
Biological Variation in Peripheral Inflammation and Oxidative Stress Biomarkers in Individuals with Gaucher Disease

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Participant Demographics
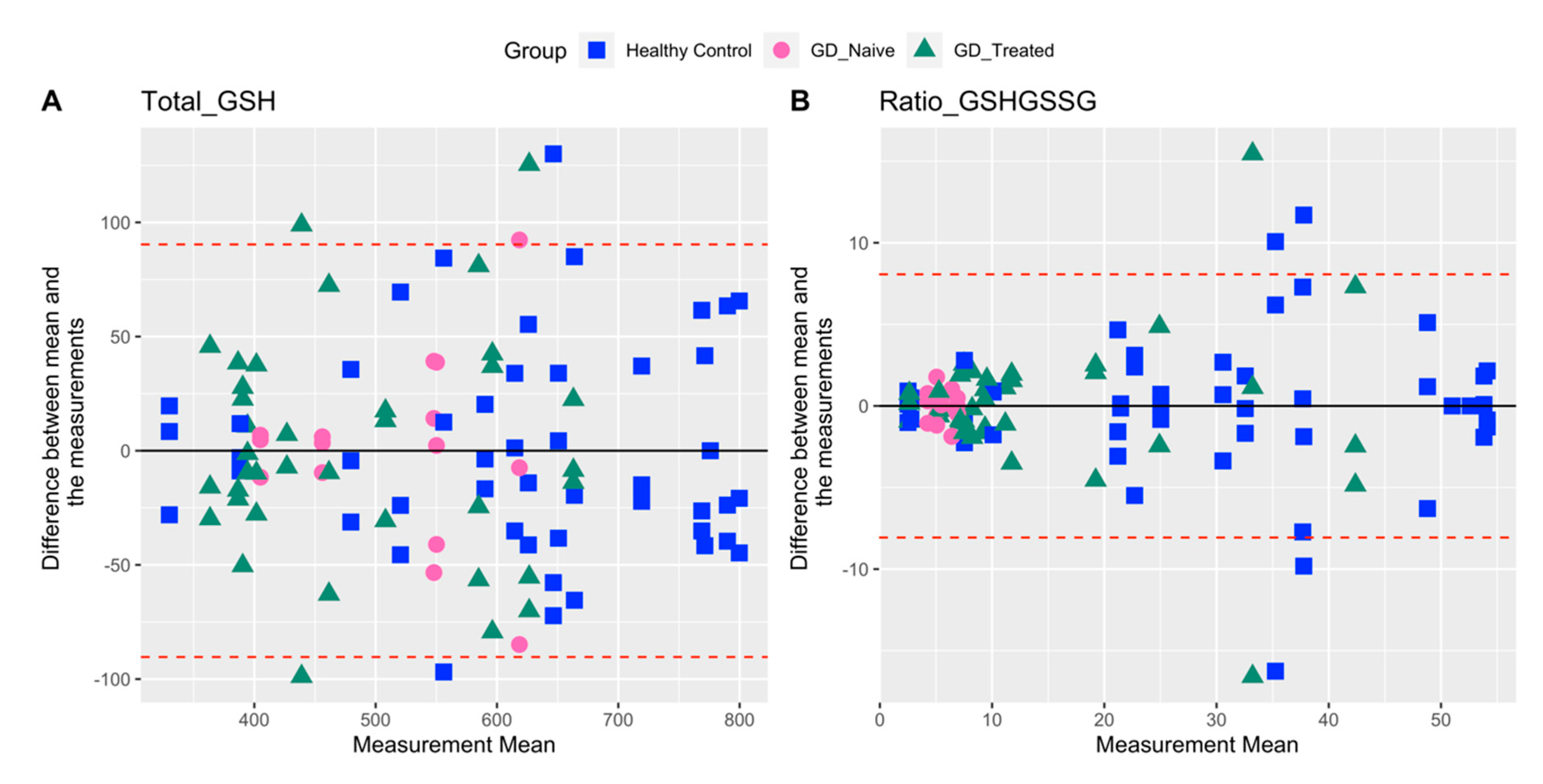
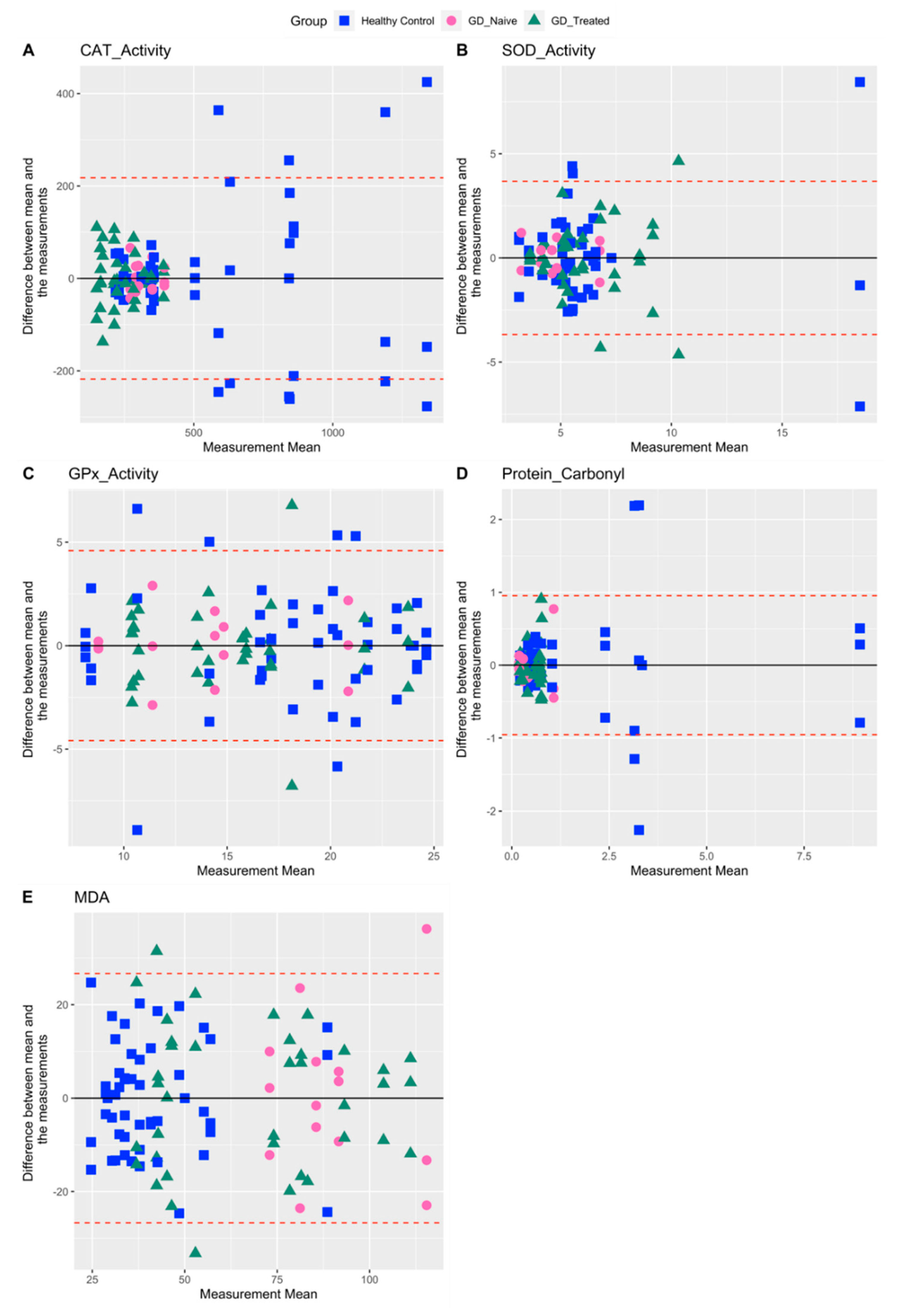
2.2. Variability in Measures Related to Oxidative Stress
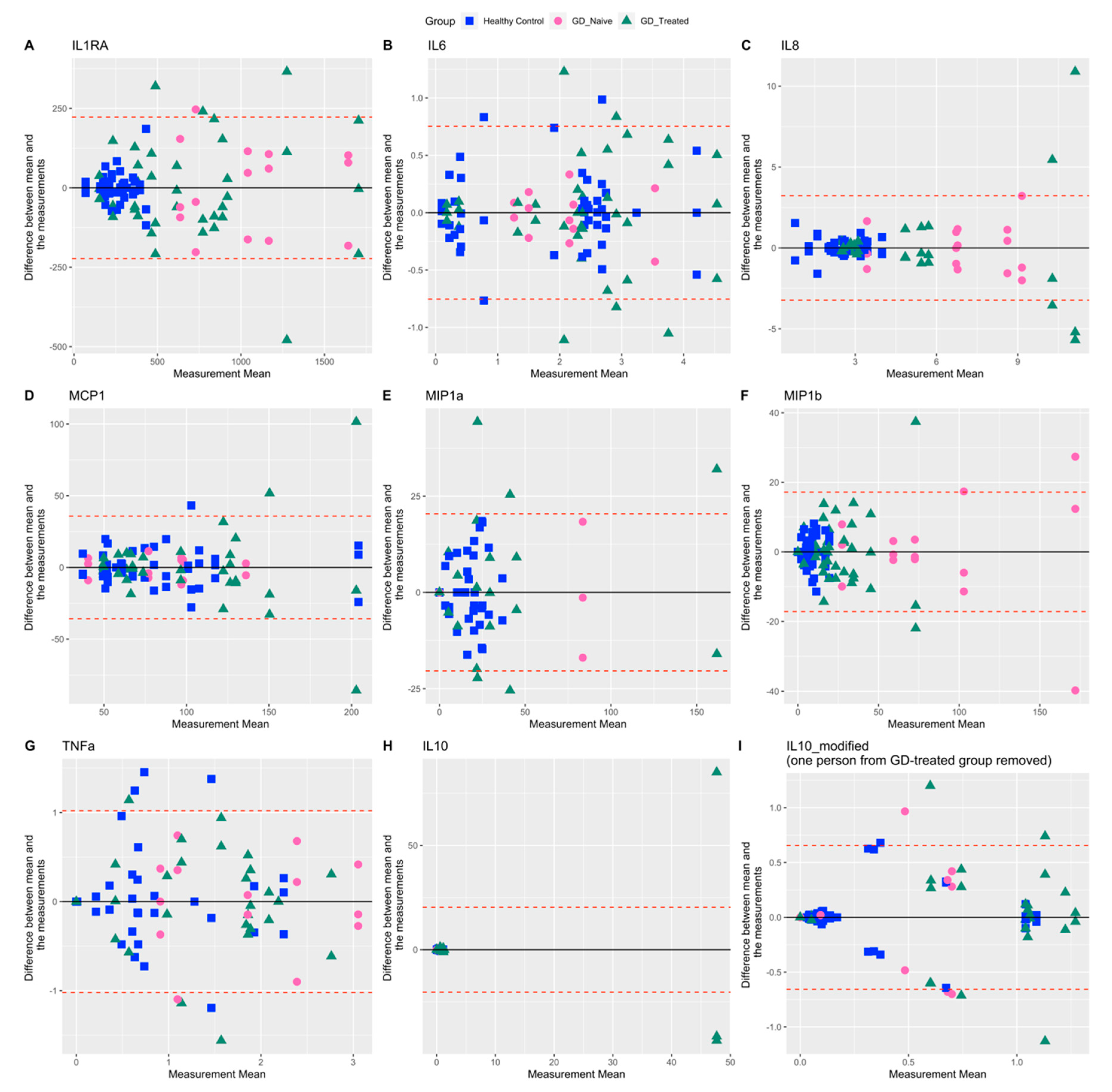
2.3. Variability in Measures Related to Inflammation
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Blood Sample Collection
- (1)
- Intracellular glutathione (GSH) status measured as total GSH and the redox ratio of reduced/oxidized glutathione (GSH/GSSG) in RBCs;
- (2)
- The activity of intracellular antioxidant enzymes-catalase (CAT), superoxide dismutase (SOD), and Glutathione peroxidase (GPx) in RBCs;
- (3)
- Plasma lipid peroxidation profile as determined by malondialdehyde (MDA) levels;
- (4)
- Oxidative modification of proteins determined as protein carbonylation (Protein carbonyl) levels in plasma.
- (1)
- Pro-inflammation cytokines: interleukin-1 receptor antagonist (IL-1RA), interleukins (IL-6, IL-8, IL-10), tumor necrosis factor-alpha (TNFa), monocyte chemoattractant protein-1 (MCP-1) measured in plasma;
- (2)
- Inflammation markers related to skeletal manifestations: macrophage inflammatory protein (MIP)-1alpha (MIP-1a) and MIP-1beta (MIP-1b), measured in plasma.
4.3. Measurement of Oxidative Stress
4.4. Measurement of Inflammation
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meikle, P.J. Prevalence of Lysosomal Storage Disorders. JAMA 1999, 281, 249. [Google Scholar] [CrossRef]
