
Selective Expansion of NKG2C+ Adaptive NK Cells Using K562 Cells Expressing HLA-E

 , ,
, ,
Abstract
:1. Introduction
2. Results
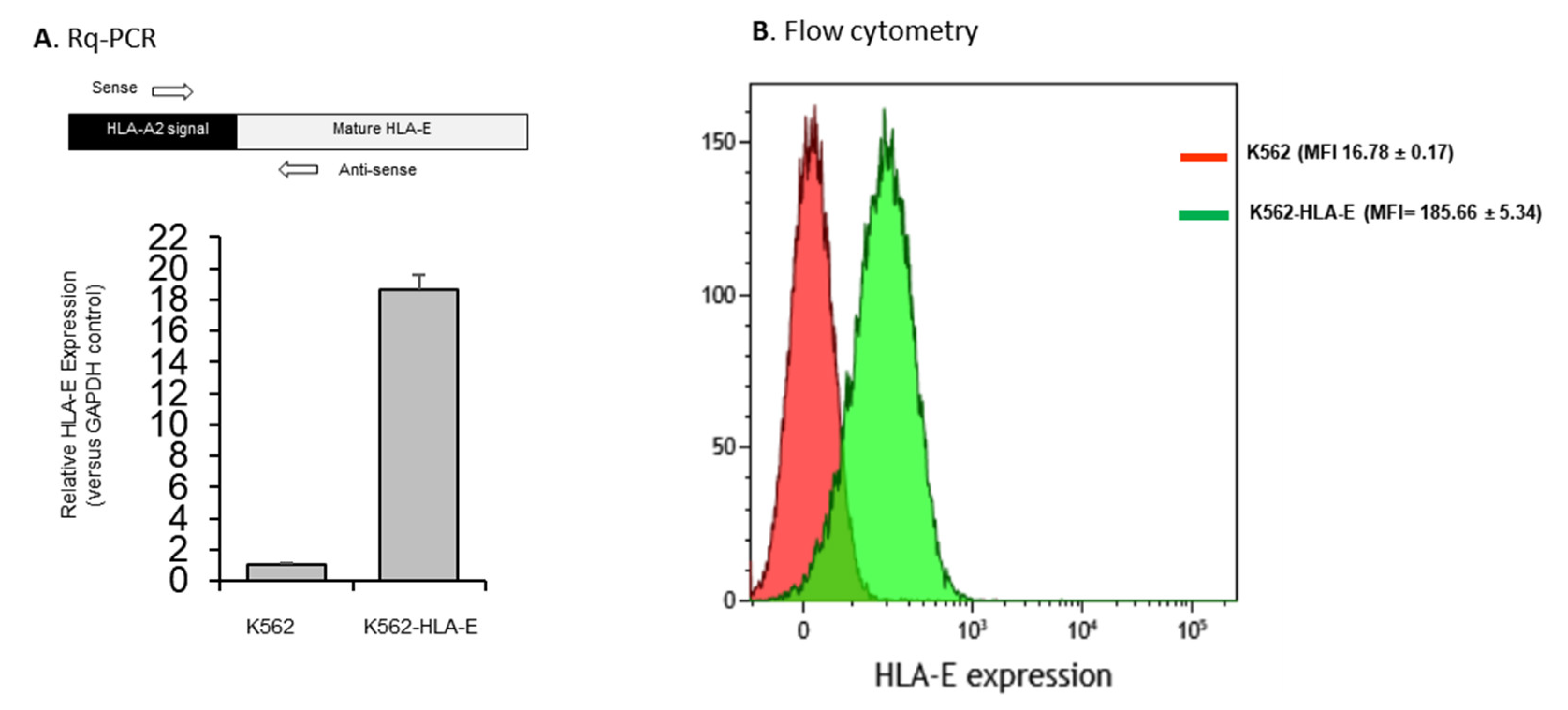
2.1. Generation of Genetically Engineered K562 Cells Expressing HLA-E
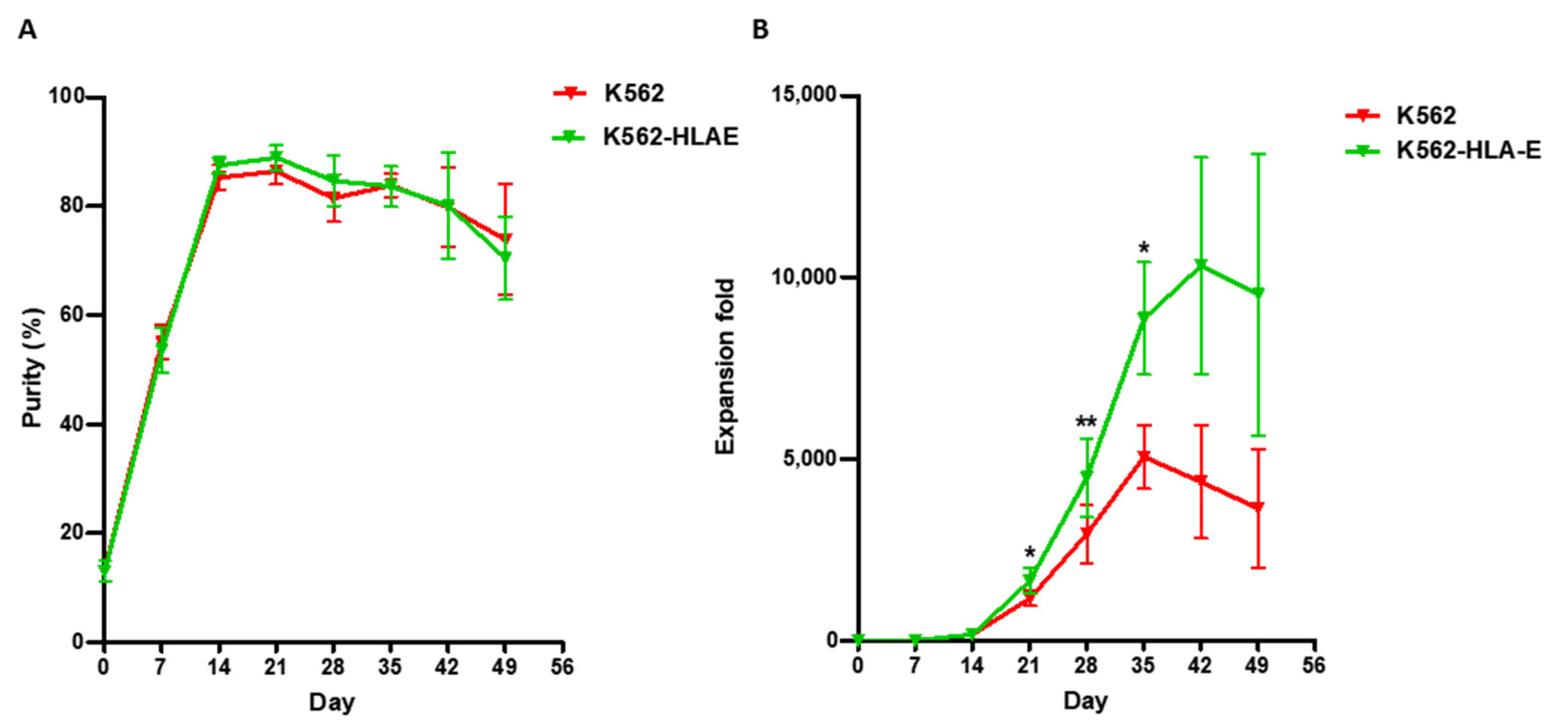
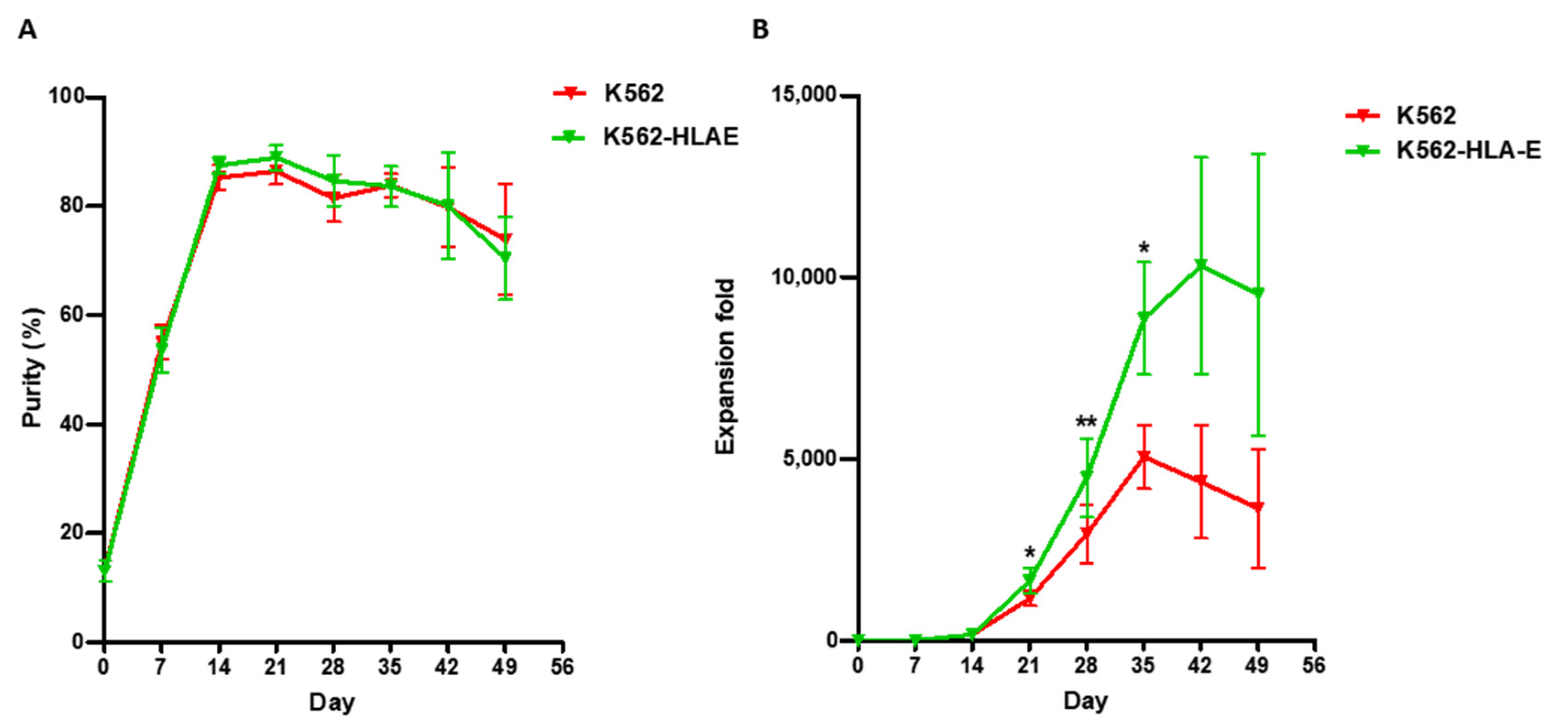
2.2. K562-HLA-E Feeder Cells Enhanced the Expansion of NK Cells
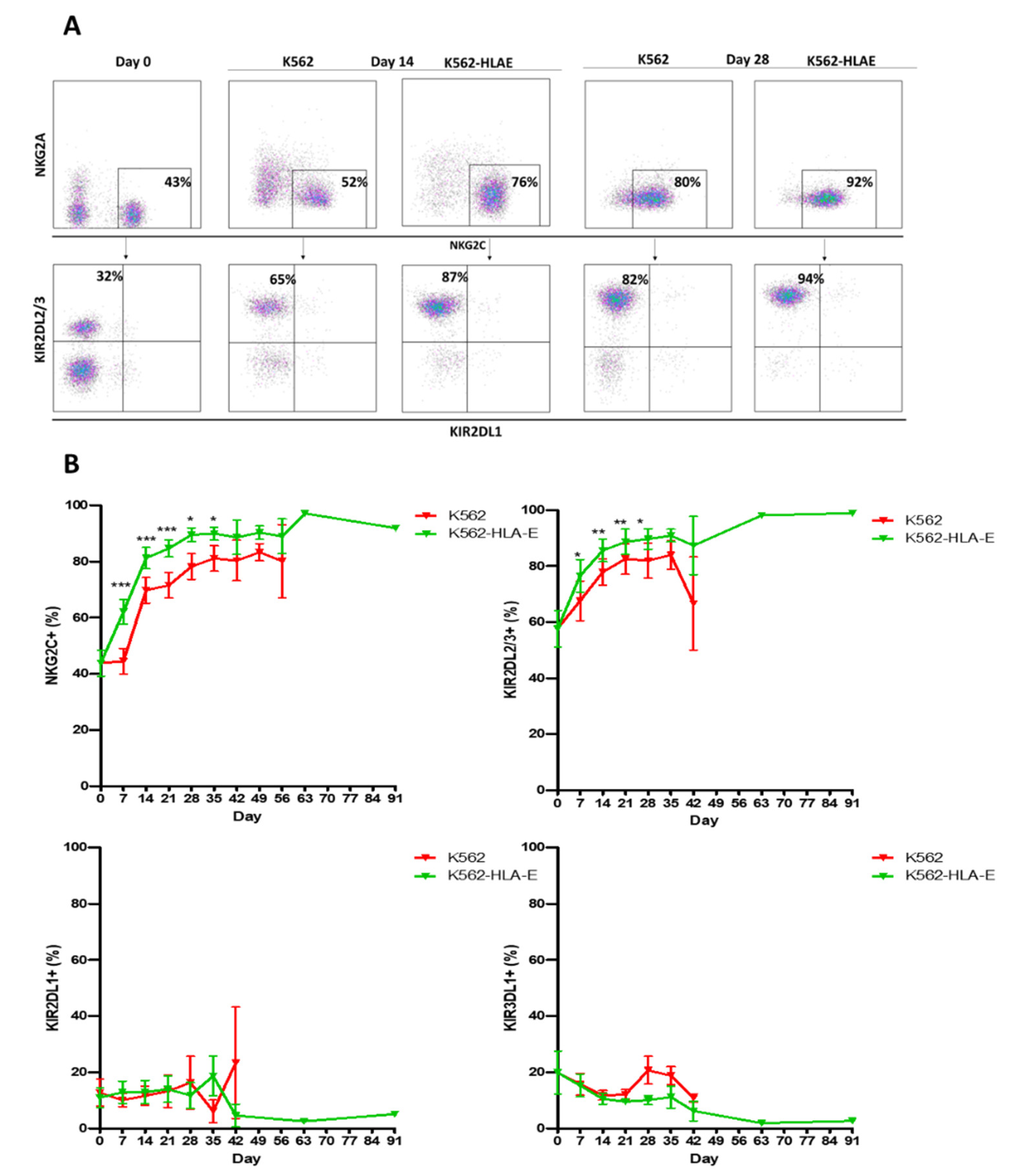
2.3. Selective Expansion of Single KIR+ Cells Co-Cultured with PBMCs on K562-HLA-E Feeder Cells
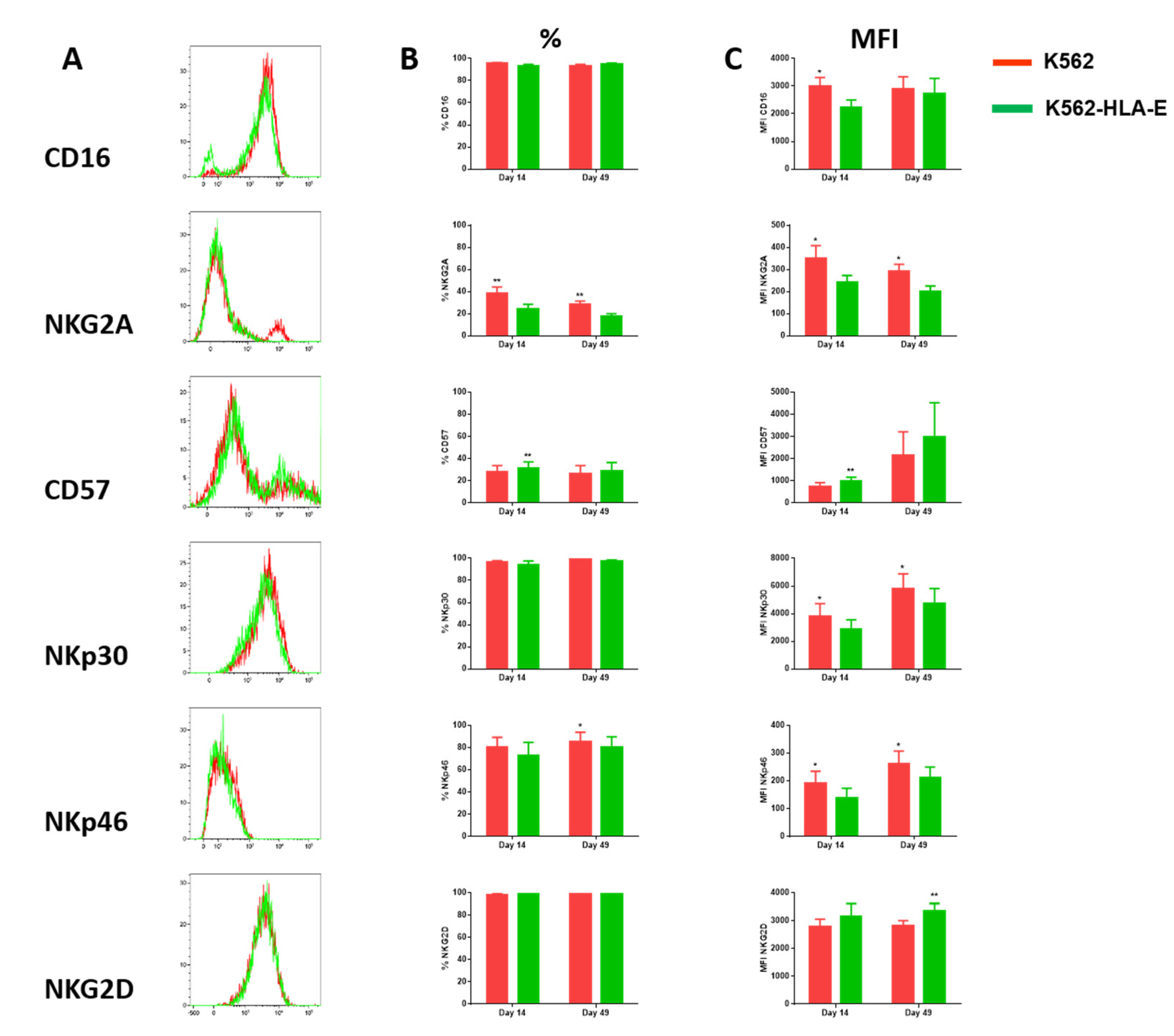
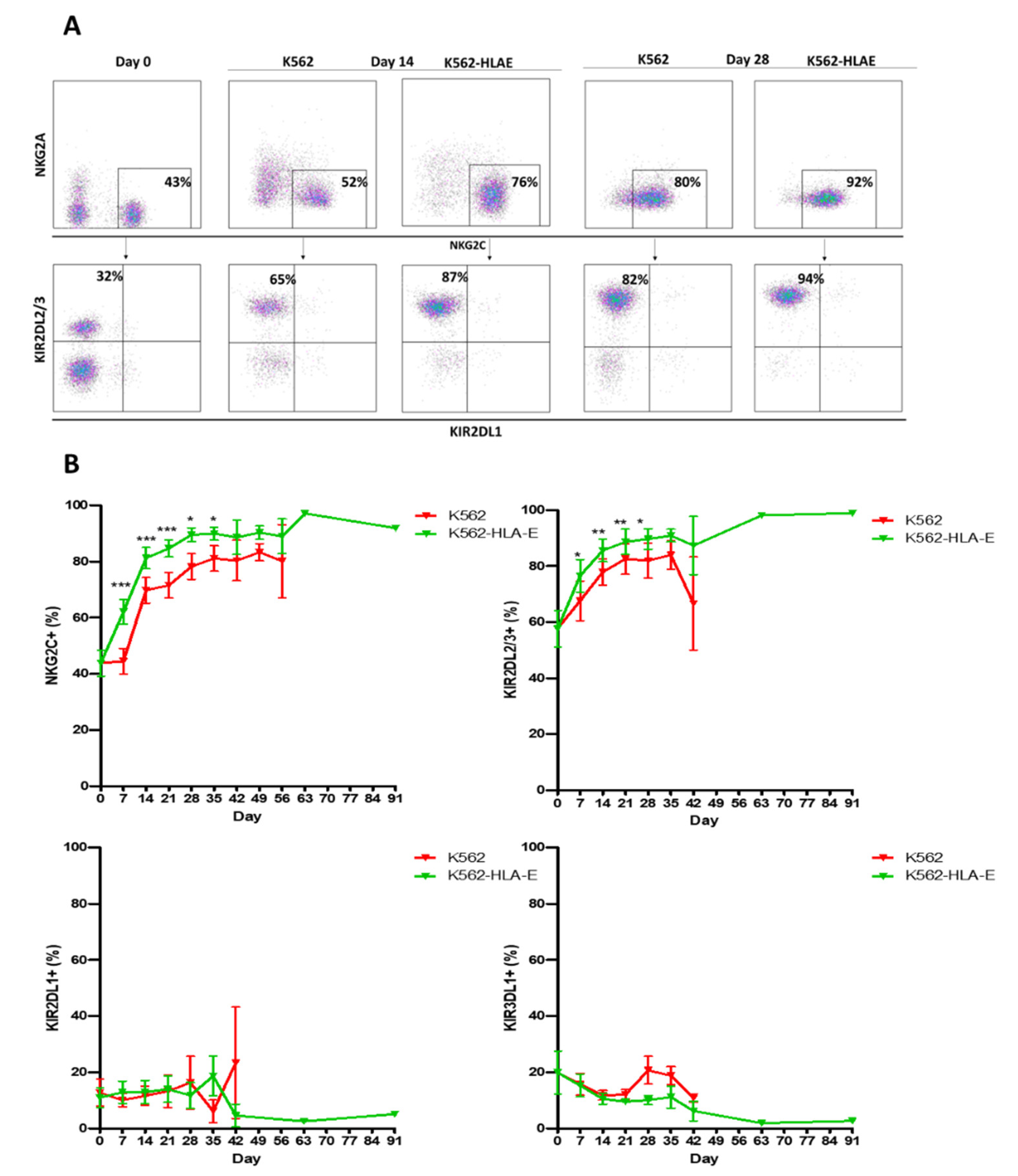
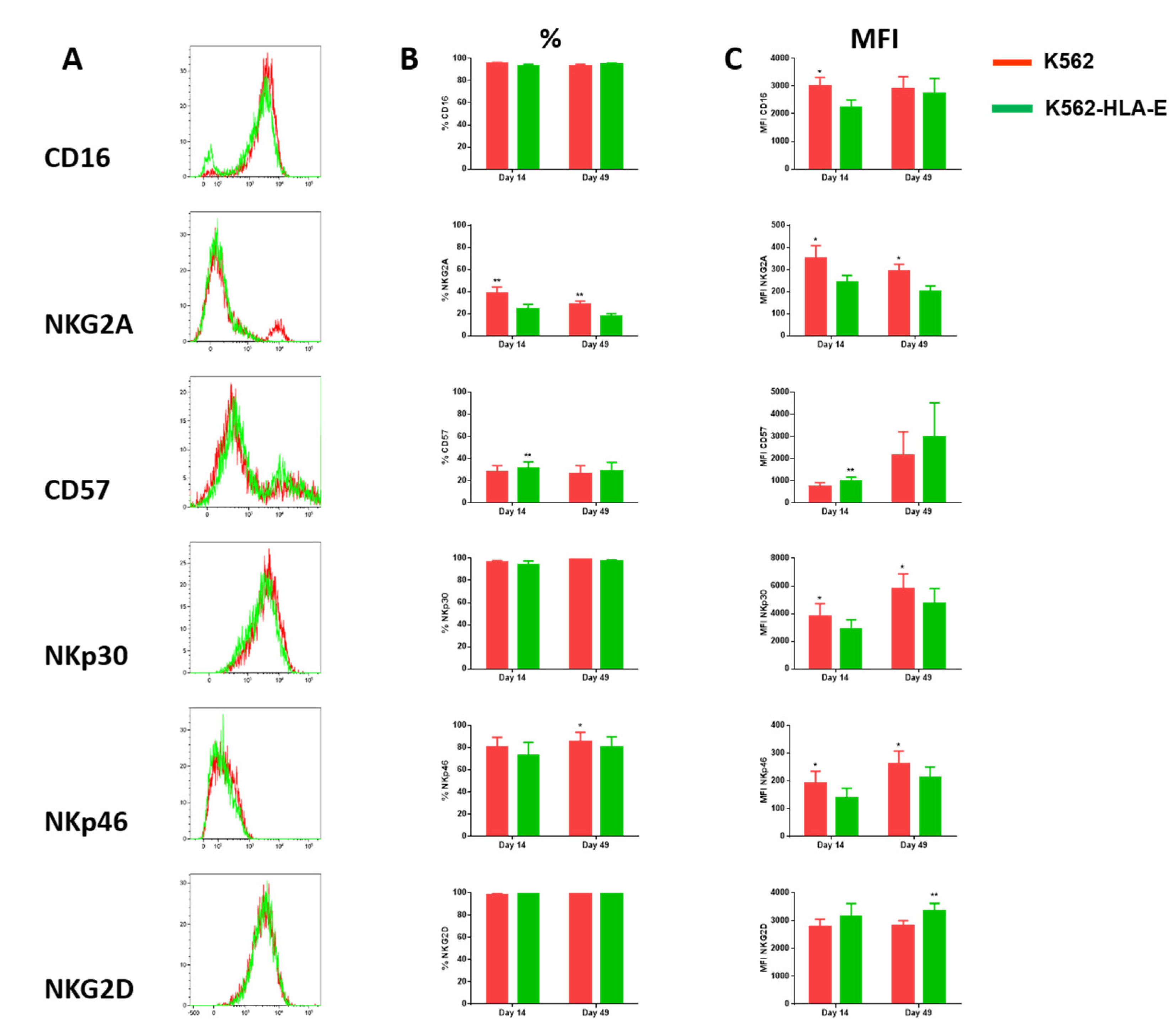
2.4. Phenotypic Characteristics of eNK Cells Using K562 and K562-HLA-E as Feeder Cells
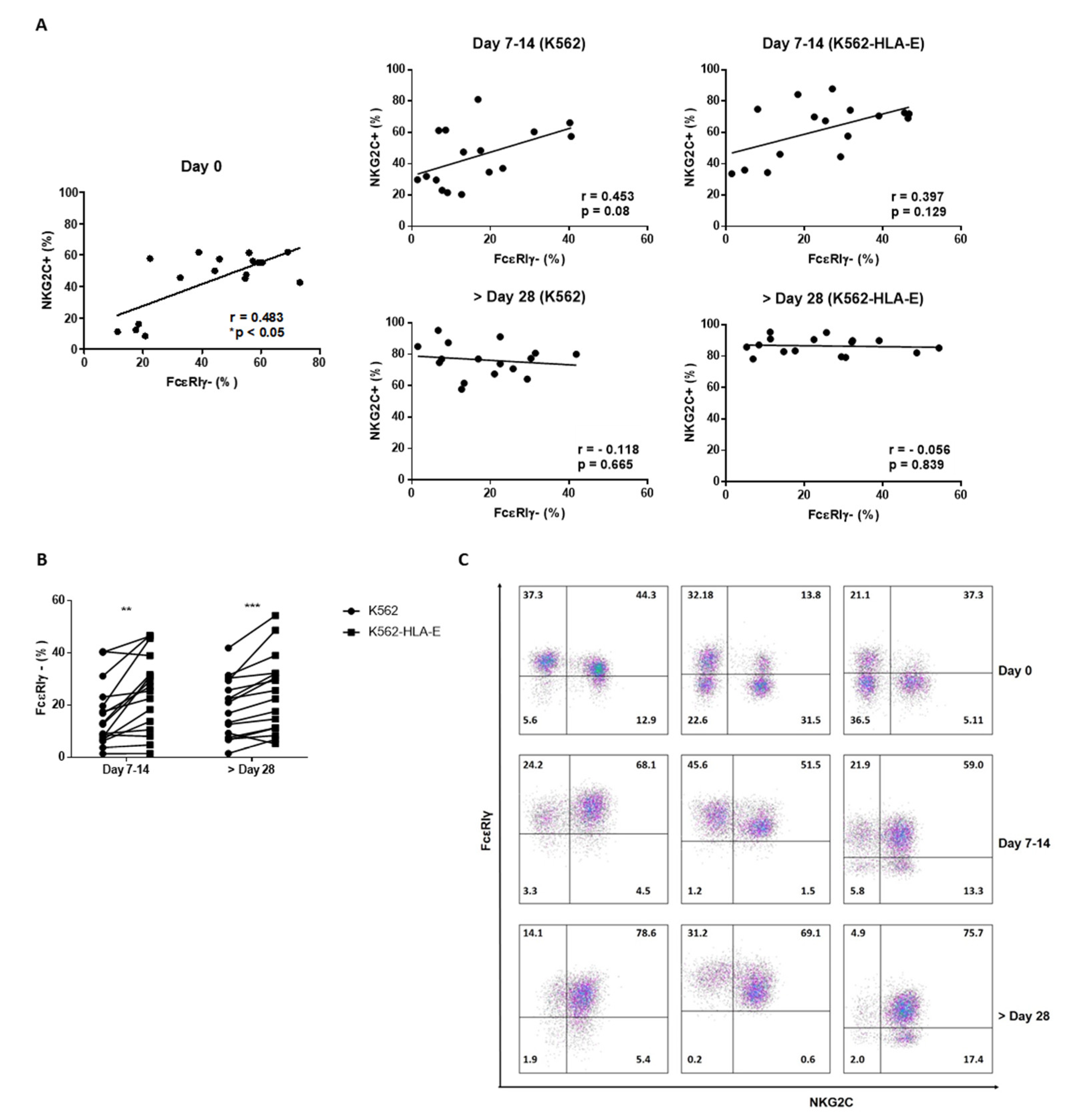
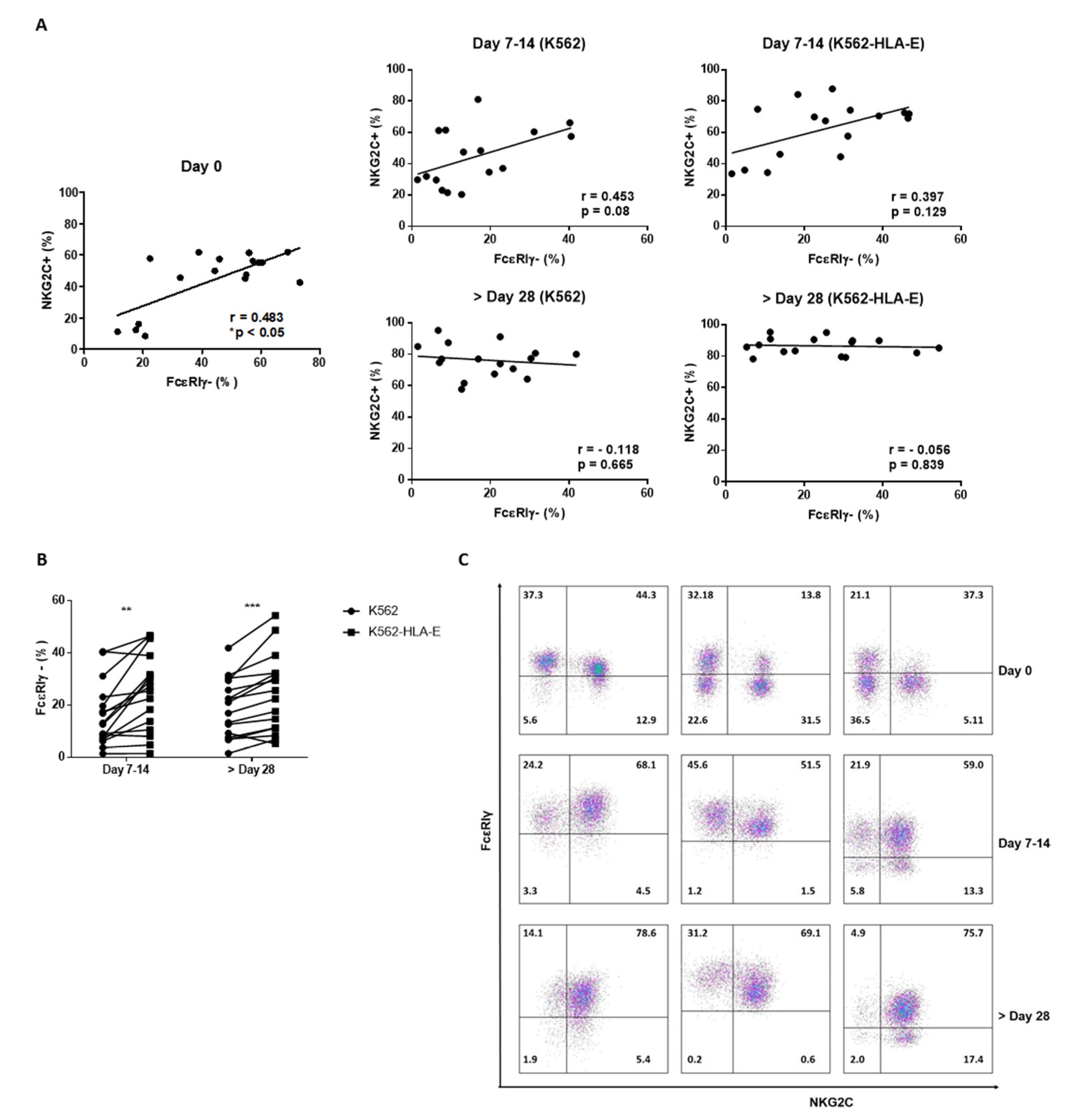
2.5. Expression of FcεRIγ and NKG2C in NK Cells during Expansion
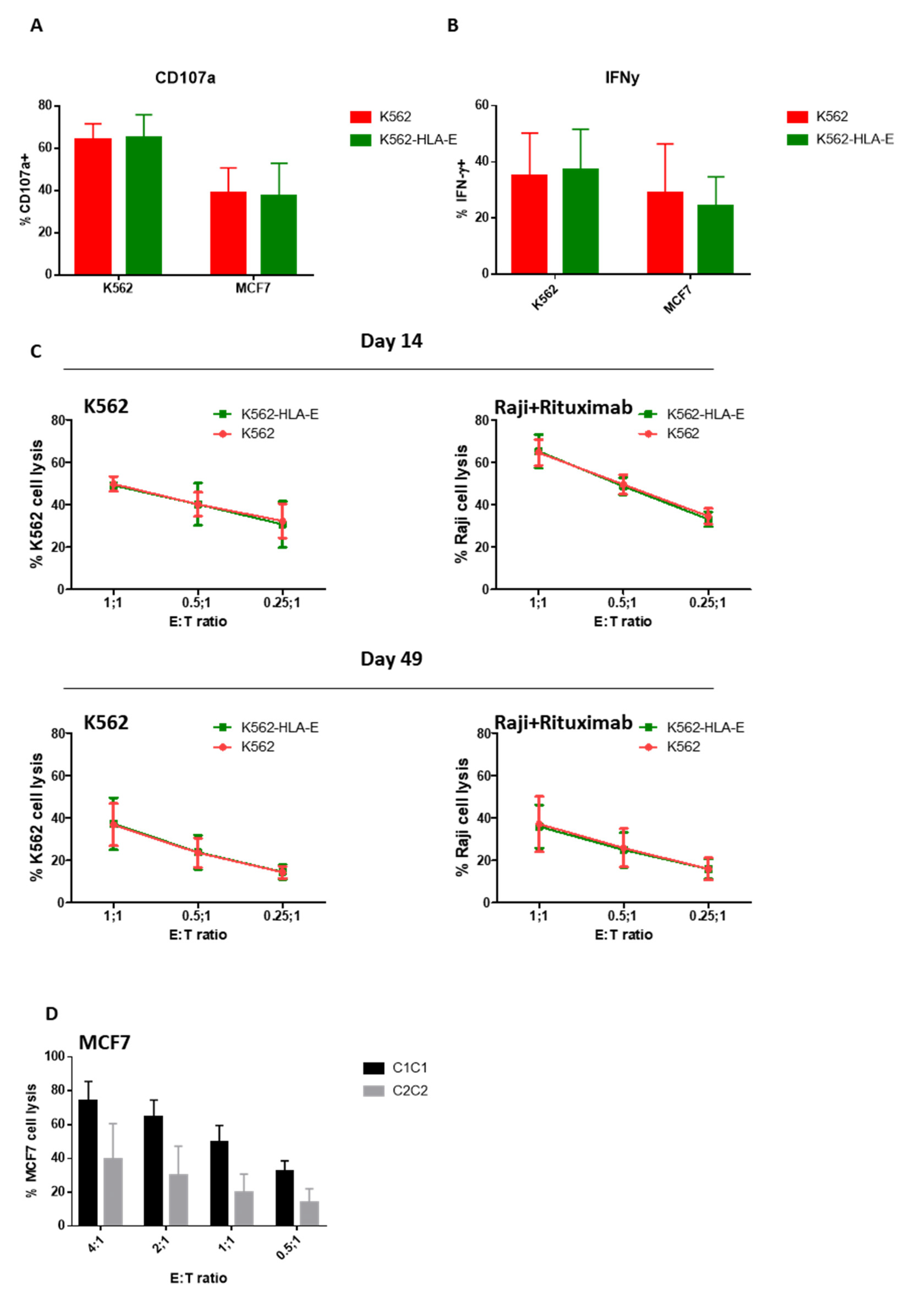
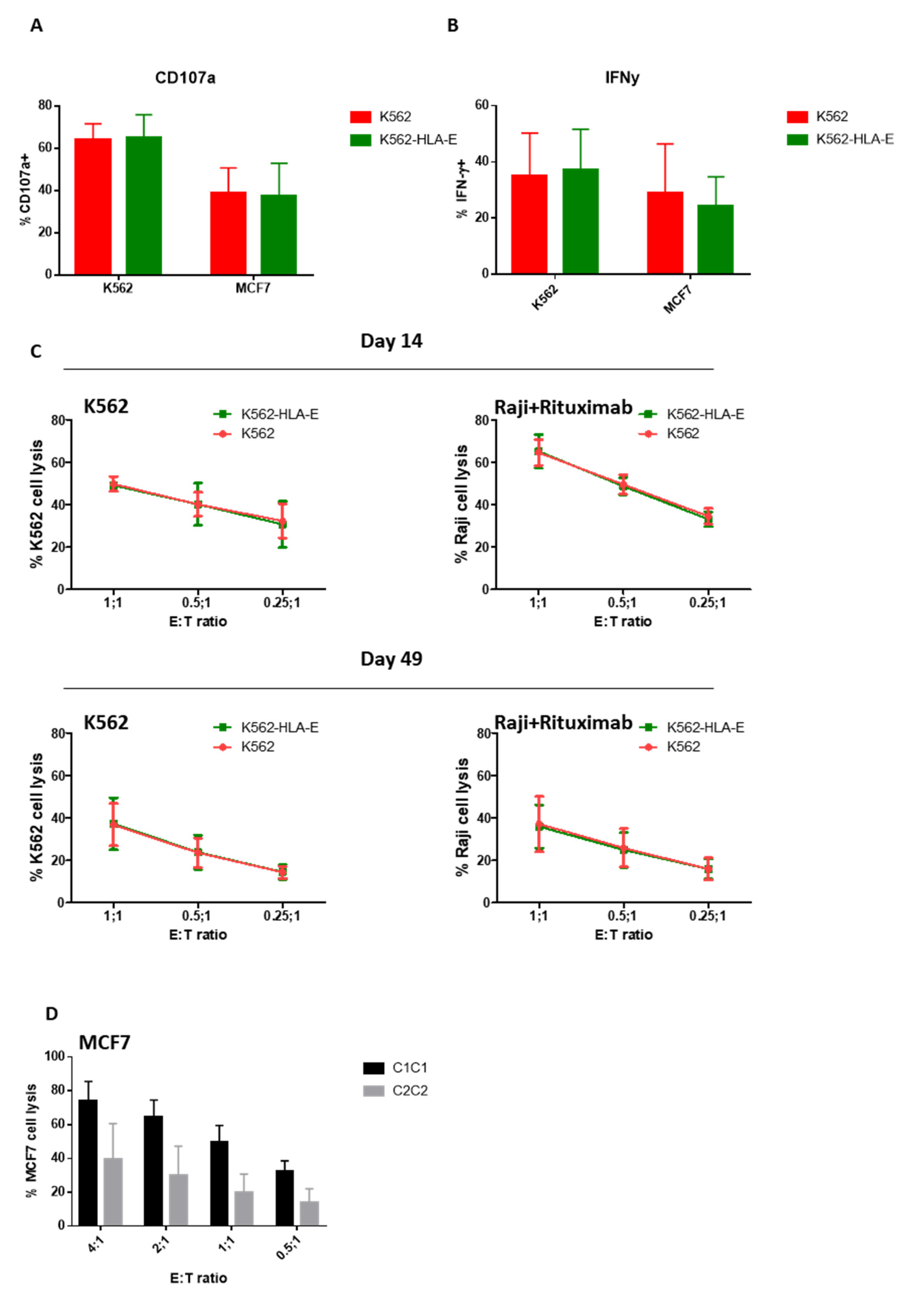
2.6. Function of In Vitro Generated eNK Cells Using K562 and K562-HLA-E Feeder Cells
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Cells and Culture
4.3. Generation of Genetically Engineered K562 Expressing HLA-E
4.4. Real-Time Quantitative Polymerase Chain Reaction (Rq-PCR) Detection of HLA-E mRNA
4.5. Cytokines and Antibodies
4.6. Ex vivo NK Cell Expansion Using K562 and K562-HLA-E Feeder Cells
4.7. Surface and Intracellular Staining Using Flow Cytometry
4.8. CD107a Degranulation
4.9. Cytotoxicity and ADCC Assays
4.10. HLA-C Genotyping and HCMV Serology
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lanier, L.L. Up on the tightrope: Natural killer cell activation and inhibition. Nat. Immunol. 2008, 9, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, W.M.; Kim, S.; French, A.R. The dynamic life of natural killer cells. Annu. Rev. Immunol. 2004, 22, 405–429. [Google Scholar] [CrossRef] [PubMed]
