
Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals




Abstract
:1. Introduction
2. Naturally Occurring Heavy Metal Hyperaccumulators
2.1. Classification and Occurrence of Hyperaccumulators
2.2. Parameters for Assessing Heavy Metal Resistance
3. Characteristics and Methods of Phytoremediation
4. Molecular Basis of Natural Heavy Metal Hyperaccumulation and Hypertolerance
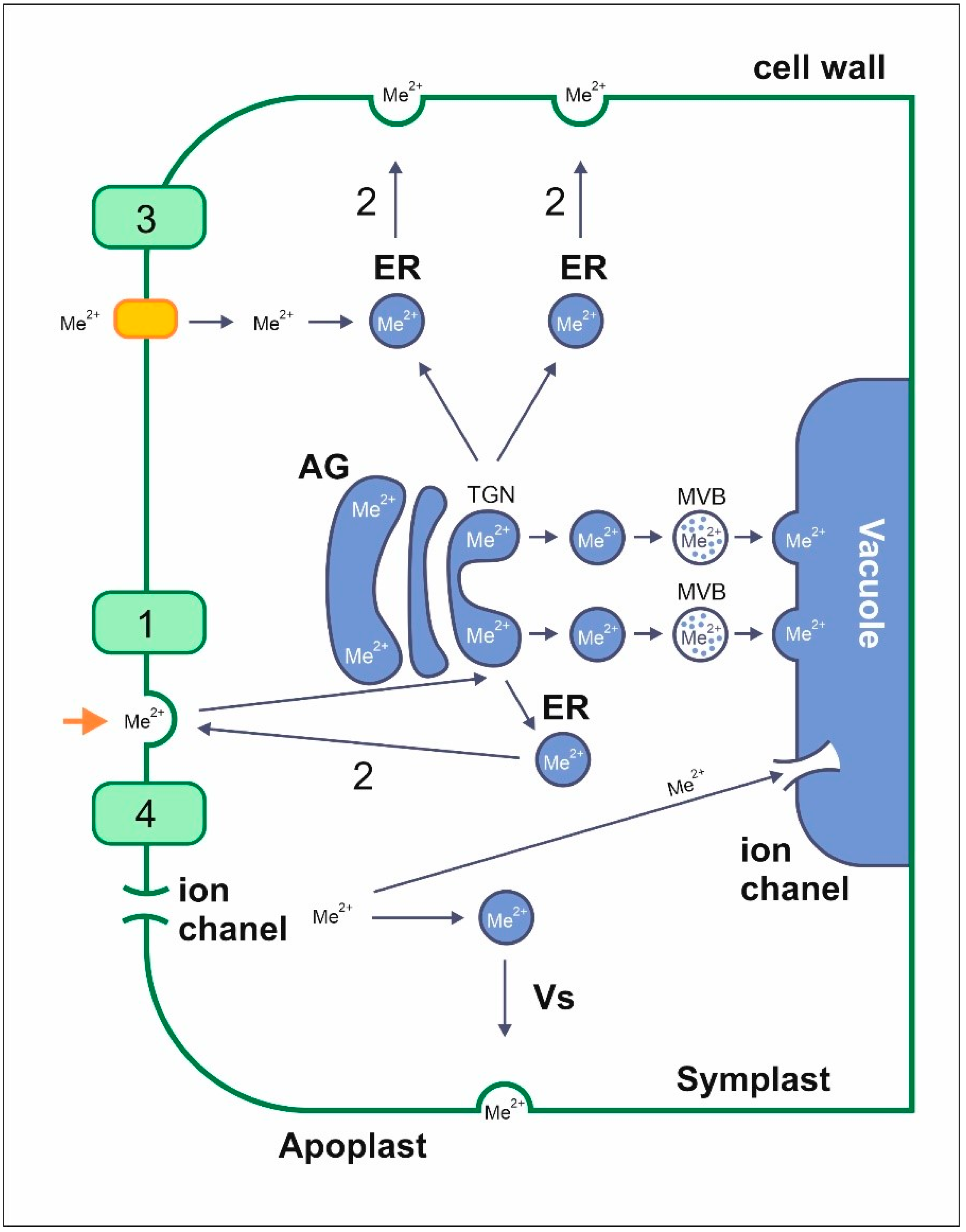
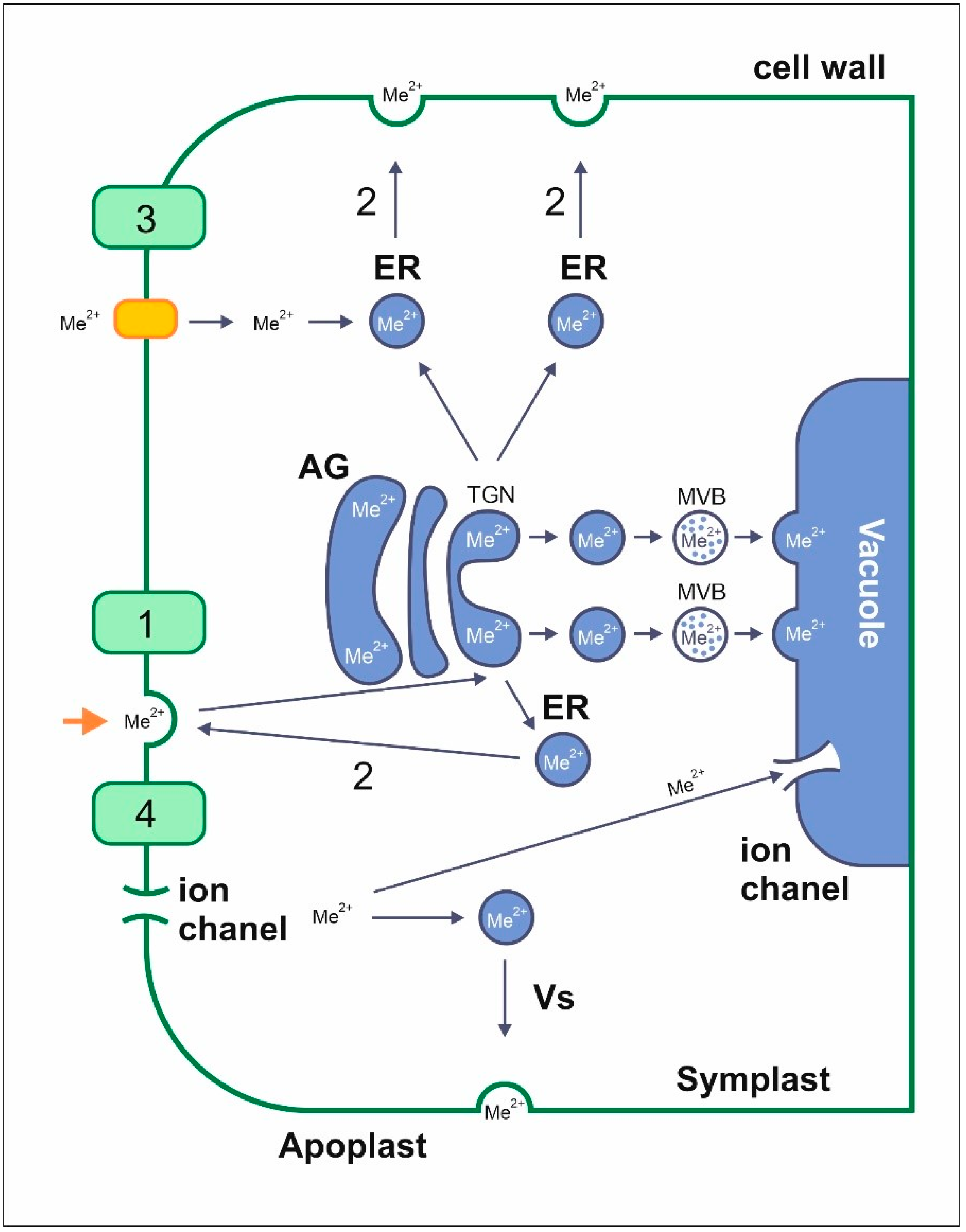
4.1. Uptake and Transport of Metals by Endocytosis
4.2. Transport of Metals to Different Plant Organs
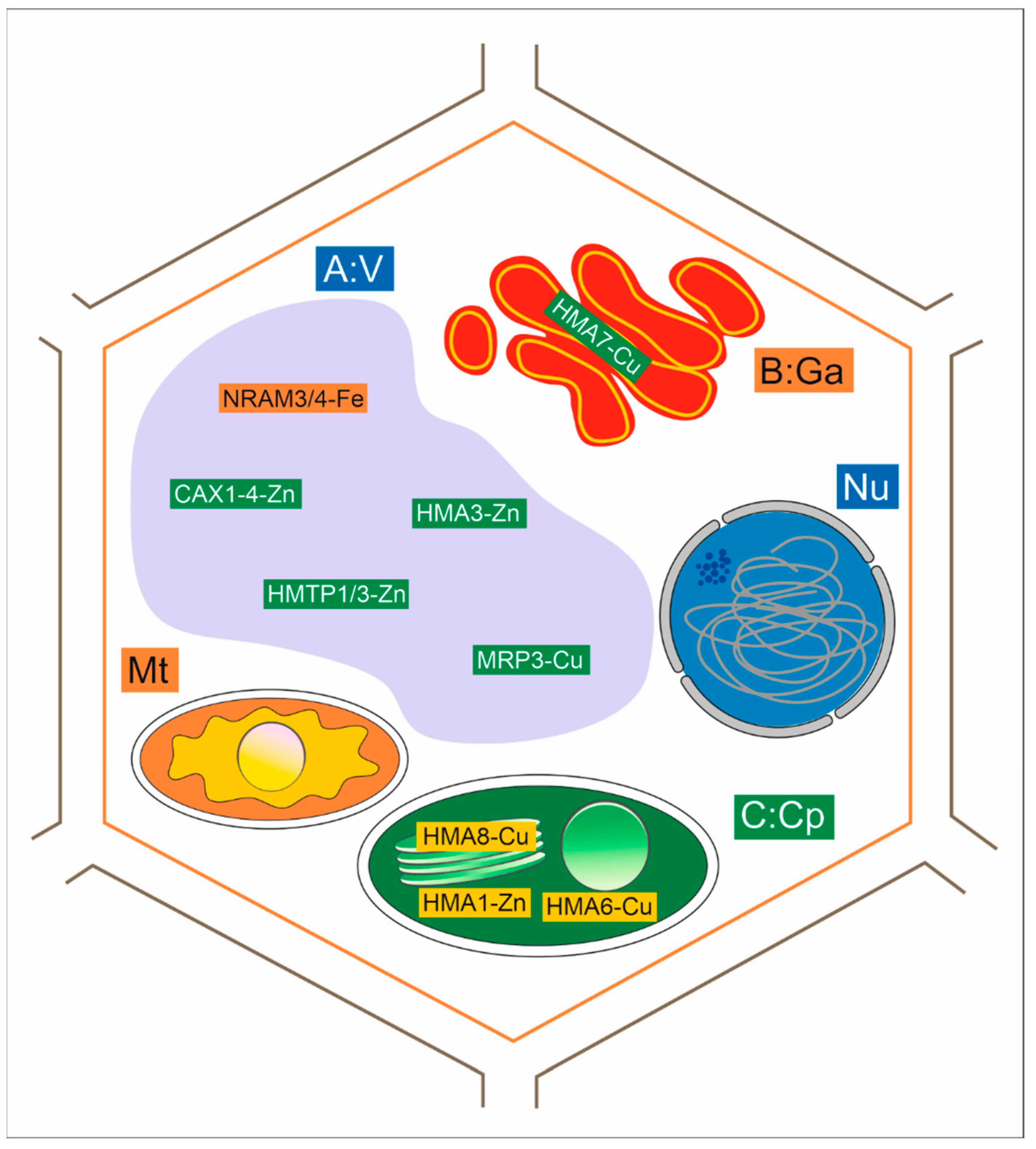
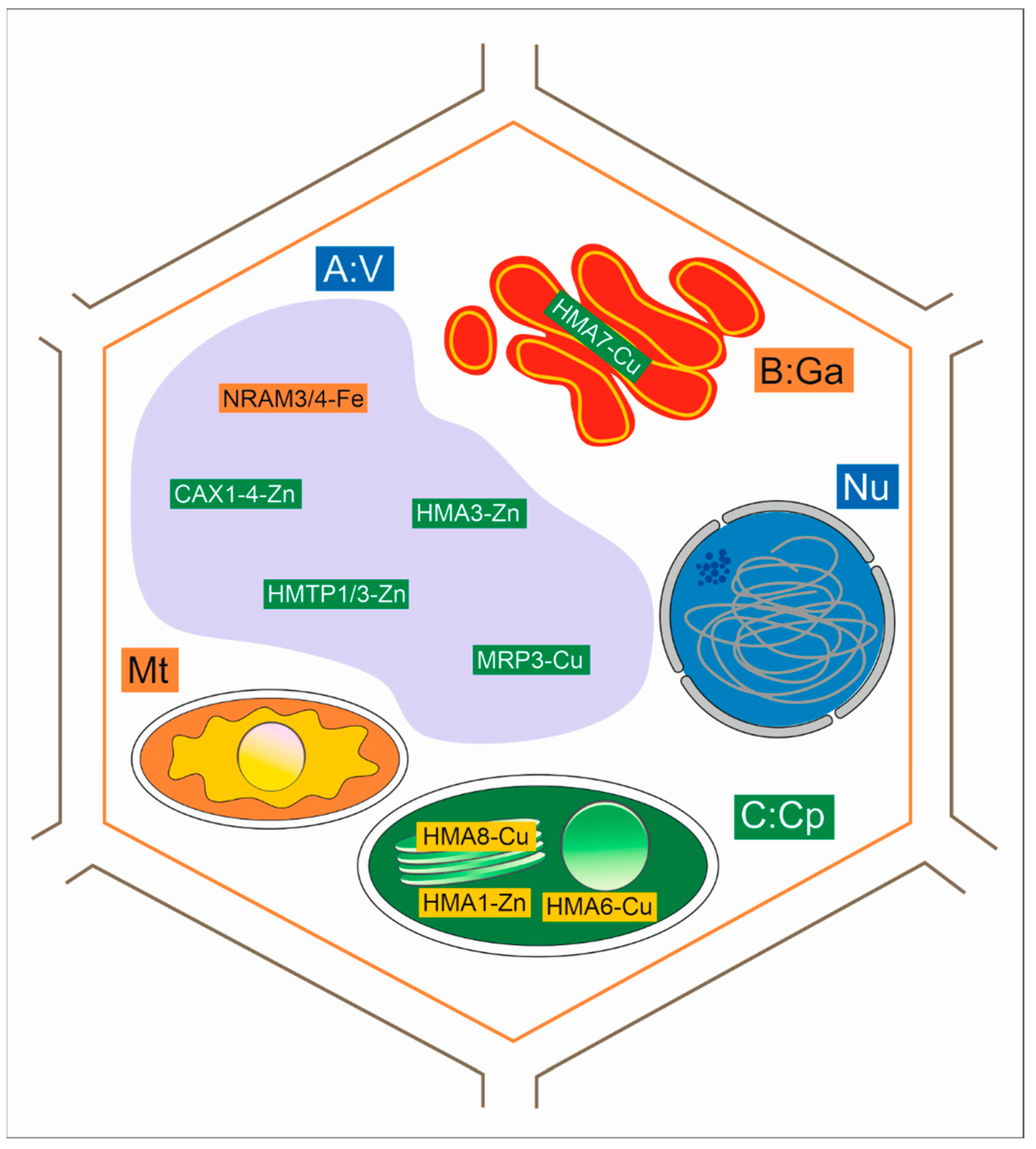
4.3. Uptake and Transport of Metals with Membrane Transporters
4.4. Strategy for Avoiding Heavy Metal Uptake
4.4.1. Symbioses with Rhizospheric Microorganisms
4.4.2. Release of Substances into the Soil and Immobilization of Metals
4.4.3. Changes in Cell Wall Permeability and Its Modifications
4.5. Strategy of Stress Tolerance
4.5.1. Chaperones
4.5.2. Chelation of Heavy Metal Ions
4.5.3. Production of Heat-Shock Proteins
4.6. Expression of Genes Associated with Heavy Metal Tolerance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Genes | Plant | Elements | References |
|---|---|---|---|
| ZIP gene family: | [106,107,108,109,110,111] | ||
| TcZNT1 | Thlaspi caerulescens | Zn | |
| AhIRT3 | Arabidopsis halleri | Fe | |
| AtZIP1-3 AtZIP5-7 AtZIP9-12 AtIRT1 AtIRT2 AtIRT3 | Arabidopsis thaliana | Fe, Mn, Cu, Zn, Cd | |
| MtZIP1 MtZIP3 MtZIP4 MtZIP5 MtZIP6 MtZIP7 | Medicago truncatula | Mn, Fe | |
| P-type ATPase (HMA) genes: | [79,112,113,114,115,116,117] | ||
| AtHMA2 AtHMA4 | Arabidopis thaliana | Zn, Cd | |
| TcHMA4 | Thlaspi caerulescens | Cd | |
| AhHMA | Arabidopsis halleri | Zn | |
| OsHMA2 | Oryza sativa | Cd, Zn | |
| HvHMA2 | Hordeum vulgare | Zn, Cd | |
| YSL gene family: | [112,117,118] | ||
| AhYSL3.1 AhYSL3.2 | Arachis hypogaea L. | Cu | |
| AtYSL2 | Arabidopsis thaliana | Cu, Fe | |
| OsYSL2 | Oryza sativa | Fe | |
| NRAMP genes: | [119,120,121,122,123] | ||
| GmNRAMP1-7 | Glycine max L. | Cd, Cu, Mn, Fe | |
| AtNRAMP1- AtNRAMP6 | Arabidopsis thaliana | Cd, Cu, Mn, Fe | |
| MtNRAMP1 | Medicago truncatula | Fe | |
| OsNRAMP3 OsNRAMP5 | Oryza sativa | Mn, Cu | |
| CDF gene family: | [124,125,126] | ||
| BrrMTP | Zn, Fe, Mn | ||
| AtMTP | Arabidopsis thaliana | Zn, Mn | |
| ShMTP | Stylosanthes hamata | Mn | |
| OsMTP11 | Oryza sativa | Mn | |
| CAX gene family: | [127,128,129,130,131] | ||
| OsCAX | Oryza sativa | Mn, Cd | |
| AtCAX2 AtCAX4 | Arabidopsis thaliana | Cd | |
| COPT gene family: | [83,132,133,134] | ||
| OsCOPT1 OsCOPT5 | Oryza sativa | Cu | |
| AtCOPT5 | Arabidopsis thaliana | Cu | |
| ABC (ATP-binding cassett) genes: | [135] | ||
| OsSTAR1 OsSTAR2 | Oryza sativa | Al, Cd | |
| YCF1 | Brassica juncea | Cd | |
| AtATM3 | Arabidopsis thaliana | Cd | |
| IREG genes: | [136,137] | ||
| FeIREG1 | Fagopyrum esculentum Moench | Al | |
| AtIREG1 AtIREG3 | Arabidopsis thaliana | Fe, Co |
| Source/Target Transgenic; Plants | Genes | Main Tissue Expression | Response in Plants | References |
|---|---|---|---|---|
| Brassica jouncea/Nicotiana tabacum | CAT; CAT3 | roots | The accumulation of the heavy metal is markedly higher in the roots than in the leaves. This lower accumulation may explain the absence of Cd2+ genotoxicity in leaves. | [138,139] |
| Brassica oleracea/Arabidopsis | CAT1; CAT2 | roots | The results demonstrate that overexpression of BoCAT1 or BoCAT2 could reduce the phytotoxicity of H2O2 caused by high temperature in Arabidopsis. Transgenic Arabidopsis plants exhibited higher levels of RNA and CAT. This is the first report suggesting that CAT-encoding gene expression in Arabidopsis is regulated by heat stress. | [140] |
| Zea mays/Brassica campestris | Cu/ZnSOD and/or CAT | roots, shoots | Under SO2 stress less reduction in photosynthetic activity than wild type. | [141] |
| Festuca arundinacea | Cu/ZnSOD and APX | leaves | The mechanisms of increased antioxidative defense in transgenic tall fescue plants is the overexpression of the CuZnSOD and APX genes, which are utilized in scavenging ROS and thus provide improved tolerance to abiotic stresses. | [142] |
| Triticum aestivum/Brasica napus | MnSOD | roots, shoots | Observed 1·5- to 2·5-fold increase in total SOD activity in transgenic B. napus plants increased oxidative resistance compared with the wild-type plants. | [143] |
| Brassica rapa/Escherichia coli | GR; BrGR | - | GR participates in protection against oxidation by maintaining the adequate redox state in the intracellular environment and, thus, regulating various cellular activities Antioxidant capacity of BrGR protein can be examined by the induction of cell protection via the introduction of a variety of stressors, including hydrogen peroxide, menadione, or heavy metals. | [144] |
| Escherichia coli/Nicotiana tabacum | DHAR/GR/GST | leaves | Overexpression of these different enzymes enhanced salt and cold tolerance. In leaves from the DHAR plants, the level of DHAR activity increased between 1.8- and 2.7-fold when compared to the wild type. The n leaves from the DHAR:GR plants, this increase was found to be greater, between 3.2- and 4-fold. Leaves of the GST plants exhibited an increase in GST activity of approximately 2-fold, while this increase was greater, from 2.9- to 3.7-fold in the GST:GR double transformants | [145] |
| Arabidopsis thaliana/Nicotiana tabacum | MDHAR/DHAR | roots | DHAR but not MDHAR enhanced Al tolerance by maintaining the ascorbate level. | [146] |
| Thlaspi caerulescens | ZIP | roots, shoots | Genes are Zn regulated and Cd influx is mainly due to Zn transporters having strong preference for Zn over Cd. Zn uptake is due to overexpression of genes belonging to the ZIP stunted growth, chlorosis, leaf curling, and death of leaf tips. | [65,147] |