- Brady, R.O.; Kanfer, J.N.; Bradley, R.M.; Shapiro, D. Demonstration of a deficiency of glucocerebroside-cleaving enzyme in Gaucher’s disease. J. Clin. Investig. 1966, 45, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Avenali, M.; Blandini, F.; Cerri, S. Glucocerebrosidase Defects as a Major Risk Factor for Parkinson’s Disease. Front. Aging Neurosci. 2020, 12, 97. [Google Scholar] [CrossRef]
- Maor, G.; Rencus-Lazar, S.; Filocamo, M.; Steller, H.; Segal, D.; Horowitz, M. Unfolded protein response in Gaucher disease: From human to Drosophila. Orphanet J. Rare Dis. 2013, 8, 140. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E. The pathology of Gaucher disease. Prog. Clin. Biol. Res. 1982, 95, 177–217. [Google Scholar] [PubMed]
- Gegg, M.E.; Schapira, A.H.V. Mitochondrial dysfunction associated with glucocerebrosidase deficiency. Neurobiol. Dis. 2016, 90, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.E.G.; Liebscher, G.; Hess, M.W.; Huber, L.A. Lysosomal size matters. Traffic 2020, 21, 60–75. [Google Scholar] [CrossRef]
- Tantawy, A.A.G. Cytokines in Gaucher disease: Role in the pathogenesis of bone and pulmonary disease. Egypt. J. Med. Hum. Genet. 2015, 16, 207–213. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals, antioxidants, and human disease: Curiosity, cause, or consequence? Lancet 1994, 344, 721–724. [Google Scholar] [CrossRef]
- Roversi, F.M.; Galdieri, L.C.; Grego, B.H.C.; Souza, F.G.; Micheletti, C.; Martins, A.M.; D’Almeida, V. Blood oxidative stress markers in Gaucher disease patients. Clin. Chim. Acta 2006, 364, 316–320. [Google Scholar] [CrossRef]
