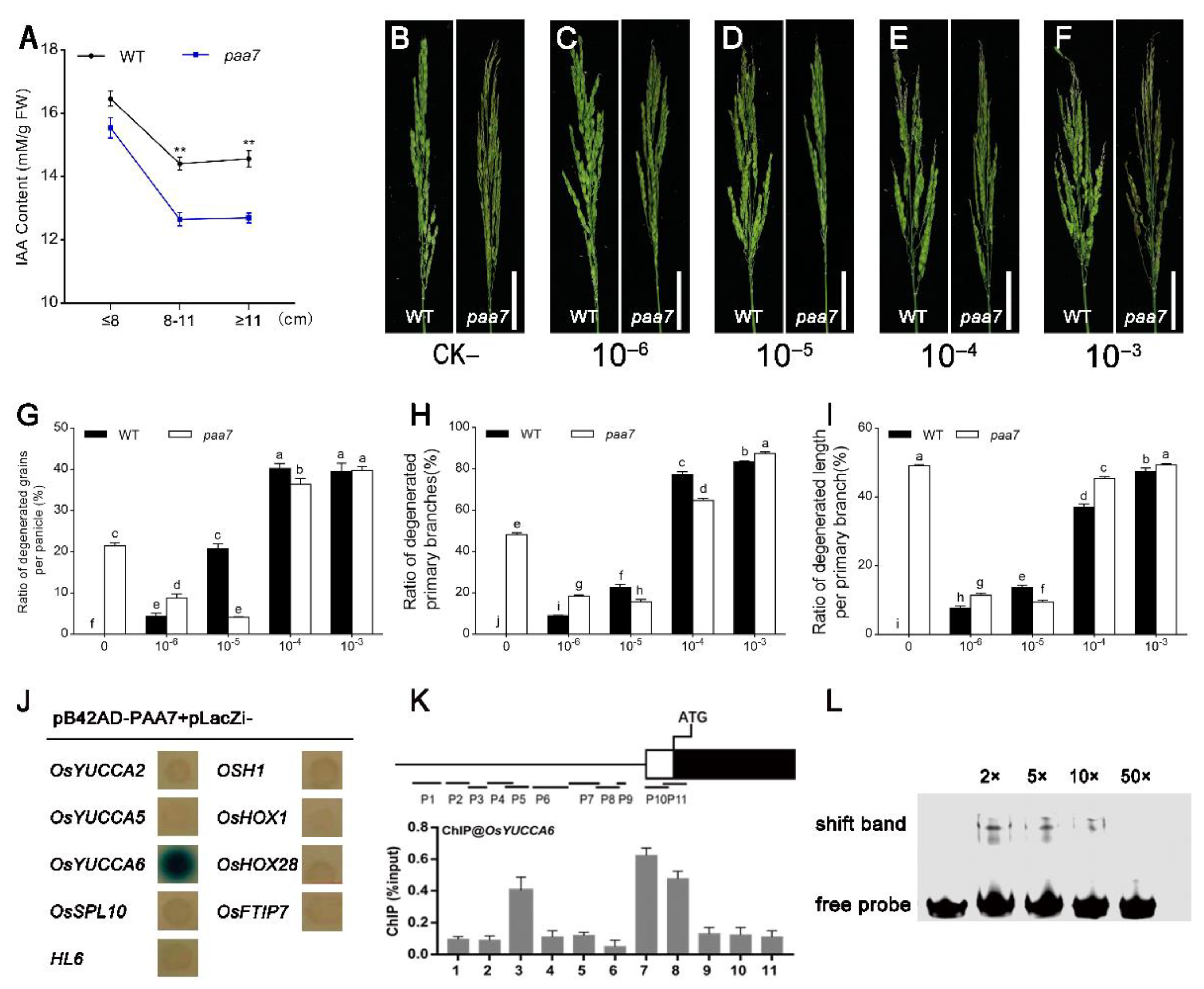
2.4. Feeding of Exogenous Auxin Alleviates Apical Spikelet Degeneration in paa7
2.5. PAA7 Activates the Expression of OsYUCCA6 to Regulate the Auxin Content in Young Panicle
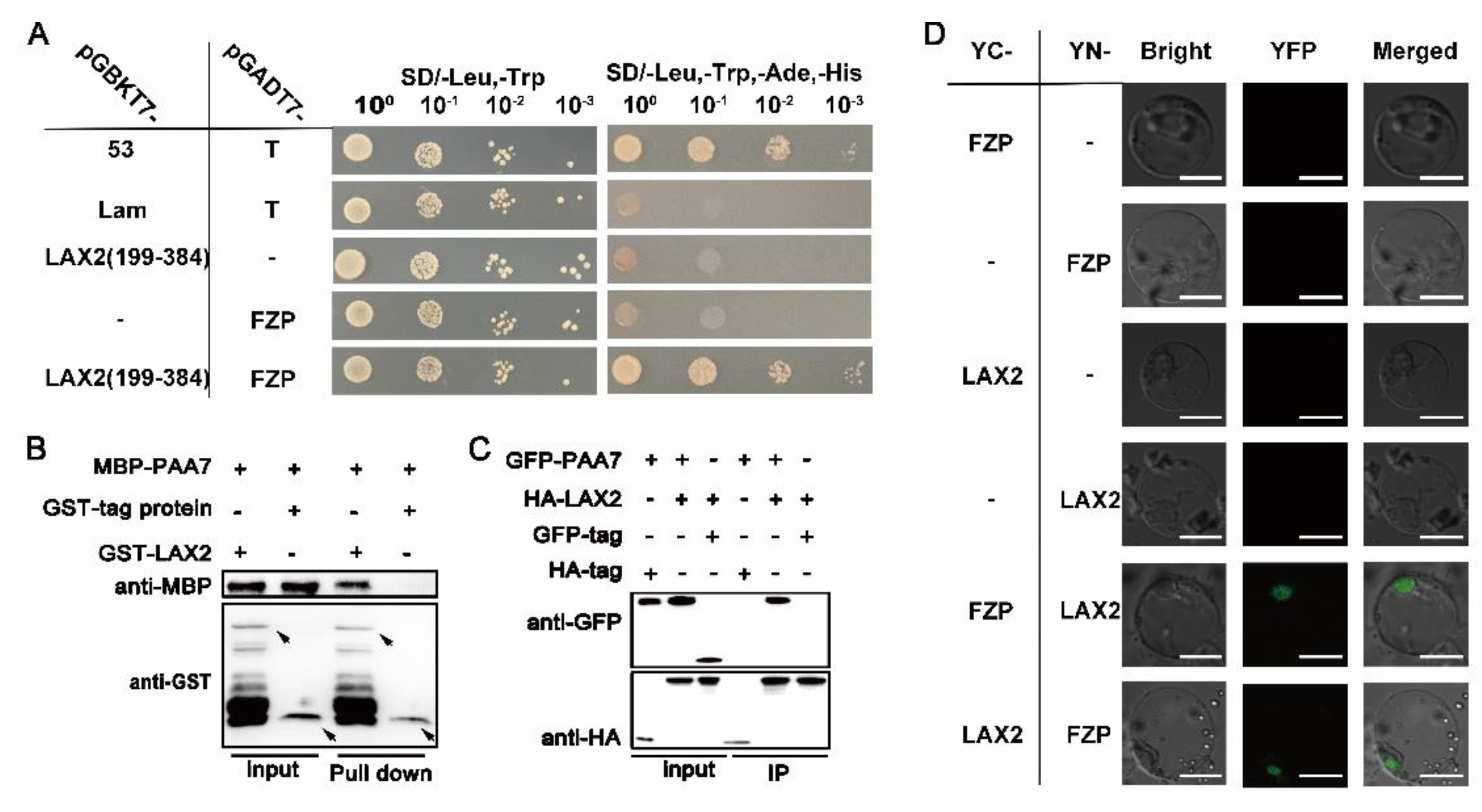
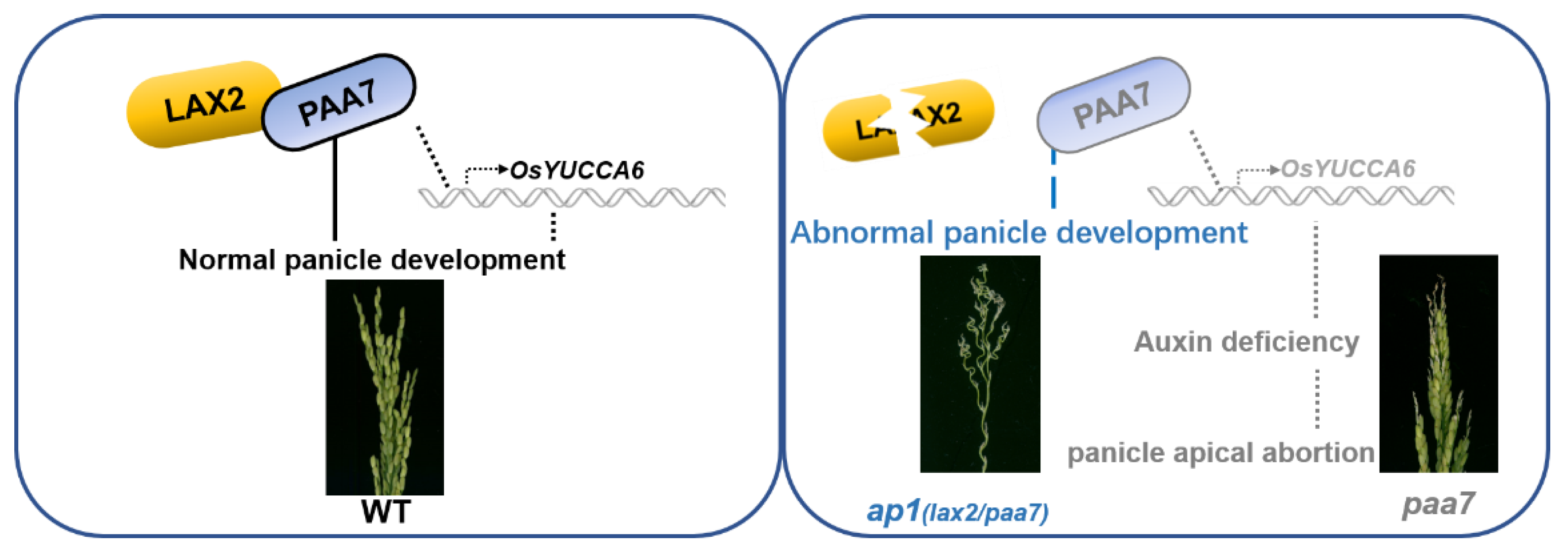
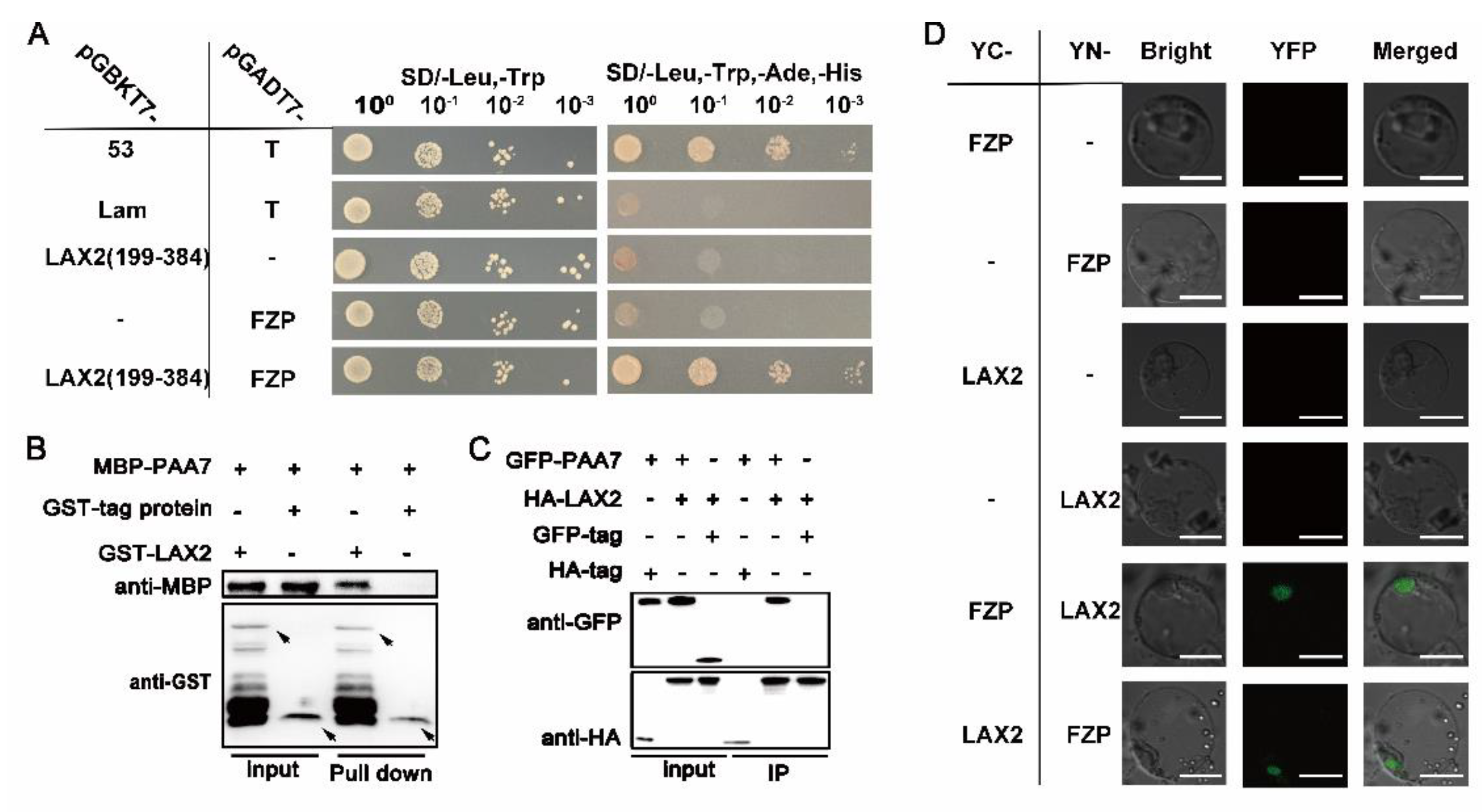
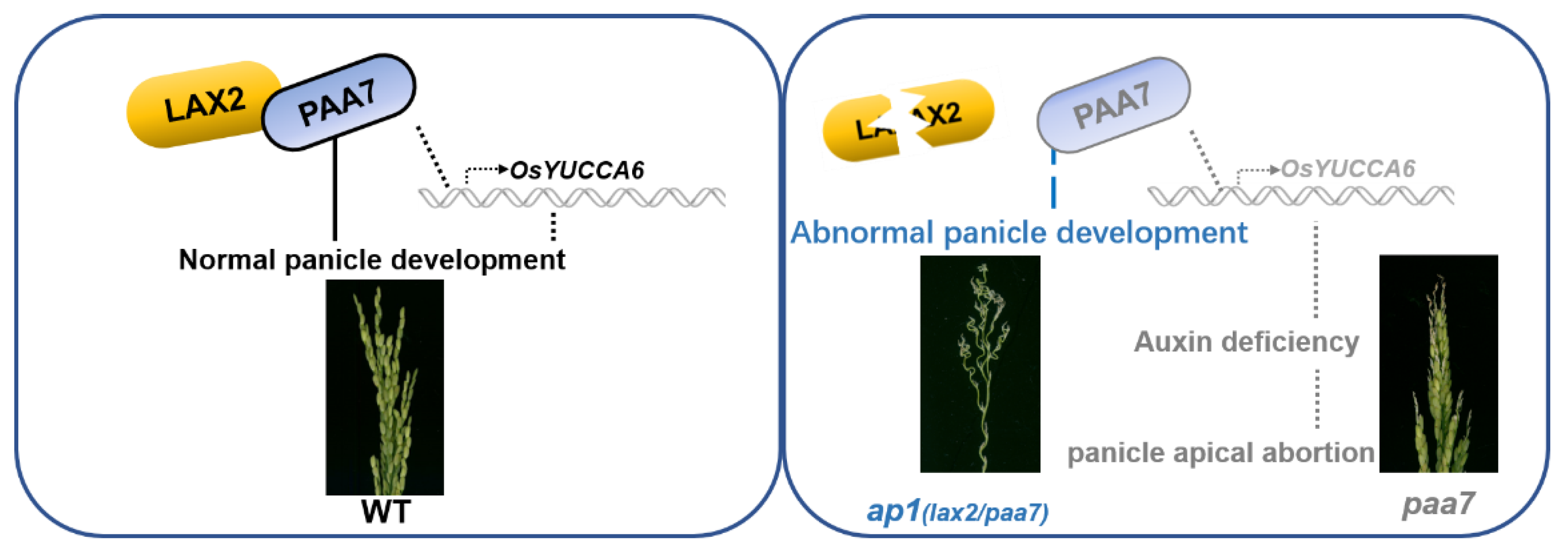
2.6. PAA7 Synergistically Works with LAX2 during Panicle Development
3. Discussion
3.1. paa7 Is a Novel Allele of fzp
3.2. Panicle Abortion in paa7 Possibly Be Caused by Down-Rugulation of OsYUCCA6
3.3. LAX2 and PAA7 Interact to Regulate Panicle Development in Rice
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Histological Analysis and TUNEL Assay
4.3. Trypan Blue and Evans Blue Staining
4.4. Comet Assay
4.5. Determination of H2O2 and Malondialdehyde (MDA) Contents
4.6. Positional Cloning of the PAA7 Gene
4.7. RNA Extraction and Quantitative Real-Time PCR Analysis
4.8. Vector Construction and Transformation
4.9. Yeast Two-Hybrid (Y2H) Assay
4.10. Pull-Down Assay
4.11. Bimolecular Fluorescence Complementation (BiFC) Assay
4.12. Coimmunoprecipitation (Co-IP) Assay
4.13. Auxin Content Determination
4.14. Yeast One-Hybrid Assay
4.15. Chromatin Immunoprecipitation-Quantitative PCR (ChIP-qPCR)
4.16. Electrophoresis Mobility Shift Assay (EMSA)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Tang, S.; Zhi, H.; Xing, L.; Zhang, H.; Tang, C.; Wang, E.; Zhao, M.; Jia, G.; Feng, B.; et al. The boron transporter SiBOR1 functions in cell wall integrity, cellular homeostasis, and panicle development in foxtail millet. Crop J. 2021, 10, 342–353. [Google Scholar] [CrossRef]
