
Heparan Sulfate Facilitates Binding of hIFNγ to Its Cell-Surface Receptor hIFNGR1


 ,
,  , ,
, ,  , and
, and 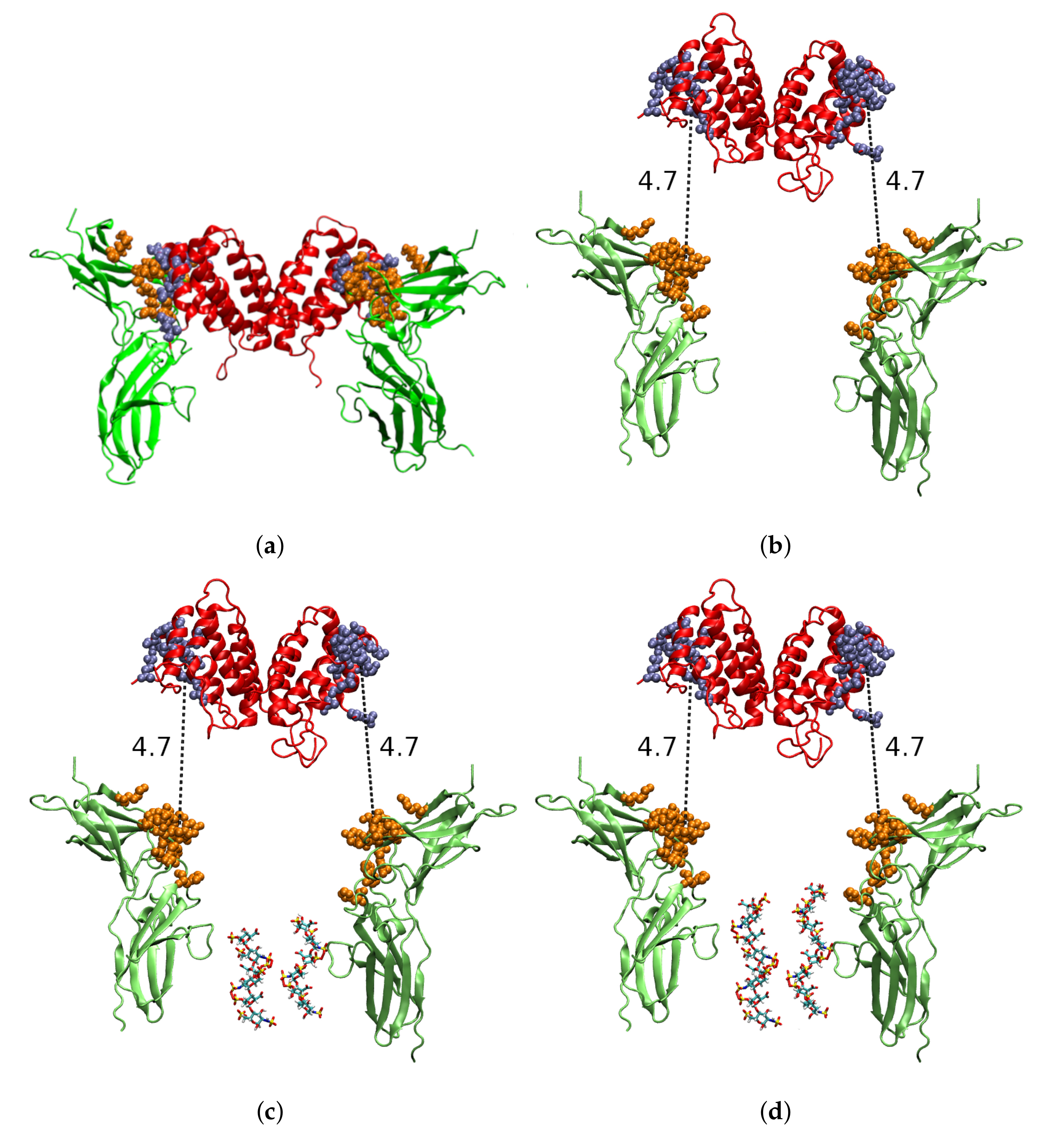{kind=link}
{kind=link}
{kind=link}
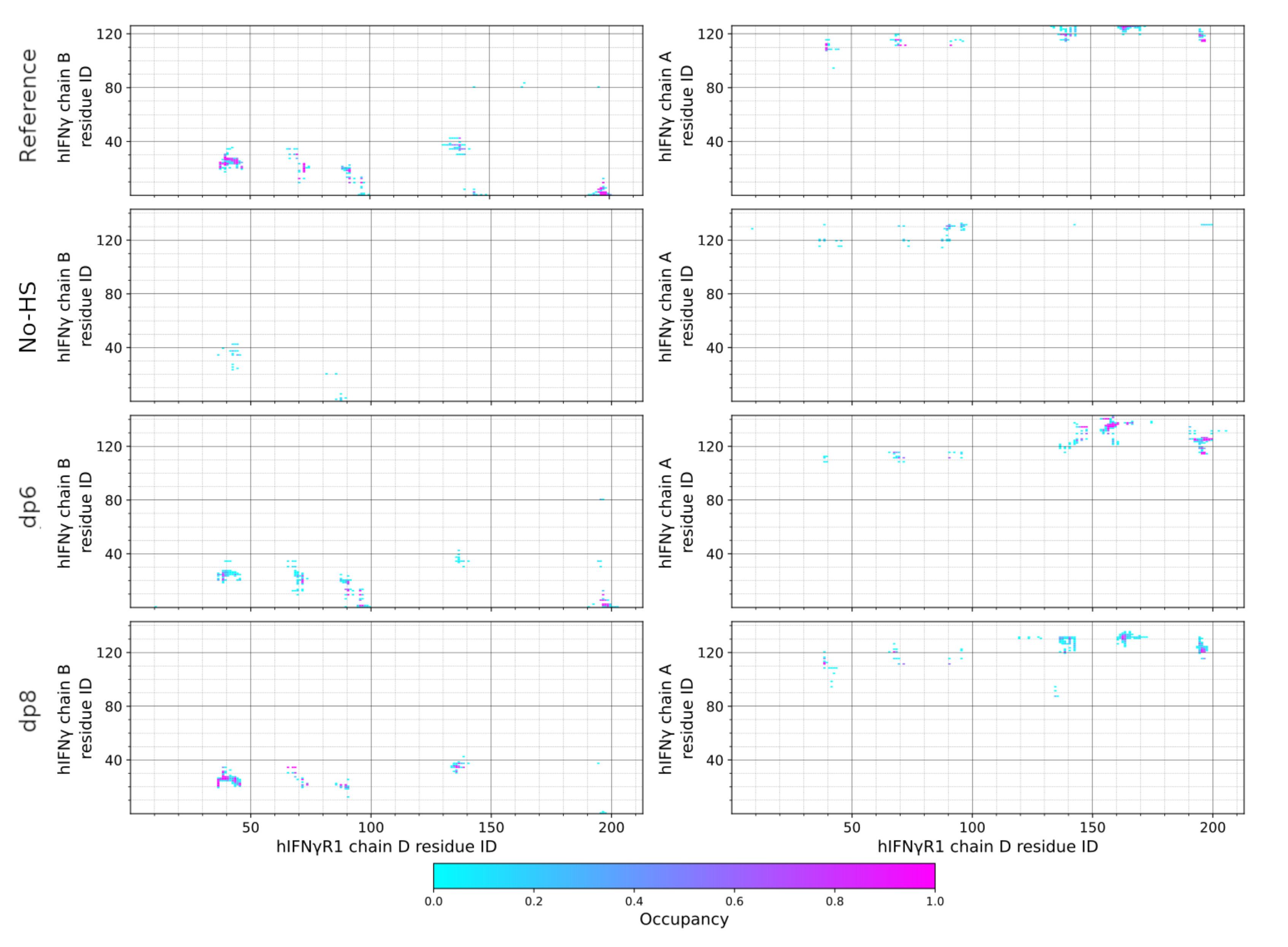{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
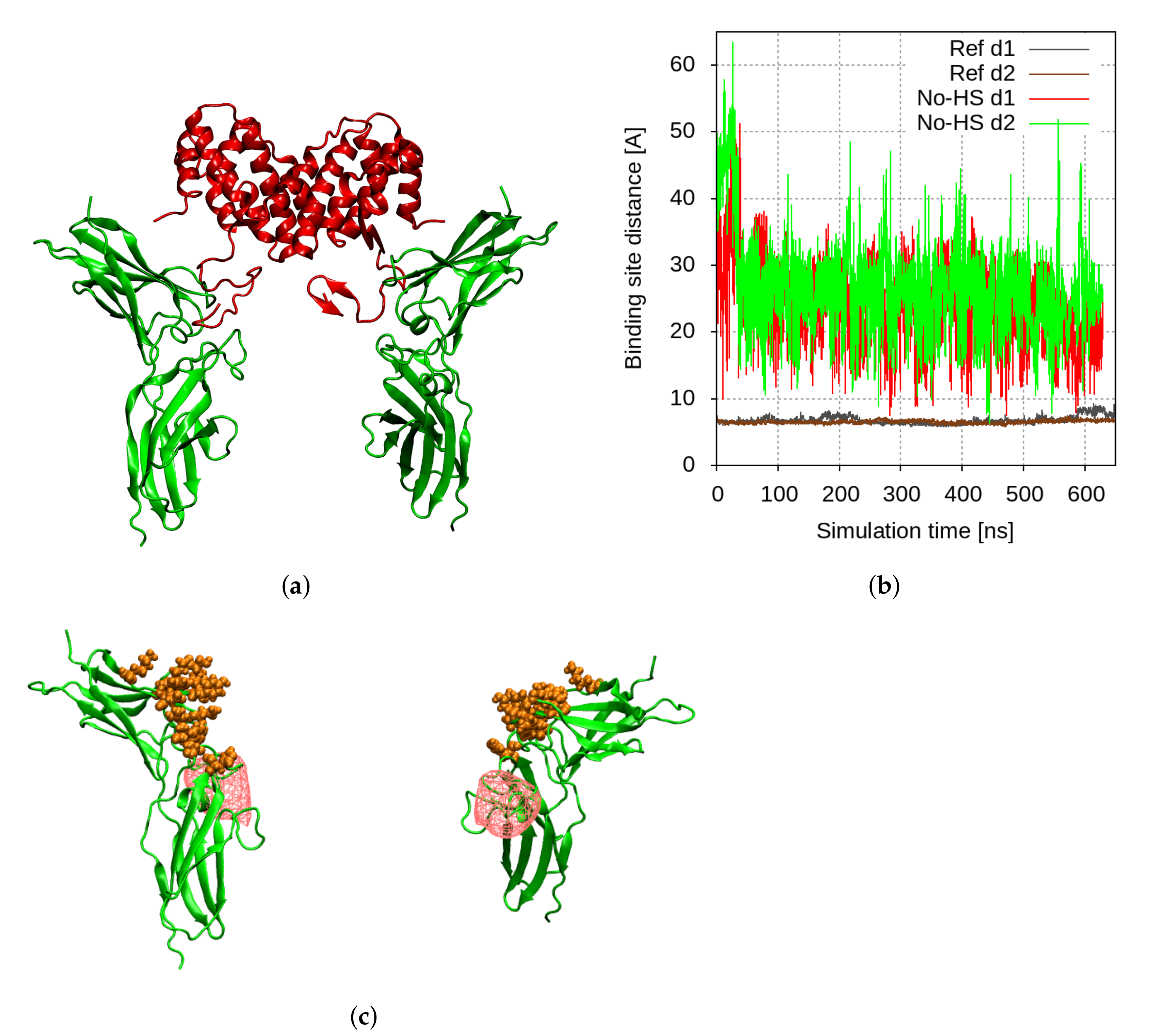
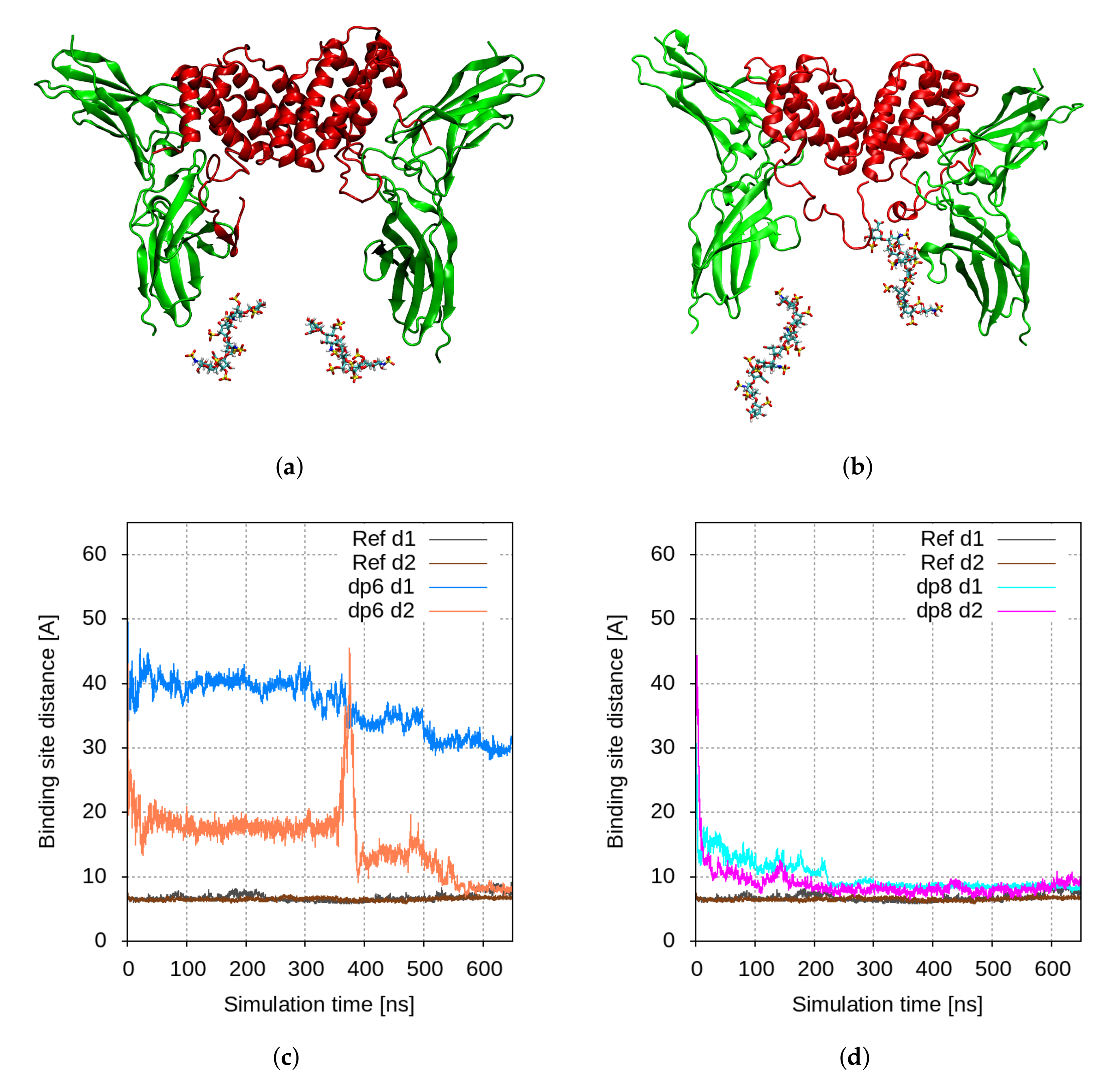
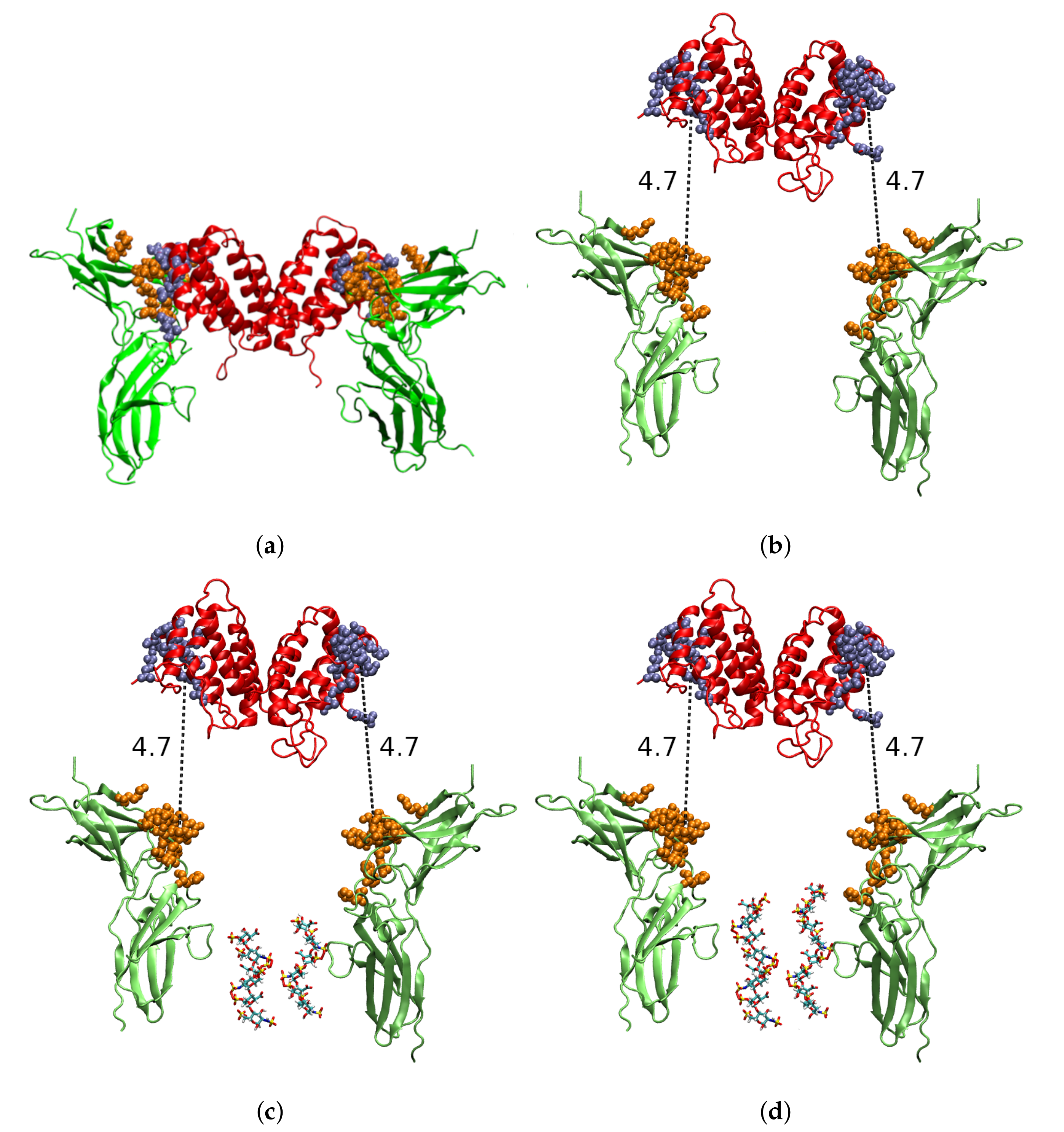
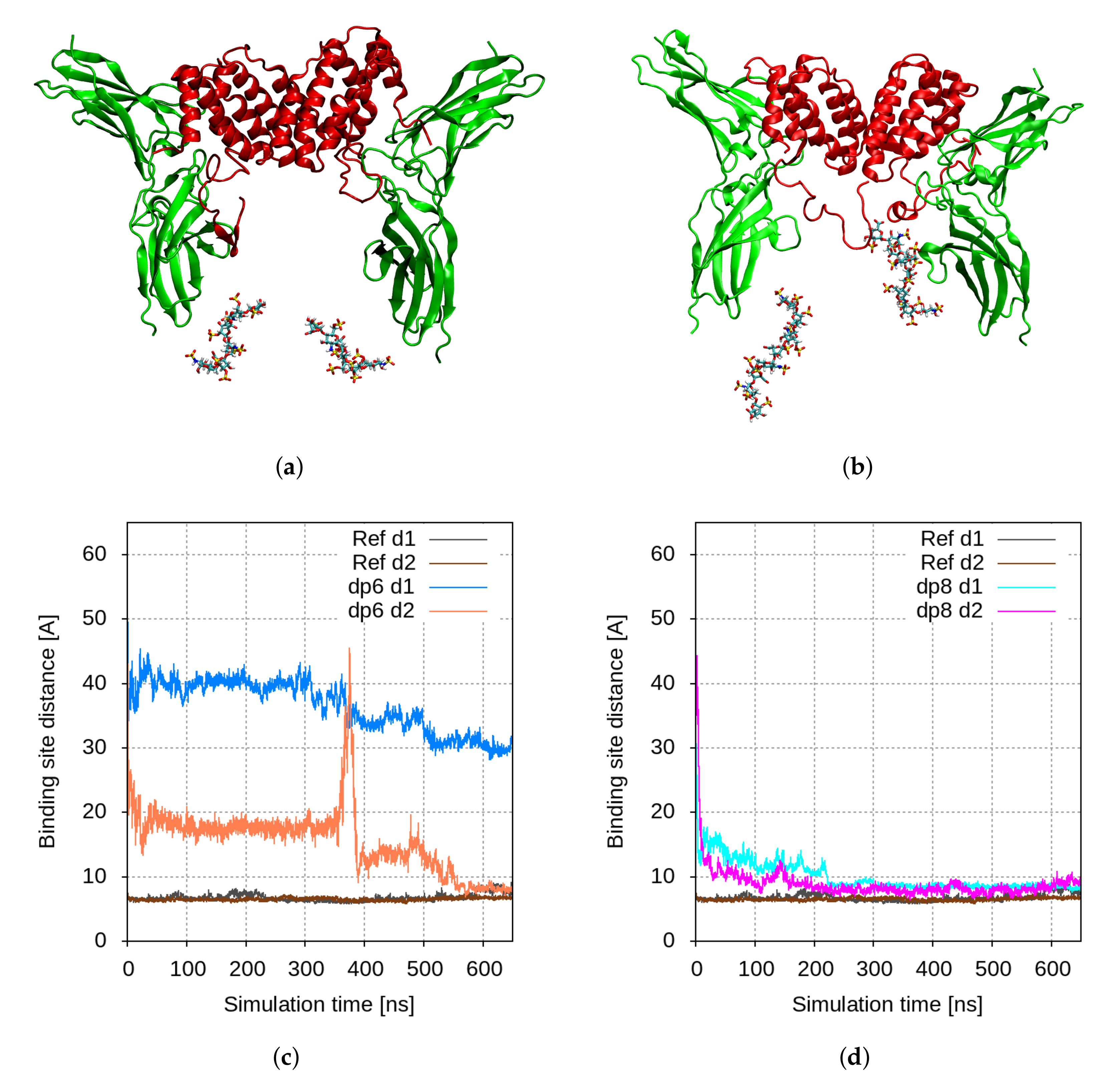
2.1. Molecular Modelling
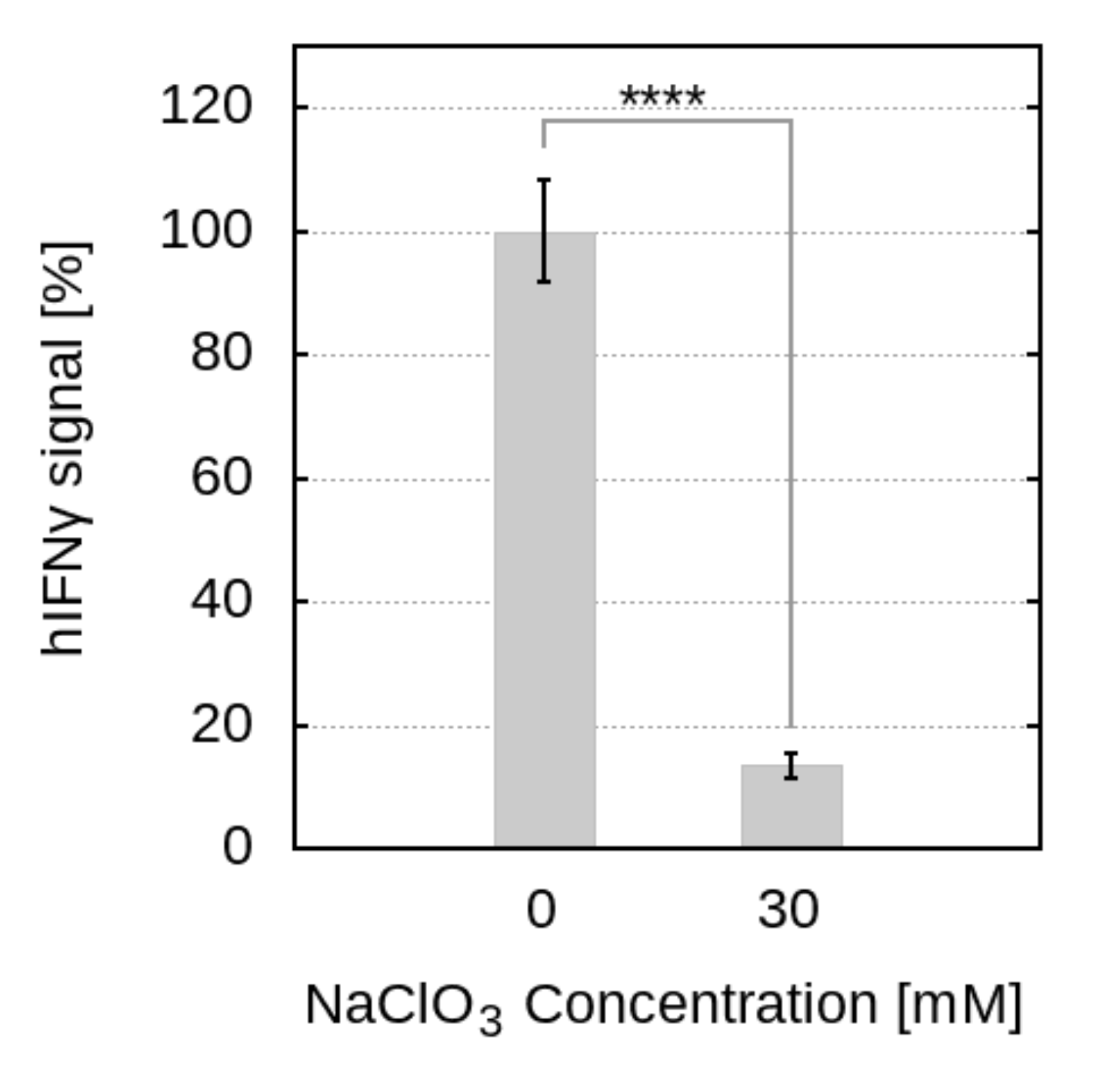
2.2. The Reduction of Sulfation of HS Drastically Reduces the Levels of Phosphorylated STAT1
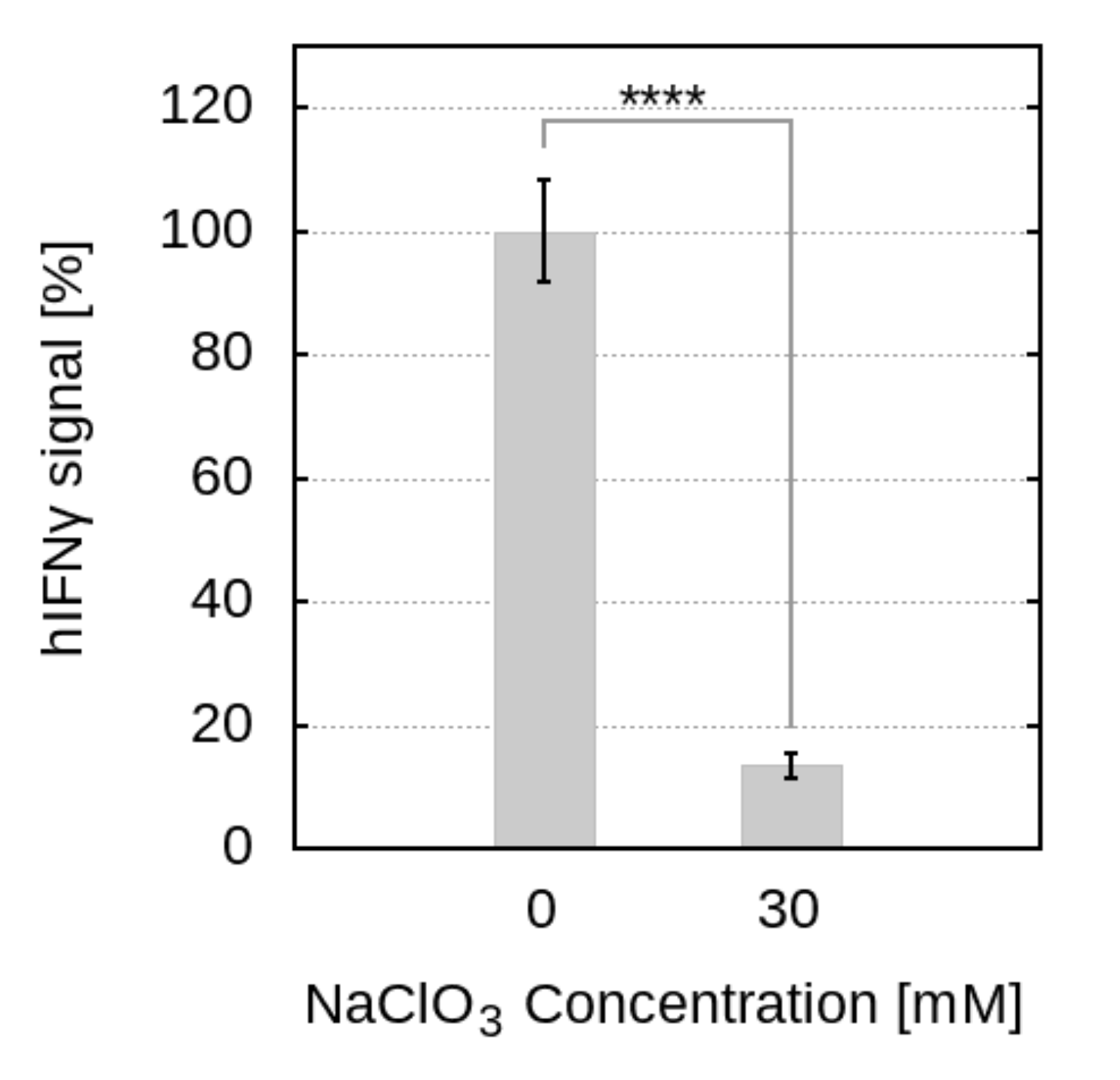
2.3. Disrupting HSPG Sulfation Decreases the Antiproliferative Activity of hIFN
3. Discussion
4. Materials and Methods
4.1. Molecular Dynamics Simulations
4.1.1. Input Structures
4.1.2. MD Simulation Protocol
4.2. In Vitro Experiments
4.2.1. Cell Culture and Phosphorylation of STAT1 after Cell Treatment with Sodium Chlorate
4.2.2. Biological Activity of hIFN after Treatment with Sodium Chlorate
4.2.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aa | Amino acid |
| CS | Chondroitin sulfate |
