
Novel Plant Extract Ameliorates Metabolic Disorder through Activation of Brown Adipose Tissue in High-Fat Diet-Induced Obese Mice


Abstract
1. Introduction
2. Results
2.1. PUE Treatment Decreased Lipid Accumulation While Promoting Expression of Browning-Specific Genes in 3T3-L1 Adipocytes
2.2. PUE Improved Body, Organ, and Adipose Tissue Weights by Regulating Lipid Metabolism
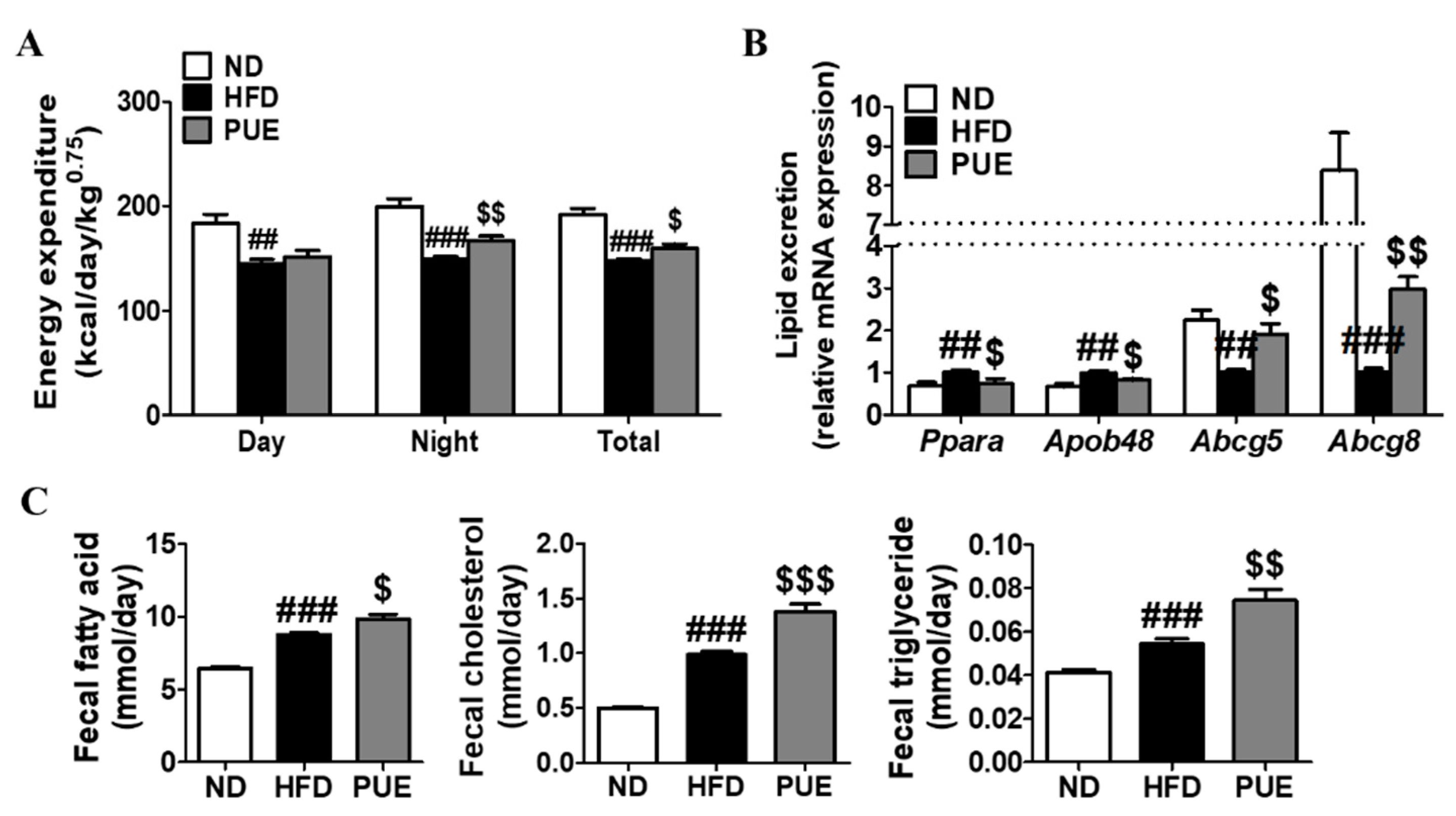
2.3. PUE Increased Energy Expenditure and Fecal Lipid Excretion Levels
2.4. PUE Supplementation Improved Plasma Lipid Levels
2.5. PUE Downregulated Hepatic Glucose Enzyme Activities and Attenuated Insulin Resistance
2.6. PUE Ameliorated Hepatic Steatosis as a Modulator of Hepatic Lipid Metabolism
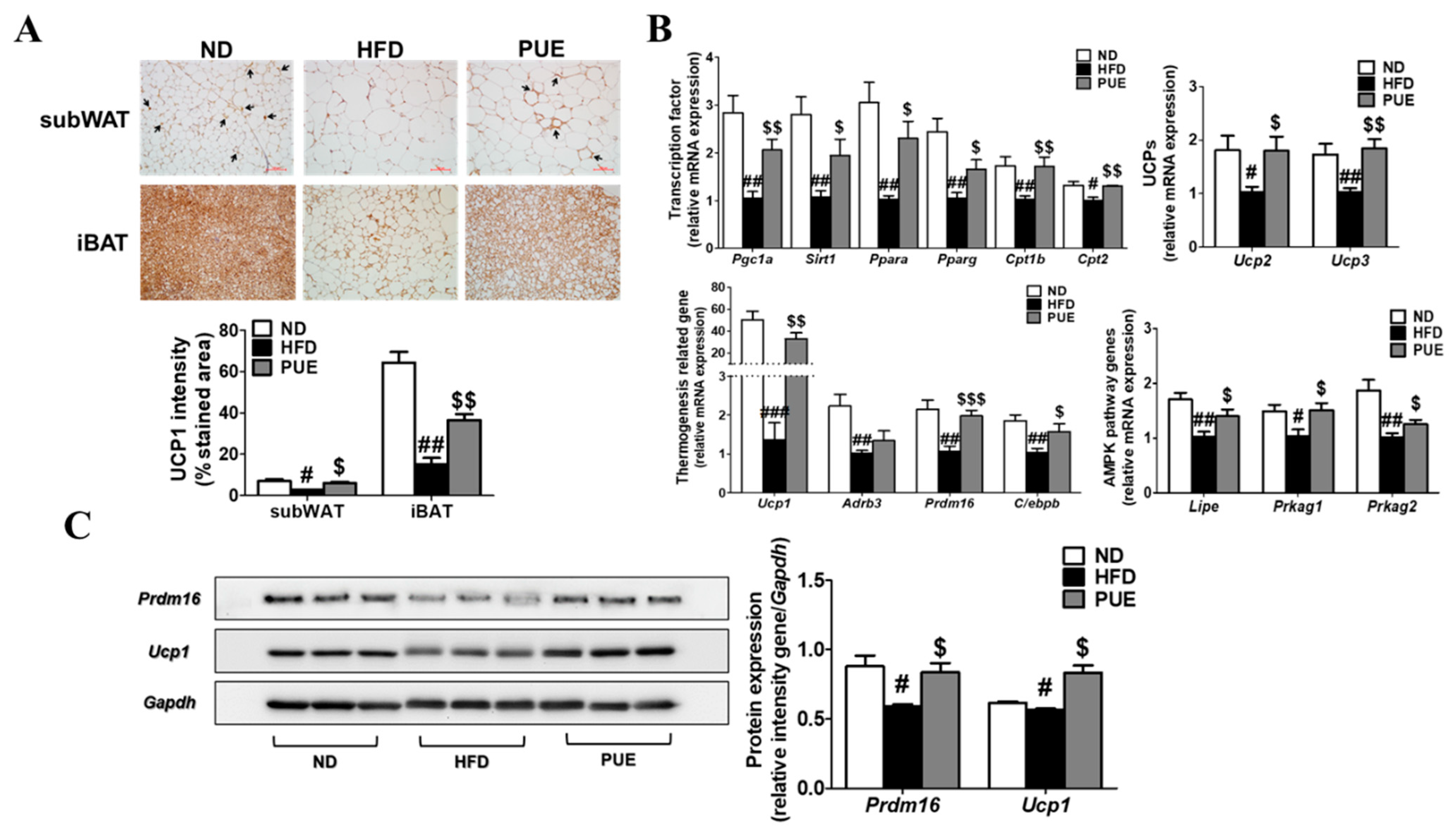
2.7. PUE Promoted BAT Activation and Browning of subWAT
3. Discussion
4. Materials and Methods
4.1. Preparation of P. umbrosa Root Ethanol Extract
4.2. Cell Culture and Cell Chemical Staining
4.2.1. Cell Culture and Treatment
4.2.2. Oil Red O Staining
4.3. Animal Experimental Design and Diet
4.4. Measurements of Energy Expenditure
4.5. Morphology of Liver and Adipose Tissues
4.6. Biochemical Parameters of Plasma, Hepatic, and Fecal Lipids
4.7. Determination of Hormones and Adipokines in Plasma
4.8. Fasting Blood Glucose and Insulin Resistance Index
4.9. Plasma Glutamic Oxaloacetic Transaminase (GOT) and Glutamic Pyruvic Transaminase (GPT) Activities
4.10. Determination of Hepatic Enzyme Activity
4.11. Real-Time qPCR Analysis
4.12. Western Blotting
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| BAT | brown adipose tissue |
| CPT | carnitine palmitoyltransferase |
| DIO | diet-induced obesity |
| EE | energy expenditure |
| eWAT | epididymal WAT |
| FAO | fatty acid oxidation |
| FAS | fatty acid synthase |
| FER | food efficiency ratio |
| GOT | glutamic oxaloacetic transaminase |
| GPT | glutamic pyruvic transaminase |
| G6PD | glucose-6-phosphate dehydrogenase |
| HFD | high-fat diet |
| HOMA-IR | homeostasis model assessment for insulin resistance |
| ME | malic enzyme |
| ND | normal diet |
| NAFLD | non-alcoholic fatty liver disease |
| PAP | phosphatidate phosphohydrolase |
| PBS | phosphate buffered saline |
| PUE | Phlomis umbrosa Turcz. root ethanol extract |
| subWAT | subcutaneous WAT |
| T2DM | type 2 diabetes mellitus |
| UCP1 | uncoupling protein 1 |
| WAT | white adipose tissue |