- Pei, Y.; Deng, Y.; Zhang, H.; Zhang, Z.; Liu, J.; Chen, Z.; Cai, D.; Li, K.; Du, Y.; Zang, J.; et al. Ear apical degeneration 1 regulates maize ear development by maintaining malate supply for apical inflorescence. Plant Cell 2022, 34, 2222–2241. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhu, X.; Wang, Q.; Zhang, J.; Chen, H.; Dong, G.; Zhu, L.; Zheng, H.; Xie, Q.; Nian, J.; et al. Rice tutou 1 encodes a suppressor of camp receptor-like protein that is important for actin organization and panicle development. Plant Physiol. 2015, 169, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.J.; Sun, Y.J.; Xu, H.S.; Yu, S.B. Identification of quantitative trait locus and epistatic interaction for degenerated spikelets on the top of panicle in rice. Plant Breed. 2011, 130, 177–184. [Google Scholar] [CrossRef]
- Cheng, Z.-J.; Mao, B.-G.; Gao, S.-W.; Zhang, L.; Wang, J.-L.; Lei, C.-L.; Zhang, X.; Wu, F.-Q.; Guo, X.-P.; Wan, J. Fine mapping of qPAA8, a gene controlling panicle apical development in rice. J. Integr. Plant Biol. 2011, 53, 710–718. [Google Scholar] [CrossRef]
- Xu, H.; Sun, Y.; Zhou, H.; Yu, S. Development and characterization of contiguous segment substitution lines with background of an elite restorer line. Acta Agron. Sin. 2007, 33, 979–986. [Google Scholar]