| dp | Degree of polymerization |
| GAG | Glycosaminoglycans |
| hIFN | Human interferon-gamma |
| hIFNGR | Human interferon-gamma receptor |
| hIFNGR1 | Human interferon-gamma receptor chain-1 |
| hIFNGR2 | Human interferon-gamma receptor chain-2 |
| H | Heparin |
| HS | Heparan sulfate |
| HSPG | Heparan sulfate proteoglycan |
| IFN | Interferon-gamma |
| IL-10 | Interleukine 10 |
| MD | Molecular dynamics |
| PG | Proteoglycans |
References
- Tsanev, R.G.; Ivanov, I. Immune Interferon: Properties and Clinical Applications; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar]
- Lees, J.R. Interferon gamma in autoimmunity: A complicated player on a complex stage. Cytokine 2015, 74, 18–26. [Google Scholar] [PubMed] [Green Version]
- Kulkarni, A.; Ganesan, P.; O’Donnell, L.A. Interferon Gamma: Influence on Neural Stem Cell Function in Neurodegenerative and Neuroinflammatory Disease. Clin. Med. Insights Pathol. 2016, 9, CPath.S40497. [Google Scholar]
- Seifert, H.A.; Collier, L.A.; Chapman, C.B.; Benkovic, S.A.; Willing, A.E.; Pennypacker, K.R. Pro-Inflammatory Interferon Gamma Signaling is Directly Associated with Stroke Induced Neurodegeneration. J. Neuroimmune Pharmacol. 2014, 9, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49. [Google Scholar] [PubMed]