- Deganuto, M.; Pittis, M.G.; Pines, A.; Dominissini, S.; Kelley, M.R.; Garcia, R.; Quadrifoglio, F.; Bembi, B.; Tell, G. Altered intracellular redox status in Gaucher disease fibroblasts and impairment of adaptive response against oxidative stress. J. Cell. Physiol. 2007, 212, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Cleeter, M.W.J.; Chau, K.-Y.; Gluck, C.; Mehta, A.; Hughes, D.A.; Duchen, M.; Wood, N.W.; Hardy, J.; Mark Cooper, J.; Schapira, A.H. Glucocerebrosidase inhibition causes mitochondrial dysfunction and free radical damage. Neurochem. Int. 2013, 62, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Belmatoug, N.; Di Rocco, M.; Fraga, C.; Giraldo, P.; Hughes, D.; Lukina, E.; Maison-Blanche, P.; Merkel, M.; Niederau, C.; Plöckinger, U.; et al. Management and monitoring recommendations for the use of eliglustat in adults with type 1 Gaucher disease in Europe. Eur. J. Intern. Med. 2017, 37, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Balwani, M.; Burrow, T.A.; Charrow, J.; Goker-Alpan, O.; Kaplan, P.; Kishnani, P.S.; Mistry, P.; Ruskin, J.; Weinreb, N. Recommendations for the use of eliglustat in the treatment of adults with Gaucher disease type 1 in the United States. Mol. Genet. Metab. 2016, 117, 95–103. [Google Scholar] [CrossRef]
- Hayes, R.P.; Grinzaid, K.A.; Duffey, E.B.; Elsas II, L.J. The impact of Gaucher disease and its treatment on quality of life. Qual. Life Res. 1998, 7, 521–534. [Google Scholar] [CrossRef]
- Chen Zion, Y.; Pappadopulos, E.; Wajnrajch, M.; Rosenbaum, H. Rethinking fatigue in Gaucher disease. Orphanet J. Rare Dis. 2016, 11, 53. [Google Scholar] [CrossRef]
- Kartha, R.V.; Terluk, M.R.; Brown, R.; Travis, A.; Mishra, U.R.; Rudser, K.; Lau, H.; Jarnes, J.R.; Cloyd, J.C.; Weinreb, N.J. Patients with Gaucher disease display systemic oxidative stress dependent on therapy status. Mol. Genet. Metab. Rep. 2020, 25, 100667. [Google Scholar] [CrossRef]