- Peng, Y.; Hou, F.; Bai, Q.; Xu, P.; Liao, Y.; Zhang, H.; Gu, C.; Deng, X.; Wu, T.; Chen, X.; et al. Rice calcineurin B-Like protein-interacting protein kinase 31 (OsCIPK31) is involved in the development of panicle apical spikelets. Front. Plant Sci. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Wang, Q.L.; Sun, A.-Z.; Chen, S.-T.; Chen, L.-S.; Guo, F.-Q. SPL6 represses signalling outputs of ER stress in control of panicle cell death in rice. Nat. Plants 2018, 4, 280–288. [Google Scholar] [CrossRef]
- Zafar, S.A.; Patil, S.B.; Uzair, M.; Fang, J.; Zhao, J.; Guo, T.; Yuan, S.; Uzair, M.; Luo, Q.; Shi, J.; et al. Degenerated panicle and partial sterility 1 (DPS 1) encodes a cystathionine β-synthase domain containing protein required for anther cuticle and panicle development in rice. New Phytol. 2020, 225, 356–375. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xiong, M.; Huang, M.; Li, Z.; Wang, Z.; Zhu, H.; Chen, R.; Lu, L.; Cheng, Q.; Wang, Y.; et al. Panicle apical abortion 3 controls panicle development and seed size in rice. Rice 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Ali, A.; Wu, T.; Zhang, H.; Xu, P.; Zafar, S.A.; Liao, Y.; Chen, X.; Zhou, H.; Liu, Y.; Wang, W.; et al. A putative Subtilisin-like serine protease 1 (SUBSrP1) regulates anther cuticle biosynthesis and panicle development in rice. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- Zhu, Z.-C.; Luo, S.; Lei, B.; Li, X.-Y.; Cheng, Z.-J. Locus tutou 2 determines the panicle apical abortion phenotype of rice (Oryza sativa L.) in tutou 2 mutant. J. Integr. Agric. 2022, 21, 621–630. [Google Scholar] [CrossRef]
