
Cryobanking European Mink (Mustela lutreola) Mesenchymal Stem Cells and Oocytes


Abstract
:1. Introduction
2. Results
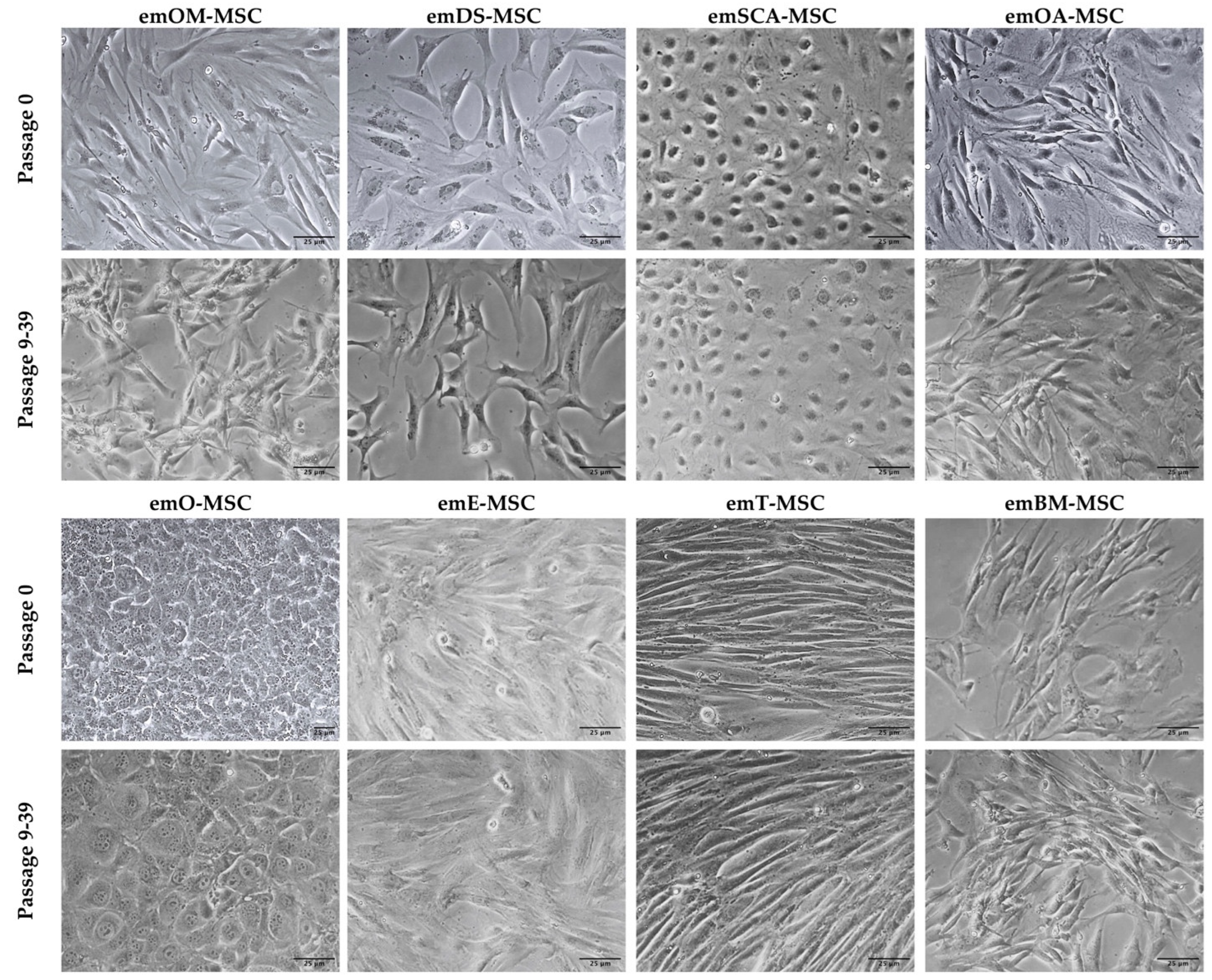
2.1. Isolation of European Mink MSC from Different Tissues
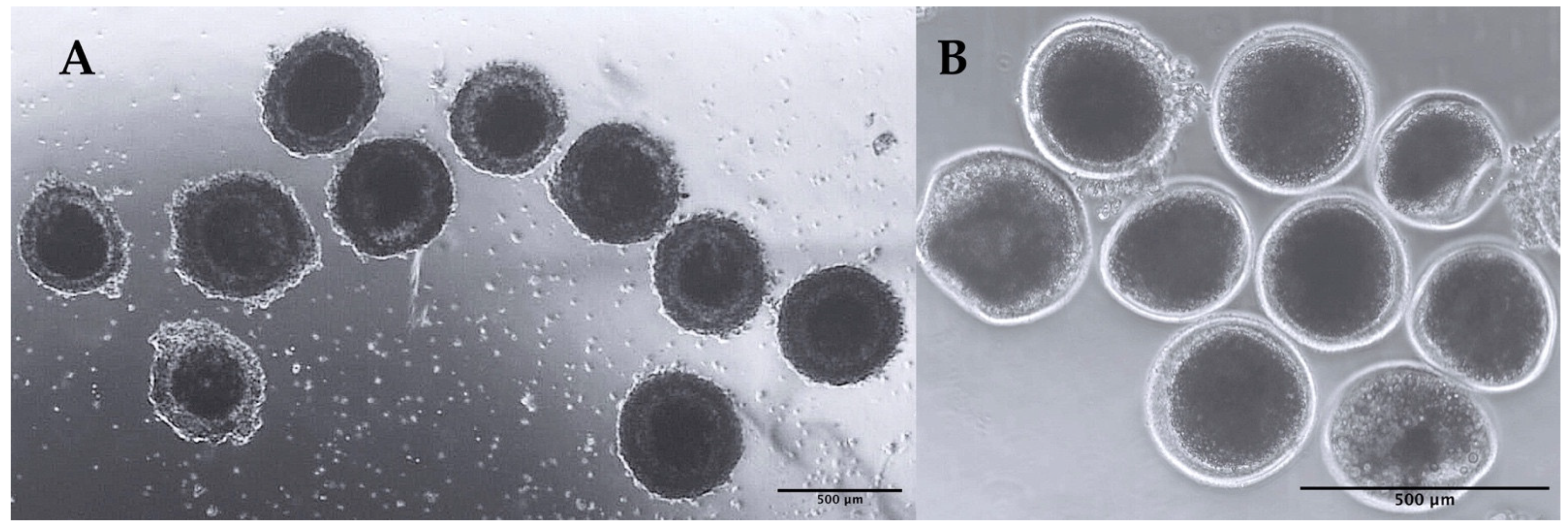
2.2. Isolation, Sorting, and Cryobanking of Immature Oocytes
2.3. Immunocytochemical Analysis by Flow Cytometry
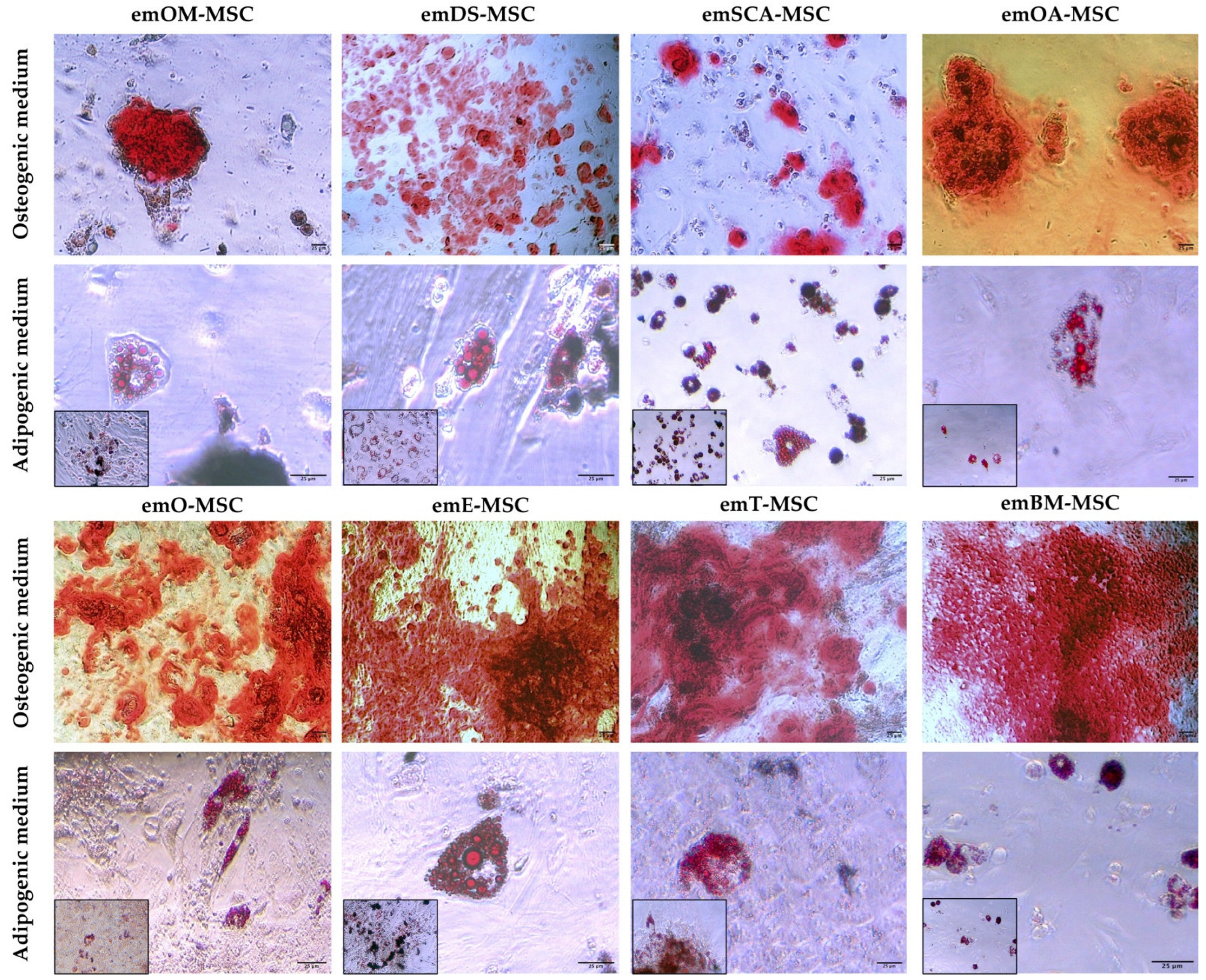
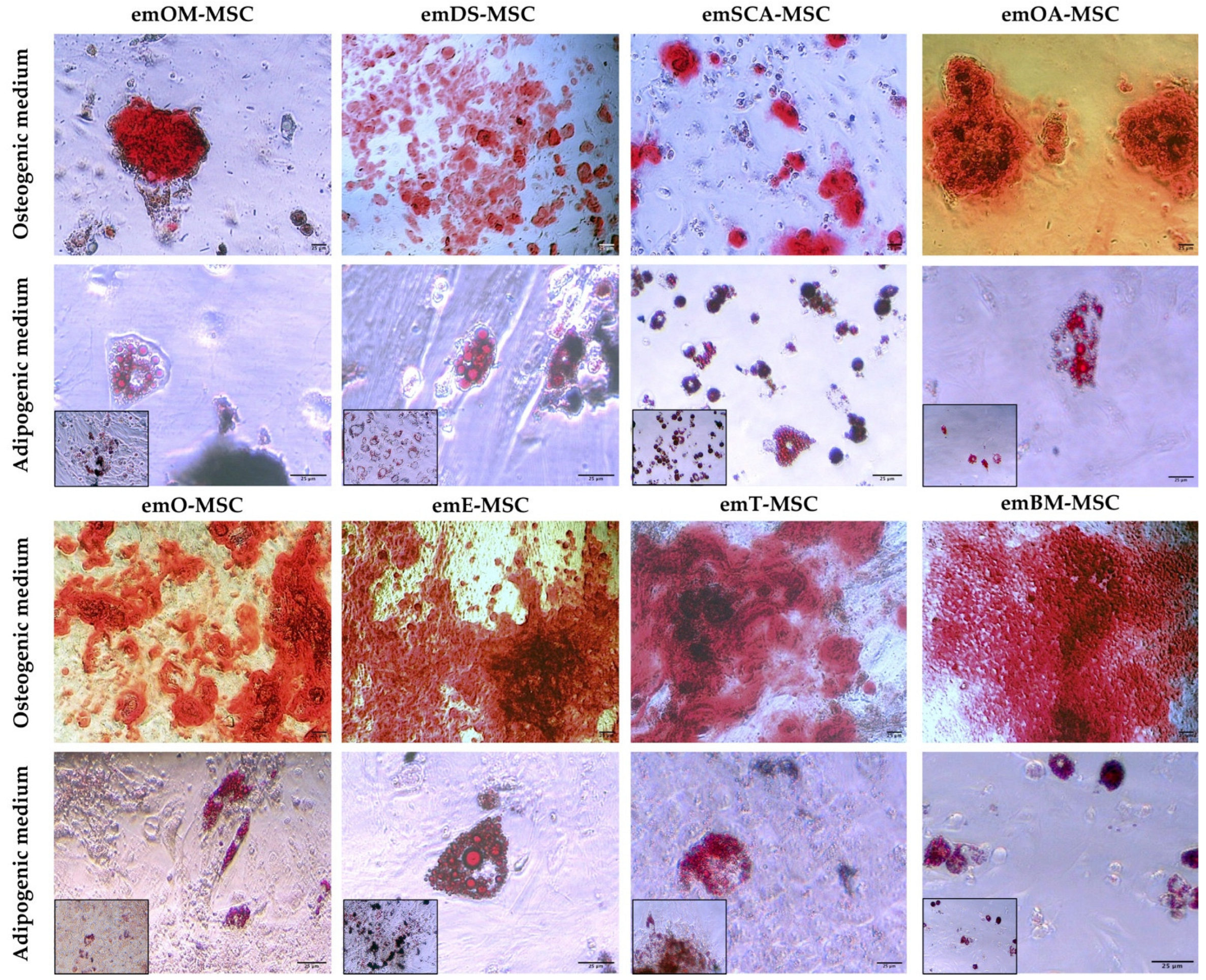
2.4. Differentiation Assay
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of European Mink Cell Lines
4.2. Collection and Vitrification of Immature Oocytes
4.3. Immunocytochemical Analysis of emMSC by Flow Cytometry
4.4. In Vitro Differentiation Potential Assay of emMSC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, T.; Sharma, A.; Ghazaleh, R.A.; Blakemore, W.E.; Ellard, F.M.; Simmons, D.L.; Newman, J.W.; Stuart, D.I.; King, A.M. Arginine-glycine-aspartic acid-specific binding by foot-and-mouth disease viruses to the purified integrin alpha(v)beta3 in vitro. J. Virol. 1997, 71, 8357–8361. [Google Scholar] [CrossRef] [PubMed]
- Skorupski, J. Fifty Years of Research on European Mink Mustela lutreola L., 1761 Genetics: Where Are We Now in Studies on One of the Most Endangered Mammals? Genes 2020, 11, 1332. [Google Scholar] [CrossRef] [PubMed]
- Lodé, T.; Cormier, J.-P.; Le Jacques, D. Decline in endangered species as an indication of anthropic pressures: The case of European mink Mustela lutreola Western population. Environ. Manag. 2001, 28, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Michaux, J.R.; Hardy, O.J.; Justy, F.; Fournier, P.; Kranz, A.; Cabria, M.; Davison, A.; Rosoux, R.; Libois, R. Conservation genetics and population history of the threatened European mink Mustela lutreola, with an emphasis on the west European population. Mol. Ecol. 2005, 14, 2373–2388. [Google Scholar] [CrossRef]
- Santiago, P.I.M.; Ruiz-Olmo, J.; Gosàlbez, I.N.J. Autumn-winter diet of three carnivores, European mink (Mustela lutreola), Eurasian otter (Lutra lutra) and small-spotted genet (Genetta genetta), in Northern Spain. Anim. Biodivers. Conserv. ABC 2009, 31, 37–43. [Google Scholar]