- Gumá, M.; Angulo, A.; Vilches, C.; Gómez-Lozano, N.; Malats, N.; López-Botet, M. Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 2004, 104, 3664–3671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Botet, M.; Muntasell, A.; Vilches, C. The CD94/NKG2C+ NK-cell subset on the edge of innate and adaptive immunity to human cytomegalovirus infection. Semin. Immunol. 2014, 26, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Cerwenka, A.; Lanier, L.L. Natural killer cell memory in infection, inflammation and cancer. Nat. Rev. Immunol. 2016, 16, 112–123. [Google Scholar] [CrossRef]
- Béziat, V.; Dalgard, O.; Asselah, T.; Halfon, P.; Bedossa, P.; Boudifa, A.; Hervier, B.; Theodorou, I.; Martinot, M.; Debré, P.; et al. CMV drives clonal expansion of NKG2C+ NK cells expressing self-specific KIRs in chronic hepatitis patients. Eur. J. Immunol. 2012, 42, 447–457. [Google Scholar] [CrossRef]
- Liu, L.L.; Pfefferle, A.; Yi Sheng, V.O.; Björklund, A.T.; Béziat, V.; Goodridge, J.P.; Malmberg, K.J. Harnessing adaptive natural killer cells in cancer immunotherapy. Mol. Oncol. 2015, 9, 1904–1917. [Google Scholar] [CrossRef] [Green Version]
- Luetke-Eversloh, M.; Killig, M.; Romagnani, C. Signatures of human NK cell development and terminal differentiation. Front. Immunol. 2013, 4, 499. [Google Scholar] [CrossRef] [Green Version]
- Béziat, V.; Liu, L.L.; Malmberg, J.A.; Ivarsson, M.A.; Sohlberg, E.; Björklund, A.T.; Retière, C.; Sverremark-Ekström, E.; Traherne, J.; Ljungman, P.; et al. NK cell responses to cytomegalovirus infection lead to stable imprints in the human KIR repertoire and involve activating KIRs. Blood 2013, 121, 2678–2688. [Google Scholar] [CrossRef]
