
Conformational Changes of α-Crystallin Proteins Induced by Heat Stress

 and
and
Abstract
1. Introduction
2. Results
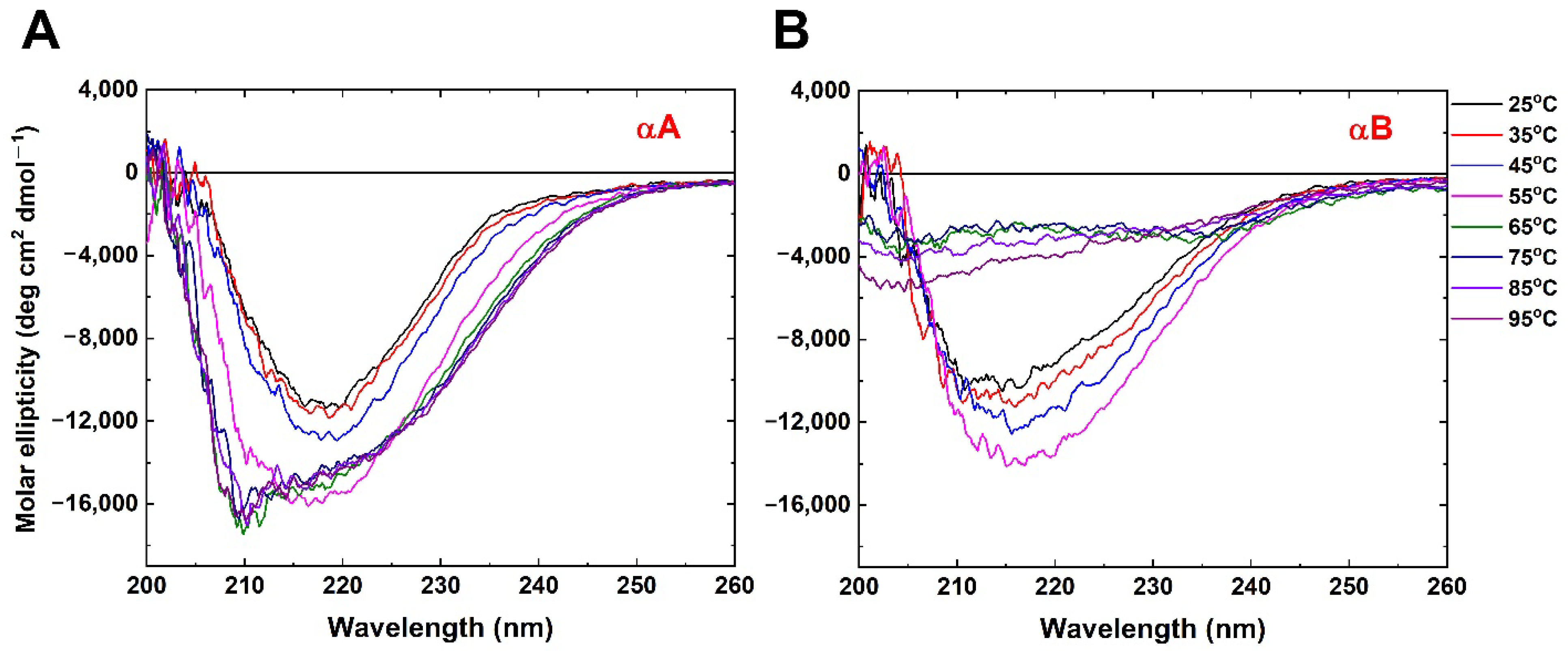
2.1. Circular Dichroism Shows Conformational Change Induced through Elevated Temperature
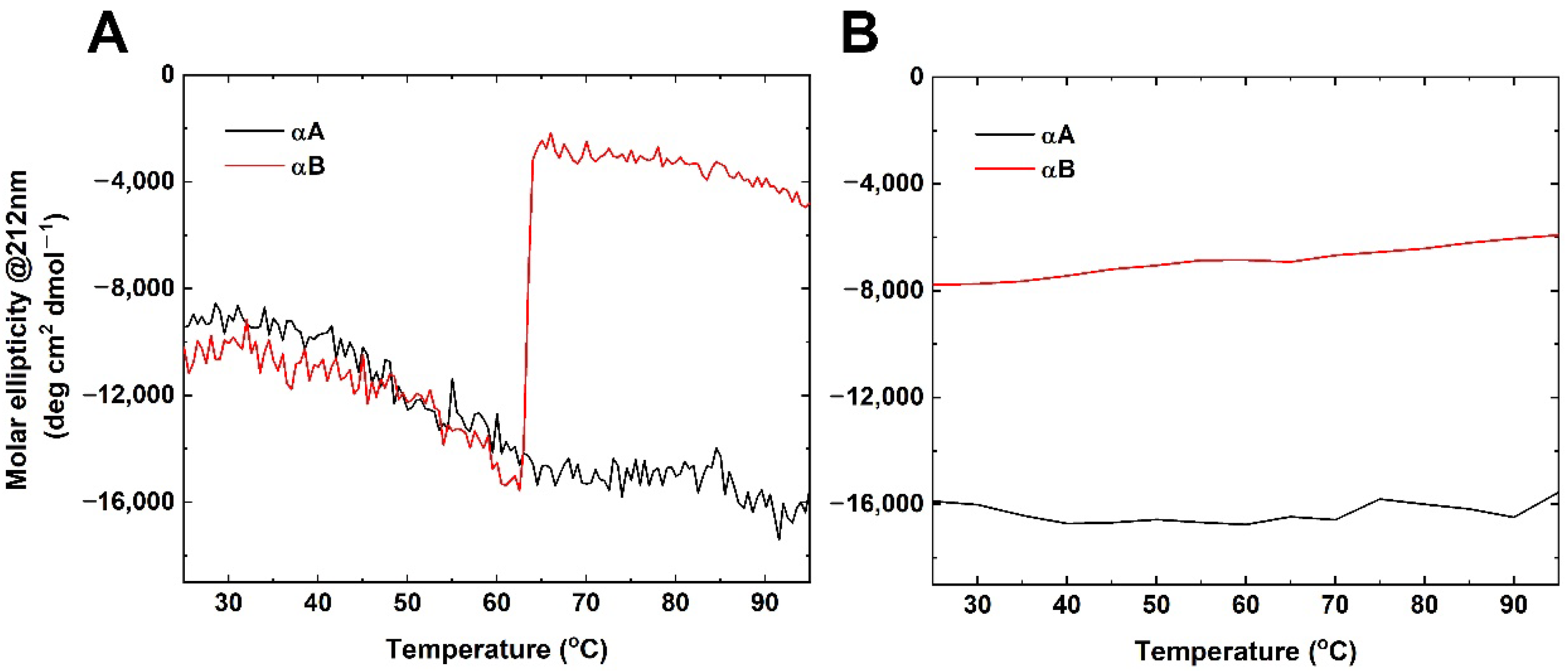
2.2. Thermal Denaturation Studies
2.3. Enhancement of Chaperone-like Activity in αA but Loss of Function in αB Crystallin at High Temperatures
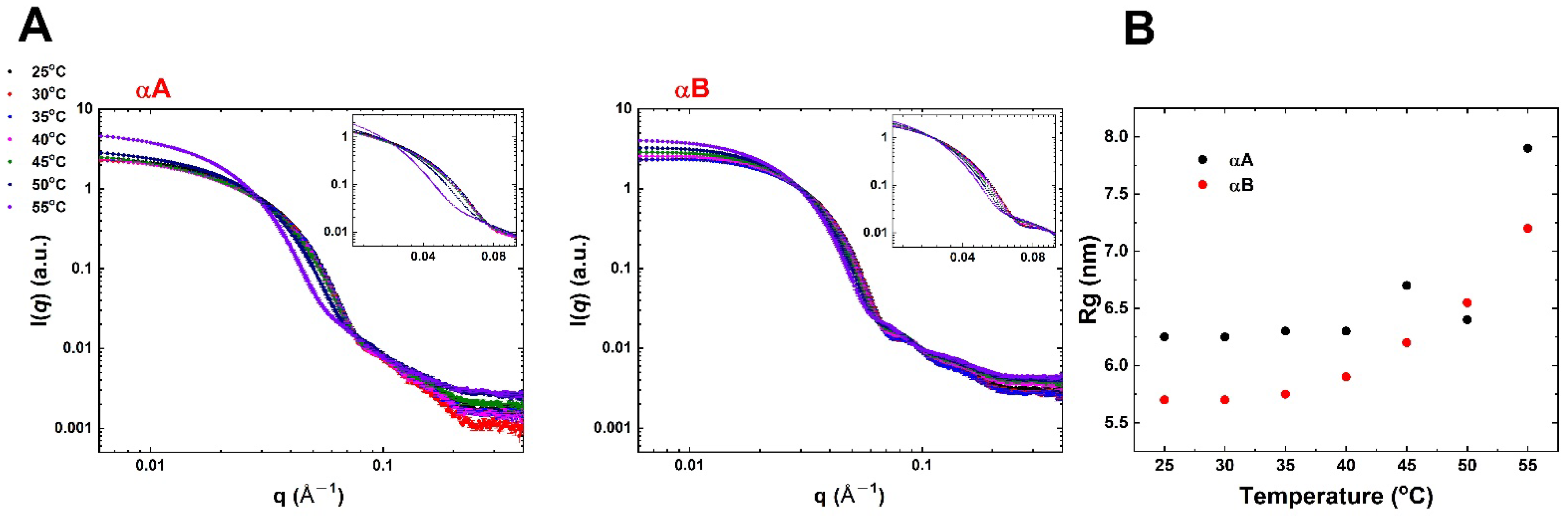
2.4. Heat-Induced Changes in Conformation and Size by SAXS Analysis
3. Discussion
4. Materials and Methods
4.1. Cloning, Protein Expression, and Purification
4.2. Circular Dichroism Spectroscopy
4.3. Chaperone Activity Assay
4.4. Small-Angle X-ray Scattering (SAXS) Measurements
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bloemendal, H. The vertebrate eye lens. Science 1977, 197, 127–138. [Google Scholar] [CrossRef]
- Ingolia, T.D.; Craig, E.A. Four small Drosophila heat shock proteins are related to each other and to mammalian alpha-crystallin. Proc. Natl. Acad. Sci. USA 1982, 79, 2360–2364. [Google Scholar] [CrossRef]
- Klein, R.; Klein, B.E. The prevalence of age-related eye diseases and visual impairment in aging: Current estimates. Investig. Ophthalmol. Vis. Sci. 2013, 54, ORSF5–ORSF13. [Google Scholar] [CrossRef] [PubMed]
- Petrash, J.M. Aging and age-related diseases of the ocular lens and vitreous body. Investig. Ophthalmol. Vis. Sci. 2013, 54, ORSF54–ORSF59. [Google Scholar] [CrossRef] [PubMed]
- Brian, G.; Taylor, H. Cataract blindness—Challenges for the 21st century. Bull. World Health Organ. 2001, 79, 249–256. [Google Scholar] [PubMed]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef]
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef]
- Horwitz, J. Proctor Lecture. The function of alpha-crystallin. Investig. Ophthalmol. Vis. Sci. 1993, 34, 10–22. [Google Scholar]
- Reddy, G.B.; Kumar, P.A.; Kumar, M.S. Chaperone-like activity and hydrophobicity of alpha-crystallin. IUBMB Life 2006, 58, 632–641. [Google Scholar] [CrossRef]