References
- WHO. Overweight and Obesity. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 6 August 2022).
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Vliora, M.; Ravelli, C.; Grillo, E.; Corsini, M.; Flouris, A.D.; Mitola, S. The impact of adipokines on vascular networks in adipose tissue. Cytokine Growth Factor Rev. 2022. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metab. Clin. Exp. 2019, 92, 82–97. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.; Li, J.; Lin, L.; Zheng, G. A flavonoid-rich Smilax china L. extract prevents obesity by upregulating the adiponectin-receptor/AMPK signalling pathway and modulating the gut microbiota in mice. Food Funct. 2021, 12, 5862–5875. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, A.I.S.; Blindauer, C.A.; Stewart, A.J. Changes in Plasma Free Fatty Acids Associated with Type-2 Diabetes. Nutrients 2019, 11, 2022. [Google Scholar] [CrossRef]
- Li, T.; Gao, J.; Du, M.; Song, J.; Mao, X. Milk Fat Globule Membrane Attenuates High-Fat Diet-Induced Obesity by Inhibiting Adipogenesis and Increasing Uncoupling Protein 1 Expression in White Adipose Tissue of Mice. Nutrients 2018, 10, 331. [Google Scholar] [CrossRef]
- Kawabe, Y.; Mori, J.; Morimoto, H.; Yamaguchi, M.; Miyagaki, S.; Ota, T.; Tsuma, Y.; Fukuhara, S.; Nakajima, H.; Oudit, G.Y.; et al. ACE2 exerts anti-obesity effect via stimulating brown adipose tissue and induction of browning in white adipose tissue. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E1140–E1149. [Google Scholar] [CrossRef]
- Park, A.; Kim, W.K.; Bae, K.H. Distinction of white, beige and brown adipocytes derived from mesenchymal stem cells. World J. Stem Cells 2014, 6, 33. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Kazak, L.; Spiegelman, B.M. New advances in adaptive thermogenesis: UCP1 and beyond. Cell Metab. 2019, 29, 27–37. [Google Scholar] [CrossRef]
- Seale, P.; Conroe, H.M.; Estall, J.; Kajimura, S.; Frontini, A.; Ishibashi, J.; Cohen, P.; Cinti, S.; Spiegelman, B.M. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J. Clin. Investig. 2011, 121, 96–105. [Google Scholar] [CrossRef]
- Nedergaard, J.; Cannon, B. The browning of white adipose tissue: Some burning issues. Cell Metab. 2014, 20, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Bae, I.Y.; Lee, J.Y.; Kwak, B.Y.; Lee, H.G. Estrogenic effects of various extracts from Chamdanggui (Angelica gigas Nakai) and sogdan (Phlomis umbrosa Turcz). Food Sci. Biotechnol. 2011, 20, 1113. [Google Scholar] [CrossRef]