- Shemesh, E.; Deroma, L.; Bembi, B.; Deegan, P.; Hollak, C.; Weinreb, N.J.; Cox, T.M. Enzyme replacement and substrate reduction therapy for Gaucher disease. In Cochrane Database of Systematic Reviews; Shemesh, E., Ed.; John Wiley & Sons, Ltd: Chichester, UK, 2013. [Google Scholar]
- FDA Center for Drug Evaluation and Research, Medical Review [CERDELGA]. 2014. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2014/205494Orig1s000MedR.pdf (accessed on 14 August 2022).
- Mistry, P.K.; Lukina, E.; Ben Turkia, H.; Amato, D.; Baris, H.; Dasouki, M.; Ghosn, M.; Mehta, A.; Packman, S.; Pastores, G.; et al. Effect of Oral Eliglustat vs Placebo on Spleen Volume in Patients with Splenomegaly and Gaucher Disease Type 1: The ENGAGE Randomized Clinical Trial. JAMA 2015, 313, 695. [Google Scholar] [CrossRef]
- Mistry, P.K.; Lukina, E.; Ben Turkia, H.; Shankar, S.P.; Feldman, H.; Ghosn, M.; Mehta, A.; Packman, S.; Lau, H.; Petakov, M.; et al. Clinical outcomes after 4.5 years of eliglustat therapy for Gaucher disease type 1: Phase 3 ENGAGE trial final results. Am. J. Hematol. 2021, 96, 1156–1165. [Google Scholar] [CrossRef]
- Pastores, G.M.; Weinreb, N.J.; Aerts, H.; Andria, G.; Cox, T.M.; Giralt, M.; Grabowski, G.A.; Mistry, P.K.; Tylki-Szymańska, A. Therapeutic goals in the treatment of Gaucher disease. Semin. Hematol. 2004, 41, 4–14. [Google Scholar] [CrossRef]
- Barak, V.; Acker, M.; Nisman, B.; Kalickman, I.; Abrahamov, A.; Zimran, A.; Yatziv, S. Cytokines in Gaucher’s disease. Eur. Cytokine Netw. 1999, 10, 205–210. [Google Scholar] [PubMed]
- Seattle Children’s Hospital Gaucher Disease Biomarker Panel. Available online: https://seattlechildrenslab.testcatalog.org/show/LAB3073-1 (accessed on 9 August 2022).