- Ealick, S.E.; Cook, W.J.; Vijay-Kumar, S.; Carson, M.; Nagabhushan, T.L.; Trotta, P.P.; Bugg, C.E. Three-dimensional structure of recombinant human interferon-γ. Science 1991, 252, 698–702. [Google Scholar] [PubMed]
- Farrar, M.A.; Schreiber, R.D. The molecular cell biology of interferon-gamma and its receptor. Annu. Rev. Immunol. 1993, 11, 571–611. [Google Scholar] [CrossRef]
- Greenlund, A.; Farrar, M.; Viviano, B.; Schreiber, R. Ligand-induced IFN gamma receptor tyrosine phosphorylation couples the receptor to its signal transduction system (p91). EMBO J. 1994, 13, 1591–1600. [Google Scholar] [CrossRef]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar]
- Walter, M.R.; Windsor, W.T.; Nagabhushan, T.L.; Lundell, D.J.; Lunn, C.A.; Zauodny, P.J.; Narula, S.K. Crystal structure of a complex between interferon-gamma and its soluble high-affinity receptor. Nature 1995, 376, 230–235. [Google Scholar] [CrossRef]
- Thiel, D.; le Du, M.H.; Walter, R.; D’Arcy, A.; Chène, C.; Fountoulakis, M.; Garotta, G.; Winkler, F.; Ealick, S. Observation of an unexpected third receptor molecule in the crystal structure of human interferon-γ receptor complex. Structure 2000, 8, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Nacheva, G.; Todorova, K.; Boyanova, M.; Berzal-Herranz, A.; Karshikoff, A.; Ivanov, I. Human Interferon gamma: Significance of the C-terminal flexible domain for its biological activity. Arch. Biochem. Biophys. 2003, 413, 91–98. [Google Scholar] [CrossRef]
- Lortat-Jacob, H.; Baltzer, F.; Grimaud, J.A. Heparin Decreases the Blood Clearance of Interferon-γ and Increases Its Activity by Limiting the Processing of Its Carboxyl-terminal Sequence. J. Biol. Chem. 1996, 271, 16139–16143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugues, L.J.; Grimaud, J.A. Interferon-γ binds to heparan sulfate by a cluster of amino acids located in the C-terminal part of the molecule. FEBS Lett. 1991, 280, 152–154. [Google Scholar]
- Camejo, E.H.; Rosengren, B.; Camejo, G.; Sartipy, P.; Fager, G.; Bondjers, G. Interferon Gamma Binds to Extracellular Matrix Chondroitin-Sulfate Proteoglycans, Thus Enhancing Its Cellular Response. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 1456–1465. [Google Scholar] [CrossRef]