- Heng, Y.; Wu, C.; Long, Y.; Luo, S.; Ma, J.; Chen, J.; Liu, J.; Zhang, H.; Ren, Y.; Wang, M.; et al. Osalmt 7 maintains panicle size and grain yield in rice by mediating malate transport. Plant Cell 2018, 30, 889–906. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. In Annual Review of Plant Biology; Merchant, S., Briggs, W.R., Ort, D., Eds.; Annual Reviews: Palo Alto, CA, USA, 2010; Volume 61, pp. 421–442. [Google Scholar]
- Li, G.; Zhang, H.; Li, J.; Zhang, Z.; Li, Z. Genetic control of panicle architecture in rice. Crop J. 2021, 9, 590–597. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, X.; Ma, X.; Xu, B.; Zhao, Y.; Ma, Z.; Li, G.; Khan, N.U.; Pan, Y.; Liang, Y.; et al. GNP6, a novel allele of moc 1, regulates panicle and tiller development in rice. Crop J. 2020, 9, 57–67. [Google Scholar] [CrossRef]
- Komatsu, M.; Chujo, A.; Nagato, Y.; Shimamoto, K.; Kyozuka, J. Frizzy panicle is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development 2003, 130, 3841–3850. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Huang, Y.; Hu, Y.; Liu, H.; Zhang, B.; Smaczniak, C.; Hu, G.; Han, Z.; Xing, Y. Duplication of an upstream silencer of FZP increases grain yield in rice. Nat. Plants 2017, 3, 885–893. [Google Scholar] [CrossRef]
- Song, S.; Wang, G.; Hu, Y.; Liu, H.; Bai, X.; Qin, R.; Xing, Y. Osmft 1 increases spikelets per panicle and delays heading date in rice by suppressing EHD1, FZP and sepallata-like genes. J. Exp. Bot. 2018, 69, 4283–4293. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, S.; He, Y.; Yan, L.; Wang, R.; Zhao, Y. Synergistic roles of LAX1 and FZP in the development of rice sterile lemma. Crop J. 2020, 8, 16–25. [Google Scholar] [CrossRef]