| Mesembryanthemum crystallinum | ZIP4, IRT2, CAX4, HMA4, PCS1 | roots, shoots | Expression of the root genes IRT2, CAX4, HMA4, PCS1, and ZIP4 salinity Cd stress-enhanced. For IRT2 and PCS1, a cumulative effect of both stressors on gene expression was found. The salt-stressed plants subjected to a 1 mM concentration accumulated more Cd compared to the NaCl-untreated plants, and the heavy metal was stored mainly in the roots. Interestingly, this relationship was also maintained under 10 mM Cd treatment, where the salt-stressed plant roots accumulated almost 2-fold more Cd in comparison to the roots of NaCl-untreated plants. The elevated Cd amounts were deposited in shoots of NaCl-untreated and salt-stressed plants only under a 10 mM concentration. | [148] |
| Mesembryanthemum crystallinum | IRT2 | roots | For IRT2, a cumulative effect of both stressors on gene expression was found. Moreover, the role of salinity stress as an upstream regulator in the halophyte IRT2 expression scheme was suggested. | [149] |
| Sedum alfredii/Arabidopsis thaliana | ZIP4 | roots, shoots | Transgenic Arabidopsis thaliana mutant ZIP4-2-expressing SaZIP4h reversed the Zn/Cd uptake defect, and wild-type A. thaliana ectopically overexpressing SaZIP4h displayed increased Zn accumulation both in roots and shoots. Together, these results suggest that SaZIP4 is an important Zn uptake transporter that takes up Zn in the roots and shoots of S. alfredii. | [150] |
| Oryza sativa | NRAMP1 NRAMP5 | roots, shoots | Detoxification/sequestration of heavy metal. Revealed their role in uptake and transport of Cd Mn, and Fe. Inhibits plant growth. | [93] |
| Oryza sativa | NRAMP1 | roots, shoots | These results suggest that OsNRAMP1 participates in cellular Cd uptake and Cd transport within plants, and the higher expression of OsNRAMP1 in the roots could lead to an increase in Cd accumulation in the shoots. | [151] |
| Oryza sativa | LCT2 | roots, shoots | When grown in Cd-contaminated paddy soils, rice plants overexpressing OsLCT2 significantly reduced Cd concentrations in the straw and grains. OSLCT2 overexpression decreased the rate of Cd translocation from roots to shoots, and reduced Cd concentrations in xylem sap and in shoots of rice. Overexpression of OsLCT2 reduces Cd accumulation in rice shoots and grains by limiting the amounts of Cd loaded into the xylem and restricting Cd translocation from roots to shoots of rice. | [152] |
| Arabidopsis halleri | FDR3 | roots, shoots | Root-to-shoot translocation of heavy metals. The high expression of FRD3 in A. halleri contributes to metal homeostasis, but not specifically to the high accumulation of Zn in shoots of A. halleri. | [153] |
| Lycopersicon esculentum Mill | GRP | roots | LeGRP transcripts predominately accumulated in roots at different developmental stages, but not in leaves or ripe fruit tissues, and their levels declined gradually during plant development. | [154] |
| Thlaspi arvense | ZNT1 ZNT2 | roots, leaves | T. arvense ZNT1 transcript was only detected in roots and leaves of plants grown at 0 mm Zn. Moreover, ZNT2 is expressed only at 0 mm Zn, although it is barely detectable after hybridization. Under these conditions, and especially in roots, the expression is much lower in T. arvense than in T. caerulescens. | [155] |
| Thlaspi caerulescens | ZNT1 ZNT2 | roots, shoots | The expression in T. caerulescensis was barely Zn-responsive, suggesting that Zn hyperaccumulation might rely on a decreased Zn-induced transcriptional downregulation of these genes. | [156] |
| Noccaea caerulescens | ZNT1 | shoots | The orthologue of the A. thaliana AtZIP4 gene. Their conclusion is that NcZNT1 plays an important role in Zn and Cd tolerance and accumulation and is involved in establishing a high metal influx into the root vasculature, important for xylem-mediated translocation of metals to the shoot. | [157] |
| Medicago truncatula | MTP1 | vegetative organs | The expression of MtMTP1 was detected in all vegetative organs with the highest level of expression observed in leaves. | [158] |
| Arabidopsis halleri | MTP1 | leaves—high roots—low shoots—completely abolished | The high level of expression in leaves and completely abolished in the shoots of the plants, but weak staining, was observed in the roots. | [159] |
| Mus musculus/Nicotiana tabacum | MTP1 | roots, stalk and leaves of tissues | The expression of mouse metallothionein in transplastomic plants increases mercury resistance, accumulation, and phytoremediation by the mechanism of chelation. The high level of expression in leaves of tissues. | [160] |
References
- Appenroth, K.J. What are “heavy metals” in plant sciences? Acta Physiol. Plant. 2010, 32, 615–619. [Google Scholar] [CrossRef]
- Szarek-Łukaszewska, G. Heavy metals hypperaccumulating plants. Kosmos 2014, 63, 443–453. [Google Scholar]
- Tibbett, M.; Green, I.; Rate, A.; De Oliveira, V.H.; Whitaker, J. The transfer of trace metals in the soil-plant-arthropod system. Sci. Total Environ. 2021, 779, 141260. [Google Scholar] [CrossRef]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation in hyperaccumulator plants. Physiol. Plant. 2021, 173, 148–166. [Google Scholar] [CrossRef] [PubMed]