- Gandhi, N.S.; Mancera, R.L. The Structure of Glycosaminoglycans and their Interactions with Proteins. Chem. Biol. Drug Des. 2008, 72, 455–482. [Google Scholar] [CrossRef]
- Lortat-Jacob, H. Interferon and heparan sulphate. Biochem. Soc. Trans. 2006, 34, 461–464. [Google Scholar] [CrossRef] [Green Version]
- Lortat-Jacob, H.; Brisson, C.; Guerret, S.; Morel, G. Non-receptor-mediated tissue localization of human interferon-gamma: Role of heparan sulfate/heparin-like molecules. Cytokine 1996, 8, 557–566. [Google Scholar] [CrossRef]
- Sadir, R.; Forest, E.; Lortat-Jacob, H. The heparan sulfate binding sequence of interferon-gamma increased the on rate of the interferon-gamma-interferon-gamma receptor complex formation. J. Biol. Chem. 1998, 273, 10919–10925. [Google Scholar] [CrossRef] [Green Version]
- McGibbon, R.T.; Beauchamp, K.A.; Harrigan, M.P.; Klein, C.; Swails, J.M.; Hernández, C.X.; Schwantes, C.R.; Wang, L.P.; Lane, T.J.; Pande, V.S. MDTraj: A Modern Open Library for the Analysis of Molecular Dynamics Trajectories. Biophys. J. 2015, 109, 1528–1532. [Google Scholar] [CrossRef] [Green Version]
- Lortat-Jacob, H.; Kleinman, H.K.; Grimaud, J.A. High-affinity binding of interferon-gamma to a basement membrane complex (matrigel). J. Clin. Investig. 1991, 87, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Safaiyan, F.; Kolset, S.O.; Prydz, K.; Gottfridsson, E.; Lindahl, U.; Salmivirta, M. Selective Effects of Sodium Chlorate Treatment on the Sulfation of Heparan Sulfate. J. Biol. Chem. 1999, 274, 36267–36273. [Google Scholar] [CrossRef] [Green Version]
- Sadir, R.; Lortat-Jacob, H.; Morel, G. Internalization and Nuclear Translocation of IFN-γ and IFN-γR: An Ultrastructural Approach. Cytokine 2000, 12, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Stockert, J.C.; Horobin, R.W.; Colombo, L.L.; Blázquez-Castro, A. Tetrazolium salts and formazan products in Cell Biology: Viability assessment, fluorescence imaging, and labeling perspectives. Acta Histochem. 2018, 120, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Loading Control Handbook. Novus Biologicals. Available online: https://images.novusbio.com/design/BR_loadingcontrolsguide_101416.pdf (accessed on 30 June 2022).