- Mañas, S.; Gómez, A.; Palazón, S.; Pǒdra, M.; Minobis, B.; Alarcia, O.E.; Casal, J.; Ruiz-Olmo, J. Are we able to affect the population structure of an invasive species through culling? A case study of the attempts to control the American mink population in the Northern Iberian Peninsula. Mammal Res. 2016, 61, 309–317. [Google Scholar] [CrossRef]
- Põdra, M.; Gómez, A.; Palazón, S. Do American mink kill European mink? Cautionary message for future recovery efforts. Eur J. Wildl. Res. 2013, 59, 431–440. [Google Scholar] [CrossRef]
- Palazón, S.; Ceña, J.; Ruiz-Olmo, J.; Ceña, A.; Gosálbez, J.; Gómez-Gayubo, A. Trends in distribution of the European mink (Mustela lutreola L, 1761) in Spain: 1950–1999. Mammalia 2003, 67, 473–484. [Google Scholar] [CrossRef]
- Amstislavsky, S.; Lindeberg, H.; Aalto, J.; Kennedy, M. Conservation of the European Mink (Mustela lutreola): Focus on Reproduction and Reproductive Technologies. Reprod. Domest. Anim. 2008, 43, 502–513. [Google Scholar] [CrossRef]
- Comizzoli, P. Biobanking efforts and new advances in male fertility preservation for rare and endangered species. Asian J. Androl. 2015, 17, 640–645. [Google Scholar] [CrossRef]
- Le Gac, S.; Ferraz, M.; Venzac, B.; Comizzoli, P. Understanding and Assisting Reproduction in Wildlife Species Using Microfluidics. Trends Biotechnol. 2021, 39, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Prentice, J.R.; Anzar, M. Cryopreservation of Mammalian Oocyte for Conservation of Animal Genetics. Veter Med. Int. 2010, 2011, 146405. [Google Scholar] [CrossRef] [PubMed]
- Easley, C.A.; Simerly, C.R.; Schatten, G. Gamete derivation from embryonic stem cells, induced pluripotent stem cells or somatic cell nuclear transfer-derived embryonic stem cells: State of the art. Reprod. Fertil. Dev. 2014, 27, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Praxedes, É.A.; Borges, A.A.; Santos, M.V.; Pereira, A.F. Use of somatic cell banks in the conservation of wild felids. Zoo Biol. 2018, 37, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Spees, J.L.; Lee, R.H.; Gregory, C.A. Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res. Ther. 2016, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- Papait, A.; Vertua, E.; Magatti, M.; Ceccariglia, S.; De Munari, S.; Silini, A.R.; Sheleg, M.; Ofir, R.; Parolini, O. Mesenchymal Stromal Cells from Fetal and Maternal Placenta Possess Key Similarities and Differences: Potential Implications for Their Applications in Regenerative Medicine. Cells 2020, 9, 127. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise review: Mesenchymal stem cells: From roots to boost. Stem Cells 2019, 37, 855–864. [Google Scholar] [CrossRef]
- An, J.-H.; Li, F.-P.; He, P.; Chen, J.-S.; Cai, Z.-G.; Liu, S.-R.; Yue, C.-J.; Liu, Y.-L.; Hou, R. Characteristics of Mesenchymal Stem Cells Isolated from the Bone Marrow of Red Pandas. Zoology 2020, 140, 125775. [Google Scholar] [CrossRef]
- Gómez, M.C.; Qin, Q.; Biancardi, M.N.; Galiguis, J.; Dumas, C.; MacLean, R.A.; Wang, G.; Pope, C.E. Characterization and Multilineage Differentiation of Domestic and Black-Footed Cat Mesenchymal Stromal/Stem Cells from Abdominal and Subcutaneous Adipose Tissue. Cell. Reprogram. 2015, 17, 376–392. [Google Scholar] [CrossRef]
- Verma, R.; Holland, M.; Temple-Smith, P.; Verma, P. Inducing pluripotency in somatic cells from the snow leopard (Panthera uncia), an endangered felid. Theriogenology 2012, 77, 220–228.e1. [Google Scholar] [CrossRef]
- De los Reyes, M.; de Lange, J.; Miranda, P.; Palominos, J.; Barros, C. Effect of human chorionic gonadotrophin supplementation during different culture periods on in vitro maturation of canine oocytes. Theriogenology 2005, 64, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Azami-Conesa, I.; Sansano-Maestre, J.; Martínez-Díaz, R.; Gómez-Muñoz, M. Invasive Species as Hosts of Zoonotic Infections: The Case of American Mink (Neovison vison) and Leishmania infantum. Microorganisms 2021, 9, 1531. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Rudaitytė-Lukošienė, E.; Sneideris, D.; Butkauskas, D. Invasive American mink (Neovison vison) as potential definitive host of Sarcocystis elongata, S. entzerothi, S. japonica, S. truncata and S. silva using different cervid species as intermediate hosts. Parasitol Res. 2021, 120, 2243–2250. [Google Scholar] [CrossRef] [PubMed]