- Skorbiłowicz, M.; Skorbiłowicz, E.; Rogowska, W. Heavy Metal Concentrations in Roadside Soils on the Białystok-Budzisko Route in Northeastern Poland. Minerals 2021, 11, 1290. [Google Scholar] [CrossRef]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Hanus-Fajerska, E.; Muszyńska, E.; Giemzik, A. Review on studies of zinc-lead waste heaps microbiota. Arch. Waste Manag. Environ. Prot. 2015, 17, 59–68. [Google Scholar]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E. Heavy metals in contaminated soils: A review of sources, chemistry, risks and best available strategies for remediation. Int. Sch. Res. Not. 2011, 2021, 402647. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and Metalloid Toxicity in Plants: An Overview on Molecular Aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta. Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef]
- Jan, S.; Parray, J.A. Metal Tolerance Strategy in Plants. In Approaches to Heavy Metal Tolerance in Plants; Springer: Singapore, 2016; pp. 19–32. [Google Scholar] [CrossRef]
- Tao, J.; Lu, L. Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants. Toxics 2022, 10, 411. [Google Scholar] [CrossRef] [PubMed]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, S.; Mathur, V.; Garg, A.; Lenka, S.; Verma, K.; Agarwal, S. Molecular mechanisms underlying heavy metal uptake, translocation and tolerance in hyperaccumulators—An analysis: Heavy metal tolerance in hyperaccumulators. Environ. Chall. 2021, 4, 100197. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Kicińska, A.; Gruszecka-Kosowska, A. Long-term changes of metal contents in two metallophyte species (Olkusz area of Zn-Pb ores, Poland). Environ. Monit. Assess. 2016, 188, 339. [Google Scholar] [CrossRef]
- Van der Pas, L.; Ingle, R.A. Towards an Understanding of the Molecular Basis of Nickel Hyperaccumulation in Plants. Plants 2019, 8, 11. [Google Scholar] [CrossRef]
- Jaffré, T.; Pillon, Y.; Thomine, S.; Merlot, S. The metal hyperaccumulators from New Caledonia can broaden our understanding of nickel accumulation in plants. Front. Plant Sci. 2013, 4, 279. [Google Scholar] [CrossRef]
- Van der Ent, A.; Baker, A.J.M.; Echevarria, G.; Simonnot, M.-O.; Morel, J.L. Agromining: Farming for Metals: Extracting Unconventional Resources Using Plants; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Lombi, E.; Zhao, F.J.; Dunham, S.J.; McGrath, S.P. Phytoremediation of Heavy Metal–Contaminated Soils: Natural Hyperaccumulation versus Chemically Enhanced Phytoextraction. J. Environ. Qual. 2001, 30, 1919–1926. [Google Scholar] [CrossRef]
- Van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil. 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Dradrach, A.; Karczewska, A.; Szopka, K.; Lewińska, K. Accumulation of Arsenic by Plants Growing in the Sites Strongly Contaminated by Historical Mining in the Sudetes Region of Poland. Int. J. Environ. Res. Public Health 2020, 17, 3342. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, B.; Szulc, W.; Łabętowicz, J.; Pikuła, D. Bioaccumulation index and contamination index as criteria for assessment of accumulation of copper and zinc in biomass of pasture plants. Fresenius Environ. Bull. 2010, 19, 620–623. [Google Scholar]
- Hołtra, A.; Zamorska-Wojdyła, D. The pollution indices of trace elements in soils and plants close to the copper and zinc smelting works in Poland’s Lower Silesia. Environ. Sci. Pollut. Res. 2020, 27, 16086–16099. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, E.; Hanus-Fajerska, E. Why are heavy metal hyperaccumulating plants so amazing? BioTechnologia 2015, 96, 265–271. [Google Scholar] [CrossRef]
- Khalid, S.; Shahid, M.; Niazi, N.K.; Murtaza, B.; Bibi, I.; Dumat, C. A comparison of technologies for remediation of heavy metal contaminated soils. J. Geochem. Explor. 2016, 182, 247–268. [Google Scholar] [CrossRef]
- Dixit, R.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.P.; Rai, J.P.; Sharma, P.K.; Lade, H.; et al. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef]
- Smolińska, B. The Methods of Cleaning Soil Polluted by Mercury. Scient. Bull. Tech. Univ. Lodz. 2010, 74, 121–136. [Google Scholar]
- Siwek, M. Phytoremediation as a biological method of cleaning up the environment. Pol. Bot. Soc. 2008, 52, 23–38. [Google Scholar]
- Grobelak, A.; Kacprzak, M.; Fijałkowski, K. Phytoremediation the underestimated potential of plants in cleaning up the environment. J. Ecol. Health 2010, 14, 276–280. [Google Scholar]
- Pulford, I.; Watson, C. Phytoremediation of heavy metal-contaminated land by trees—A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef]
- Volk, T.A.; Abrahamson, L.P.; Nowak, C.A.; Smart, L.B.; Tharakan, P.J.; White, E.H. The development of short-rotation willow in the northeastern United States for bioenergy and bioproducts, agroforestry and phytoremediation. Biomass Bioenergy 2006, 30, 715–727. [Google Scholar] [CrossRef]
- Ruttens, A.; Boulet, J.; Weyens, N.; Smeets, K.; Adriaensen, K.; Meers, E.; van Slycken, S.; Tack, F.; Meiresonne, L.; Thewys, T.; et al. Short rotation coppice culture of willows and poplars as energy crops on metal contaminated agricultural soils. Int. J. Phytorem. 2011, 13, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yu, N.; Wang, L.; Gupta, D.K.; He, Z.; Wang, K.; Zhu, Z.; Yan, X.; Li, T.; Yang, X.E. The phytoremediation potential of bioenergy crop Ricinus communis for DDTs and cadmium co-contaminated soil. Bioresour. Technol. 2011, 102, 11034–11038. [Google Scholar] [CrossRef]
- Wei, S.H.; Zhou, Q.X.; Wang, X. Cadmium-hyperaccumulator Solanum nigrum L. and its accumulating characteristics. Environ. Sci. 2005, 26, 167–171. [Google Scholar]
- Meers, E.; van Slycken, S.; Adriaensen, K.; Ruttens, A.; Vangronsveld, J.; Du Laing, G.; Witters, N.; Thewys, T.; Tack, F.M. The use of bio-energy crops (Zea mays) for “phytoattenuation” of heavy metals on moderately contaminated soils: A field experiment. Chemosphere 2010, 78, 35–41. [Google Scholar] [CrossRef]
