
RNA-As-Graphs Motif Atlas—Dual Graph Library of RNA Modules and Viral Frameshifting-Element Applications



Abstract
:1. Introduction
2. Results
2.1. RNA Database
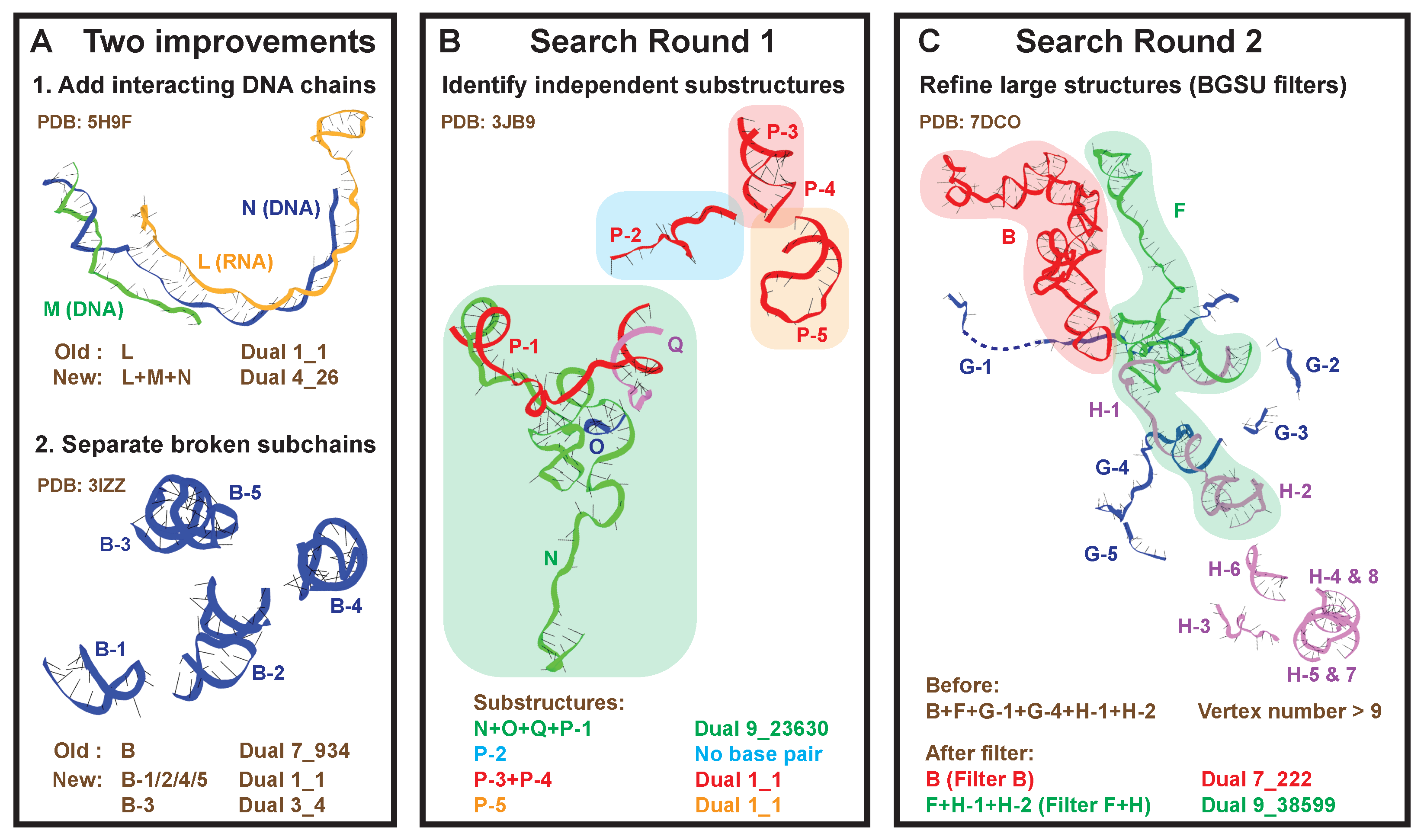