- Giner, J.; Villanueva-Saz, S.; Fernández, A.; Gómez, M.A.; Podra, M.; Lizarraga, P.; Lacasta, D.; Ruiz, H.; Aranda, M.D.C.; Jimenez, M.D.L.; et al. Detection of Anti–Leishmania infantum Antibodies in Wild European and American Mink (Mustela lutreola and Neovison vison) from Northern Spain, 2014–2020. J. Wildl. Dis. 2022, 58, 198–204. [Google Scholar] [CrossRef]
- Kiik, K.; Maran, T.; Nemvalts, K.; Sandre, S.-L.; Tammaru, T. Reproductive parameters of critically endangered European mink (Mustela lutreola) in captivity. Anim. Reprod. Sci. 2017, 181, 86–92. [Google Scholar] [CrossRef]
- Kiik, K.; Maran, T.; Nagl, A.; Ashford, K.; Tammaru, T. The causes of the low breeding success of european mink (Mustela lutreola) in captivity. Zoo Biol. 2013, 32, 387–393. [Google Scholar] [CrossRef]
- Łupicka, M.; Bodek, G.; Shpigel, N.; Elnekave, E.; Korzekwa, A.J. Identification of pluripotent cells in bovine uterus: In situ and in vitro studies. Reproduction 2015, 149, 317–327. [Google Scholar] [CrossRef]
- Hugo, H.; Ackland, L.; Blick, T.; Lawrence, M.G.; Clements, J.; Williams, E.D.; Thompson, E.W. Epithelial—mesenchymal and mesenchymal—epithelial transitions in carcinoma progression. J. Cell. Physiol. 2007, 213, 374–383. [Google Scholar] [CrossRef]
- Calle, A.; López-Martín, S.; Monguió-Tortajada, M.; Borràs, F.E.; Yáñez-Mó, M.; Ramírez, M. Bovine endometrial MSC: Mesenchymal to epithelial transition during luteolysis and tropism to implantation niche for immunomodulation. Stem Cell Res. Ther. 2019, 10, 239. [Google Scholar] [CrossRef]
- Calle, A.; Zamora-Ceballos, M.; Bárcena, J.; Blanco, E.; Ramírez, M. Comparison of Biological Features of Wild European Rabbit Mesenchymal Stem Cells Derived from Different Tissues. Int. J. Mol. Sci. 2022, 23, 6420. [Google Scholar] [CrossRef]
- Calle, A.; Gutiérrez-Reinoso, M.; Re, M.; Blanco, J.; De La Fuente, J.; Monguió-Tortajada, M.; Borràs, F.E.; Yáñez-Mó, M.; Ramírez, M. Bovine peripheral blood MSCs chemotax towards inflammation and embryo implantation stimuli. J. Cell. Physiol. 2021, 236, 1054–1067. [Google Scholar] [CrossRef] [PubMed]