- Komatsu, K.; Maekawa, M.; Ujiie, S.; Satake, Y.; Furutani, I.; Okamoto, H.; Shimamoto, K.; Kyozuka, J. LAX and SPA: Major regulators of shoot branching in rice. Proc. Natl. Acad. Sci. USA 2003, 100, 11765–11770. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Xie, D.; Tang, Z.; Shi, D.; Yang, W. Pinoid regulates floral organ development by modulating auxin transport and interacts with mads16 in rice. Plant Biotechnol. J. 2020, 18, 1778–1795. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, J.; Tang, Z.; Sun, X.; Zhang, H.; Yu, J.; Yao, G.; Li, G.; Guo, H.; Li, J.; et al. Gnp4/LAX2, a rawul protein, interferes with the Osiaa3–Osarf25 interaction to regulate grain length via the auxin signaling pathway in rice. J. Exp. Bot. 2018, 69, 4723–4737. [Google Scholar] [CrossRef]
- Tabuchi, H.; Zhang, Y.; Hattori, S.; Omae, M.; Shimizu-Sato, S.; Oikawa, T.; Qian, Q.; Nishimura, M.; Kitano, H.; Xie, H.; et al. Lax panicle2 of rice encodes a novel nuclear protein and regulates the formation of axillary meristems. Plant Cell 2011, 23, 3276–3287. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Sunohara, H.; Nagato, Y. Developmental course of inflorescence and spikelet in rice. Breed. Sci. 2004, 54, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Speit, G.; Rothfuss, A. The Comet Assay: A Sensitive Genotoxicity Test for the Detection of DNA Damage and Repair; Humana Press: Totowa, NJ, USA, 2006. [Google Scholar]