- Della Chiesa, M.; Falco, M.; Bertaina, A.; Muccio, L.; Alicata, C.; Frassoni, F.; Locatelli, F.; Moretta, L.; Moretta, A. Human cytomegalovirus infection promotes rapid maturation of NK cells expressing activating killer Ig-like receptor in patients transplanted with NKG2C-/- umbilical cord blood. J. Immunol. 2014, 192, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Foley, B.; Cooley, S.; Verneris, M.R.; Pitt, M.; Curtsinger, J.; Luo, X.; Lopez-Vergès, S.; Lanier, L.L.; Weisdorf, D.; Miller, J.S. Cytomegalovirus reactivation after allogeneic transplantation promotes a lasting increase in educated NKG2C+ natural killer cells with potent function. Blood 2012, 119, 2665–2674. [Google Scholar] [CrossRef] [PubMed]
- Diermayr, S.; Himmelreich, H.; Durovic, B.; Mathys-Schneeberger, A.; Siegler, U.; Langenkamp, U.; Hofsteenge, J.; Gratwohl, A.; Tichelli, A.; Paluszewska, M.; et al. NKG2D ligand expression in AML increases in response to HDAC inhibitor valproic acid and contributes to allorecognition by NK-cell lines with single KIR-HLA class I specificities. Blood 2008, 111, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Siegler, U.; Meyer-Monard, S.; Jörger, S.; Stern, M.; Tichelli, A.; Gratwohl, A.; Wodnar-Filipowicz, A.; Kalberer, C.P. Good manufacturing practice-compliant cell sorting and large-scale expansion of single KIR-positive alloreactive human natural killer cells for multiple infusions to leukemia patients. Cytotherapy 2010, 12, 750–763. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Béziat, V.; Oei, V.Y.S.; Pfefferle, A.; Schaffer, M.; Lehmann, S.; Hellström-Lindberg, E.; Söderhäll, S.; Heyman, M.; Grandér, D.; et al. Ex Vivo Expanded Adaptive NK Cells Effectively Kill Primary Acute Lymphoblastic Leukemia Cells. Cancer Immunol. Res. 2017, 5, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Bigley, A.B.; Spade, S.; Agha, N.H.; Biswas, S.; Tang, S.; Malik, M.H.; Dai, L.; Masoumi, S.; Patiño-Escobar, B.; Hale, M.; et al. FcεRIγ-negative NK cells persist in vivo and enhance efficacy of therapeutic monoclonal antibodies in multiple myeloma. Blood Adv. 2021, 5, 3021–3031. [Google Scholar] [CrossRef]
- Cho, H.; Kim, K.H.; Lee, H.; Kim, C.G.; Chung, H.; Choi, Y.S.; Park, S.H.; Cheong, J.W.; Min, Y.H.; Shin, E.C.; et al. Adaptive Natural Killer Cells Facilitate Effector Functions of Daratumumab in Multiple Myeloma. Clin. Cancer Res. 2021, 27, 2947–2958. [Google Scholar] [CrossRef]
- Rölle, A.; Pollmann, J.; Ewen, E.M.; Le, V.T.; Halenius, A.; Hengel, H.; Cerwenka, A. IL-12-producing monocytes and HLA-E control HCMV-driven NKG2C+ NK cell expansion. J. Clin. Investig. 2014, 124, 5305–5316. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Scott, J.M.; Hwang, I.; Kim, S. Cutting edge: Antibody-dependent memory-like NK cells distinguished by FcRγ deficiency. J. Immunol. 2013, 190, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Baek, H.J.; Kim, D.W.; Phan, M.T.; Kim, J.S.; Yang, J.H.; Choi, J.I.; Lee, J.J.; Shin, M.G.; Ryang, D.W.; Kim, S.K.; et al. Comparison of FcRγ-Deficient and CD57+ Natural Killer Cells Between Cord Blood and Adult Blood in the Cytomegalovirus-Endemic Korean Population. Ann. Lab. Med. 2015, 35, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Zhang, T.; Scott, J.M.; Kim, A.R.; Lee, T.; Kakarla, T.; Kim, A.; Sunwoo, J.B.; Kim, S. Identification of human NK cells that are deficient for signaling adaptor FcRγ and specialized for antibody-dependent immune functions. Int. Immunol. 2012, 24, 793–802. [Google Scholar] [CrossRef]