- Calle, A.; Barrajón-Masa, C.; Gómez-Fidalgo, E.; Martín-Lluch, M.; Cruz-Vigo, P.; Sánchez-Sánchez, R.; Ramírez, M. Iberian pig mesenchymal stem/stromal cells from dermal skin, abdominal and subcutaneous adipose tissues, and peripheral blood: In vitro characterization and migratory properties in inflammation. Stem Cell Res. Ther. 2018, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Corradetti, B.; Meucci, A.; Bizzaro, D.; Cremonesi, F.; Lange-Consiglio, A. Mesenchymal stem cells from amnion and amniotic fluid in the bovine. Reproduction 2013, 145, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, T.C.; Ferrari, H.F.; Garcia, A.F.; Novais, J.B.; Silva-Frade, C.; Ferrarezi, M.C.; Andrade, A.L.; Gameiro, R. Isolation and characterization of Wharton’s jelly-derived multipotent mesenchymal stromal cells obtained from bovine umbilical cord and maintained in a defined serum-free three-dimensional system. BMC Biotechnol. 2012, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Lorente, R.; Bejar, M.T.; Badimon, L. Notch Signaling Pathway Activation in Normal and Hyperglycemic Rats Differs in the Stem Cells of Visceral and Subcutaneous Adipose Tissue. Stem Cells Dev. 2014, 23, 3034–3048. [Google Scholar] [CrossRef] [PubMed]
- Tchkonia, T.; Giorgadze, N.; Pirtskhalava, T.; Thomou, T.; DePonte, M.; Koo, A.; Forse, R.A.; Chinnappan, D.; Martin-Ruiz, C.; von Zglinicki, T.; et al. Fat Depot–Specific Characteristics Are Retained in Strains Derived from Single Human Preadipocytes. Diabetes 2006, 55, 2571–2578. [Google Scholar] [CrossRef]
- Przekora, A.; Juszkiewicz, L. The Effect of Autologous Adipose Tissue–Derived Mesenchymal Stem Cells’ Therapy in the Treatment of Chronic Posttraumatic Spinal Cord Injury in a Domestic Ferret Patient. Cell Transplant. 2020, 29, 963689720928982. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Hsu, S.-C.; Yao, M.; Huang, D.-M. CD9 Upregulation-Decreased CCL21 Secretion in Mesenchymal Stem Cells Reduces Cancer Cell Migration. Int. J. Mol. Sci. 2021, 22, 1738. [Google Scholar] [CrossRef]