- Bittsanszkya, A.; Kömives, T.; Gullner, G.; Gyulai, G.; Kiss, J.; Heszky, L.; Radimszky, L.; Rennenberg, H. Ability of transgenic poplars with elevated glutathione content to tolerate zinc(2+) stress. Environ. Int. 2005, 31, 251–254. [Google Scholar] [CrossRef]
- Jamil, S.; Abhilash, P.C.; Singh, N.; Sharma, P.N. Jatropha curcas: A potential crop for phytoremediation of coal fly ash. J. Hazard. Mater. 2009, 172, 269–275. [Google Scholar] [CrossRef]
- Adelodun, A.A.; Afolabi, N.O.; Chaúque, E.F.C.; Akinwumiju, A.S. The potentials of Eichhornia crassipes for Pb, Cu, and Fe removal from polluted waters. SN Appl. Sci. 2020, 2, 1646. [Google Scholar] [CrossRef]
- Che, D.; Meagher, R.B.; Heaton, A.C.; Lima, A.; Rugh, C.L.; Merkle, S.A. Expression of mercuric ion reductase in Eastern cottonwood (Populus deltoides) confers mercuric ion reduction and resistance. Plant Biotechnol. J. 2003, 1, 311–319. [Google Scholar] [CrossRef]
- Tong, X.; Yuan, L.; Luo, L.; Yin, X. Characterization of a selenium-tolerant rhizosphere strain from a novel Se-hyperaccumulating plant Cardamine hupingshanesis. Sci. World J. 2014, 2014, 108562. [Google Scholar] [CrossRef] [PubMed]
- Nematian, M.A.; Kazemeini, F. Accumulation of Pb, Zn, Cu and Fe in plants and hyperaccumulator choice in Galali iron mine area, Iran. Int. J. Agric. Crop Sci. 2013, 5, 426–432. [Google Scholar]
- Bani, A.; Pavlova, D.; Echevarria, G.; Mullaj, A.; Reeves, R.D.; Morel, J.L.; Sulce, S. Nickel hyperaccumulation by the species of Alyssum and Thlaspi (Brassicaceae) from the ultramafic soils of the Balkans. Bot. Serbica 2010, 34, 3–14. [Google Scholar]
- Yadav, R.; Singh, S.; Kumar, A.; Sing, A.N. Phytoremediation: A wonderful cost-effective tool. In Advances in Environmental Pollution Research. Cost Effective Technologies for Solid Waste and Wastewater Treatment; Kathi, S., Devipriya, S., Thamaraiselvi, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 179–208. [Google Scholar] [CrossRef]
- Conesa, H.M.; Faz, A.; Arnaldos, R. Heavy metal accumulation and tolerance in plants from mine tailings of the semiarid Cartagena-La Union Mining District (Se Spain). Sci. Total Environ. 2006, 366, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.C.; Gonzalez-Chavez, M.C.A. Metal accumulation in wild plants surrounding mining wastes: Soil and sediment remediation (SSR). Environ. Pollut. 2006, 144, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.O.; Glenn, E.P.; Maier, R.M. Phytostabilization potential of quailbush for mine tailings: Growth, metal accumulation, and microbial community changes. J. Environ. Qual. 2007, 36, 245–253. [Google Scholar] [CrossRef]
- Ahalya, N.; Ramachandra, T.V. Phytoremediation: Processes and mechanisms. J. Ecobiol. 2006, 18, 33–38. [Google Scholar]
- Suman, J.; Uhlik, O.; Viktorova, J.; Macek, T. Phytoextraction of heavy metals: A Promising Tool for Clean-Up of Polluted Environment? Front. Plant Sci. 2018, 9, 1476. [Google Scholar] [CrossRef]
- Asgari Lajayer, B.; Moghadam, N.K.; Maghsoodi, M.R.; Ghorbanpour, M.; Kariman, K. Phytoextraction of heavy metals from contaminated soil, water and atmosphere using ornamental plants: Mechanisms and efficiency improvement strategies. Environ. Sci. Pollut. Res. 2019, 26, 8468–8484. [Google Scholar] [CrossRef]
- Lasat, M.M. Phytoextraction of toxic metals: A review of biological mechanisms. J. Environ. Qual. 2002, 31, 109–120. [Google Scholar] [CrossRef]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Role of hyperaccumulators in phytoextraction of metals from contaminated mining sites: A Review. Crit. Rev. Environ. Sci. Technol. 2011, 41, 168–214. [Google Scholar] [CrossRef]
- Chatterjee, S.; Mitra, A.; Datta, S.; Veer, V. Phytoremediation Protocols: An Overview. In Plant-Based Remediation Processes. Soil Biology; Gupta, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; p. 35. [Google Scholar] [CrossRef]
- Rugh, C.L. Mercury detoxification with transgenic plants and other biotechnological breakthroughs for phytoremediation. In Vitro Cell. Dev. Biol. Plant. 2001, 37, 321–325. [Google Scholar] [CrossRef]
- Muthusaravanan, S.; Sivarajasekar, N.; Vivek, J.S.; Paramasivan, T.; Naushad, M.; Prakashmaran, J.; Gayathri, V.; Al-Duaij, O.K. Phytoremediation of heavy metals: Mechanisms, methods and enhancements. Environ. Chem. Lett. 2018, 16, 1339–1359. [Google Scholar] [CrossRef]
- Arthur, E.L.A.; Rice, P.J.; Rice, P.J.; Anderson, T.A.; Baladi, S.A.; Henderson, K.L.D.; Coats, J.R. Phytoremediation—An Overview. Crit. Rev. Plant Sci. 2005, 24, 109–122. [Google Scholar] [CrossRef]
- Bolan, N.S.; Park, J.H.; Robinson, B.; Naidu, R.; Huh, K.Y. Phytostabilization: A Green Approach to Contaminant Containment. Adv. Agron. 2011, 112, 145–204. [Google Scholar] [CrossRef]
- Montiel-Rozas, M.M.; Madejón, E.; Madejón, P. Effect of heavy metals and organic matter on root exudates (low molecular weight organic acids) of herbaceous species: An assessment in sand and soil conditions under different levels of contamination. Environ. Pollut. 2016, 216, 273–281. [Google Scholar] [CrossRef]
- Pajević, S.; Borišev, M.; Nikolić, N.; Arsenov, D.D.; Orlović, S.; Župunski, M. Phytoextraction of Heavy Metals by Fast-Growing Trees: A Review. In Phytoremediation; Ansari, A., Gill, S., Gill, R., Lanza, G., Newman, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Singh, O.V.; Jain, R.K. Phytoremediation of toxic aromatic pollutants from soil. Appl. Microbiol. Biotechnol. 2003, 63, 128–135. [Google Scholar] [CrossRef]
- Nouri, J.; Khorasani, N.; Lorestani, B.; Karami, M.; Hassani, A.H.; Yousefi, N. Accumulation of heavy metals in soil and uptake by plant species with phytoremediation potential. Environ. Earth Sci. 2009, 59, 315–323. [Google Scholar] [CrossRef]
- Taylor, G.; Crowder, A. Copper and nickel tolerance in Typha latifolia clones from contaminated and uncontaminated environments. Can. J. Bot. 2011, 62, 1304–1308. [Google Scholar] [CrossRef]