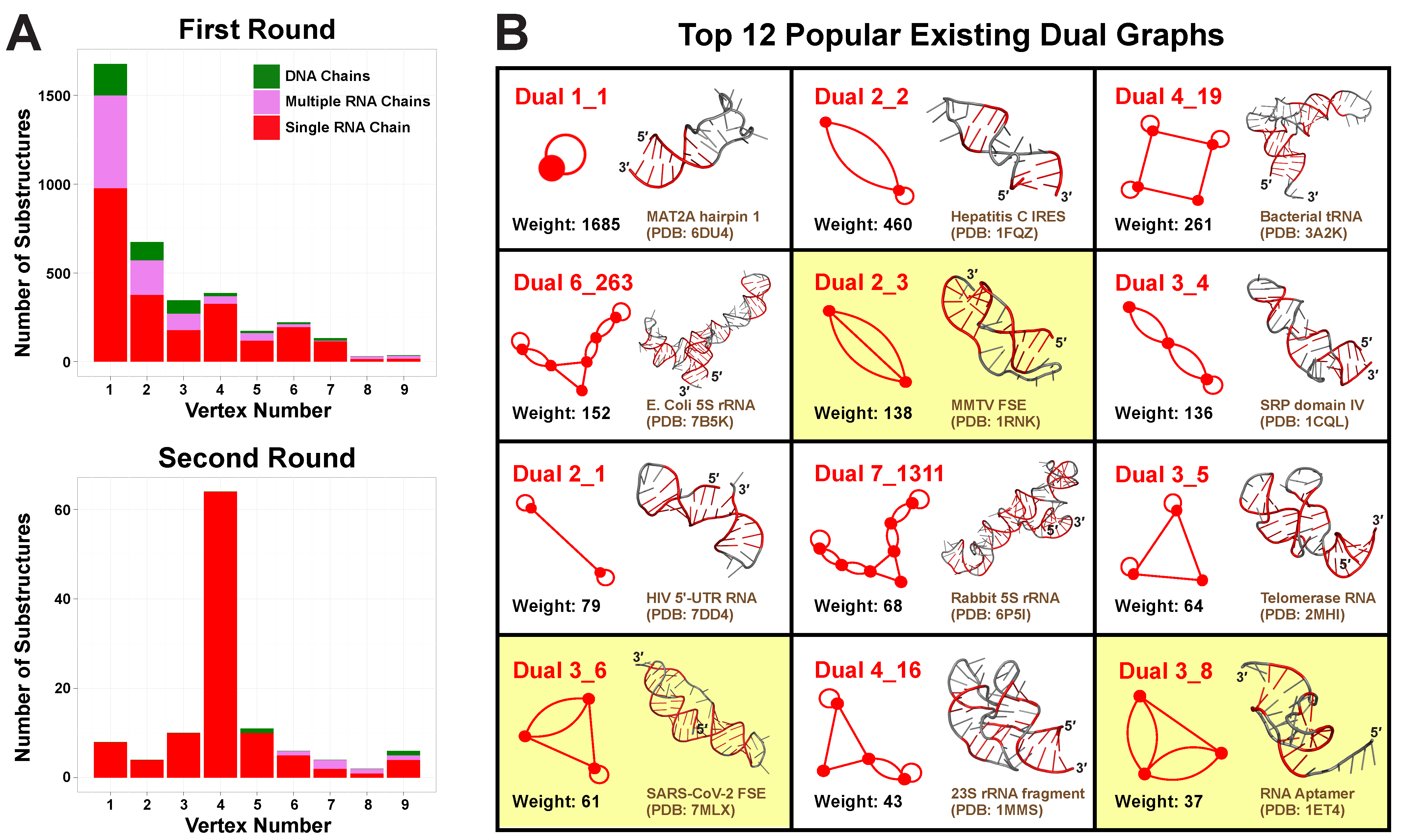
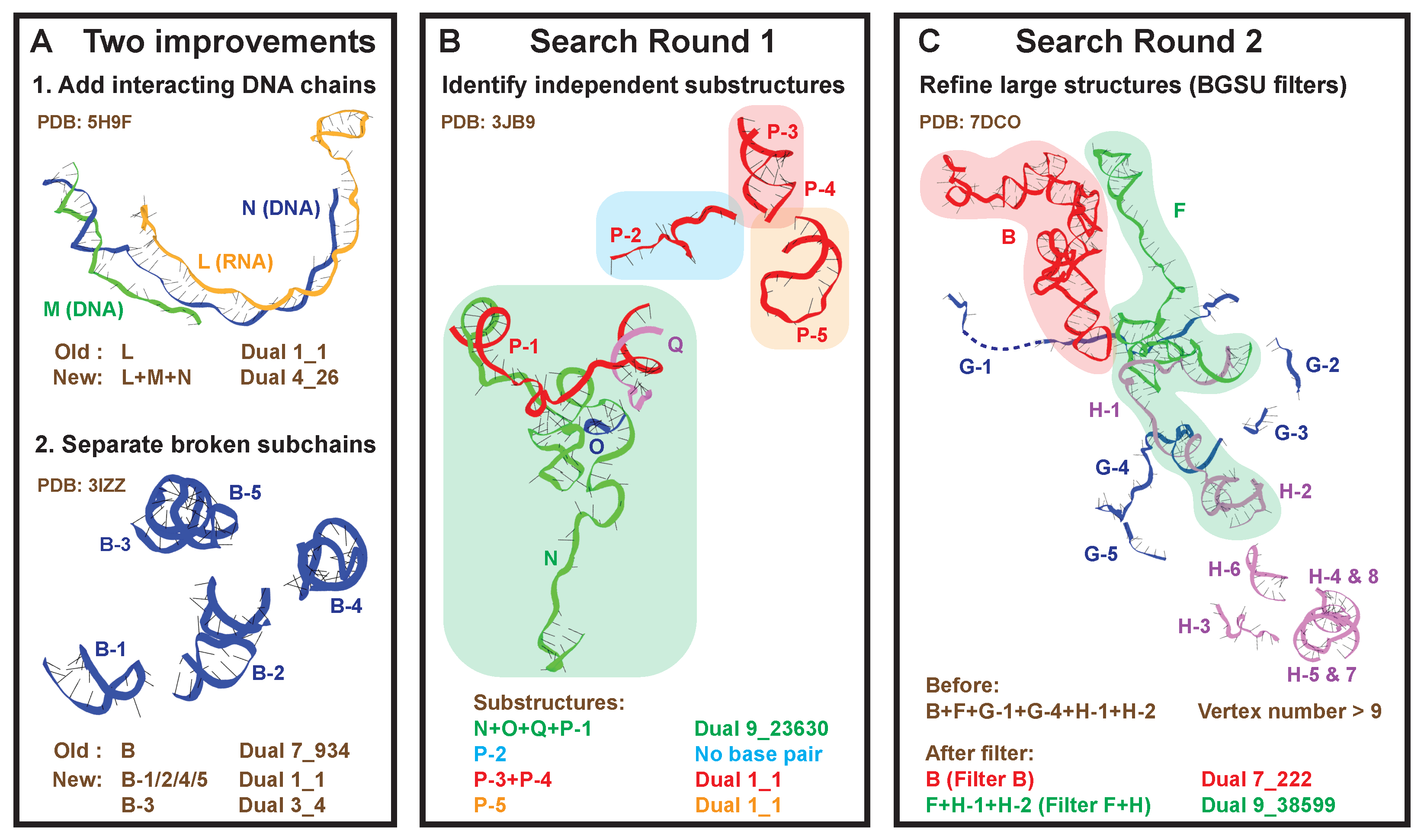
2.2. Round 1: Substructure Search
2.3. Round 2: Add Refinement for Large Substructures
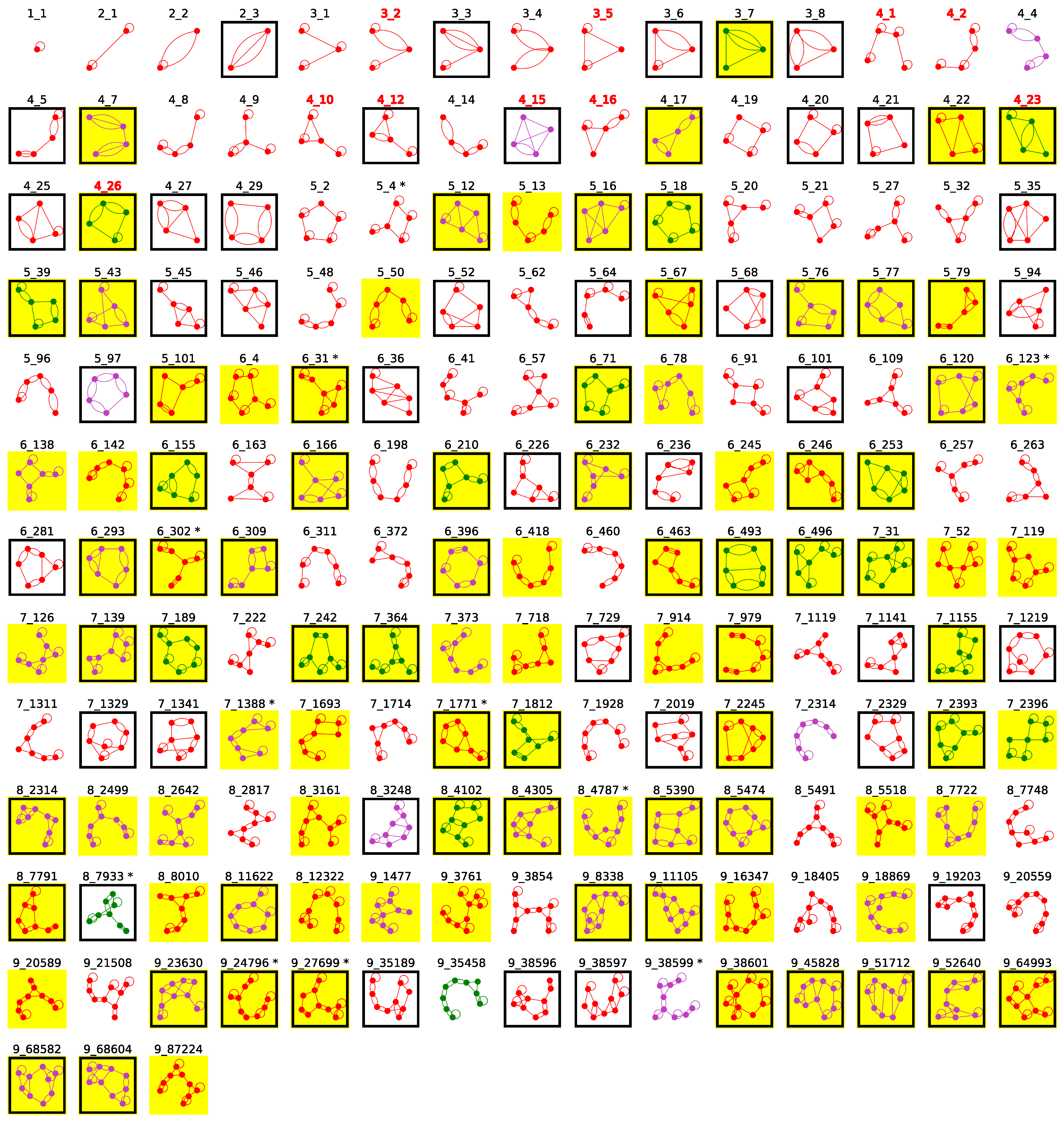
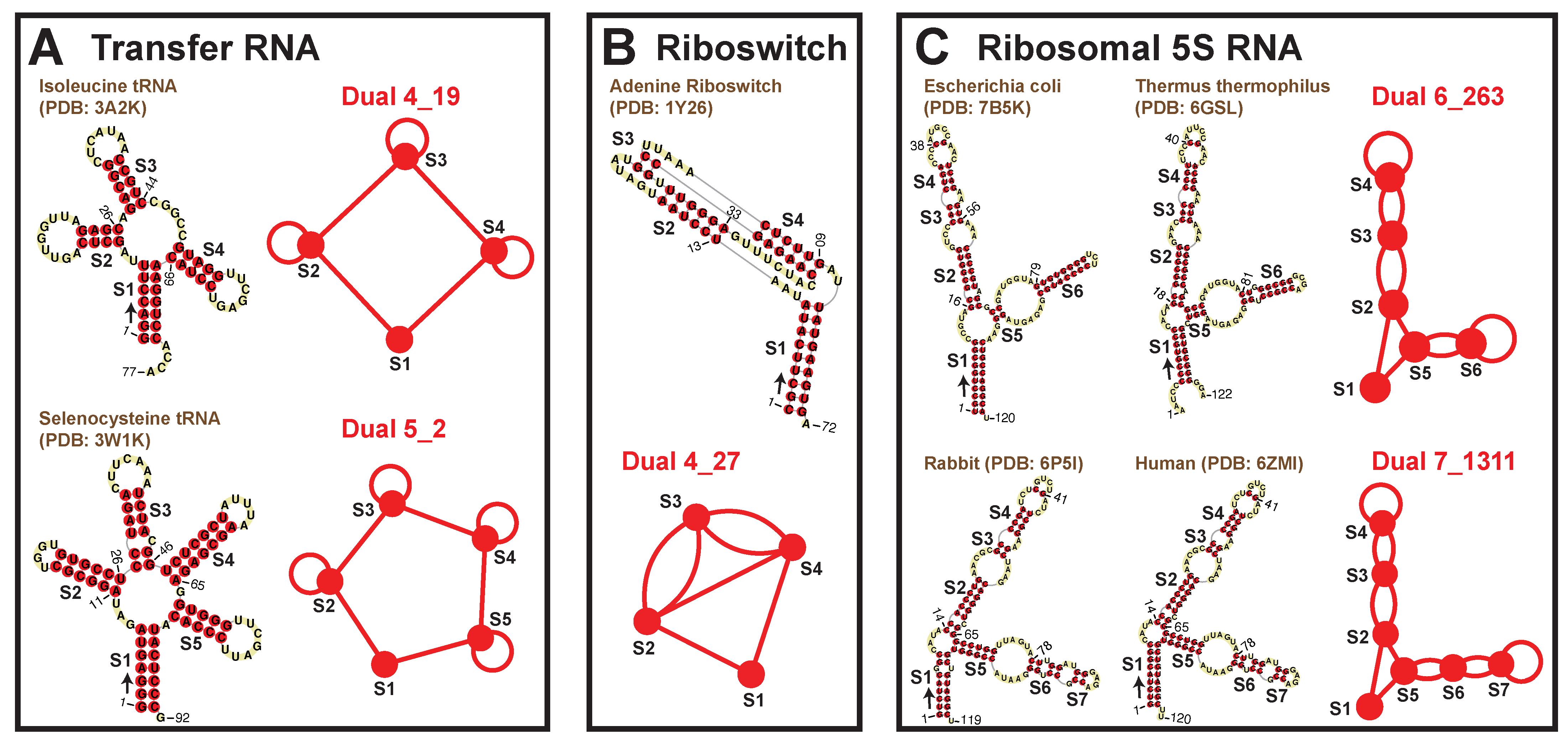