- Halfon, S.; Abramov, N.; Grinblat, B.; Ginis, I. Markers Distinguishing Mesenchymal Stem Cells from Fibroblasts Are Downregulated with Passaging. Stem Cells Dev. 2011, 20, 53–66. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yu, J.M.; Joo, H.J.; Kim, H.K.; Cho, H.H.; Bae, Y.C.; Jung, J.S. Role of CD9 in proliferation and proangiogenic action of human adipose-derived mesenchymal stem cells. Pflugers Arch. 2007, 455, 283–296. [Google Scholar] [CrossRef]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell. Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Holley, R.J.; Tai, G.; Williamson, A.J.; Taylor, S.; Cain, S.A.; Richardson, S.M.; Merry, C.L.; Whetton, A.D.; Kielty, C.M.; Canfield, A.E. Comparative Quantification of the Surfaceome of Human Multipotent Mesenchymal Progenitor Cells. Stem Cell Rep. 2015, 4, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, V.; Rezende, N.C.; Scotland, K.B.; Shaffer, S.M.; Persson, J.L.; Gudas, L.J.; Mongan, N.P. Regulation of Stem Cell Pluripotency and Differentiation Involves a Mutual Regulatory Circuit of the Nanog, OCT4, and SOX2 Pluripotency Transcription Factors with Polycomb Repressive Complexes and Stem Cell microRNAs. Stem Cells Dev. 2009, 18, 1093–1108. [Google Scholar] [CrossRef]
- Wang, D.-H.; Wu, X.-M.; Chen, J.-S.; Cai, Z.-G.; An, J.-H.; Zhang, M.-Y.; Li, Y.; Li, F.-P.; Hou, R.; Liu, Y.-L. Isolation and characterization mesenchymal stem cells from red panda (Ailurus fulgens styani) endometrium. Conserv. Physiol. 2022, 10, coac004. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, F.; Cai, Z.; Wang, D.; Hou, R.; Zhang, H.; Zhang, M.; Yie, S.; Wu, K.; Zeng, C.; et al. Isolation and characterization of mesenchymal stem cells from umbilical cord of giant panda. Tissue Cell 2021, 71, 101518. [Google Scholar] [CrossRef] [PubMed]
- Melotti, L.; Martinello, T.; Perazzi, A.; Martines, E.; Zuin, M.; Modenese, D.; Cordaro, L.; Ferro, S.; Maccatrozzo, L.; Iacopetti, I.; et al. Could cold plasma act synergistically with allogeneic mesenchymal stem cells to improve wound skin regeneration in a large size animal model? Res. Veter Sci. 2021, 136, 97–110. [Google Scholar] [CrossRef]