- Gervas-Arruga, J.; Cebolla, J.J.; de Blas, I.; Roca, M.; Pocovi, M.; Giraldo, P. The Influence of Genetic Variability and Proinflammatory Status on the Development of Bone Disease in Patients with Gaucher Disease. PLoS ONE 2015, 10, e0126153. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Branagan, A.R.; Liu, J.; Boddupalli, C.S.; Mistry, P.K.; Dhodapkar, M.V. Clonal Immunoglobulin against Lysolipids in the Origin of Myeloma. N. Engl. J. Med. 2016, 374, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Grabowski, G.A. Immunological Cells and Functions in Gaucher Disease. Crit. Rev. Oncog. 2013, 18, 197–220. [Google Scholar] [CrossRef]
- Jmoudiak, M.; Futerman, A.H. Gaucher disease: Pathological mechanisms and modern management. Br. J. Haematol. 2005, 129, 178–188. [Google Scholar] [CrossRef]
- De Fost, M.; Out, T.A.; de Wilde, F.A.; Tjin, E.P.M.; Pals, S.T.; van Oers, M.H.J.; Boot, R.G.; Aerts, J.F.M.G.; Maas, M.; vom Dahl, S.; et al. Immunoglobulin and free light chain abnormalities in Gaucher disease type I: Data from an adult cohort of 63 patients and review of the literature. Ann. Hematol. 2008, 87, 439–449. [Google Scholar] [CrossRef]
- Allen, M.J.; Myer, B.J.; Khokher, A.M.; Rushton, N.; Cox, T.M. Pro-inflammatory cytokines and the pathogenesis of Gaucher’s disease: Increased release of interleukin-6 and interleukin-10. QJM 1997, 90, 19–25. [Google Scholar] [CrossRef]
- Hollak, C.E.M.; Evers, L.; Aerts, J.M.F.G.; van Oers, M.H.J. Elevated Levels of M-CSF, sCD14 and IL8 in Type 1 Gaucher Disease. Blood Cells Mol. Dis. 1997, 23, 201–212. [Google Scholar] [CrossRef]
- Van Breemen, M.J.; de Fost, M.; Voerman, J.S.A.; Laman, J.D.; Boot, R.G.; Maas, M.; Hollak, C.E.M.; Aerts, J.M.; Rezaee, F. Increased plasma macrophage inflammatory protein (MIP)-1α and MIP-1β levels in type 1 Gaucher disease. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 788–796. [Google Scholar] [CrossRef]
- Mello, A.S.; da Silva Garcia, C.; de Souza Machado, F.; da Silva Medeiros, N.; Wohlenberg, M.F.; Marinho, J.P.; Dani, C.; Funchal, C.; Coelho, J.C. Oxidative stress parameters of Gaucher disease type I patients. Mol. Genet. Metab. Rep. 2015, 4, 1–5. [Google Scholar] [CrossRef]
- Zahran, A.M.; Elsayh, K.I.; El-Deek, S.E.M.; El-Baz, M.A.H. Oxidative Stress, Trace Elements, and Circulating Microparticles in Patients with Gaucher Disease Before and After Enzyme Replacement Therapy. Clin. Appl. Thromb. 2015, 21, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, H.; Khatami, S.; Kiani, A.; Rahimi, Z.; Vaisi-Raygani, A.; Afsharnaderi, A.; Alaei, M.R. Oxidative Stress Parameters, Trace Elements, and Lipid Profile in Iranian Patients with Gaucher Disease. Biol. Trace Elem. Res. 2020, 193, 130–137. [Google Scholar] [CrossRef] [PubMed]
- SRT in Comparison to ERT on Immune Aspects and Bone Involvement in Gaucher Disease [NCT02605603]. Available online: https://www.clinicaltrials.gov/ct2/show/NCT02605603?cond=Gaucher+Disease&draw=2 (accessed on 4 July 2022).
- Immune Biomarkers Related to Bone Pathology in Patients with Type 1 Gaucher Disease [NCT04055831]. Available online: https://www.clinicaltrials.gov/ct2/show/NCT04055831?cond=Gaucher+Disease&draw=2&rank=2 (accessed on 4 July 2022).
- Trials and Research: Gaucher Online Disease Platform. Available online: https://gaucherdiseaseplatform.org/trials-and-research/ (accessed on 4 July 2022).