- Lee, D.; Kim, Y.S.; Song, J.; Kim, H.S.; Lee, H.J.; Guo, H.; Kim, H. Effects of Phlomis umbrosa root on longitudinal bone growth rate in adolescent female rats. Molecules 2016, 21, 461. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Lee, H.; Kim, M.H.; Yang, W.M. Osteogenic effects of Phlomis umbrosa via up-regulation of Runx2 in osteoporosis. Biomed Rep. 2019, 10, 17–22. [Google Scholar] [CrossRef]
- Shang, X.; Wang, J.; Li, M.; Miao, X.; Pan, H.; Yang, Y.; Wang, Y. Antinociceptive and anti-inflammatory activities of Phlomis umbrosa Turcz extract. Fitoterapia 2011, 82, 716–721. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Le, D.D.; Zhao, B.T.; Ma, E.S.; Min, B.S.; Woo, M.H. Antioxidant Compounds Isolated from the Roots of Phlomis umbrosa Turcz. Nat. Prod. Sci. 2018, 24, 119–124. [Google Scholar] [CrossRef]
- Chun, J.M.; Lee, A.Y.; Moon, B.C.; Choi, G.; Kim, J.S. Effects of Dipsacus asperoides and Phlomis umbrosa Extracts in a Rat Model of Osteoarthritis. Plants 2021, 10, 2030. [Google Scholar] [CrossRef]
- Yun, J.; Kim, J.; Choi, J.; Kwon, K.; Jo, C.H. Simultaneous determination of Phlomis umbrosa and Dipsacus asperoides in foods using LC-MS/MS methods. Korean J. Food Sci. Technol. 2016, 48, 531–535. [Google Scholar] [CrossRef]
- Stojanović, O.; Altirriba, J.; Rigo, D.; Spiljar, M.; Evrard, E.; Roska, B.; Fabbiano, S.; Zamboni, N.; Maechler, P.; Rohner-Jeanrenaud, F.; et al. Dietary excess regulates absorption and surface of gut epithelium through intestinal PPARα. Nat. Commun. 2021, 12, 7031. [Google Scholar] [CrossRef]
- Han, Y.; Han, H.J.; Kim, A.H.; Choi, J.Y.; Cho, S.J.; Park, Y.B.; Jung, U.J.; Choi, M.S. d-Allulose supplementation normalized the body weight and fat-pad mass in diet-induced obese mice via the regulation of lipid metabolism under isocaloric fed condition. Mol. Nutr. Food. Res. 2016, 60, 1695–1706. [Google Scholar] [CrossRef]
- Bolsoni-Lopes, A.; Alonso-Vale, M.I.C. Lipolysis and lipases in white adipose tissue–an update. Arch. Endocrinol. Metab. 2015, 59, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, S.; Zhai, A.; Zhang, B.; Tian, G. AMPK-mediated regulation of lipid metabolism by phosphorylation. Biol. Pharm. Bull. 2018, 41, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Barua, S.; Jeong, Y.J.; Lee, J.E. Adiponectin: The potential regulator and therapeutic target of obesity and Alzheimer’s disease. Int. J. Mol. Sci. 2020, 21, 6419. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef]