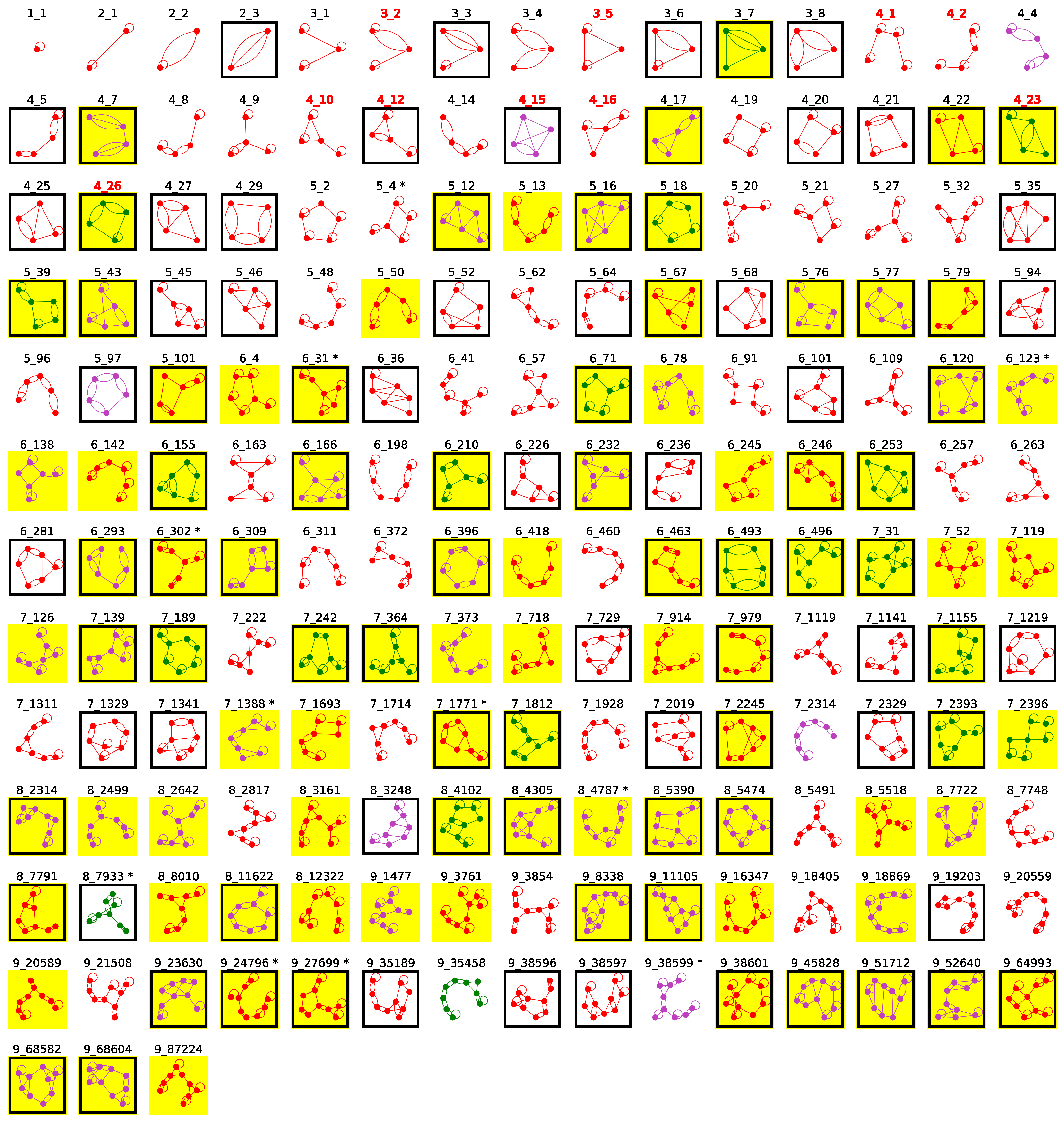
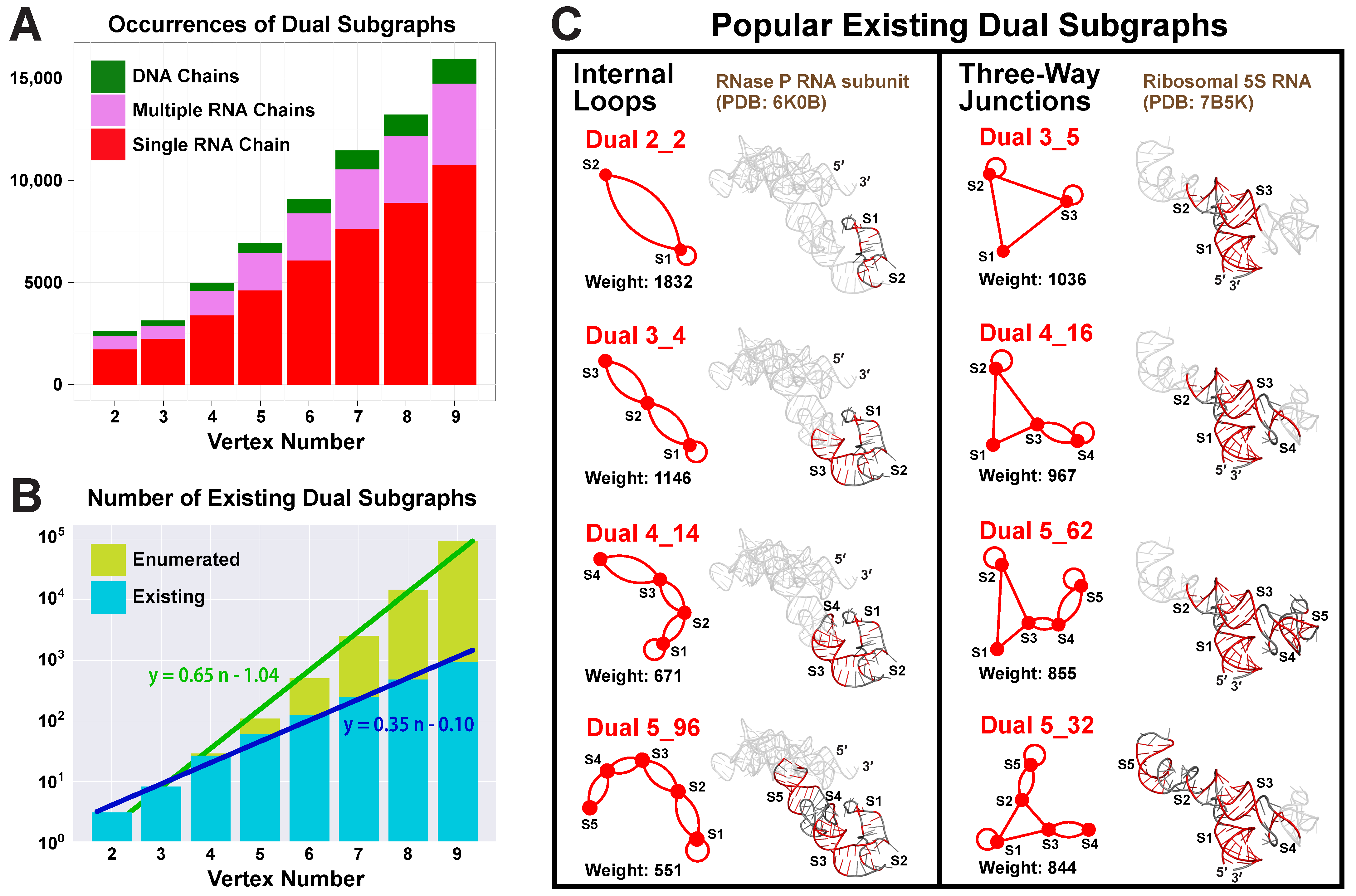
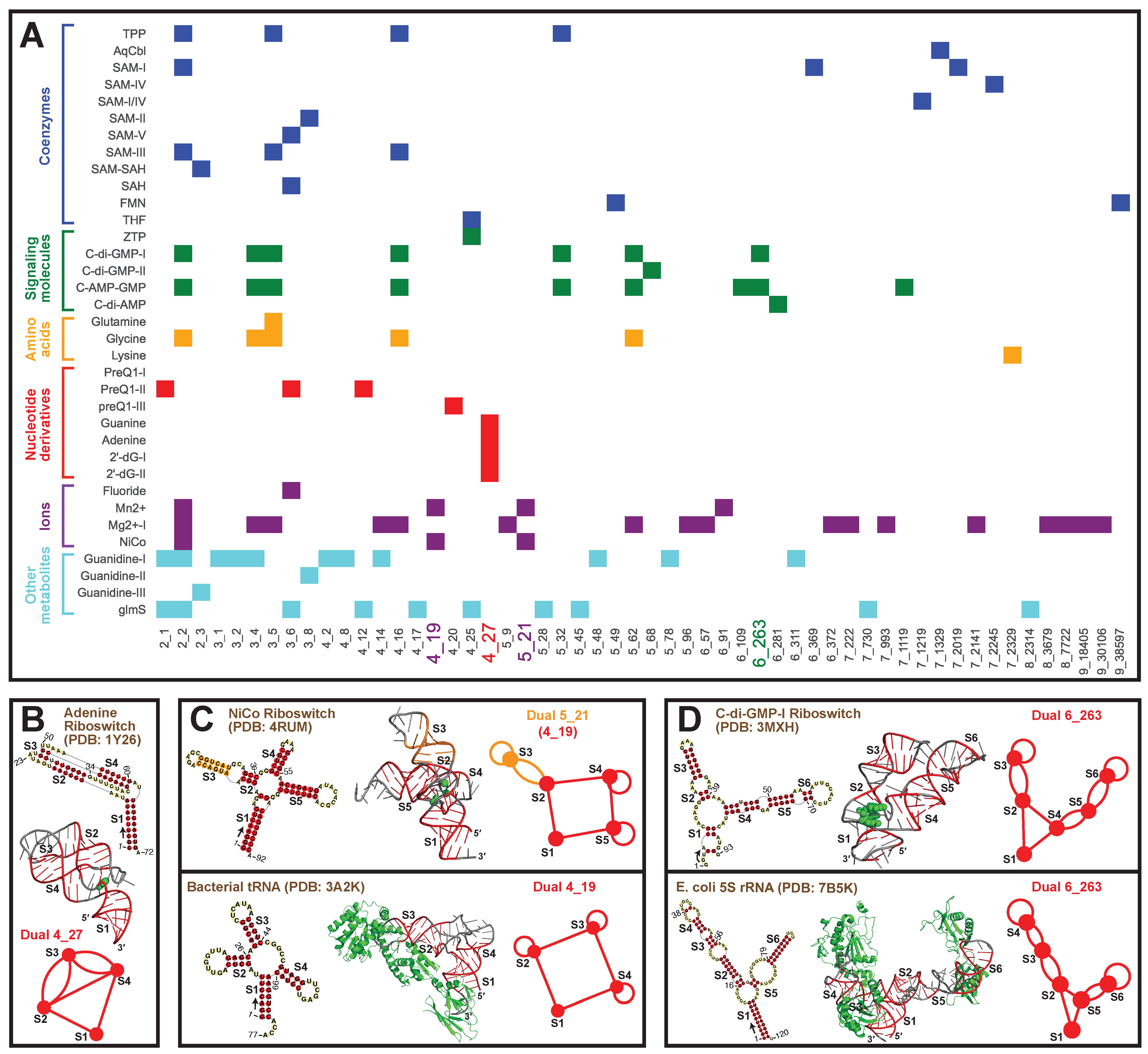
2.4. Popular RNA Motifs
2.5. Updated Existing Dual Graph Library
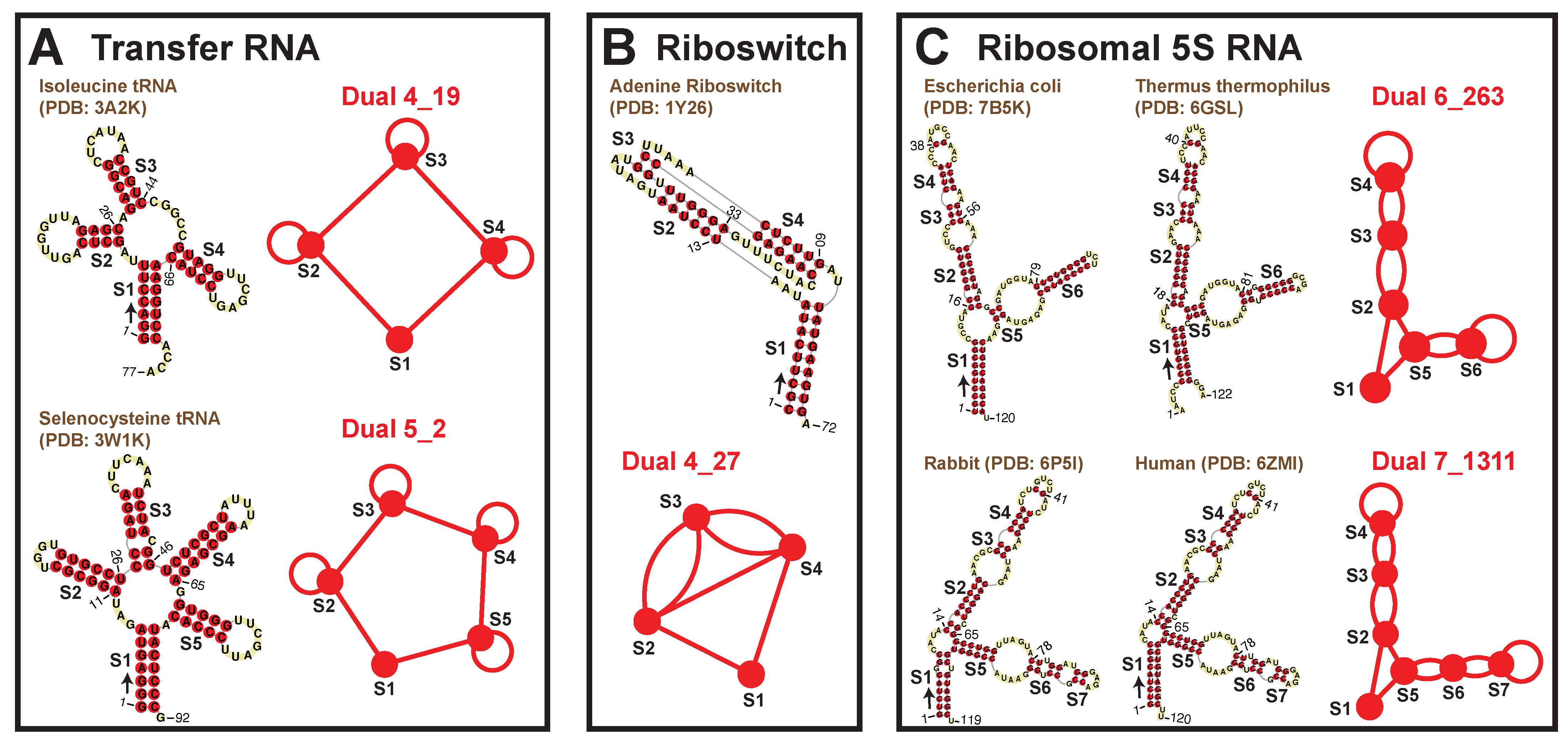