- Hasan, M.K.; Cheng, Y.; Kanwar, M.K.; Chu, X.-Y.; Ahammed, G.J.; Qi, Z.-Y. Responses of Plant Proteins to Heavy Metal Stress—A Review. Front. Plant Sci. 2017, 8, 1492. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Barea, J.M.; Pozo, M.J.; Azcón, R.; Azcón-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Gorbushina, A.A.; Krumbein, W. Role of Microorganisms in Wear Down of Rocks and Minerals. In Microorganisms in Soils: Roles in Genesis and Functions. Soil Biology; Varma, A., Buscot, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar] [CrossRef]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.; Anli, M.; Meddich, A.; Oufdou, K. Use of Rhizobacteria and Mycorrhizae Consortium in the Open Field as a Strategy for Improving Crop Nutrition, Productivity and Soil Fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef]
- Policarpo Tonelli, F.C.; Policarpo Tonelli, F.M.; Lemos, M.S.; de Melo Nunes, N.A. Mechanisms of phytoremediation. Phytoremediation 2022, 37–64. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead Toxicity in Plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Tangahu, B.V.; Abdullah, S.R.S.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M. A Review on Heavy Metals (As, Pb, and Hg) Uptake by Plants through Phytoremediation. Int. J. Chem. Eng. 2011, 2011, 939161. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Gautam, S.; Landi, M.; Kandhol, N.; Araniti, F.; Ramakrishnan, M.; Satish, L.; Singh, V.P.; Sharma, P.; et al. Heavy metal induced regulation of plant biology: Recent insights. Physiol. Plant. 2022, 174, e13688. [Google Scholar] [CrossRef]
- Horst, W.J.; Wang, Y.; Eticha, D. The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: A review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef]
- Hu, Y.T.; Ming, F.; Chen, W.W.; Yan, J.Y.; Xu, Z.Y.; Li, G.X.; Xu, C.Y.; Yang, J.L.; Zheng, S.J. TcOPT3, a member of oligopeptide transporters from the hyperaccumulator Thlaspi caerulescens, is a novel Fe/Zn/Cd/Cu transporter. PLoS ONE 2012, 7, e38535. [Google Scholar] [CrossRef]
- Kerkeb, L.; Krämer, U. The role of free histidine in xylem loading of nickel in Alyssum lesbiacum and Brassica juncea. Plant Physiol. 2003, 131, 716–724. [Google Scholar] [CrossRef]
- Colangelo, E.P.; Guerinot, M.L. Put the metal to the petal: Metal uptake and transport throughout plants. Curr. Opin. Plant Biol. 2006, 9, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Hanikenne, M.; Talke, I.N.; Haydon, M.J.; Lanz, C.; Nolte, A.; Motte, P.; Kroymann, J.; Weigel, D.; Krämer, U. Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4. Nature 2008, 453, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Pandey, A.; Pandey, G.K. Calcium homeostasis: Role of CAXs transporters in plant signaling. Plant Stress 2012, 6, 60–69. [Google Scholar]
- Cailliatte, R.; Schikora, A.; Briat, J.F.; Mari, S.; Curie, C. Hight affinity manganese uptake by the metal transporter NRAMP1 is essential for Arabidopsis growth in low manganese conditions. Plant Cell. 2010, 22, 904–917. [Google Scholar] [CrossRef]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef]
- Puig, S. Function and Regulation of the Plant COPT Family of High-Affinity Copper Transport Proteins. Adv. Bot. 2014, 2014, 476917. [Google Scholar] [CrossRef]
- Cai, Z.; Xian, P.; Lin, R.; Cheng, Y.; Lian, T.; Ma, Q.; Nian, H. Characterization of the Soybean GmIREG Family Genes and the Function of GmIREG3 in Conferring Tolerance to Aluminum Stress. Int. J. Mol. Sci. 2020, 21, 497. [Google Scholar] [CrossRef]
- Lane, T.S.; Rempe, C.S.; Davitt, J.; Staton, M.E.; Peng, Y.; Soltis, D.E.; Melkonian, M.; Deyholos, M.; Leebens-Mack, J.H.; Chase, M.; et al. Diversity of ABC transporter genes across the plant kingdom and their potential utility in biotechnology. BMC Biotechnol. 2016, 16, 47. [Google Scholar] [CrossRef]
- Checcucci, A.; Marchetti, M. The Rhizosphere Talk Show: The Rhizobia on Stage. Front. Agron. 2020, 2020, 591494. [Google Scholar] [CrossRef]
- Colpaert, J.V.; Van Assche, J.A. Zinc toxicity in ectomycorrhizal Pinus sylvestris. Plant Soil. 1992, 143, 201–211. [Google Scholar] [CrossRef]
- Hagerberg, D.; Manique, N.; Brandt, K.K.; Larsen, J.; Nybroe, O.; Olsson, S. Low Concentration of Copper Inhibits Colonization of Soil by the Arbuscular Mycorrhizal Fungus Glomus Intraradices and Changes the Microbial Community Structure. Microb. Ecol. 2011, 61, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.J.; Evans, D.E. Aluminium-silicon interactions in higher plants: An update. J. Exp. Bot. 2020, 71, 6719–6729. [Google Scholar] [CrossRef]
- Wang, P.; Yang, B.; Wan, H.; Fang, X.; Yang, C. The differences of cell wall in roots between two contrasting soybean cultivars exposed to cadmium at young seedlings. Environ. Sci. Pollut. Res. 2018, 25, 29705–29714. [Google Scholar] [CrossRef] [PubMed]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.F.; Aarts, M.G. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2015, 6, 1143. [Google Scholar] [CrossRef]
- Jain, S.; Muneer, S.; Guerriero, G.; Liu, S.; Vishwakarma, K.; Chauhan, D.K.; Dubey, N.K.; Tripathi, D.K.; Sharma, S. Tracing the role of plant proteins in the response to metal toxicity: A comprehensive review. Plant Signal. Behav. 2018, 13, e1507401. [Google Scholar] [CrossRef]
- Attallah, C.V.; Welchen, E.; Gonzalez, D.H. The promoters of Arabidopsis thaliana genes AtCOX17-1 and -2, encoding a copper chaperone involved in cytochrome c oxidase biogenesis, are preferentially active in roots and anthers and induced by biotic and abiotic stress. Physiol. Plant. 2006, 129, 123–134. [Google Scholar] [CrossRef]