- Gao, F.; Zhou, Z.; Lin, Y.; Shu, G.; Yin, G.; Zhang, T. Biology and Clinical Relevance of HCMV-Associated Adaptive NK Cells. Front. Immunol. 2022, 13, 830396. [Google Scholar] [CrossRef] [PubMed]
- Thangaraj, J.L.; Phan, M.T.; Kweon, S.; Kim, J.; Lee, J.M.; Hwang, I.; Park, J.; Doh, J.; Lee, S.H.; Vo, M.C.; et al. Expansion of cytotoxic natural killer cells in multiple myeloma patients using K562 cells expressing OX40 ligand and membrane-bound IL-18 and IL-21. Cancer Immunol. Immunother. 2022, 71, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Kweon, S.; Phan, M.T.; Chun, S.; Yu, H.; Kim, J.; Kim, S.; Lee, J.; Ali, A.K.; Lee, S.H.; Kim, S.K.; et al. Expansion of Human NK Cells Using K562 Cells Expressing OX40 Ligand and Short Exposure to IL-21. Front. Immunol. 2019, 10, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, M.T.; Lee, S.H.; Kim, S.K.; Cho, D. Expansion of NK Cells Using Genetically Engineered K562 Feeder Cells. Methods Mol. Biol. 2016, 1441, 167–174. [Google Scholar] [CrossRef]
- Lim, D.P.; Jang, Y.Y.; Kim, S.; Koh, S.S.; Lee, J.J.; Kim, J.S.; Thi Phan, M.T.; Shin, D.J.; Shin, M.G.; Lee, S.H.; et al. Effect of exposure to interleukin-21 at various time points on human natural killer cell culture. Cytotherapy 2014, 16, 1419–1430. [Google Scholar] [CrossRef]
- Koh, S.K.; Park, J.; Kim, S.E.; Lim, Y.; Phan, M.T.; Kim, J.; Hwang, I.; Ahn, Y.O.; Shin, S.; Doh, J.; et al. Natural Killer Cell Expansion and Cytotoxicity Differ Depending on the Culture Medium Used. Ann. Lab. Med. 2022, 42, 638–649. [Google Scholar] [CrossRef]