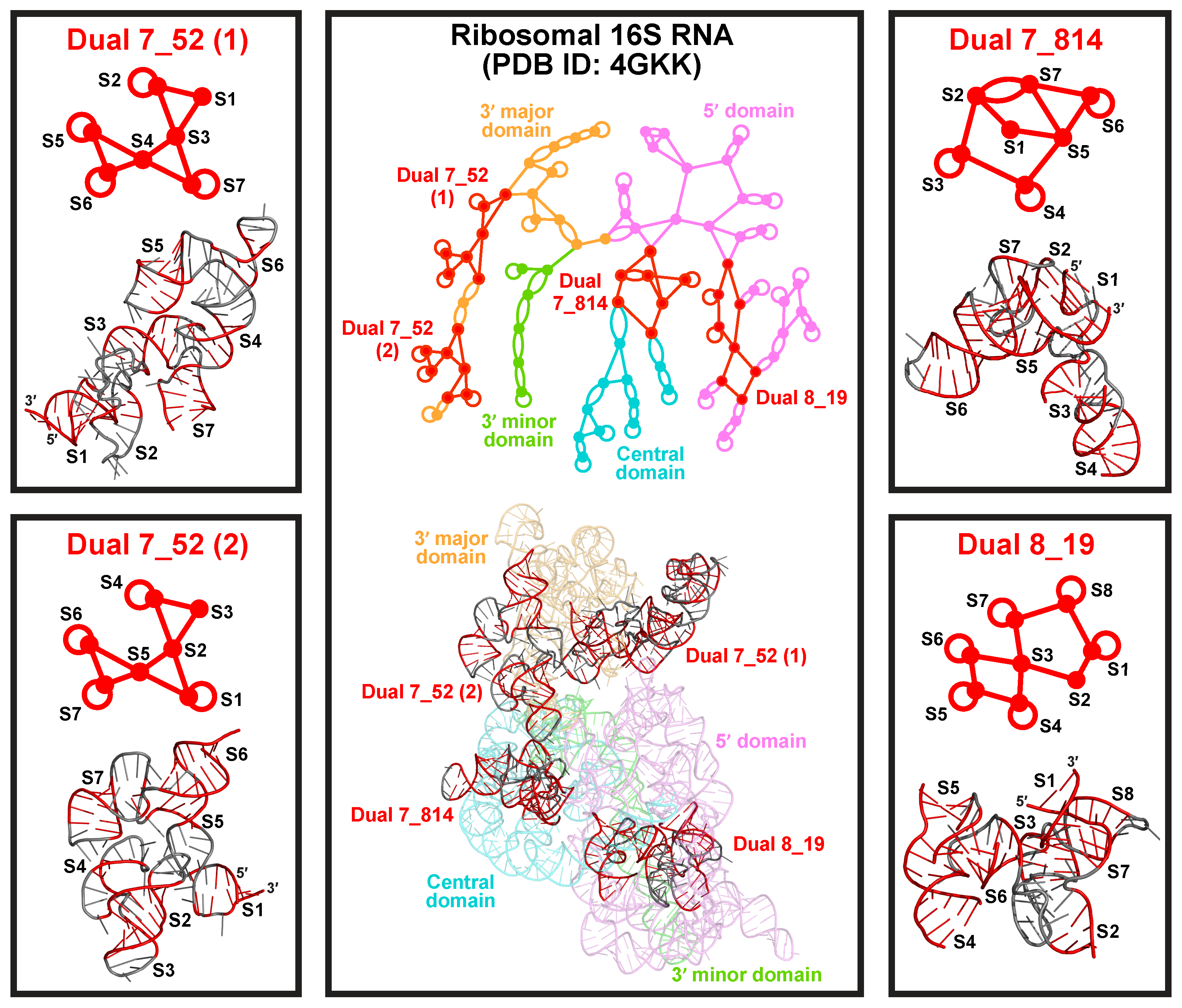
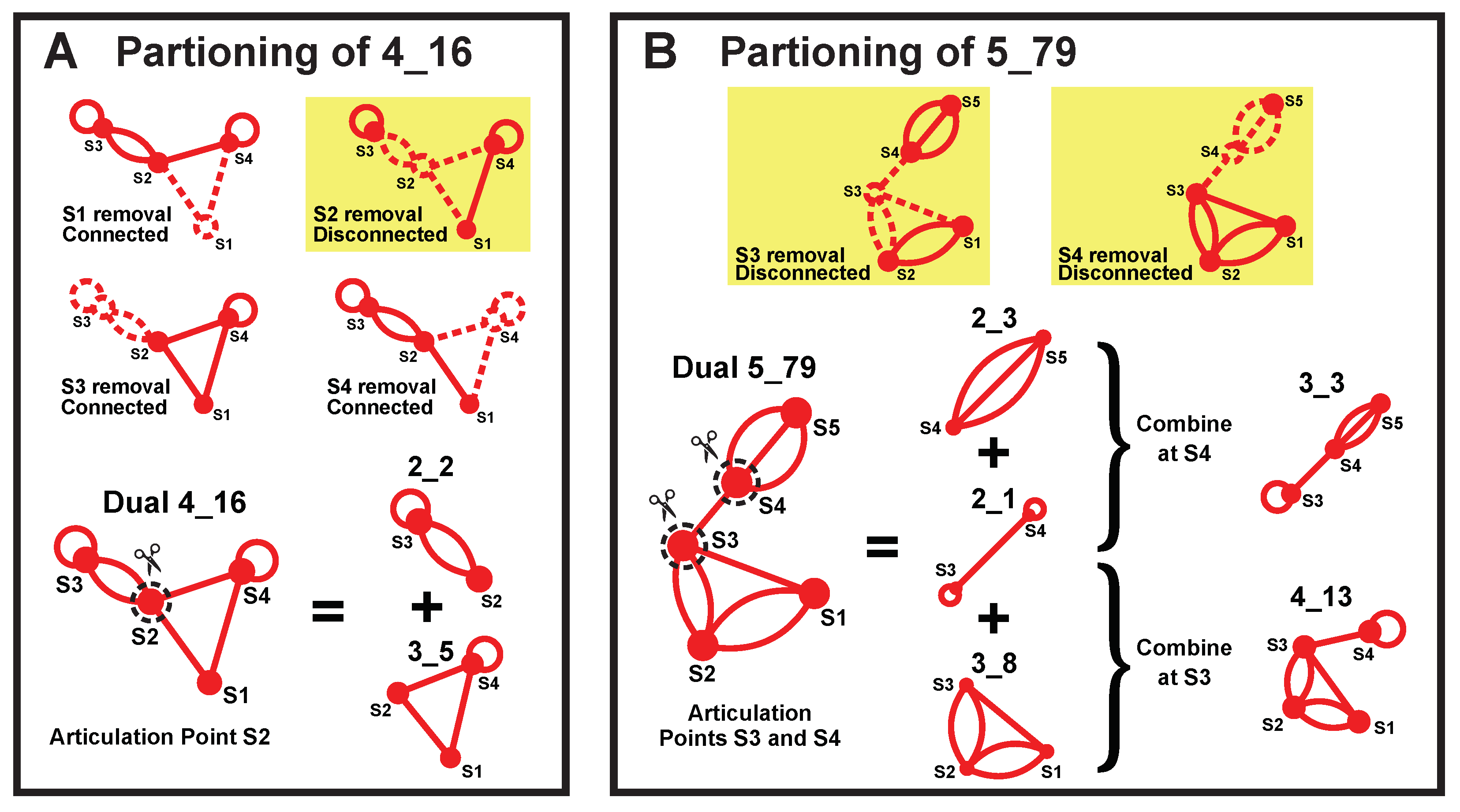
2.6. Subgraphs of Existing Dual Graph Motifs
2.7. Popular RNA Submotifs
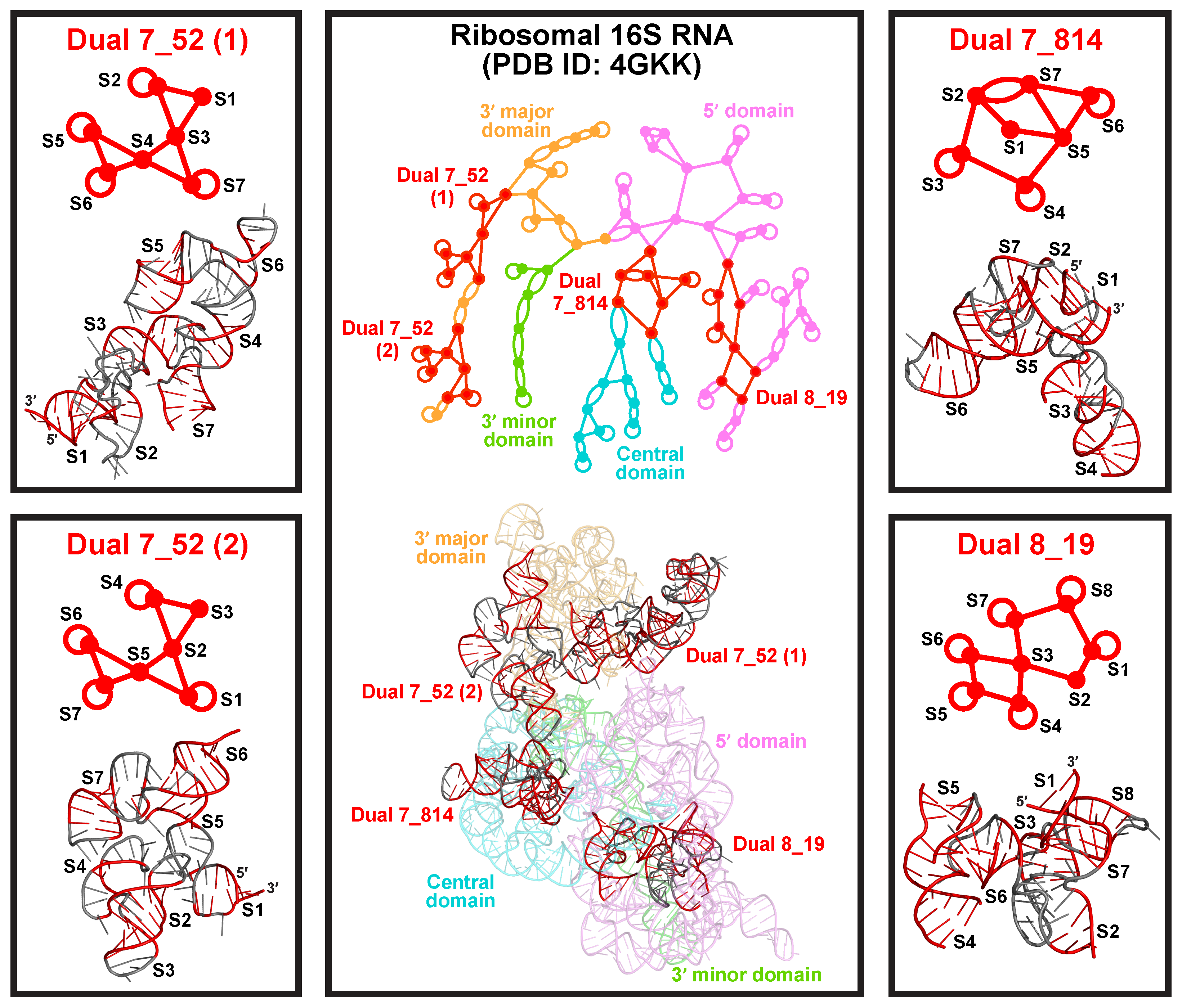