- Oxidative Stress and Inflammatory Biomarkers in Gaucher Disease [NCT02437396]. Available online: https://clinicaltrials.gov/ct2/show/NCT02437396 (accessed on 4 July 2022).
- Altman, D.G.; Bland, J.M. Measurement in Medicine: The Analysis of Method Comparison Studies. Stat. 1983, 32, 307. [Google Scholar] [CrossRef]
- Martin Bland, J.; Altman, D. Statistical Methods for Assessing Agreement Between Two Methods of Clinical Measurement. Lancet 1986, 327, 307–310. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. Statistics notes: Calculating correlation coefficients with repeated observations: Part 1-correlation within subjects. BMJ 1995, 310, 446. [Google Scholar] [CrossRef]
- Schirmer, M.; Kumar, V.; Netea, M.G.; Xavier, R.J. The causes and consequences of variation in human cytokine production in health. Curr. Opin. Immunol. 2018, 54, 50–58. [Google Scholar] [CrossRef]
- Dabrosin, C.; Öllinger, K.; Ungerstedt, U.; Hammar, M. Variability of Glutathione Levels in Normal Breast Tissue and Subcutaneous Fat during the Menstrual Cycle: An in Vivo Study with Microdialysis Technique. J. Clin. Endocrinol. Metab. 1997, 82, 1382–1384. [Google Scholar] [CrossRef][Green Version]
- Blanco, R.A.; Ziegler, T.R.; Carlson, B.A.; Cheng, P.-Y.; Park, Y.; Cotsonis, G.A.; Accardi, C.J.; Jones, D.P. Diurnal variation in glutathione and cysteine redox states in human plasma. Am. J. Clin. Nutr. 2007, 86, 1016–1023. [Google Scholar] [CrossRef]
- Woods, J.A.; Vieira, V.J.; Keylock, K.T. Exercise, Inflammation, and Innate Immunity. Neurol. Clin. 2006, 24, 585–599. [Google Scholar] [CrossRef]
- Suzuki, K.; Nakaji, S.; Yamada, M.; Totsuka, M.; Sato, K.; Sugawara, K. Systemic inflammatory response to exhaustive exercise. Cytokine kinetics. Exerc. Immunol. Rev. 2002, 8, 6–48. [Google Scholar]
- Elokda, A.S.; Nielsen, D.H. Effects of exercise training on the glutathione antioxidant system. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 630–637. [Google Scholar] [CrossRef] [PubMed]
- De Jager, W.; Bourcier, K.; Rijkers, G.T.; Prakken, B.J.; Seyfert-Margolis, V. Prerequisites for cytokine measurements in clinical trials with multiplex immunoassays. BMC Immunol. 2009, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Petrovsky, N.; McNair, P.; Harrison, L.C. Diurnal Rhythms of Pro-Inflammatory Cytokines: Regulation by Plasma Cortisol and Therapeutic Implications. Cytokine 1998, 10, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Petrovsky, N.; Harrison, L.C. The Chronobiology of Human Cytokine Production. Int. Rev. Immunol. 1998, 16, 635–649. [Google Scholar] [CrossRef]
- Smaaland, R.; Sothern, R.B.; Laerum, O.D.; Abrahamsen, J.F. Rhythms in human bone marrow and blood cells. Chronobiol. Int. 2002, 19, 101–127. [Google Scholar] [CrossRef]
- Chiolero, A.; Paradis, G.; Rich, B.; Hanley, J.A. Assessing the Relationship between the Baseline Value of a Continuous Variable and Subsequent Change Over Time. Front. Public Health 2013, 1, 29. [Google Scholar] [CrossRef]
- Food and Drug Administration. Analytical Procedures and Methods Validation for Drugs and Biologics Guidance for Industry. 2015. Available online: https://www.fda.gov/media/87801/download (accessed on 4 July 2022).