- Herbette, S.; Taconnat, L.; Hugouvieux, V.; Piette, L.; Magniette, M.-L.M.; Cuine, S.; Auroy, P.; Richaud, P.; Forestier, C.; Bourguignon, J.; et al. Genome-wide transcriptome profiling of the early cadmium response of Arabidopsis roots and shoots. Biochimie 2006, 88, 1751–1765. [Google Scholar] [CrossRef]
- Contreras, R.A.; Pizarro, M.; Köhler, H.; Sáez, C.A.; Zúñiga, G.E. Copper stress induces antioxidant responses and accumulation of sugars and phytochelatins in Antarctic Colobanthus quitensis (Kunth) Bartl. Biol. Res. 2018, 51, 48. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic Stresses: General Defenses of Land Plants and Chances for Engineering Multistress Tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef]
- Çelik, Ö.; Akdaş, E.Y. Tissue-specific transcriptional regulation of seven heavy metal stress-responsive miRNAs and their putative targets in nickel indicator castor bean (R. communis L.) plants. Ecotoxicol. Environ. Saf. 2019, 170, 682–690. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, Z.; Zhu, C. Microarray-based analysis of cadmium-responsive microRNAs in rice (Oryza sativa). J. Exp. Bot. 2011, 62, 3563–3573. [Google Scholar] [CrossRef]
- Song, Z.; Pan, F.; Lou, X.; Wang, D.; Yang, C.; Zhang, B.; Zhang, H. Genome-wide identification and characterization of Hsp70 gene family in Nicotiana tabacum. Mol. Biol. Rep. 2019, 46, 1941–1954. [Google Scholar] [CrossRef]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Han, Y.; Fan, T.; Zhu, X.; Wu, X.; Ouyang, J.; Jiang, L.; Cao, S. WRKY12 represses GSH1 expression to negatively regulate cadmium tolerance in Arabidopsis. Plant Mol. Biol. 2019, 99, 149–159. [Google Scholar] [CrossRef]
- Cevher-Keskin, B.; Yıldızhan, Y.; Yüksel, B.; Dalyan, E.; Memon, A.R. Characterization of differentially expressed genes to Cu stress in Brassica nigra by Arabidopsis genome arrays. Environ. Sci. Pollut. Res. 2019, 26, 299–311. [Google Scholar] [CrossRef]
- Milner, M.J.; Kochian, L.V. Investigating heavy-metal hyperaccumulation using Thlaspi caerulescens as a model system. Ann. Bot. 2008, 102, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, V.; Lo, J.C.; Wu, C.L.; Wang, S.L.; Lai, C.C.; Connolly, E.L.; Huang, J.L.; Yeh, K.-C. Differential expression and regulation of iron-regulated metal transporters in Arabidopsis halleri and Arabidopsis thaliana—The role in zinc tolerance. New Phytol. 2011, 190, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Assunção, A.G.; Herrero, E.; Lin, Y.F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.H.; van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Trampczynska, A.; Clemens, S. Comparative transcriptome analysis of toxic metal responses in Arabidopsis thaliana and the Cd2+-hypertolerant facultative metallophyte Arabidopsis halleri. Plant Cell. Environ. 2006, 29, 950–963. [Google Scholar] [CrossRef] [PubMed]
- López-Millán, A.F.; Ellis, D.R.; Grusak, M.A. Identification and characterization of several new members of the ZIP family of metal ion transporters in Medicago truncatula. Plant. Mol. Biol. 2004, 54, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef]
- Dai, J.; Wang, N.; Xiong, H.; Qiu, W.; Nakanishi, H.; Kobayashi, T.; Nishizawa, N.K.; Zuo, Y. The Yellow Stripe-Like (YSL) Gene Functions in Internal Copper Transport in Peanut. Genes 2018, 9, 635. [Google Scholar] [CrossRef]
- Papoyan, A.; Kochian, L.V. Identification of Thlaspi caerulescens genes that may be involved in heavy metal hyperaccumulation and tolerance. Characterization of a novel heavy metal transporting ATPase. Plant Physiol. 2004, 136, 3814–3823. [Google Scholar] [CrossRef]
- Kajala, K.; Walker, K.L.; Mitchell, G.S.; Krämer, U.; Cherry, S.R.; Brady, S.M. Real-time whole-plant dynamics of heavy metal transport in Arabidopsis halleri and Arabidopsis thaliana by gamma-ray imaging. Plant Direct. 2019, 3, e00131. [Google Scholar] [CrossRef]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Feng, M.J. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef]
- Mills, R.F.; Peaston, K.A.; Runions, J.; Williams, L.E. HvHMA2, a P(1B)-ATPase from barley, is highly conserved among cereals and functions in Zn and Cd transport. PLoS ONE 2012, 7, e42640. [Google Scholar] [CrossRef] [PubMed]
- DiDonato, R.J., Jr.; Roberts, L.A.; Sanderson, T.; Eisley, R.B.; Walker, E.L. Arabidopsis Yellow Stripe-Like2 (YSL2): A metal-regulated gene encoding a plasma membrane transporter of nicotianamine-metal complexes. Plant J. 2004, 39, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Inoue, H.; Mizuno, D.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem. Plant J. 2004, 39, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Lelièvre, F.; Bolte, S.; Hamès, C.; Alcon, C.; Neumann, D.; Vansuyt, G.; Curie, C.; Schröder, A.; Krämer, U.; et al. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005, 24, 4041–4051. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Jiménez, M.; Castro-Rodríguez, R.; Kryvoruchko, I.; Lucas, M.M.; Udvardi, M.; Imperial, J.; González-Guerrero, M. Medicago truncatula natural resistance-sssociated macrophage protein1 is required for iron uptake by rhizobia-infected nodule cells. Plant Physiol. 2015, 168, 258–272. [Google Scholar] [CrossRef]
- Qin, L.; Han, P.; Chen, L.; Walk, T.C.; Li, Y.; Hu, X.; Xie, L.; Liao, H.; Liao, X. Genome-Wide Identification and Expression Analysis of NRAMP Family Genes in Soybean (Glycine max L.). Front. Plant Sci. 2017, 8, 1436. [Google Scholar] [CrossRef]
- Yamaji, N.; Sasaki, A.; Xia, J.X.; Yokosho, K.; Ma, J.F. A node-based switch for preferential distribution of manganese in rice. Nat. Commun. 2013, 4, 2442. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Transporters involved in mineral nutrient uptake in rice. J. Exp. Bot. 2016, 67, 3645–3653. [Google Scholar] [CrossRef]