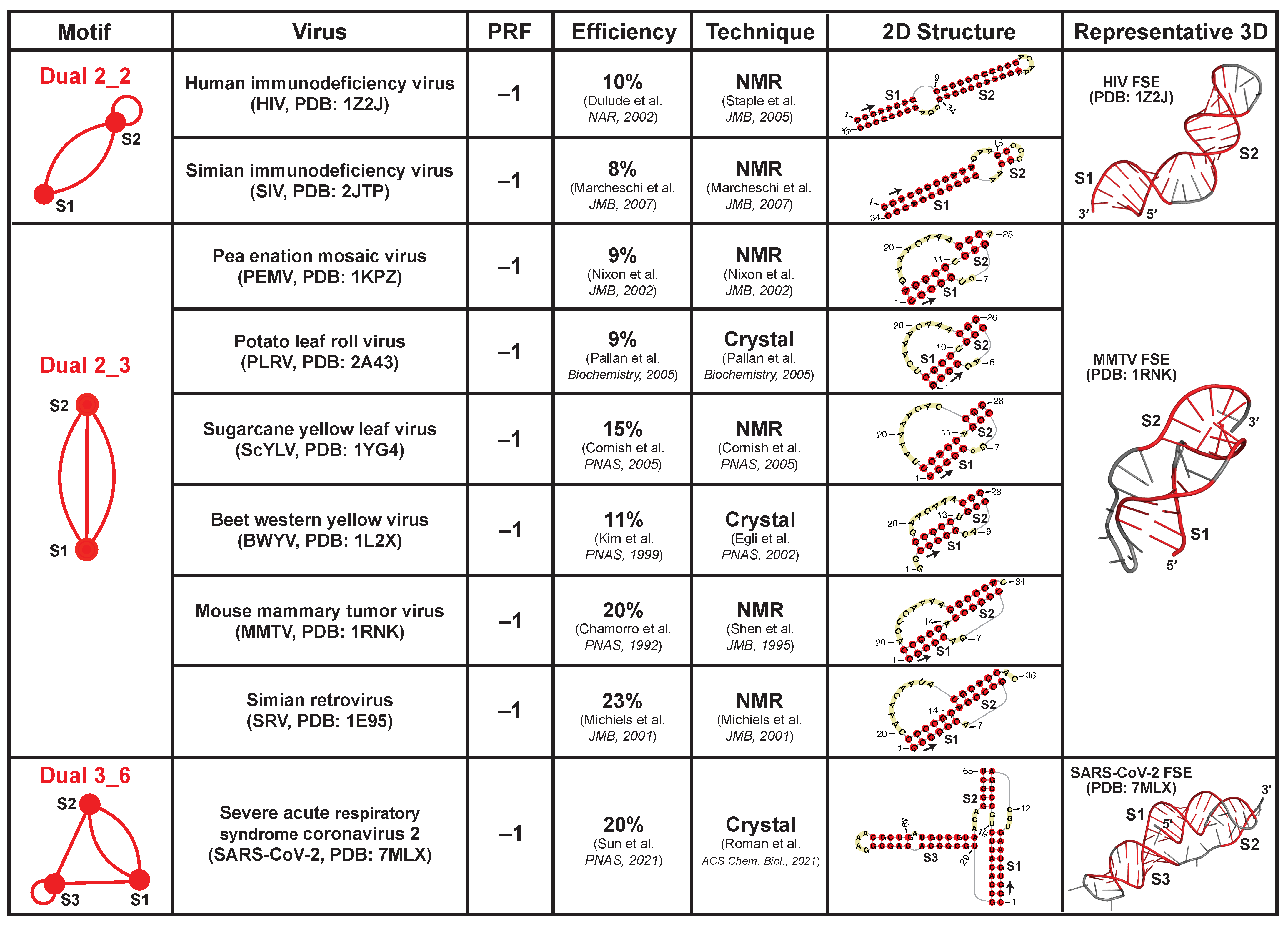
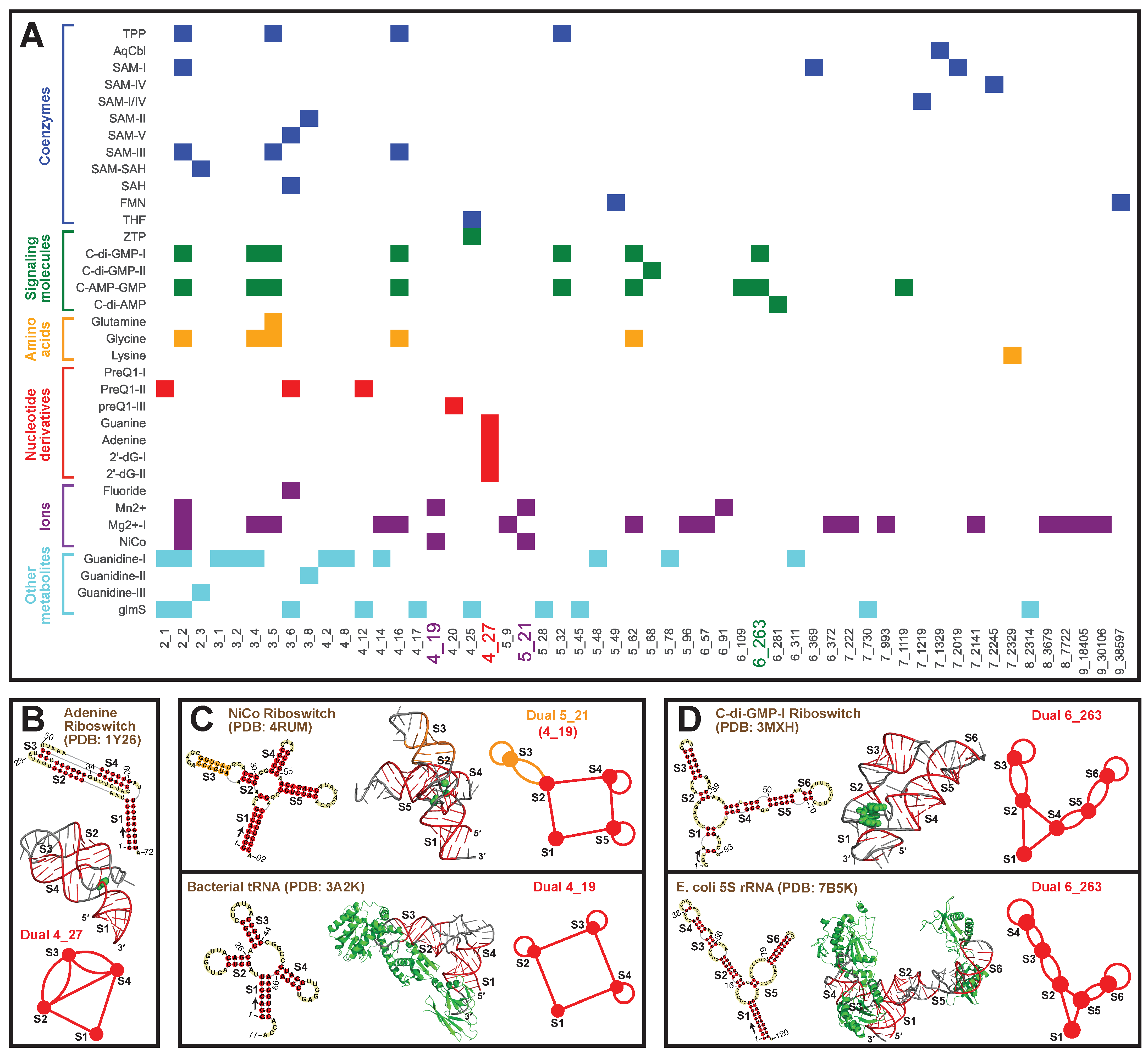
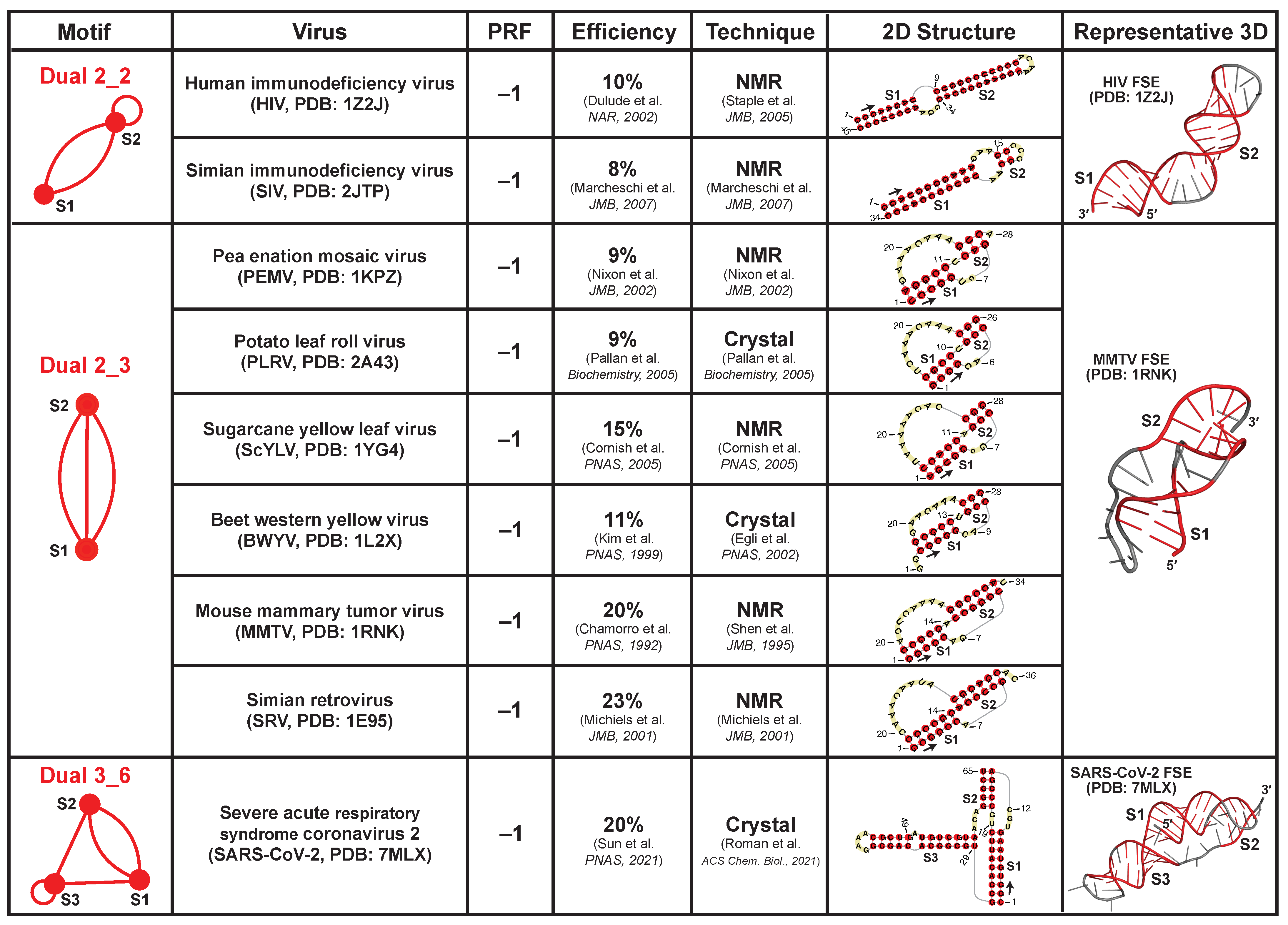
2.8. Applications: Delineating Frameshifting Element and Riboswitch Motifs
3. Discussion
4. Materials and Methods