- Dai, D.; Chen, J.; Du, C.; Liang, M.; Wu, M.; Mou, T.; Zhang, H.; Ma, L. A 2-MB chromosome inversion interrupted transcription of lax2-4 and generated pleiotropic phenotypes in rice. J. Plant Growth Regul. 2021, 41, 2328–2337. [Google Scholar] [CrossRef]
- Ren, D.; Hu, J.; Xu, Q.; Cui, Y.; Zhang, Y.; Zhou, T.; Rao, Y.; Xue, D.; Zeng, D.; Zhang, G.; et al. FZP determines grain size and sterile lemma fate in rice. J. Exp. Bot. 2018, 69, 4853–4866. [Google Scholar] [CrossRef]
- Fujishiro, Y.; Agata, A.; Ota, S.; Ishihara, R.; Takeda, Y.; Kunishima, T.; Ikeda, M.; Kyozuka, J.; Hobo, T.; Kitano, H. Comprehensive panicle phenotyping reveals that qSrn7/FZP influences higher-order branching. Sci. Rep. 2018, 8, 12511. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, S.; Fu, Y.; Sun, H.; Ma, X.; Tan, L.; Liu, F.; Sun, X.; Sun, H.; Gu, P.; et al. Variation in the regulatory region of FZP causes increases in secondary inflorescence branching and grain yield in rice domestication. Plant J. 2018, 96, 716–733. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.-H.; Hoque, M.S.; Dennis, E.S.; Upadhyaya, N.M. Ds tagging of branched floretless 1 (BFL1) that mediates the transition from spikelet to floret meristem in rice (Oryza sativa L). BMC Plant Biol. 2003, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Mignone, F.; Gissi, C.; Liuni, S.; Pesole, G. Untranslated regions of mRNAs. Genome Biol. 2002, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin biosynthesis by the yucca genes in rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-Y.; Li, J.-J.; Yao, G.-X.; Zhang, H.-L.; Dou, H.-J.; Shi, H.-L.; Sun, X.-M.; Li, Z.