- Putra, A.; Ibrahim, S.; Muhar, A.M.; Kuntardjo, N.; Dirja, B.T.; Pasongka, Z.; Tunru, I.S. Topical gel of mesenchymal stem cells-conditioned medium under TNF-α precondition accelerates wound closure healing in full-thickness skin defect animal model. J. Med. Life 2022, 15, 214–221. [Google Scholar] [CrossRef]
- Vázquez, A.; Fernández-Sevilla, L.M.; Jiménez, E.; Pérez-Cabrera, D.; Yañez, R.; Subiza, J.L.; Varas, A.; Valencia, J.; Vicente, A. Involvement of Mesenchymal Stem Cells in Oral Mucosal Bacterial Immunotherapy. Front. Immunol. 2020, 11, 567391. [Google Scholar] [CrossRef]
- Enders, R.K. Reproduction in the Mink (Mustela Vison). Proc. Am. Philos. Soc. 1952, 96, 691–755. [Google Scholar]
- Echeverry, D.M.; Asenjo, P.A.; Rojas, D.M.; Aguilera, C.J.; Rodríguez-Álvarez, L.; Castro, F.O. Characterization of mesenchymal stem cells derived from adipose tissue of a cougar (Puma concolor). Anim. Reprod. 2020, 17, e20190109. [Google Scholar] [CrossRef]
- Avanzi, S.; Leoni, V.; Rotola, A.; Alviano, F.; Solimando, L.; Lanzoni, G.; Bonsi, L.; Di Luca, D.; Marchionni, C.; Alvisi, G.; et al. Susceptibility of Human Placenta Derived Mesenchymal Stromal/Stem Cells to Human Herpesviruses Infection. PLoS ONE 2013, 8, e71412. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.A.; Travis, A.J. Stem cells in veterinary medicine. Stem Cell Res. Ther. 2011, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Cagliani, J.; Grande, D.; Molmenti, E.P.; Miller, E.J.; Rilo, H.L. Immunomodulation by Mesenchymal Stromal Cells and Their Clinical Applications. J. Stem Cell Regen. Biol. 2017, 3, 1–14. [Google Scholar] [CrossRef]
- Turathum, B.; Saikhun, K.; Sangsuwan, P.; Kitiyanant, Y. Effects of vitrification on nuclear maturation, ultrastructural changes and gene expression of canine oocytes. Reprod. Biol. Endocrinol. 2010, 8, 70. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source (European Mink Tissue) | N (Primary Cultures) | N (Cell Lines) | Animal ID | Cell Passages | Nº Vials |
|---|---|---|---|---|---|