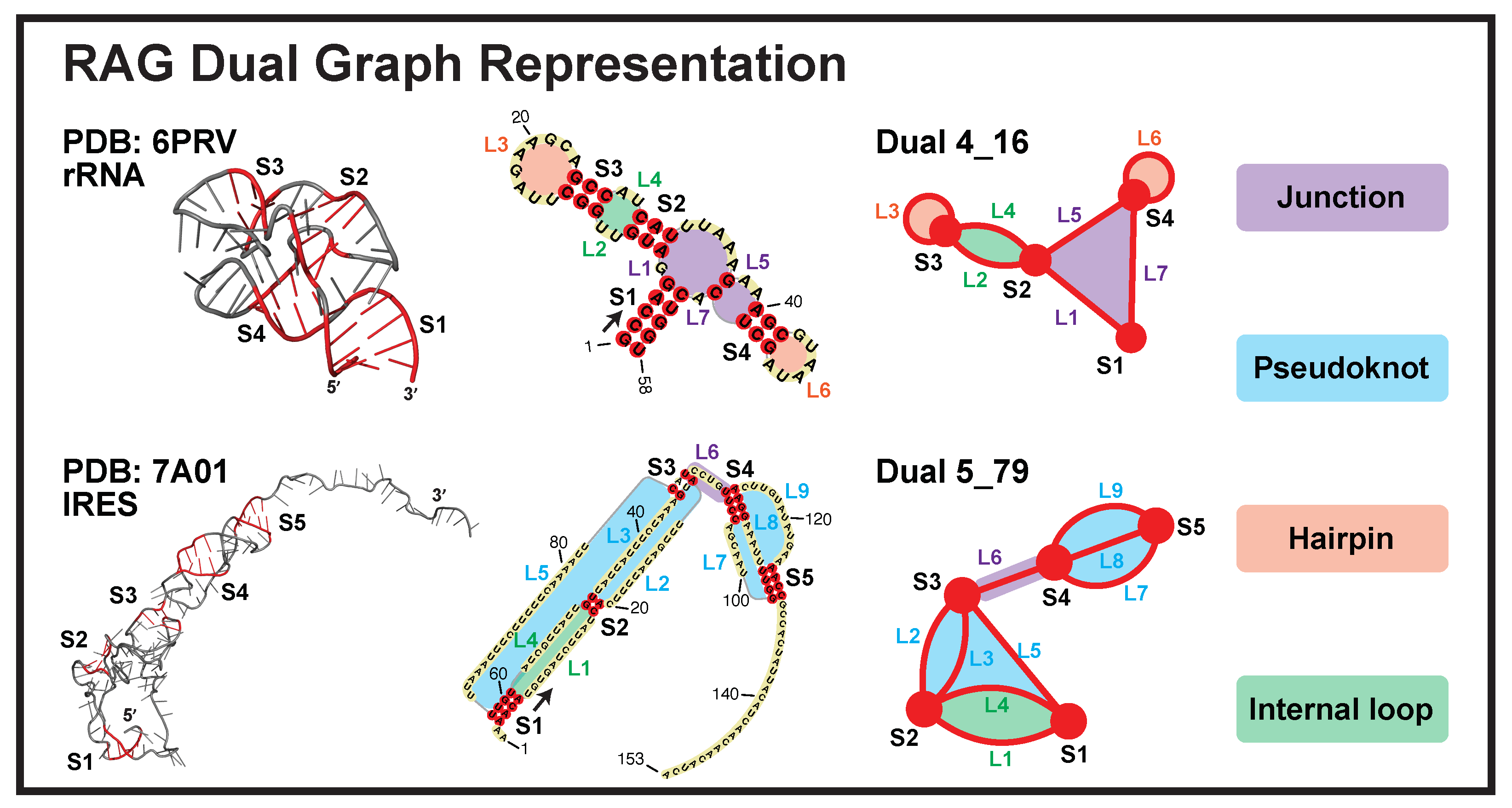
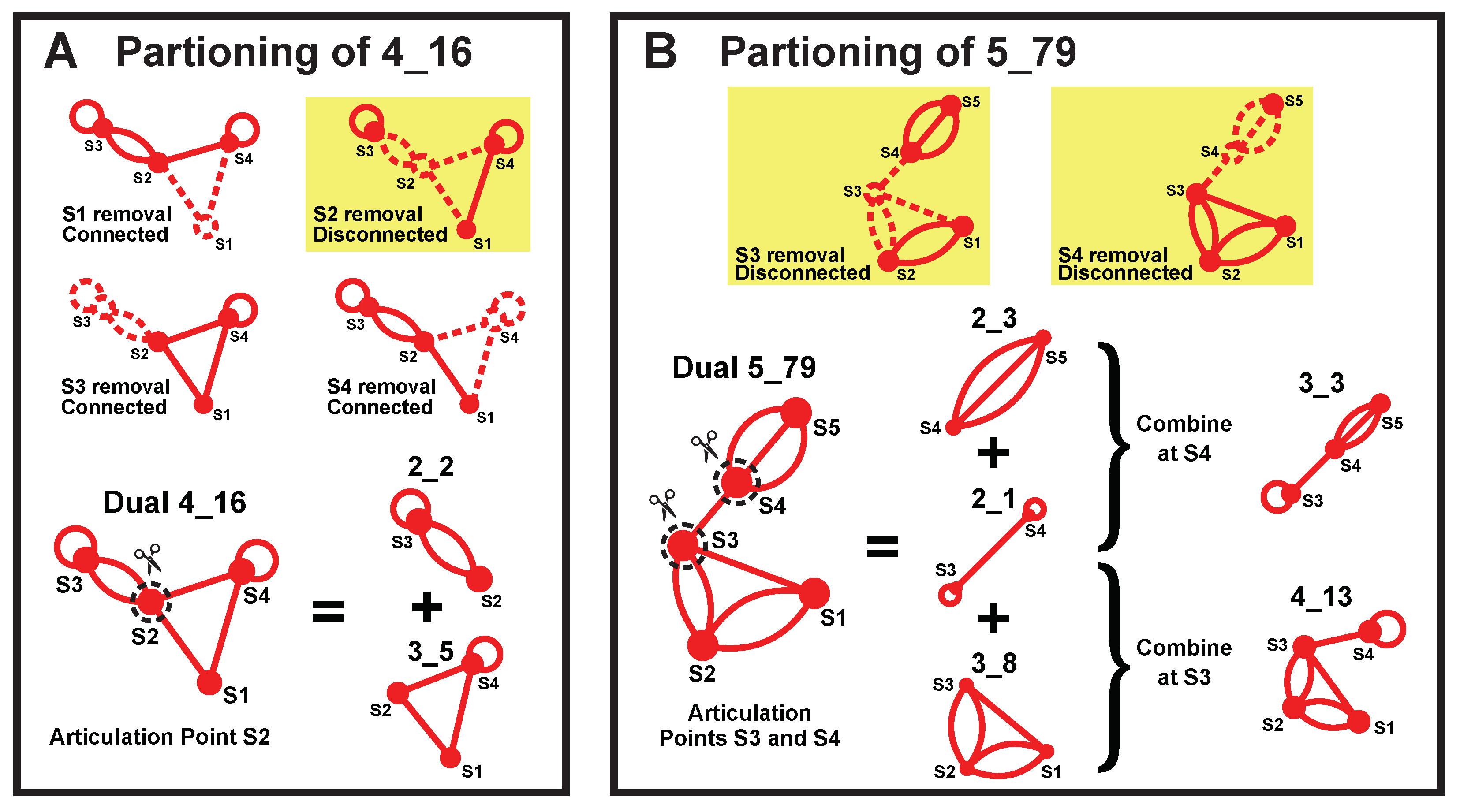
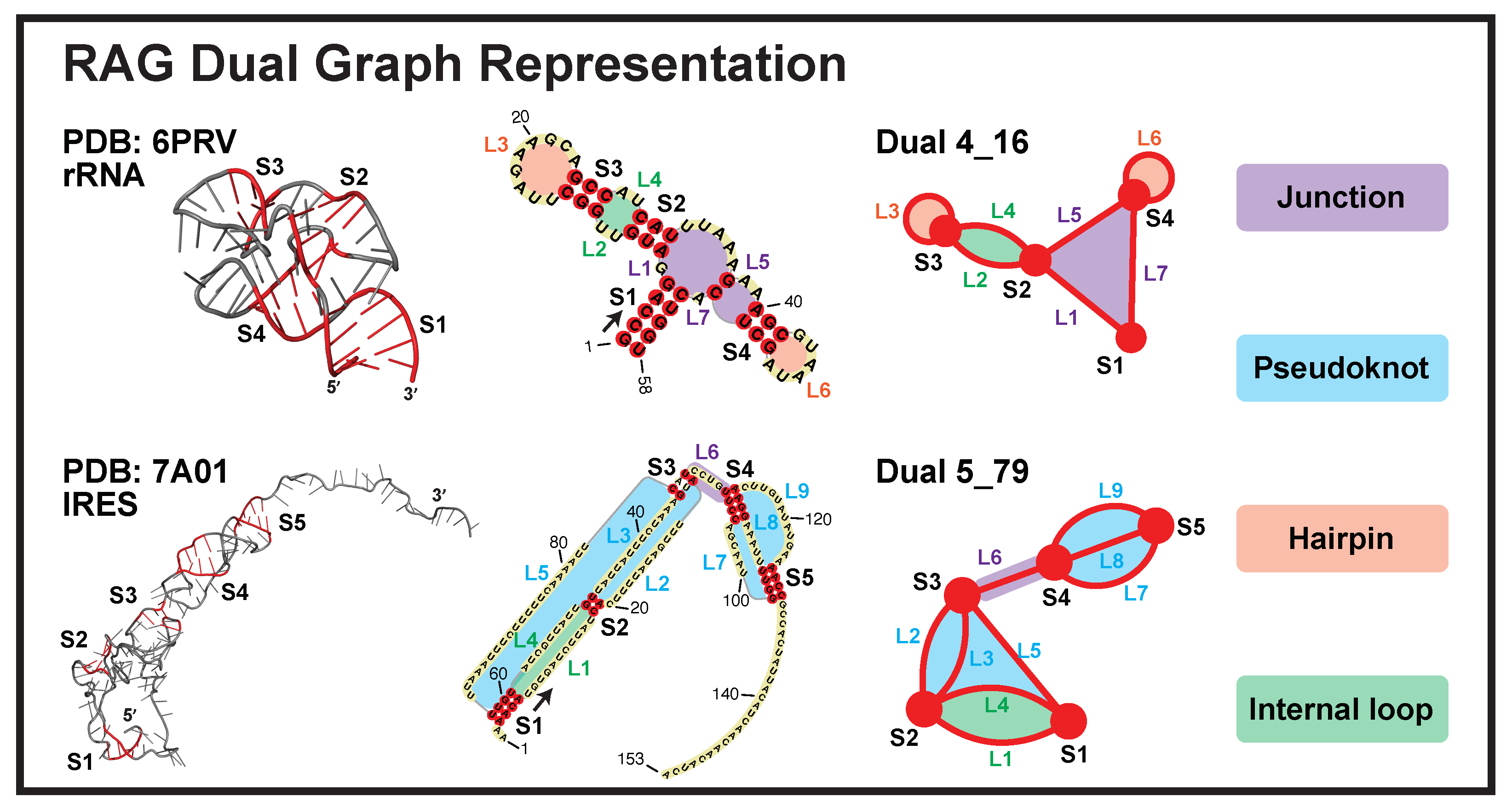
4.1. Dual Graph Definitions
- Each junction loop is denoted as an edge (e.g., purple loops in Figure 1).