- Wu, Z.; Sinzger, C.; Frascaroli, G.; Reichel, J.; Bayer, C.; Wang, L.; Schirmbeck, R.; Mertens, T. Human cytomegalovirus-induced NKG2C(hi) CD57(hi) natural killer cells are effectors dependent on humoral antiviral immunity. J. Virol. 2013, 87, 7717–7725. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Badeti, S.; Tseng, H.C.; Ma, M.T.; Liu, T.; Jiang, J.G.; Liu, C.; Liu, D. Superior Expansion and Cytotoxicity of Human Primary NK and CAR-NK Cells from Various Sources via Enriched Metabolic Pathways. Mol. Ther. Methods Clin. Dev. 2020, 18, 428–445. [Google Scholar] [CrossRef]
- Romee, R.; Leong, J.W.; Fehniger, T.A. Utilizing cytokines to function-enable human NK cells for the immunotherapy of cancer. Scientifica 2014, 2014, 205796. [Google Scholar] [CrossRef] [Green Version]
- Moseman, J.E.; Foltz, J.A.; Sorathia, K.; Heipertz, E.L.; Lee, D.A. Evaluation of serum-free media formulations in feeder cell-stimulated expansion of natural killer cells. Cytotherapy 2020, 22, 322–328. [Google Scholar] [CrossRef]
- Gumá, M.; Budt, M.; Sáez, A.; Brckalo, T.; Hengel, H.; Angulo, A.; López-Botet, M. Expansion of CD94/NKG2C+ NK cells in response to human cytomegalovirus-infected fibroblasts. Blood 2006, 107, 3624–3631. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.C.; Lopez-Verges, S.; Kim, C.C.; DeRisi, J.L.; Lanier, L.L. NK cells and immune “memory”. J. Immunol. 2011, 186, 1891–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.C.; Beilke, J.N.; Lanier, L.L. Adaptive immune features of natural killer cells. Nature 2009, 457, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Freud, A.G.; Mundy-Bosse, B.L.; Yu, J.; Caligiuri, M.A. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity 2017, 47, 820–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Zhang, T.; Hwang, I.; Kim, A.; Nitschke, L.; Kim, M.; Scott, J.M.; Kamimura, Y.; Lanier, L.L.; Kim, S. Epigenetic modification and antibody-dependent expansion of memory-like NK cells in human cytomegalovirus-infected individuals. Immunity 2015, 42, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Schlums, H.; Cichocki, F.; Tesi, B.; Theorell, J.; Beziat, V.; Holmes, T.D.; Han, H.; Chiang, S.C.; Foley, B.; Mattsson, K.; et al. Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function. Immunity 2015, 42, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Rho, E.Y.; In, J.W.; Kim, I.; Yoon, S.S.; Park, S.; Shin, S.; Park, K.U.; Song, E.Y. The impact of HLA and KIR ligand mismatching on unrelated allogeneic hematopoietic stem cell transplantation in Korean adult patients. Ann. Lab. Med. 2015, 35, 111–117. [Google Scholar] [CrossRef]
- Lee, N.; Goodlett, D.R.; Ishitani, A.; Marquardt, H.; Geraghty, D.E. HLA-E surface expression depends on binding of TAP-dependent peptides derived from certain HLA class I signal sequences. J. Immunol. 1998, 160, 4951–4960. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMVG | CMVM | KIR2DL2/3 | KIR2DL1 | KIR3DL1 | HLA-C Genotyping |
|---|---|---|---|---|---|
| Positive | Negative | 68.7 | 10.7 | 3.87 | C1C1 |
| Positive | Negative | 61.9 | 49.1 | 7.53 | C1C2 |
| Positive | Negative | 68.9 | 4.5 | 71.32 | C1C1 |
| Positive | Negative | 79.3 | 3.3 | 5.14 | C1C1 |
| Positive | Negative | 83.2 | 29.3 | 0.1 | C1C1 |
| Positive | Negative | 45.9 | 10.4 | 1.97 | C1C1 |
| Positive | Negative | 83.9 | 17.9 | 5.25 | C1C1 |
| Positive | Negative | 36.7 | 87.4 | 6.77 | C1C2 |