- Christopher, K.L.; Pedler, M.G.; Shieh, B.; Ammar, D.A.; Petrash, J.M.; Mueller, N.H. Alpha-crystallin-mediated protection of lens cells against heat and oxidative stress-induced cell death. Biochim. Biophys. Acta 2014, 1843, 309–315. [Google Scholar] [CrossRef]
- Augusteyn, R.C. Alpha-crystallin: A review of its structure and function. Clin. Exp. Optom. 2004, 87, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Satoh, T.; Shinoda, H.; Samejima, T.; Wu, S.-H.; Chiou, S.-H. Effect of heat-induced structural perturbation of secondary and tertiary structures on the chaperone activity of alpha-crystallin. Biochem. Biophys. Res. Commun. 1997, 237, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Burgio, M.R.; Kim, C.J.; Dow, C.C.; Koretz, J.F. Correlation between the chaperone-like activity and aggregate size of alpha-crystallin with increasing temperature. Biochem. Biophys. Res. Commun. 2000, 268, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, V.; Maulucci, G.; Maiorana, A.; Papi, M.; De Spirito, M. Alpha-crystallin modulates its chaperone activity by varying the exposed surface. ChemBioChem 2013, 14, 2362–2370. [Google Scholar] [CrossRef] [PubMed]
- Wistow, G.J.; Piatigorsky, J. Lens crystallins: The evolution and expression of proteins for a highly specialized tissue. Annu. Rev. Biochem. 1988, 57, 479–504. [Google Scholar] [CrossRef]
- Wistow, G. Lens crystallins: Gene recruitment and evolutionary dynamism. Trends Biochem. Sci. 1993, 18, 301–306. [Google Scholar] [CrossRef]
- Sun, T.X.; Liang, J.J. Intermolecular exchange and stabilization of recombinant human alphaA- and alphaB-crystallin. J. Biol. Chem. 1998, 273, 286–290. [Google Scholar] [CrossRef]
- Van Boekel, M.A.; de Lange, F.; de Grip, W.J.; de Jong, W.W. Eye lens alphaA- and alphaB-crystallin: Complex stability versus chaperone-like activity. Biochim. Biophys. Acta 1999, 1434, 114–123. [Google Scholar] [CrossRef]
- Kim, K.K.; Kim, R.; Kim, S.-H. Crystal structure of a small heat-shock protein. Nature 1998, 394, 595–599. [Google Scholar] [CrossRef]