- Knight, V.; Long, T.; Meng, Q.H.; Linden, M.A.; Rhoads, D.D. Variability in the Laboratory Measurement of Cytokines. Arch. Pathol. Lab. Med. 2020, 144, 1230–1233. [Google Scholar] [CrossRef]
- Rudež, G.; Meijer, P.; Spronk, H.M.H.; Leebeek, F.W.G.; Ten Cate, H.; Kluft, C.; de Maat, M.P.M. Biological variation in inflammatory and hemostatic markers. J. Thromb. Haemost. 2009, 7, 1247–1255. [Google Scholar] [CrossRef]
- Meijers, W.C.; van der Velde, A.R.; Muller Kobold, A.C.; Dijck-Brouwer, J.; Wu, A.H.; Jaffe, A.; de Boer, R.A. Variability of biomarkers in patients with chronic heart failure and healthy controls. Eur. J. Heart Fail. 2017, 19, 357–365. [Google Scholar] [CrossRef]
- Mallard, A.R.; Hollekim-Strand, S.M.; Ingul, C.B.; Coombes, J.S. High day-to-day and diurnal variability of oxidative stress and inflammation biomarkers in people with type 2 diabetes mellitus and healthy individuals. Redox Rep. 2020, 25, 64–69. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Healthy Control (N = 18) | Gaucher Disease Type 1 (GD1) | |
|---|---|---|---|
| GD-Naïve (N = 5) | GD-Treated (N = 13) | ||
| Female | 9 (50%) | 3 (60.0%) | 8 (61.5%) |
| Caucasian | 12 (75.0%) | 5 (100%) | 12 (92.3%) |
| Age (in years) | 40.8 (15.3) | 60.0 (9.3) | 46.9 (12.0) |
| GD1 biomarkers | |||
| CHITO (nmoles/h/mL) | <78.5 * | 3590 (4182.5) 1 | 220 (224.2) 7 |
| ACE (IU/L) | 32.8–107.9 * | 95.6 (86.7) 2 | 62.8 (35.2) 5 |
| TRAP (IU/L) | 0.28–9.84 * | 25.2 (11.7) 3 | 7.18 (2.6) 8 |
| Complete blood count | |||
| Hemoglobin (g/L) | ND | 13.6 (1.7) | 14.4 (1.7) 2 |
| Hematocrit (%) | 39.4 (5.6) 1 | 42.3 (4.9) 2 | |
| Neutrophil (%) | 72.5 (0.7) 3 | 55.2 (7.4) 6 | |
| Lymphocyte (%) | 23.3 (9.2) 2 | 34.0 (6.8) 5 | |
| Eosinophils (%) | 1.5 (0.7) 3 | 2.83 (3.4) 6 | |
| WBC (×109/L) | 5.62 (1.0) | 6.02 (2.5) 3 | |
| Platelets (×109/L) | 122 (28.1) | 189 (65.5) 3 | |
| Mutational status | |||
| N370S/N370S | NA | 4 (80%) | 5 (38.5%) |
| N370S/L444P | 5 (38.5%) | ||
| N370S/unknown | 1 (7.7%) | ||
| N370S/R463C | 1 (7.7%) | ||
| Unknown | 1 (20%) | 1 (7.7%) | |
| GD1 therapy | |||
| Years on therapy | NA | NA | 16.1 (8.3) |
| ERT | NA | 6 (46.1%) | |
| SRT | NA | 7 (53.8%) | |
| Biomarker | %CV Overall | %CV Healthy Control | %CV GD-Naïve | %CV GD-Treated |
|---|---|---|---|---|
| Total GSH | 25.5 | 23.2 | 17.2 | 23.6 |
| Ratio GSH/GSSG | 82.6 | 58.6 | 16.7 | 86.7 |
| CAT Activity | 74.0 | 65.9 | 17.4 | 38.4 |
| SOD Activity | 55.7 | 66.4 | 29.7 | 42.7 |
| GPx Activity | 33.3 | 32.2 | 31.9 | 30.9 |
| Protein Carbonyl | 162.3 | 143.9 | 104.7 | 63.6 |
| MDA | 51.1 | 46.3 | 25.6 | 42.6 |
| IL1RA | 81.5 | 41.6 | 38.0 | 66.0 |
| IL-6 | 60.4 | 69.8 | 39.3 | 56.5 |
| IL-8 | 76.3 | 38.1 | 36.5 | 82.2 |
| MCP1 | 51.0 | 50.6 | 36.7 | 54.1 |
| MIP-1a | 164.7 | 81.3 | 205.9 | 173.1 |
| MIP-1b | 138.1 | 87.5 | 60.9 | 84.0 |
| TNFa | 85.9 | 113.7 | 52.7 | 67.1 |
| IL-10 | 701.6 * | 129.4 | 138.5 | 484.4 * |
| Biomarker | Healthy Control | GD-Naïve | GD-Treated |
|---|---|---|---|
| N = 18 | N = 5 | N = 13 | |
| Total_GSH (µg/mL) | |||
| Mean (SD) | 620 (144) | 516 (89) | 483 (114) |
| Median (IQR) | 614 (189) | 509 (122) | 439 (163) |
| Unknown | 6 | 0 | 2 |
| Ratio_GSHGSSG | |||
| Mean (SD) | 29 (17) | 6 (1) | 15 (13) |
| Median (IQR) | 27 (28) | 6 (2) | 10 (15) |
| Unknown | 5 | 0 | 2 |