| Positive | Negative | 47.6 | 82.9 | 37.88 | C1C2 |
| Positive | Negative | 96.4 | 1.0 | 1.51 | C1C1 |
| Positive | Negative | 64.6 | 29.9 | 5.94 | C1C2 |
| Positive | Negative | 78.2 | 73.3 | 10.84 | C2C2 |
| Positive | Negative | 75.7 | 10.7 | 4.01 | C1C1 |
| Positive | Negative | 43.0 | 15.3 | 24.94 | C1C1 |
| Positive | Negative | 18.8 | 84.0 | 0.9 | C2C2 |
| Positive | Negative | 52.2 | 86.5 | 62.55 | C2C2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phan, M.-T.T.; Kim, J.; Koh, S.K.; Lim, Y.; Yu, H.; Lee, M.; Lee, J.-M.; Kang, E.-S.; Kim, H.-Y.; Kim, S.-K.; et al. Selective Expansion of NKG2C+ Adaptive NK Cells Using K562 Cells Expressing HLA-E. Int. J. Mol. Sci. 2022, 23, 9426. https://doi.org/10.3390/ijms23169426
Phan M-TT, Kim J, Koh SK, Lim Y, Yu H, Lee M, Lee J-M, Kang E-S, Kim H-Y, Kim S-K, et al. Selective Expansion of NKG2C+ Adaptive NK Cells Using K562 Cells Expressing HLA-E. International Journal of Molecular Sciences. 2022; 23(16):9426. https://doi.org/10.3390/ijms23169426
Chicago/Turabian StylePhan, Minh-Trang Thi, Jinho Kim, Seung Kwon Koh, Yuree Lim, Hongbi Yu, Mijeong Lee, Jong-Min Lee, Eun-Suk Kang, Hyun-Young Kim, Sang-Ki Kim, and et al. 2022. "Selective Expansion of NKG2C+ Adaptive NK Cells Using K562 Cells Expressing HLA-E" International Journal of Molecular Sciences 23, no. 16: 9426. https://doi.org/10.3390/ijms23169426
APA StylePhan, M.-T. T., Kim, J., Koh, S. K., Lim, Y., Yu, H., Lee, M., Lee, J.-M., Kang, E.-S., Kim, H.-Y., Kim, S.-K., Hwang, I., & Cho, D. (2022). Selective Expansion of NKG2C+ Adaptive NK Cells Using K562 Cells Expressing HLA-E. International Journal of Molecular Sciences, 23(16), 9426. https://doi.org/10.3390/ijms23169426