- Morrow, G.; Tanguay, R.M. Small heat shock protein expression and functions during development. Int. J. Biochem. Cell Biol. 2012, 44, 1613–1621. [Google Scholar] [CrossRef]
- Pelton, T.J.; McLean, L.R. Spectroscopic methods for analysis of protein secondary structure. Anal. Biochem. 2000, 277, 167–176. [Google Scholar] [CrossRef]
- Narberhaus, F. α-Crystallin-type heat shock proteins: Socializing minichaperones in the context of a multichaperone network. Microbiol. Mol. Biol. Rev. 2002, 66, 64–93. [Google Scholar] [CrossRef] [PubMed]
- Bloemendal, H.; de Jong, W.; Jaenicke, R.; Lubsen, N.H.; Slingsby, C.; Tardieu, A. Ageing and vision: Structure, stability and function of lens crystallins. Prog. Biophys. Mol. Biol. 2004, 86, 407–485. [Google Scholar] [CrossRef] [PubMed]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Hendrick, J.P.; Hartl, F.U. Molecular chaperone functions of heat-shock proteins. Annu. Rev. Biochem. 1993, 62, 349–384. [Google Scholar] [CrossRef] [PubMed]
- Haley, D.A.; Horwitz, J.; Stewart, P.L. The small heat-shock protein, alphaB-crystallin, has a variable quaternary structure. J. Mol. Biol. 1998, 277, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Hura, G.L.; Menon, A.L.; Hammel, M.; Rambo, R.P.; Ii, F.L.P.; Tsutakawa, S.E.; Jenney, F.E., Jr.; Classen, S.; A Frankel, K.; Hopkins, R.C.; et al. Robust, high-throughput solution structural analyses by small angle X-ray scattering (SAXS). Nat. Methods 2009, 6, 606–612. [Google Scholar] [CrossRef]
- Skou, S.; Gillilan, R.E.; Ando, N. Synchrotron-based small-angle X-ray scattering of proteins in solution. Nat. Protoc. 2014, 9, 1727–1739. [Google Scholar] [CrossRef]
- Neylon, C. Small angle neutron and X-ray scattering in structural biology: Recent examples from the literature. Eur. Biophys. J. 2008, 37, 531–541. [Google Scholar] [CrossRef]
- Nagar, B.; Kuriyan, J. SAXS and the working protein. Structure 2005, 13, 169–170. [Google Scholar] [CrossRef][Green Version]