- Ferre, T.; Riu, E.; Franckhauser, S.; Agudo, J.; Bosch, F. Long-term overexpression of glucokinase in the liver of transgenic mice leads to insulin resistance. Diabetologia 2003, 46, 1662–1668. [Google Scholar] [CrossRef]
- Kwon, E.Y.; Jung, U.J.; Park, T.; Yun, J.W.; Choi, M.S. Luteolin attenuates hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue in mice with diet-induced obesity. Diabetes 2015, 64, 1658–1669. [Google Scholar] [CrossRef]
- Fabbrini, E.; Magkos, F. Hepatic steatosis as a marker of metabolic dysfunction. Nutrients 2015, 7, 4995–5019. [Google Scholar] [CrossRef]
- Mottillo, E.P.; Desjardins, E.M.; Crane, J.D.; Smith, B.K.; Green, A.E.; Ducommun, S.; Henriksen, T.I.; Rebalka, I.A.; Razi, A.; Sakamoto, K.; et al. Lack of adipocyte AMPK exacerbates insulin resistance and hepatic steatosis through brown and beige adipose tissue function. Cell Metab. 2016, 24, 118–129. [Google Scholar] [CrossRef]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef]
- Zhang, G.; Sun, Q.; Liu, C. Influencing factors of thermogenic adipose tissue activity. Front. Physiol. 2016, 7, 29. [Google Scholar] [CrossRef]
- Shen, W.; Wang, Y.; Lu, S.F.; Hong, H.; Fu, S.; He, S.; Li, Q.; Yue, J.; Xu, B.; Zhu, B.M. Acupuncture promotes white adipose tissue browning by inducing UCP1 expression on DIO mice. BMC Complement. Altern Med. 2014, 14, 501. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.M.; Suh, K.S.; Jung, W.W.; Park, S.Y.; Chin, S.O.; Rhee, S.Y.; Kim Pak, Y.; Chon, S. Glabridin attenuates antiadipogenic activity induced by 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin in murine 3T3-L1 adipocytes. J. Appl. Toxicol. 2018, 38, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, S.J.; Kwon, E.Y.; Choi, M.S. Physcion reduces lipid accumulation and prevents the obesity in mice. Nutr. Metab. 2019, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Barseem, N.F.; Helwa, M.A. Homeostatic model assessment of insulin resistance as a predictor of metabolic syndrome: Consequences of obesity in children and adolescents. Egypt Pediatric Assoc. Gaz. 2015, 63, 19–24. [Google Scholar] [CrossRef][Green Version]