- Boyanova, M.; Tsanev, R.; Ivanov, I. A modified kynurenine bioassay for quantitative determination of human interferon-γ. Anal. Biochem. 2002, 308, 178–181. [Google Scholar] [CrossRef]
- Miller, C.H.; Maher, S.G.; Young, H.A. Clinical Use of Interferon-gamma. Ann. N. Y. Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef]
- Nacheva, G.; Boyanova, M.; Todorova, K.A.; Kyurkchiev, S.D.; Ivanov, I. Characterization of functional domains of human interferon gamma by specific monoclonal antibodies. Folia Biol. 2002, 48, 96–101. [Google Scholar]
- Hogrefe, H.H.; McPhie, P.; Bekisz, J.B.; Enterline, J.C.; Dyer, D.; Webb, D.S.A.; Gerrard, T.L.; Zoon, K.C. Amino Terminus Is Essential to the Structural Integrity of Recombinant Human Interferon-γ. J. Biol. Chem. 1989, 264, 12179–12186. [Google Scholar] [CrossRef]
- Arakawa, T.; Narachi, M.A.; Hsu, Y.R.; Everett, R.R.; Lai, P.H.; Fish, E.N. The effect of C-terminal processing on the activity of human interferon-gamma. Drug Des. Deliv. 1989, 4, 217–225. [Google Scholar]
- Honda, S.; Asano, T.; Kajio, T.; Nakagawa, S.; Ikeyama, S.; Ichimori, Y.; Sugino, H.; Nara, K.; Kakinuma, A.; Kung, H.F. Differential Purification by Immunoaffinity Chromatography of Two Carboxy-Terminal Portion-Deleted Derivatives of Recombinant Human Interferon-γ from Escherichia coli. J. Interferon Res. 1987, 7, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Slodowski, O.; Böhm, J.; Schöne, B.; Otto, B. Carboxy-terminal truncated rhuIFN-γ with a substitution of Gln133 or Ser132 to leucine leads to higher biological activity than in the wild type. Eur. J. Biochem. 1991, 202, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.; Perry, L.J.; Veilleux, C.; Chang, G. Mutational analysis of the C-terminus of human interferon-γ. Protein Eng. Des. Sel. 1990, 3, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.C.E.; Stern, A.S.; Familletti, P.C.; Chizzonite, R.; Khan, F.R. Structural characterization of human interferon γ Heterogeneity of the carboxyl terminus. Eur. J. Biochem. 1987, 166, 145–149. [Google Scholar] [CrossRef]
- Haelewyn, J.; Michiels, L.; Verhaert, P.; Hoylaerts, M.F.; Witters, R.; De Ley, M. Interaction of truncated human interferon gamma variants with the interferon gamma receptor: Crucial importance of Arg-129. Biochem. J. 1997, 324, 591–595. [Google Scholar] [CrossRef]
- Oyler-Yaniv, J.; Oyler-Yaniv, A.; Shakiba, M.; Min, N.K.; Chen, Y.H.; Cheng, S.Y.; Krichevsky, O.; Altan-Bonnet, N.; Altan-Bonnet, G. Catch and Release of Cytokines Mediated by Tumor Phosphatidylserine Converts Transient Exposure into Long-Lived Inflammation. Mol. Cell 2017, 66, 635–647.e7. [Google Scholar] [CrossRef] [Green Version]
- Vanhaverbeke, C.; Simorre, J.P.; Sadir, R.; Gans, P.; Lortat-Jacob, H. NMR characterization of the interaction between the C-terminal domain of interferon-γ and heparin-derived oligosaccharides. Biochem. J. 2004, 384, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Lortat-Jacob, H.; Turnbull, J.E.; Grimaud, J.A. Molecular organization of the interferon γ-binding domain in heparan sulphate. Biochem. J. 1995, 310, 497–505. [Google Scholar] [CrossRef]
- Hayashida, K.; Aquino, R.S.; Park, P.W. Coreceptor functions of cell surface heparan sulfate proteoglycans. Am. J.-Physiol.-Cell Physiol. 2022, 322, C896–C912. [Google Scholar] [CrossRef]
- Hurt-Camejo, E.; Rosengren, B.; Sartipy, P.; Elfsberg, K.; Camejo, G.; Svensson, L. CD44, a Cell Surface Chondroitin Sulfate Proteoglycan, Mediates Binding of Interferon-γ and Some of Its Biological Effects on Human Vascular Smooth Muscle Cells. J. Biol. Chem. 1999, 274, 18957–18964. [Google Scholar] [CrossRef] [Green Version]
- Castilla-Casadiego, D.A.; García, J.R.; García, A.J.; Almodovar, J. Heparin/Collagen Coatings Improve Human Mesenchymal Stromal Cell Response to Interferon Gamma. ACS Biomater. Sci. Eng. 2019, 5, 2793–2803. [Google Scholar] [CrossRef] [PubMed]
- Zdanov, A.; Schalk-Hihi, C.; Gustchina, A.; Tsang, M.; Weatherbee, J.; Wlodawer, A. Crystal structure of interleukin-10 reveals the functional dimer with an unexpected topological similarity to interferon γ. Structure 1995, 3, 591–601. [Google Scholar] [CrossRef]
- Salek-Ardakani, S.; Arrand, J.R.; Shaw, D.; Mackett, M. Heparin and heparan sulfate bind interleukin-10 and modulate its activity. Blood 2000, 96, 1879–1888. [Google Scholar] [CrossRef] [PubMed]
- Litov, L.; Petkov, P.; Rangelov, M.; Ilieva, N.; Lilkova, E.; Todorova, N.; Krachmarova, E.; Malinova, K.; Gospodinov, A.; Hristova, R.; et al. Molecular Mechanism of the Anti-Inflammatory Action of Heparin. Int. J. Mol. Sci. 2021, 22, 10730. [Google Scholar] [CrossRef]