- Das, K.P.; Surewicz, W.K. Temperature-induced exposure of hydrophobic surfaces and its effect on the chaperone activity of alpha-crystallin. FEBS Lett. 1995, 369, 321–325. [Google Scholar] [CrossRef]
- Raman, B.; Ramakrishna, T.; Rao, C.M. Temperature dependent chaperone-like activity of alpha-crystallin. FEBS Lett. 1995, 365, 133–136. [Google Scholar] [CrossRef]
- Gesierich, U.; Pfeil, W. The conformational stability of alpha-crystallin is rather low: Calorimetric results. FEBS Lett. 1996, 393, 151–154. [Google Scholar] [CrossRef][Green Version]
- Walsh, M.T.; Sen, A.C.; Chakrabarti, B. Micellar subunit assembly in a three-layer model of oligomeric alpha-crystallin. J. Biol. Chem. 1991, 266, 20079–20084. [Google Scholar] [CrossRef]
- Surewicz, W.K.; Olesen, P.R. On the thermal stability of alpha-crystallin: A new insight from infrared spectroscopy. Biochemistry 1995, 34, 9655–9660. [Google Scholar] [CrossRef] [PubMed]
- Maiti, M.; Kono, M.; Chakrabarti, B. Heat-induced changes in the conformation of alpha- and beta-crystallins: Unique thermal stability of alpha-crystallin. FEBS Lett. 1988, 236, 109–114. [Google Scholar] [CrossRef]
- Das, B.K.; Liang, J.J.; Chakrabarti, B. Heat-induced conformational change and increased chaperone activity of lens alpha-crystallin. Curr. Eye Res. 1997, 16, 303–309. [Google Scholar] [CrossRef]
- Rasmussen, T.; van de Weert, M.; Jiskoot, W.; Kasimova, M.R. Thermal and acid denaturation of bovine lens alpha-crystallin. Proteins 2011, 79, 1747–1758. [Google Scholar] [CrossRef]
- Zhang, K.; Zhu, X.; Lu, Y. Effect of mild heating on human lens epithelial cells: A possible model of lens aging. Sci. Rep. 2016, 6, 33917. [Google Scholar] [CrossRef]
- Tjondro, H.C.; Xi, Y.B.; Chen, X.J.; Su, J.T.; Yan, Y.B. Membrane insertion of αA-crystallin is oligomer-size dependent. Biochem. Biophys. Res. Commun. 2016, 473, 1–7. [Google Scholar] [CrossRef]