- Hulcher, F.H.; Oleson, W.H. Simplified spectrophotometric assay for microsomal 3-hydroxy-3-methylglutaryl CoA reductase by measurement of coenzyme A. J. Lipid Res. 1973, 14, 625–631. [Google Scholar] [CrossRef]
- Nepokroeff, C.M.; Lakshmanan, M.R.; Porter, J.W. [6] Fatty acid synthase from rat liver. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1975; Volume 35, pp. 37–44. [Google Scholar]
- Ochoa, S.; Mehleb, A.H.; Kobnbebg, A. Biosynthesis of dicarboxylic acids by carbon dioxide fixation. 1. Isolation and properties of an enzyme from pigeon liver catalyzing the reversible oxidative decarboxylation of L-malic acid. J. Biol. Chem. 1948, 174, 979–1000. [Google Scholar] [CrossRef]
- Pitkänen, E.; Pitkänen, O.; Uotila, L. Enzymatic determination of unbound D-mannose in serum. Clin. Chem. Lab. Med. 1997, 35, 761–766. [Google Scholar] [CrossRef]
- Walton, P.A.; Possmayer, F. Mg2+-dependent phosphatidate phosphohydrolase of rat lung: Development of an assay employing a defined chemical substrate which reflects the phosphohydrolase activity measured using membrane-bound substrate. Anal. Biochem. 1985, 151, 479–486. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; McGroarty, E.J.; Bieber, L.L.; Tolbert, N.E. The subcellular distribution of carnitine acyltransferases in mammalian liver and kidney a new peroxisomal enzyme. J. Biol. Chem. 1973, 248, 3426–3432. [Google Scholar] [CrossRef]
- Lazarow, P.B. [19] Assay of peroxisomal β-oxidation of fatty acids. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1981; Volume 72, pp. 315–319. [Google Scholar]
- Newgard, C.B.; Hirsch, L.J.; Foster, D.W.; McGarry, J.D. Studies on the mechanism by which exogenous glucose is converted into liver glycogen in the rat. A direct or an indirect pathway? J. Biol. Chem. 1983, 258, 8046–8052. [Google Scholar] [CrossRef]
- Alegre, M.; Ciudad, C.J.; Fillat, C.; Guinovart, J.J. Determination of glucose-6-phosphatase activity using the glucose dehydrogenase-coupled reaction. Anal. Biochem. 1988, 173, 185–189. [Google Scholar] [CrossRef]
- Drewnowski, A.; Specter, S.E. Poverty and obesity: The role of energy density and energy costs. Am. J. Clin. Nutr. 2004, 79, 6–16. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND | HFD | PUE | |
|---|---|---|---|
| TG (mmol/L) | 1.09 ± 0.06 | 1.35 ± 0.06 ## | 1.25 ± 0.11 |
| TC (mmol/L) | 3.70 ± 0.14 | 5.84 ± 0.17 ### | 5.16 ± 0.22 $ |