-C. Fine mapping and cloning of the grain number per-panicle gene (Gnp4) on chromosome 4 in rice (Oryza sativa L.). Agric. Sci. China 2011, 10, 1825–1833. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Y.; Ma, L.; Sang, X.; Ling, Y.; Wang, Y.; Yu, P.; Zhuang, H.; Huang, J.; Wang, N.; et al. Lateral floret 1 induced the three-florets spikelet in rice. Proc. Natl. Acad. Sci. USA 2017, 114, 9984–9989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Zhang, S.; Qiu, Z.; Zhao, J.; Nie, W.; Lin, H.; Zhu, Z.; Zeng, D.; Qian, Q.; Zhu, L. Fructokinase-like protein 1 interacts with TRXz to regulate chloroplast development in rice. J. Integr. Plant Biol. 2018, 60, 94–111. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wang, C.; Fu, Y.; Liu, Q.; Jiao, X.; Wang, K. Expanding the range of crispr/cas9 genome editing in rice. Mol. Plant 2016, 9, 943–945. [Google Scholar] [CrossRef] [Green Version]
- Toki, S.; Hara, N.; Ono, K.; Onodera, H.; Tagiri, A.; Oka, S.; Tanaka, H. Early infection of scutellum tissue with agrobacterium allows high-speed transformation of rice. Plant J. 2006, 47, 969–976. [Google Scholar] [CrossRef]
- Teng, N.; Wang, J.; Chen, T.; Wu, X.; Wang, Y.; Lin, J. Elevated CO2 induces physiological, biochemical and structural changes in leaves of arabidopsis thaliana. New Phytol. 2006, 172, 92–103. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Wang, L.; Liu, L.; Li, L.; Sun, L.; Rao, Q.; Zhang, J.; Huang, S. JMJ704 positively regulates rice defense response against xanthomonas oryzae pv. oryzae infection via reducing H3K4me2/3 associated with negative disease resistance regulators. BMC Plant Biol. 2015, 15, 286. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hou, Y.; Qiu, J.; Wang, H.; Wang, S.; Tang, L.; Tong, X.; Zhang, J. Abscisic acid promotes jasmonic acid biosynthesis via a ‘SAPK10-bZIP72-AOC’ pathway to synergistically inhibit seed germination in rice (Oryza sativa). New Phytol. 2020, 228, 1336–1353. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, D.; Zhang, H.; He, L.; Chen, J.; Du, C.; Liang, M.; Zhang, M.; Wang, H.; Ma, L. Panicle Apical Abortion 7 Regulates Panicle Development in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2022, 23, 9487. https://doi.org/10.3390/ijms23169487
Dai D, Zhang H, He L, Chen J, Du C, Liang M, Zhang M, Wang H, Ma L. Panicle Apical Abortion 7 Regulates Panicle Development in Rice (Oryza sativa L.). International Journal of Molecular Sciences. 2022; 23(16):9487. https://doi.org/10.3390/ijms23169487
Chicago/Turabian StyleDai, Dongqing, Huali Zhang, Lei He, Junyu Chen, Chengxing Du, Minmin Liang, Meng Zhang, Huimei Wang, and Liangyong Ma. 2022. "Panicle Apical Abortion 7 Regulates Panicle Development in Rice (Oryza sativa L.)" International Journal of Molecular Sciences 23, no. 16: 9487. https://doi.org/10.3390/ijms23169487
APA StyleDai, D., Zhang, H., He, L., Chen, J., Du, C., Liang, M., Zhang, M., Wang, H., & Ma, L. (2022). Panicle Apical Abortion 7 Regulates Panicle Development in Rice (Oryza sativa L.). International Journal of Molecular Sciences, 23(16), 9487. https://doi.org/10.3390/ijms23169487