- Selivanova, O.M.; Galzitskaya, O.V. Structural and Functional Peculiarities of α-Crystallin. Biology 2020, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.J.; Ramalli, S.G.; Wallace, B.A. DichroWeb, a website for calculating protein secondary structure from circular dichroism spectroscopic data. Protein Sci. 2022, 31, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Konarev, P.V.; Volkov, V.V.; Sokolova, A.V.; Koch, M.H.J.; Svergun, D.I. PRIMUS: A windows PC-based system for small-angle scattering data analysis. J. Appl. Crystallogr. 2003, 36, 1277–1282. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| αA Crystallin | αB Crystallin | |||||
|---|---|---|---|---|---|---|
| α-Helix * | β-Sheet ** | Unordered | α-Helix * | β-Sheet ** | Unordered | |
| 25 °C | 0.54 | 0.24 | 0.22 | 0.53 | 0.29 | 0.18 |
| 35 °C | 0.51 | 0.27 | 0.22 | 0.52 | 0.32 | 0.16 |
| 45 °C | 0.48 | 0.31 | 0.21 | 0.54 | 0.28 | 0.18 |
| 55 °C | 0.50 | 0.30 | 0.20 | 0.51 | 0.30 | 0.19 |
| 65 °C | 0.51 | 0.32 | 0.17 | 0.61 | 0.18 | 0.21 |
| 75 °C | 0.52 | 0.30 | 0.18 | 0.61 | 0.21 | 0.18 |
| 85 °C | 0.55 | 0.27 | 0.18 | 0.61 | 0.24 | 0.15 |
| 95 °C | 0.54 | 0.27 | 0.19 | 0.64 | 0.22 | 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-Y.; Hsieh, M.-H.; Huang, Y.-C.; Chen, C.-J.; Lee, M.-T. Conformational Changes of α-Crystallin Proteins Induced by Heat Stress. Int. J. Mol. Sci. 2022, 23, 9347. https://doi.org/10.3390/ijms23169347
Chang Y-Y, Hsieh M-H, Huang Y-C, Chen C-J, Lee M-T. Conformational Changes of α-Crystallin Proteins Induced by Heat Stress. International Journal of Molecular Sciences. 2022; 23(16):9347. https://doi.org/10.3390/ijms23169347
Chicago/Turabian StyleChang, Yu-Yung, Meng-Hsuan Hsieh, Yen-Chieh Huang, Chun-Jung Chen, and Ming-Tao Lee. 2022. "Conformational Changes of α-Crystallin Proteins Induced by Heat Stress" International Journal of Molecular Sciences 23, no. 16: 9347. https://doi.org/10.3390/ijms23169347
APA StyleChang, Y.-Y., Hsieh, M.-H., Huang, Y.-C., Chen, C.-J., & Lee, M.-T. (2022). Conformational Changes of α-Crystallin Proteins Induced by Heat Stress. International Journal of Molecular Sciences, 23(16), 9347. https://doi.org/10.3390/ijms23169347