| Peripheral blood tissue derived cells (emPB-C) | 1 | 89 ♂ | 5 | 7 | |
| Bone marrow mesenchymal stem cells (emBM-MSC) | 2 | 89 ♂ | 12 | 15 | |
| 90 ♂ | 4 | 4 | |||
| Abdominal adipose tissue-derived cells (emAA-C) | 90 ♀ | 1 | 0 | ||
| 1 | 79 ♀ | 1 | 2 | ||
| Subcutaneous adipose mesenchymal stem cells (emSCA-MSC) | 2 | 78 ♂ | 25 | 34 | |
| 89 ♂ | 6 | 10 | |||
| Dermal skin mesenchymal stem cells (emDS-MSC) | 6 | 75 ♀ | 33 | 29 | |
| 77 ♀ | 4 | 2 | |||
| 78 ♂ | 12 | 14 | |||
| 80 ♂ | 3 | 1 | |||
| 89 ♂ | 5 | 3 | |||
| 90 ♀ | 1 | 1 | |||
| Oral mucose mesenchymal stem cells (emOM-MSC) | 5 | 78 ♂ | 15 | 23 | |
| 79 ♀ | 3 | 1 | |||
| 80 ♀ | 6 | 8 | |||
| 89 ♂ | 3 | 2 | |||
| 90 ♀ | 14 | 10 | |||
| Pericardial adipose tissue-derived cells (emPA-C) | 1 | 90 ♀ | 5 | 5 | |
| Oviductal mesenchymal stem cells (emO-MSC) | 4 | 69B♀ | 11 | 21 | |
| 71 ♀ | 49 | 56 | |||
| 79 ♀ | 7 | 7 | |||
| 90 ♀ | 6 | 6 | |||
| Endometrial mesenchymal stem cells (emE-MSC) | 2 | 79 ♀ | 5 | 10 | |
| 90 ♀ | 9 | 16 | |||
| Ovarian adipose mesenchymal stem cells (emOA-MSC) | 2 | 69B♀ | 5 | 2 | |
| 77 ♀ | 42 | 53 | |||
| Testicular mesenchymal stem cells (emT-MSC) | 3 | 69 ♂ | 45 | 53 | |
| 78 ♂ | 4 | 4 | |||
| 89 ♂ | 6 | 6 | |||
| TOTAL | 3 | 26 | 11 ♀/10 ♂ | 405 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calle, A.; Ramírez, M.Á. Cryobanking European Mink (Mustela lutreola) Mesenchymal Stem Cells and Oocytes. Int. J. Mol. Sci. 2022, 23, 9319. https://doi.org/10.3390/ijms23169319
Calle A, Ramírez MÁ. Cryobanking European Mink (Mustela lutreola) Mesenchymal Stem Cells and Oocytes. International Journal of Molecular Sciences. 2022; 23(16):9319. https://doi.org/10.3390/ijms23169319
Chicago/Turabian StyleCalle, Alexandra, and Miguel Ángel Ramírez. 2022. "Cryobanking European Mink (Mustela lutreola) Mesenchymal Stem Cells and Oocytes" International Journal of Molecular Sciences 23, no. 16: 9319. https://doi.org/10.3390/ijms23169319
APA StyleCalle, A., & Ramírez, M. Á. (2022). Cryobanking European Mink (Mustela lutreola) Mesenchymal Stem Cells and Oocytes. International Journal of Molecular Sciences, 23(16), 9319. https://doi.org/10.3390/ijms23169319