- Each single strand in a pseudoknot is denoted as an edge (e.g., blue loops).
- Hairpin loops are denoted as self-edges (e.g., orange loops).
- A single-stranded internal loop/bulge is denoted as two edges (e.g., green loops). A bulge of 1-nt or an internal loop of 1-nt on both sides is ignored.
- A stem of ≥2 consecutive base pairs (i.e., no loop in between) is denoted as a vertex. An isolated single base pair is ignored.
- The dangling 5 and 3 ends are ignored.
4.2. RAG Library
4.3. New Substructure Search Algorithm
- 1.
- Identify the set of all subchains C using 3DNA-DSSR [30]. For each subchain x, find the set of subchains that interacts with it (>1 base pairs). Subchains that have >92% sequence and 2D structure similarity to subchain x are not included in to avoid polymers.
- 2.
- For each subchain x not assigned to any substructure, set its initial substructure , and add subchains that interact with any subchain in . For those newly added subchains, again include their interacting subchains if not contained in yet. This process is repeated until no new subchains are added. Below is the corresponding pseudocode (Algorithm 1):
| Algorithm 1 Pseudocode for identifying independently folded nucleic acid substructures. |
| Input: Set of all subchains C, Set of interacting subchains for each subchain x |
| Output: Independently folded |
| ←C ▹ records all subchains not assigned to substructures |
| forx in C do |
| if x in then ▹ Find the substructure containing subchain x |
| ▹ records newly added subchains in each loop below |
| while not empty do |
| ▹ updates the substructure in each loop |
| for z in do |
| ▹ Include interacting subchains for each newly added subchain |
| end for |
| end while |
| ← ▹ Remove subchains in from |
| Add to |
| end if |
| end for |
4.4. Subgraph Partitioning
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fire, A.; Xu, S.; Montgomery, M.; Kostas, S.; Driver, S.; Mello, C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Ma, Y.; Chan, C.; He, M. RNA interference and antiviral therapy. World J. Gastroenterol. 2007, 13, 5169–5179. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.; Yang, C.; Ueda, M.; Kristen, A.; Tournev, M.D.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. Spinal muscular atrophy approval boosts antisense drugs. Nat. Biotechnol. 2017, 35, 99–100. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.R.; Rodino-Klapac, L.R.; Sahenk, Z.; Roush, K.; Bird, L.; Lowes, L.P.; Alfano, L.; Gomez, A.M.; Lewis, S.; Kota, J.; et al. Eteplirsen for the treatment of Duchenne muscular dystrophy. Ann. Neurol. 2013, 74, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, J.; Kowalski, P.; Anderson, D. Advances in the delivery of RNA therapeutics: From concept to clinical reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef]
- Fellmann, C.; Gowen, B.G.; Lin, P.C.; Doudna, J.A.; Corn, J.E. Cornerstones of CRISPR–Cas in drug discovery and therapy. Nat. Rev. Drug. Discov. 2017, 16, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Keefe, A.; Pai, S.; Ellington, A. Aptamers as therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550. [Google Scholar] [CrossRef]
- Waterman, M.; Smith, T. RNA secondary structure: A complete mathematical analysis. Math. Biosci. 1978, 42, 257–266. [Google Scholar] [CrossRef]
- Shapiro, B. An algorithm for comparing multiple RNA secondary structures. Bioinformatics 1988, 4, 387–393. [Google Scholar] [CrossRef]
- Le, S.; Owens, J.; Nussinov, R.; Chen, J.; Shapiro, B.; Maizel, J. RNA secondary structures: Comparison and determination of frequently recurring substructures by consensus. Bioinformatics 1989, 5, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Pasquali, S.; Schlick, T. Exploring the repertoire of RNA secondary motifs using graph theory; implications for RNA design. Nucleic Acids Res. 2003, 31, 2926–2943. [Google Scholar] [CrossRef]
- Kim, N.; Laing, C.; Elmetwaly, S.; Jung, S.; Curuksu, J.; Schlick, T. Graph-based sampling for approximating global helical topologies of RNA. Proc. Natl. Acad. Sci. USA 2014, 111, 4079–4084. [Google Scholar] [CrossRef]
- Jain, S.; Saju, S.; Petingi, L.; Schlick, T. An extended dual graph library and partitioning algorithm applicable to pseudoknotted RNA structures. Methods 2019, 162, 74–84. [Google Scholar] [CrossRef]
- Baba, N.; Elmetwaly, S.; Kim, N.; Schlick, T. Predicting large RNA-like topologies by a knowledge-based clustering approach. J. Mol. Biol. 2016, 428, 811–821. [Google Scholar] [CrossRef]
- Zhu, Q.; Schlick, T. A fiedler vector scoring approach for novel RNA motif selection. J. Phys. Chem. B 2021, 125, 1144–1155. [Google Scholar] [CrossRef]
- Jain, S.; Laederach, A.; Ramos, S.; Schlick, T. A pipeline for computational design of novel RNA-like topologies. Nucleic Acids Res. 2018, 46, 7040–7051. [Google Scholar] [CrossRef]
- Kim, N.; Zheng, Z.; Elmetwaly, S.; Schlick, T. RNA graph partitioning for the discovery of RNA modularity: A novel application of graph partition algorithm to biology. PLoS ONE 2014, 9, e106074. [Google Scholar] [CrossRef]
- Petingi, L.; Schlick, T. Partitioning and classification of RNA secondary structures into pseudonotted and pseudoknot-free regions using a graph- theoretical approach. IAENG Int. J. Comput. Sci. 2017, 44, 241–246. [Google Scholar]
- Jain, S.; Schlick, T. F-RAG: Generating atomic models from RNA graphs using fragment assembly. J. Mol. Biol. 2017, 429, 3587–3605. [Google Scholar] [CrossRef]
- Jain, S.; Tao, Y.; Schlick, T. Inverse folding with RNA-As-Graphs produces a large pool of candidate sequences with target topologies. J. Struct. Biol. 2020, 209, 107438. [Google Scholar] [CrossRef]
- Zahran, M.; Bayrak, C.; Elmetwaly, S.; Schlick, T. RAG-3D: A search tool for RNA 3D substructures. Nucleic Acids Res. 2015, 43, 9474–9488. [Google Scholar] [CrossRef]
- Schlick, T.; Zhu, Q.; Jain, S.; Yan, S. Structure-altering mutations of the SARS-CoV-2 frameshifting RNA element. Biophys. J. 2021, 120, 1040–1053. [Google Scholar] [CrossRef]
- Schlick, T.; Zhu, Q.; Dey, A.; Jain, S.; Yan, S.; Laederach, A. To knot or not to knot: Multiple conformations of the SARS-CoV-2 frameshifting RNA element. J. Amer. Chem. Soc. 2021, 143, 11404–11422. [Google Scholar] [CrossRef]
- Shuting, Y.; Zhu, Q.; Jain, S.; Schlick, T. Length-dependent motions of SARS-CoV-2 frameshifting RNA pseudoknot and alternative conformations suggest avenues for frameshifting suppression. Nat. Commun. 2022, 13, 4284. [Google Scholar]
- Laing, C.; Wen, D.; Wang, J.; Schlick, T. Predicting coaxial helical stacking in RNA junctions. Nucleic Acids Res. 2012, 40, 487–498. [Google Scholar] [CrossRef]
- Hohl, J. Unraveling the Conformational Landscapes for the Frameshifting Element of β-Coronaviruses by Graph Theory and Modeling. Master’s Thesis, Applied Mathematics, Courant Institute of Mathematical Sciences, New York University, New York, NY, USA, 2022. Advisor: T. Schlick. In preparation. [Google Scholar]
- Kim, N.; Shiffeldrim, N.; Gan, H.; Schlick, T. Candidates for novel RNA topologies. J. Mol. Biol. 2004, 341, 1129–1144. [Google Scholar] [CrossRef]
- Leontis, N.; Zirbel, C. Nonredundant 3D structure datasets for RNA knowledge extraction and benchmarking. In RNA 3D Structure Analysis and Prediction; Leontis, N., Zirbel, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 27, pp. 281–298. [Google Scholar]
- Lu, X.; Olson, W. 3dna: A software package for the analysis, rebuilding and visualization of three-dimensional nucleic acid structures. Nucleic Acids Res. 2003, 31, 5108–5121. [Google Scholar] [CrossRef]
- Krahn, N.; Fischer, J.; Söll, D. Naturally occurring tRNAs with non-canonical structures. Front. Microbiol. 2020, 11, 596914. [Google Scholar] [CrossRef]
- Yang, H.; Jossinet, F.; Leontis, N.; Chen, L.; Westbrook, J.; Berman, H.; Westhof, E. Tools for the automatic identification and classification of RNA base pairs. Nucleic Acids Res. 2003, 31, 3450–3460. [Google Scholar] [CrossRef]
- Lemieux, S.; Major, F. RNA canonical and non-canonical base pairing types: A recognition method and complete repertoire. Nucleic Acids Res. 2002, 30, 4250–4263. [Google Scholar] [CrossRef]
- Izzo, J.; Kim, N.; Elmetwaly, S.; Schlick, T. RAG: An update to the RNA-As-Graphs resource. BMC Bioinform. 2011, 12, 291. [Google Scholar] [CrossRef]
- Namy, O.; Moran, S.; Stuart, D.; Gilbert, R.; Brierley, I. A mechanical explanation of RNA pseudoknot function in programmed ribosomal frameshifting. Nature 2006, 441, 244–247. [Google Scholar] [CrossRef]
- Ritchie, D.; Foster, D.; Woodside, M. Programmed 𢀒1 frameshifting efficiency correlates with RNA pseudoknot conformational plasticity, not resistance to mechanical unfolding. Proc. Nat. Acad. Sci. USA 2012, 109, 16167–16172. [Google Scholar] [CrossRef]