| PL (mg/dL) | 6.50 ± 1.67 | 33.55 ± 2.24 ### | 20.68 ± 2.39 $$ |
| HDL-C (mmol/L) | 0.77 ± 0.04 | 1.13 ± 0.04 ### | 1.06 ± 0.03 |
| Non-HDL-C (mmol/L) | 2.93 ± 0.13 | 4.71 ± 0.14 ### | 4.10 ± 0.18 $ |
| HTR (%) | 20.96 ± 1.19 | 19.73 ± 1.09 | 21.27 ± 1.28 |
| AI | 3.89 ± 0.26 | 4.20 ± 0.33 | 3.82 ± 0.30 |
| Apo A-1 (mg/dL) | 83.02 ± 1.16 | 78.65 ± 0.64 # | 81.02 ± 1.03 |
| Apo B (mg/dL) | 6.88 ± 0.39 | 10.29 ± 1.96 | 5.82 ± 0.69 |
| Apo A-1/Apo B | 12.06 ± 0.59 | 7.64 ± 1.31 # | 13.92 ± 1.78 $ |
| Primer | Primer Direction | Sequence |
|---|---|---|
| Gapdh: Glyceraldehyde-3-phosphate dehydrogenase | Forward Reverse | 5′-AAGGTCATCCCAGAGCTGAA-3′ 5′-CTGCTTCACCACCTTCTTGA-3′ |
| Abcg5: ATP-binding cassette sub-family G member 5 | Forward Reverse | 5′-AGAGGGCCTCACATCAACAGA-3′ 5′-CTGACGCTGTAGGACACATGC-3′ |
| Abcg8: ATP-binding cassette sub-family G member 8 | Forward Reverse | 5′-TGGTCAGTCCAACACTCTGG-3′ 5′-ACTGGGTTGCCCATTTATCC-3′ |
| Acat: Acetyl-coenzyme A acetyltransferase | Forward Reverse | 5′-AGAAATCAAGCAAAGGTCCA-3′ 5′-AGGAGTCCTTGGGTAGTTGT-3′ |
| Acc1: Acetyl-CoA carboxylase 1 | Forward Reverse | 5′-GCCTCTTCCTGACAAACGAG-3′ 5′-TGACTGCCGAAACATCTCTG-3′ |
| Adrb3: Adrenergic receptor beta 3 | Forward Reverse | 5′-ACCAACGTGTTCGTGACT-3′ 5′-ACAGCTAGGTAGCGGTCC-3′ |
| Apob48: Apolipoprotein B 48 | Forward Reverse | 5′-TGGCTCTGATCCCAAATCCCT-3′ 5′-CCGTGCATTCATTGTCGATCT-3′ |
| Cidea: Cell death-inducing DNA fragmentation factor alpha subunit-like effector A | Forward Reverse | 5′-TTTCAAACCATGACCGAAGTAGCC-3′ 5′-CCTCCAGCACCAGCGTAACC-3′ |
| Cpt1a: Carnitine palmitoyl-CoA transferase 1-alpha | Forward Reverse | 5′-ATCTGGATGGCTATGGTCAAGGTC-3′ 5′-GTGCTGTCATGCGTTGGAAGTC-3′ |
| Cpt1b: Carnitine palmitoyl-CoA transferase 1-beta | Forward Reverse | 5′-TGCCTTTACATCGTCTCCAA-3 5′-AGACCCCGTAGCCATCATC-3′ |
| Cpt2: Carnitine palmitoyl-CoA transferase 2 | Forward Reverse | 5′-CAACTCGTATACCCAAACCCAGTC-3′ 5′-GTTCCCATCTTGATCGAGGACATC-3′ |
| C/ebpb: CCAAT/enhancer-binding protein beta | Forward Reverse | 5′-GGAGACGCAGCACAAGGT-3′ 5′-AGCTGCTTGAACAAGTTCCG-3′ |
| Fas: Fatty acid synthase | Forward Reverse | 5′-GCTGCGGAAACTTCAGGAAAT-3′ 5′-AGAGACGTGTCACTCCTGGACTT-3′ |
| Lipe: Lipase E | Forward Reverse | 5′-GGCTCACAGTTACCATCTCACC-3′ 5′-GAGTACCTTGCTGTCCTGTCC-3′ |
| Pgc1a: Peroxisome proliferator-activated receptor gamma coactivator 1-alpha | Forward Reverse | 5′-AAGTGTGGAACTCTCTGGAACTG-3′ 5′-GGGTTATCTTGGTTGGCTTTATG-3′ |