- Li, X.; Wu, Y.; Li, B.; He, W.; Yang, Y.; Yang, Y. Genome-Wide Identification and Expression Analysis of the Cation Diffusion Facilitator Gene Family in Turnip Under Diverse Metal Ion Stresses. Front. Genet. 2018, 9, 103. [Google Scholar] [CrossRef]
- Van der Zaal, B.J.; Neuteboom, L.W.; Pinas, J.E.; Chardonnens, A.N.; Schat, H.; Verkleij, J.A.; Hooykaas, P.J. Overexpression of a novel Arabidopsis gene related to putative zinc-transporter genes from animals can lead to enhanced zinc resistance and accumulation. Plant Physiol. 1999, 119, 1047–1055. [Google Scholar] [CrossRef]
- Delhaize, E.; Kataoka, T.; Hebb, D.M.; White, R.G.; Ryan, P.R. Genes encoding proteins of the cation diffusion facilitator family that confer manganese tolerance. Plant Cell 2003, 15, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, B. Identification of a rice metal tolerance protein OsMTP11 as a manganese transporter. PLoS ONE 2017, 12, e0174987. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Shigaki, T.; Hirschi, K.D. Plant cation/H+ exchangers (CAXs): Biological functions and genetic manipulations. Plant Biol. 2011, 13, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Pittman, J.K.; Hirschi, K.D. CAX-ing a wide net: Cation/H(+) transporters in metal remediation and abiotic stress signalling. Plant Biol. 2016, 18, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Koren’kov, V.; Park, S.; Cheng, N.H.; Sreevidya, C.; Lachmansingh, J.; Morris, J.; Hirschi, K.; Wagner, G.J. Enhanced Cd2+ -selective root-tonoplast-transport in tobaccos expressing Arabidopsis cation exchangers. Planta 2007, 225, 403–411. [Google Scholar] [CrossRef]
- Zou, W.; Chen, J.; Meng, L.; Chen, D.; He, H.; Ye, G. The rice cation/H+ exchanger family involved in Cd tolerance and transport. Int. J. Mol. Sci. 2021, 22, 8186. [Google Scholar] [CrossRef]
- Yuan, M.; Li, X.; Xiao, J.; Wang, S. Molecular and functional analyses of COPT/Ctr-type copper transporter-like gene family in rice. BMC Plant Biol. 2011, 11, 69. [Google Scholar] [CrossRef]
- Carrió-Seguí, A.; Romero, P.; Curie, C.; Mari, S.; Peñarrubia, L. Copper transporter COPT5 participates in the crosstalk between vacuolar copper and iron pools mobilization. Sci. Rep. 2019, 9, 4648. [Google Scholar] [CrossRef]
- Gayomba, S.R.; Jung, H.I.; Yan, J.; Danku, J.; Rutzke, M.A.; Bernal, M.; Krämer, U.; Kochian, L.V.; Salt, D.E.; Vatamaniuk, O.K. The CTR/COPT-dependent copper uptake and SPL7-dependent copper deficiency responses are required for basal cadmium tolerance in A. thaliana. Metallomics 2013, 5, 1262–1275. [Google Scholar] [CrossRef]
- Song, W.Y.; Park, J.; Eisenach, C.; Maeshima, M.; Lee, Y.; Martinoia, E. ABC Transporters and Heavy Metals. In Plant ABC Transporters; Geisler, M., Ed.; Springer: Cham, Switzerland, 2014; pp. 1–17. [Google Scholar] [CrossRef]
- Yokosho, K.; Yamaji, N.; Mitani-Ueno, N.; Shen, R.F.; Ma, J.F. An Aluminum-Inducible IREG Gene is Required for Internal Detoxification of Aluminum in Buckwheat. Plant Cell Physiol. 2016, 57, 1169–1178. [Google Scholar] [CrossRef]
- Morrissey, J.; Baxter, I.R.; Lee, J.; Li, L.; Lahner, B.; Grotz, N.; Kaplan, J.; Salt, D.E.; Guerinot, M.L. The ferroportin metal efflux proteins function in iron and cobalt homeostasis in Arabidopsis. Plant Cell 2009, 21, 3326–3338. [Google Scholar] [CrossRef] [PubMed]
- Gichner, T.; Patkova, Z.; Szakova, J.; Demnerova, K. Cadmium induces DNA damages in tobacco roots, but no DNA damage, somatic mutations orhomologous recombinations in tobacco leaves. Mutat. Res. Genet. Toxicol. Environ. Mut. 2004, 559, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Chai, T.; Zhang, Y.; Xu, J.; Wei, W. Enhancement of Cd tolerance in transgenic tobacco plants overexpressing a Cd-induced catalase cDNA. Chemosphere 2009, 76, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.M.; Chen, S.P.; Chen, L.F.O.; Chiang, M.C.; Chien, H.L.; Lin, K.H. Expression of the broccoli catalase gene (BoCAT) enhances heat tolerance in transgenic Arabidopsis. J. Plant Biochem. Biotechnol. 2013, 23, 266–277. [Google Scholar] [CrossRef]
- Tseng, M.J.; Liu, C.W.; Yiu, J.C. Enhanced tolerance to sulfur dioxide and salt stress of transgenic Chinese cabbage plants expressing both superoxide dismutase and catalase in chloroplasts. Plant Physiol. Biochem. 2007, 45, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Ahsan, N.; Lee, K.W.; Kim, D.H.; Lee, D.G.; Kwak, S.S.; Kwon, S.Y.; Kim, T.H.; Lee, B.H. Simultaneous overexpression of both CuZn superoxide dismutase and ascorbate peroxidase in transgenic tall fescue plants confers increased tolerance to a wide range of abiotic stresses. J. Plant Physiol. 2007, 164, 1626–1638. [Google Scholar] [CrossRef]
- Basu, U.; Good, A.G.; Taylor, G.J. Transgenic Brassica napus plants overexpressing aluminium-induced mitochondrial manganese superoxide dismutase cDNA are resistant to aluminium. Plant Cell Environ. 2001, 24, 1269–1278. [Google Scholar] [CrossRef]
- Kim, I.S.; Shin, S.Y.; Kim, Y.S.; Kim, H.Y.; Yoon, H.S. Expression of a glutathione reductase from Brassica rapa subsp. pekinensis enhanced cellular redox homeostasis by modulating antioxidant proteins in Escherichia coli. Mol. Cells. 2009, 28, 479–487. [Google Scholar] [CrossRef]
- Le Martret, B.; Poage, M.; Shiel, K.; Nugent, G.D.; Dix, P.J. Tobacco chloroplast transformants expressing genes encoding dehydroascorbate reductase, glutathione reductase, and glutathione-S-transferase, exhibit altered anti-oxidant metabolism and improved abiotic stress tolerance. Plant Biotechnol. J. 2011, 9, 661–673. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Eltayeb, A.E.; Uddin, M.I.; Yamamoto, Y.; Tsuji, W.; Takeuchi, Y.; Tanaka, K. Overexpression of dehydroascorbate reductase, but not monodehydroascorbate reductase, confers tolerance to aluminum stress in transgenic tobacco. Planta 2010, 231, 609–621. [Google Scholar] [CrossRef]