- Qu, X.; Wen, J.; Lancaster, L.; Noller, H.; Bustamante, C.; Tinoco, I. The ribosome uses two active mechanisms to unwind messenger RNA during translation. Nature 2011, 475, 118–121. [Google Scholar] [CrossRef]
- Moomau, C.; Musalgaonkar, S.; Khan, Y.; Jones, J.; Dinman, J. Structural and functional characterization of programmed ribosomal frameshift signals in West Nile virus strains reveals high structural plasticity among cis-acting RNA elements. J. Biol. Chem. 2016, 291, 15788–15795. [Google Scholar] [CrossRef]
- Yang, L.; Toh, D.; Krishna, M.; Zhong, Z.; Liu, Y.; Wang, S.; Gong, Y.; Chen, G. Tertiary base triple formation in the SRV-1 frameshifting pseudoknot stabilizes secondary structure components. Biochemistry 2020, 59, 4429–4438. [Google Scholar] [CrossRef]
- Staple, D.; Butcher, S. Solution structure and thermodynamic investigation of the HIV-1 frameshift inducing element. J. Mol. Biol. 2005, 349, 1011–1023. [Google Scholar] [CrossRef]
- Marcheschi, R.; Staple, D.; Butcher, S. Programmed ribosomal frameshifting in SIV is induced by a highly structured RNA stem-loop. J. Mol. Biol. 2007, 373, 652–663. [Google Scholar] [CrossRef]
- Nixon, P.; Rangan, A.; Kim, Y.; Rich, A.; Hoffman, D.; Hennig, M.; Giedroc, D. Solution structure of a luteoviral P1-P2 frameshifting mRNA pseudoknot. J. Mol. Biol. 2002, 322, 621–633. [Google Scholar] [CrossRef]
- Pallan, P.; Marshall, W.; Harp, J.; Jewett, F.; Wawrzak, Z.; Brown, B.; Rich, R.; Egli, M. Crystal structure of a luteoviral RNA pseudoknot and model for a minimal ribosomal frameshifting motif. Biochemistry 2005, 44, 11315–11322. [Google Scholar] [CrossRef]
- Cornish, P.; Hennig, M.; Giedroc, D. A loop 2 cytidine-stem 1 minor groove interaction as a positive determinant for pseudoknot-stimulated -1 ribosomal frameshifting. Proc. Natl. Acad. Sci. USA 2005, 102, 12694–12699. [Google Scholar] [CrossRef]
- Egli, M.; Minasov, G.; Su, L.; Rich, A. Metal ions and flexibility in a viral RNA pseudoknot at atomic resolution. Proc. Natl. Acad. Sci. USA 2002, 99, 4302–4307. [Google Scholar] [CrossRef]
- Shen, L.; Tinoco, I. The structure of an RNA pseudoknot that causes efficient frameshifting in Mouse Mammary Tumor Virus. J. Mol. Biol. 1995, 247, 963–978. [Google Scholar] [CrossRef]
- Michiels, P.; Versleijen, A.; Verlaan, P.; Pleij, C.; Hilbers, C.; Heus, H. Solution structure of the pseudoknot of SRV-1 RNA, involved in ribosomal frameshifting. J. Mol. Biol. 2001, 310, 1109–1123. [Google Scholar] [CrossRef]
- Roman, C.; Lewicka, A.; Koirala, D.; Li, N.; Piccirilli, J. The SARS-CoV-2 programmed −1 ribosomal frameshifting element crystal structure solved to 2.09 Å using chaperone-assisted RNA crystallography. ACS Chem. Biol. 2021, 16, 1469–1481. [Google Scholar] [CrossRef]
- Dulude, D.; Baril, M.; Brakier-Gingras, L. Characterization of the frameshift stimulatory signal controlling a programmed −1 ribosomal frameshift in the human immunodeficiency virus type 1. Nucleic Acids Res. 2002, 30, 5094–5102. [Google Scholar] [CrossRef]
- Kim, Y.; Maas, L.; O’Neil, A.; Rich, A. Specific mutations in a viral RNA pseudoknot drastically change ribosomal frameshifting efficiency. Proc. Natl. Acad. Sci. USA 1999, 96, 14234–14239. [Google Scholar] [CrossRef]
- Chamorro, M.; Parkin, N.; Varmus, H. An RNA pseudoknot and an optimal heptameric shift site are required for highly efficient ribosomal frameshifting on a retroviral messenger RNA. Proc. Natl. Acad. Sci. USA 1992, 89, 713–717. [Google Scholar] [CrossRef]
- Sun, Y.; Abriola, L.; Niederer, R.; Pedersen, S.; Alfajaro, M.; Monteiro, V.; Wilen, C.; Ho, Y.; Gilbert, W.; Surovtseva, Y.; et al. Restriction of SARS-CoV-2 replication by targeting programmed −1 ribosomal frameshifting. Proc. Natl. Acad. Sci. USA 2021, 118, e2023051118. [Google Scholar] [CrossRef]
- Sharer, L.; Baskin, G.; Cho, E.; Murphey-Corb, M.; Blumberg, B.; Epstein, L. Comparison of simian immunodeficiency virus and human immunodeficiency virus encephalitides in the immature host. Ann. Neurol. 1988, 23, S108. [Google Scholar] [CrossRef]
- McCown, P.; Corbino, K.; Stav, S.; Sherlock, M.; Breaker, R. Riboswitch diversity and distribution. RNA 2017, 23, 995–1011. [Google Scholar] [CrossRef]
- Kalvari, I.; Nawrocki, E.; Ontiveros-Palacios, J.A.N.; Lamkiewicz, K.; Marz, M.; Griffiths-Jones, S.; Toffano-Nioche, C.; Gautheret, D.; Weinberg, Z.; Rivas, E.; et al. Rfam 14: Expanded coverage of metagenomic, viral and microRNA families. Nucleic Acids Res. 2021, 49, 192–200. [Google Scholar] [CrossRef]
- Mignone, F.; Grillo, G.; Licciulli, F.; Iacono, M.; Liuni, S.; Kersey, P.; Duarte, J.; Saccone, C.; Pesole, G. UTRdb and UTRsite: A collection of sequences and regulatory motifs of the untranslated regions of eukaryotic mRNAs. Nucleic Acids Res. 2005, 33, 141–146. [Google Scholar] [CrossRef]
- Stamm, S.; Riethoven, J.; Le, T.; Gopalakrishnan, C.; Kumanduri, V.; Tang, Y.; Barbosa-Morais, N.; Thanaraj, T. ASD: A bioinformatics resource on alternative splicing. Nucleic Acids Res. 2006, 34, 46–55. [Google Scholar] [CrossRef]
- Matys, V.; Fricke, E.; Geffers, R.; Gossling, E.; Haubrock, M.; Hehl, R.; Hornischer, K.; Karas, D.; Kel, A.; Kel-Margoulis, O. TRANSFAC: Transcriptional regulation, from patterns to profiles. Nucleic Acids Res. 2003, 31, 374–378. [Google Scholar] [CrossRef]
- Hendrix, D.; Brennerand, S.; Holbrook, S. RNA structural motifs: Building blocks of a modular biomolecule. Q. Rev. Biophys. 2006, 38, 221–243. [Google Scholar] [CrossRef]
- Jain, S.; Bayrak, C.; Petingi, L.; Schlick, T. Dual graph partitioning highlights a small group of pseudoknot-containing RNA submotifs. Genes 2018, 9, 371. [Google Scholar] [CrossRef]
- Fiedler, M. Algebraic connectivity of graphs. Czechoslovak Math. J. 1973, 23, 298–305. [Google Scholar] [CrossRef]
- Yassin, A.; Haque, M.; Datta, P.; Elmore, K.; Banavali, N.; Spremulli, L.; Agrawal, R. Insertion domain within mammalian mitochondrial translation initiation factor 2 serves the role of eubacterial initiation factor 1. Proc. Natl. Acad. Sci. USA 2011, 108, 3918–3923. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | RAG Development | Refs. |
|---|---|---|
| 2003 | Launch of RAG: planar tree and dual graphs | [12] |
| 2011 | RNA junction coaxial stacking prediction | [26] |
| 2014 | Tree graph partitioning using Fiedler vectors | [18] |
| 2014 | RAG 3D tree graph: sampling RNA 3D structures | [13] |
| 2015 | Laplacian spectrum based graph feature selection and clustering | [15] |
| 2015 | RAG-3D Database: searching for similar RNA fragments | [22] |
| 2017 | Fragment assembly (F-RAG): generating atomic models for tree graphs | [20] |
| 2017 | Dual graph partitioning algorithm | [19] |
| 2018 | Novel RNA motif design pipeline | [17] |
| 2019 | Extended dual graph library and RAG-3Dual database | [14] |
| 2020 | Tree graph inverse folding (RAG-IF) | [21] |
| 2021 | Fiedler vector based graph feature selection and scoring | [16] |
| 2021 | Dual graph inverse folding (Dual-RAG-IF) and | [23] |
| SARS-CoV-2 frameshifting element (FSE) conformational landscape | [24] | |
| 2022 | SARS-CoV-2 FSE dynamics and Coronavirus conformational landscape | [25,27] |
| Vertex | Graphs | Current Existing Graphs | Prior Existing | |||
|---|---|---|---|---|---|---|
| Rd 1 | Rd 2 | Combined | Pknot | Total/(Common) | ||
| 1 | 1 | 1 | 1 | 1 | 0 | 1 (1) |
| 2 | 3 | 3 | 3 | 3 | 1 | 3 (3) |
| 3 | 8 | 8 | 4 | 8 | 4 | 7 (7) |
| 4 | 29 | 22 | 5 | 22 | 13 | 17 (17) |
| 5 | 110 | 28 | 6 | 29 | 18 | 20 (17) |
| 6 | 508 | 36 | 5 | 39 | 21 | 22 (16) |
| 7 | 2551 | 31 | 4 | 33 | 18 | 21 (13) |
| 8 | 14,670 | 18 | 3 | 20 | 9 | 14 (5) |
| 9 | 92,788 | 25 | 4 | 28 | 16 | 17 (10) |
| Total | 110,668 | 172 | 35 | 183 | 100 | 122 (89) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Petingi, L.; Schlick, T. RNA-As-Graphs Motif Atlas—Dual Graph Library of RNA Modules and Viral Frameshifting-Element Applications. Int. J. Mol. Sci. 2022, 23, 9249. https://doi.org/10.3390/ijms23169249
Zhu Q, Petingi L, Schlick T. RNA-As-Graphs Motif Atlas—Dual Graph Library of RNA Modules and Viral Frameshifting-Element Applications. International Journal of Molecular Sciences. 2022; 23(16):9249. https://doi.org/10.3390/ijms23169249
Chicago/Turabian StyleZhu, Qiyao, Louis Petingi, and Tamar Schlick. 2022. "RNA-As-Graphs Motif Atlas—Dual Graph Library of RNA Modules and Viral Frameshifting-Element Applications" International Journal of Molecular Sciences 23, no. 16: 9249. https://doi.org/10.3390/ijms23169249
APA StyleZhu, Q., Petingi, L., & Schlick, T. (2022). RNA-As-Graphs Motif Atlas—Dual Graph Library of RNA Modules and Viral Frameshifting-Element Applications. International Journal of Molecular Sciences, 23(16), 9249. https://doi.org/10.3390/ijms23169249