- Perez Sanchez, H.; Tatarenko, K.; Nigen, M.; Pavlov, G.; Imberty, A.; Lortat-Jacob, H.; Garcia de la Torre, J.; Ebel, C. Organization of human interferon gamma-heparin complexes from solution properties and hydrodynamics. Biochemistry 2006, 45, 13227–13238. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Petkov, P.; Lilkova, E.; Ilieva, N.; Nacheva, G.; Ivanov, I.; Litov, L. Computational Modelling of the Full Length hIFN-γ Homodimer. In Large-Scale Scientific Computing LSSC 2017; Lirkov, I., Margenov, S., Eds.; Lecture Notes in Computer Science; Springer: Cham, Switzerland, 2018; Volume 10665. [Google Scholar] [CrossRef]
- Lilkova, E.; Petkov, P.; Ilieva, N.; Krachmarova, E.; Nacheva, G.; Litov, L. Molecular modeling of the effects of glycosylation on the structure and dynamics of human interferon-gamma. J. Mol. Model. 2019, 25, 127. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Lee, J.; Qi, Y.; Kern, N.R.; Lee, H.S.; Jo, S.; Joung, I.; Joo, K.; Lee, J.; Im, W. CHARMM-GUI Glycan Modeler for modeling and simulation of carbohydrates and glycoconjugates. Glycobiology 2019, 29, 320–331. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Guvench, O.; Mallajosyula, S.S.; Raman, E.P.; Hatcher, E.; Vanommeslaeghe, K.; Foster, T.J.; Jamison, F.W.; MacKerell, A.D. CHARMM Additive All-Atom Force Field for Carbohydrate Derivatives and Its Utility in Polysaccharide and Carbohydrate–Protein Modeling. J. Chem. Theory Comput. 2011, 7, 3162–3180. [Google Scholar] [CrossRef] [Green Version]
- Case, D.A.; Betz, R.M.; Cerutti, D.S.; Cheatham, T.E., III; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Homeyer, N.; et al. AMBER 2016; University of California: San Francisco, CA, USA, 2016; Available online: https://ambermd.org/AmberTools.php (accessed on 1 January 2016).
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmüller, H.; MacKerell, A.D., Jr. CHARMM36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2016, 14, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684. [Google Scholar] [CrossRef] [Green Version]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, M.; Rahman, A. Crystal Structure and Pair Potentials: A Molecular-Dynamics Study. Phys. Rev. Lett. 1980, 45, 1196. [Google Scholar] [CrossRef]
- Hockney, R.; Goel, S.; Eastwood, J. Quiet high-resolution computer models of a plasma. J. Comput. Phys. 1974, 14, 148–158. [Google Scholar] [CrossRef]
- Hess, B. P-LINCS: A Parallel Linear Constraint Solver for Molecular Simulation. J. Chem. Theory Comput. 2008, 4, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- Pestka, S.; Langer, J.A.; Zoon, K.C.; Samuel, C.E. Interferons and their actions. Annu. Rev. Biochem. 1987, 56, 727–777. [Google Scholar] [CrossRef]
- Meek, W.D.; Davis, W.L. Fine Structure and Immunofluorescent Studies of the WISH Cell Line. Vitr. Cell. Dev. Biol. 1986, 22, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Tileva, M.; Krachmarova, E.; Ivanov, I.; Maskos, K.; Nacheva, G. Production of aggregation prone human interferon gamma and its mutant in highly soluble and biologically active form by SUMO fusion technology. Protein Expr. Purif. 2016, 117, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Däubener, W.; Wanagat, N.; Pilz, K.; Seghrouchni, S.; Fischer, H.G.; Hadding, U. A new, simple, bioassay for human IFN-gamma. J. Immunol. Methods 1994, 168, 39–47. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miladinova, E.; Lilkova, E.; Krachmarova, E.; Malinova, K.; Petkov, P.; Ilieva, N.; Nacheva, G.; Litov, L. Heparan Sulfate Facilitates Binding of hIFNγ to Its Cell-Surface Receptor hIFNGR1. Int. J. Mol. Sci. 2022, 23, 9415. https://doi.org/10.3390/ijms23169415
Miladinova E, Lilkova E, Krachmarova E, Malinova K, Petkov P, Ilieva N, Nacheva G, Litov L. Heparan Sulfate Facilitates Binding of hIFNγ to Its Cell-Surface Receptor hIFNGR1. International Journal of Molecular Sciences. 2022; 23(16):9415. https://doi.org/10.3390/ijms23169415
Chicago/Turabian StyleMiladinova, Elisaveta, Elena Lilkova, Elena Krachmarova, Kristina Malinova, Peicho Petkov, Nevena Ilieva, Genoveva Nacheva, and Leandar Litov. 2022. "Heparan Sulfate Facilitates Binding of hIFNγ to Its Cell-Surface Receptor hIFNGR1" International Journal of Molecular Sciences 23, no. 16: 9415. https://doi.org/10.3390/ijms23169415
APA StyleMiladinova, E., Lilkova, E., Krachmarova, E., Malinova, K., Petkov, P., Ilieva, N., Nacheva, G., & Litov, L. (2022). Heparan Sulfate Facilitates Binding of hIFNγ to Its Cell-Surface Receptor hIFNGR1. International Journal of Molecular Sciences, 23(16), 9415. https://doi.org/10.3390/ijms23169415