| Ppara: Peroxisome proliferator-activated receptor alpha | Forward Reverse | 5′-CCTGAACATCGAGTGTCGAATAT-3′ 5′-GGTCTTCTTCTGAATCTTGCAGCT-3′ |
| Pparg: Peroxisome proliferator-activated receptor gamma | Forward Reverse | 5′-GCATGGTGCCTTCGCTGA-3′ 5′-TGGCATCTCTGTGTCAACCATG-3′ |
| Prdm16: PR domain containing 16 | Forward Reverse | 5′-CAGCACGGTGAAGCCATTC-3′ 5′-GCGTGCATGCGCTTGTG-3′ |
| Prkag1: Protein kinase AMP-activated non-catalytic subunit gamma 1 | Forward Reverse | 5′-TCTCCGCCTTACCTGTAGTGGA-3′ 5′-GCAGGGCTTTTGTCACAGACAC-3′ |
| Prkag2: Protein kinase AMP-activated non-catalytic subunit gamma 2 | Forward Reverse | 5′-CTCCTCATCCAAAGAGTCTTCGC-3′ 5′-TGGGTGTTGACGGAGAAGAGGA-3′ |
| Scd1: Steroly-CoA desaturase 1 | Forward Reverse | 5′-CCCCTGCGGATCTTCCTTAT-3′ 5′-AGGGTCGGCGTGTGTTTCT-3′ |
| Sirt1: Sirtuin 1 | Forward Reverse | 5′-TGTGAAGTTACTGCAGTGTAA-3′ 5′-GCATAGATACCGTCTCTTGATCTG-3′ |
| Ucp1: Uncoupling protein 1 | Forward Reverse | 5′-ACTGCCACCCCTCCAGTCATT-3′ 5′-CTTTGCCTCACTGAGGATTGG-3′ |
| Ucp2: Uncoupling protein 2 | Forward Reverse | 5′-ACCAAGGGCTCAGAGCATGCA-3′ 5′-TGGCTTTCAGGAGAGTATCTTTG-3′ |
| Ucp3: Uncoupling protein 3 | Forward Reverse | 5′-GGATTTGTGCCCTCCTTTCTG-3′ 5′-AGATTCCCGCAGTACCTGGAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-W.; Yang, Y.-M.; Kwon, E.-Y.; Choi, J.-Y. Novel Plant Extract Ameliorates Metabolic Disorder through Activation of Brown Adipose Tissue in High-Fat Diet-Induced Obese Mice. Int. J. Mol. Sci. 2022, 23, 9295. https://doi.org/10.3390/ijms23169295
Kim J-W, Yang Y-M, Kwon E-Y, Choi J-Y. Novel Plant Extract Ameliorates Metabolic Disorder through Activation of Brown Adipose Tissue in High-Fat Diet-Induced Obese Mice. International Journal of Molecular Sciences. 2022; 23(16):9295. https://doi.org/10.3390/ijms23169295
Chicago/Turabian StyleKim, Ji-Won, Young-Mo Yang, Eun-Young Kwon, and Ji-Young Choi. 2022. "Novel Plant Extract Ameliorates Metabolic Disorder through Activation of Brown Adipose Tissue in High-Fat Diet-Induced Obese Mice" International Journal of Molecular Sciences 23, no. 16: 9295. https://doi.org/10.3390/ijms23169295
APA StyleKim, J.-W., Yang, Y.-M., Kwon, E.-Y., & Choi, J.-Y. (2022). Novel Plant Extract Ameliorates Metabolic Disorder through Activation of Brown Adipose Tissue in High-Fat Diet-Induced Obese Mice. International Journal of Molecular Sciences, 23(16), 9295. https://doi.org/10.3390/ijms23169295