| CAT_Activity (nmol/min/mL/mg) | |||
| Mean (SD) | 563 (371) | 321 (56) | 245 (94) |
| Median (IQR) | 388 (534) | 326 (71) | 244 (126) |
| Unknown | 4 | 0 | 2 |
| SOD_Activity (U/mL/mg) | |||
| Mean (SD) | 6.01 (3.99) | 4.71 (1.40) | 6.18 (2.64) |
| Median (IQR) | 4.95 (2.69) | 4.43 (1.36) | 5.68 (4.11) |
| Unknown | 4 | 0 | 2 |
| GPx_Activity (nmol/min/mL/mg) | |||
| Mean (SD) | 18.0 (5.8) | 14.1 (4.5) | 15.2 (4.7) |
| Median (IQR) | 19.2 (7.8) | 14.4 (5.7) | 15.0 (5.4) |
| Unknown | 4 | 0 | 2 |
| Protein_Carbonyl (nmol/mg) | |||
| Mean (SD) | 1.55 (2.23) | 0.43 (0.45) | 0.55 (0.35) |
| Median (IQR) | 0.64 (0.80) | 0.27 (0.33) | 0.54 (0.39) |
| Unknown | 4 | 0 | 1 |
| MDA (nM/mg) | |||
| Mean (SD) | 41 (19) | 90 (23) | 68 (29) |
| Median (IQR) | 38 (23) | 88 (17) | 65 (45) |
| Unknown | 4 | 1 | 1 |
| IL1RA (pg/mL) | |||
| Mean (SD) | 255 (106) | 1043 (396) | 694 (458) |
| Median (IQR) | 256 (155) | 1000 (513) | 619 (540) |
| Unknown | 5 | 0 | 1 |
| IL-6 (pg/mL) | |||
| Mean (SD) | 1.82 (1.27) | 2.14 (0.84) | 2.30 (1.30) |
| Median (IQR) | 2.29 (2.25) | 2.08 (0.95) | 2.26 (1.53) |
| Unknown | 5 | 0 | 1 |
| IL-8 (pg/mL) | |||
| Mean (SD) | 2.52 (0.96) | 6.94 (2.53) | 4.65 (3.82) |
| Median (IQR) | 2.56 (0.90) | 7.05 (2.35) | 3.13 (2.52) |
| Unknown | 5 | 0 | 1 |
| MCP1 (pg/mL) | |||
| Mean (SD) | 81 (41) | 90 (33) | 98 (53) |
| Median (IQR) | 68 (43) | 89 (32) | 78 (62) |
| Minimum–Maximum | 32–219 | 31–139 | 45–304 |
| Unknown | 5 | 0 | 1 |
| MIP-1a (pg/mL) | |||
| Mean (SD) | 16 (13) | 17 (35) | 26 (45) |
| Median (IQR) | 15 (22) | 0 (0) | 1 (36) |
| Minimum–Maximum | 0–43 | 0–102 | 0–194 |
| Unknown | 5 | 0 | 1 |
| MIP-1b (pg/mL) | |||
| Mean (SD) | 8 (7) | 87 (53) | 25 (21) |
| Median (IQR) | 7 (11) | 71 (51) | 22 (22) |
| Unknown | 5 | 0 | 1 |
| TNFa (pg/mL) | |||
| Mean (SD) | 0.73 (0.83) | 1.86 (0.98) | 1.40 (0.94) |
| Median (IQR) | 0.27 (1.27) | 1.84 (1.33) | 1.58 (1.35) |
| Unknown | 5 | 0 | 1 |
| IL-10 (U/mL) | |||
| Mean (SD) | 0.34 (0.44) | 0.39 (0.54) | 4.42 (21.41) * |
| Median (IQR) | 0.08 (0.91) | 0.08 (1.00) | 1.02 (1.17) * |
| Unknown | 5 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahasrabudhe, S.A.; Terluk, M.R.; Rudser, K.D.; Cloyd, J.C.; Kartha, R.V. Biological Variation in Peripheral Inflammation and Oxidative Stress Biomarkers in Individuals with Gaucher Disease. Int. J. Mol. Sci. 2022, 23, 9189. https://doi.org/10.3390/ijms23169189
Sahasrabudhe SA, Terluk MR, Rudser KD, Cloyd JC, Kartha RV. Biological Variation in Peripheral Inflammation and Oxidative Stress Biomarkers in Individuals with Gaucher Disease. International Journal of Molecular Sciences. 2022; 23(16):9189. https://doi.org/10.3390/ijms23169189
Chicago/Turabian StyleSahasrabudhe, Siddhee A., Marcia R. Terluk, Kyle D. Rudser, James C. Cloyd, and Reena V. Kartha. 2022. "Biological Variation in Peripheral Inflammation and Oxidative Stress Biomarkers in Individuals with Gaucher Disease" International Journal of Molecular Sciences 23, no. 16: 9189. https://doi.org/10.3390/ijms23169189
APA StyleSahasrabudhe, S. A., Terluk, M. R., Rudser, K. D., Cloyd, J. C., & Kartha, R. V. (2022). Biological Variation in Peripheral Inflammation and Oxidative Stress Biomarkers in Individuals with Gaucher Disease. International Journal of Molecular Sciences, 23(16), 9189. https://doi.org/10.3390/ijms23169189