- Pence, N.S.; Larsen, P.B.; Ebbs, S.D.; Lasat, M.M.; Letham, D.L.D.; Garvin, D.F.; Eide, D.; Kochian, L.V. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc. Natl. Acad. Sci. USA 2000, 97, 4956–4960. [Google Scholar] [CrossRef] [PubMed]
- Nosek, M.; Kaczmarczyk, A.; Jędrzejczyk, R.J.; Supel, P.; Kaszycki, P.; Miszalski, Z. Expression of Genes Involved in Heavy Metal Trafficking in Plants Exposed to Salinity Stress and Elevated Cd Concentrations. Plants 2020, 9, 475. [Google Scholar] [CrossRef]
- Yang, Q.; Ma, X.; Luo, S.; Gao, J.; Yang, X.; Feng, Y. SaZIP4, an uptake transporter of Zn/Cd hyperaccumulator Sedum alfredii Hance. Environ. Exp. Bot. 2018, 155, 107–117. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Dong, J.; Tan, L.; Ji, Z.; Li, Y.; Sun, Y.; Chen, C.; Lv, Q.; Mao, B.; Hu, Y.; et al. Overexpression of OsLCT2, a Low-Affinity Cation Transporter Gene, Reduces Cadmium Accumulation in Shoots and Grains of Rice. Rice 2021, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.F.; Hassan, Z.; Talukdar, S.; Schat, H.; Aarts, M.G. Expression of the ZNT1 Zinc Transporter from the Metal Hyperaccumulator Noccaea caerulescens Confers Enhanced Zinc and Cadmium Tolerance and Accumulation to Arabidopsis thaliana. PLoS ONE 2016, 11, e0149750. [Google Scholar] [CrossRef]
- Talke, I.N.; Hanikenne, M.; Krämer, U. Zinc-Dependent Global Transcriptional Control, Transcriptional Deregulation, and Higher Gene Copy Number for Genes in Metal Homeostasis of the Hyperaccumulator Arabidopsis halleri. Plant Physiol. 2006, 142, 148–167. [Google Scholar] [CrossRef]
- Lin, W.-C.; Cheng, M.-L.; Wu, J.-W.; Yang, N.-S.; Cheng, C.-P. A glycine-rich protein gene family predominantly expressed in tomato roots, but not in leaves and ripe fruit. Plant Sci. 2005, 168, 283–295. [Google Scholar] [CrossRef]
- Assunção, A.G.L.; Da Costa Martins, P.; De Folter, S.; Vooijs, R.; Schat, H.; Aarts, M.G.M. Elevated expression of metal transporter genes in three accessions of the metal hyperaccumulator Thlaspi caerulescens. Plant Cell Environ. 2001, 24, 217–226. [Google Scholar] [CrossRef]
- Milner, M.J.; Craft, E.; Yamaji, N.; Koyama, E.; Ma, J.F.; Kochian, L.V. Characterization of the high affinity Zn transporter from Noccaea caerulescens, NcZNT1, and dissection of its promoter for its role in Zn uptake and hyperaccumulation. New Phytol. 2012, 195, 113–123. [Google Scholar] [CrossRef]
- Chen, M.; Shen, X.; Li, D.; Ma, L.; Dong, J.; Wang, T. Identification and characterization of MtMTP1, a Zn transporter of CDF family, in the Medicago truncatula. Plant Physiol. Biochem. 2009, 47, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Fasani, E.; DalCorso, G.; Varotto, C.; Li, M.; Visioli, G.; Mattarozzi, M.; Furini, A. TheMTP1 promoters from Arabidopsis helleri reveal cis-regulating elements for the evolution of metal tolerance. New Phytol. 2017, 214, 1614–1630. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, O.N.; Alvarez, D.; Torres, C.; Roman, L.; Daniell, H. Metallothionein expression in chloroplasts enhances mercury accumulation and phytoremediation capability. Plant Biotechnol. J. 2011, 9, 609–617. [Google Scholar] [CrossRef] [PubMed]
| # | Type of Strategy | Description |
|---|---|---|
| 1 | Strategy for avoiding heavy metal uptake |
|
| 2 | Strategy of plant tolerance to heavy metals (ion uptake and neutralization) |
|
| Heavy Metals | Hyperaccumulators | Methods of Phytoremediation | References |
|---|---|---|---|
| Cd, Cu, Pb, Zn | Salix viminalis, Salix fragilis | phytoextraction | [33,34,35] |
| Cd | Ricinus communis, Thlaspi caerulescens, Arabidopsis halleriandSolanum nigrum L. | phytoextraction | [36,37] |
| Cd, Pb, Zn | Zea mays | phytoextraction “phytoattenuation” | [38] |
| Cd, Cu, Pb, Zn | Populus deltoides, Populus nigra, Populus trichocarpa | phytoextraction, phytostabilization | [35,39] |
| Cd, Cu, Ni, Pb, Hg | Jatropha curcas L. | phytoextraction | [40] |
| Cu, Pb, Fe | Eichhornia crassipes | rhizofiltration | [41] |
| Hg | Populus deltoides | phytovolatilization | [42] |
| Se | Brassica juncea, Astragalus bisulcatus, Astragalus racemosus, Cardamine hupingshanesis | phytovolatilization | [43] |
| Zn | Populus nigra, Populus canescens | phytoextraction | [44] |
| Ni | Alyssum murale, Berkheya coddii, Thlaspi goesingense | phytoextraction | [45] |
| Pb, Cd, Cu, Ni, Zn, Cr | Thlaspi caerulescens, Brassica juncea, Pteris vittata, Arabis paniculata, Lolium italicum, Alyssum heldreichii | rhizofiltration | [46] |
| Cu, As, Cd, Pb, Zn | Piptatherum miliaceum, Euphorbia sp., Atriplex lentiformis | phytostabilization | [47,48,49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skuza, L.; Szućko-Kociuba, I.; Filip, E.; Bożek, I. Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals. Int. J. Mol. Sci. 2022, 23, 9335. https://doi.org/10.3390/ijms23169335
Skuza L, Szućko-Kociuba I, Filip E, Bożek I. Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals. International Journal of Molecular Sciences. 2022; 23(16):9335. https://doi.org/10.3390/ijms23169335
Chicago/Turabian StyleSkuza, Lidia, Izabela Szućko-Kociuba, Ewa Filip, and Izabela Bożek. 2022. "Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals" International Journal of Molecular Sciences 23, no. 16: 9335. https://doi.org/10.3390/ijms23169335
APA StyleSkuza, L., Szućko-Kociuba, I., Filip, E., & Bożek, I. (2022). Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals. International Journal of Molecular Sciences, 23(16), 9335. https://doi.org/10.3390/ijms23169335