
Aberrant Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem Cells of a Retinitis Pigmentosa Patient with the PRPF6 Mutation

 , ,
, ,
Abstract
:1. Introduction
2. Results
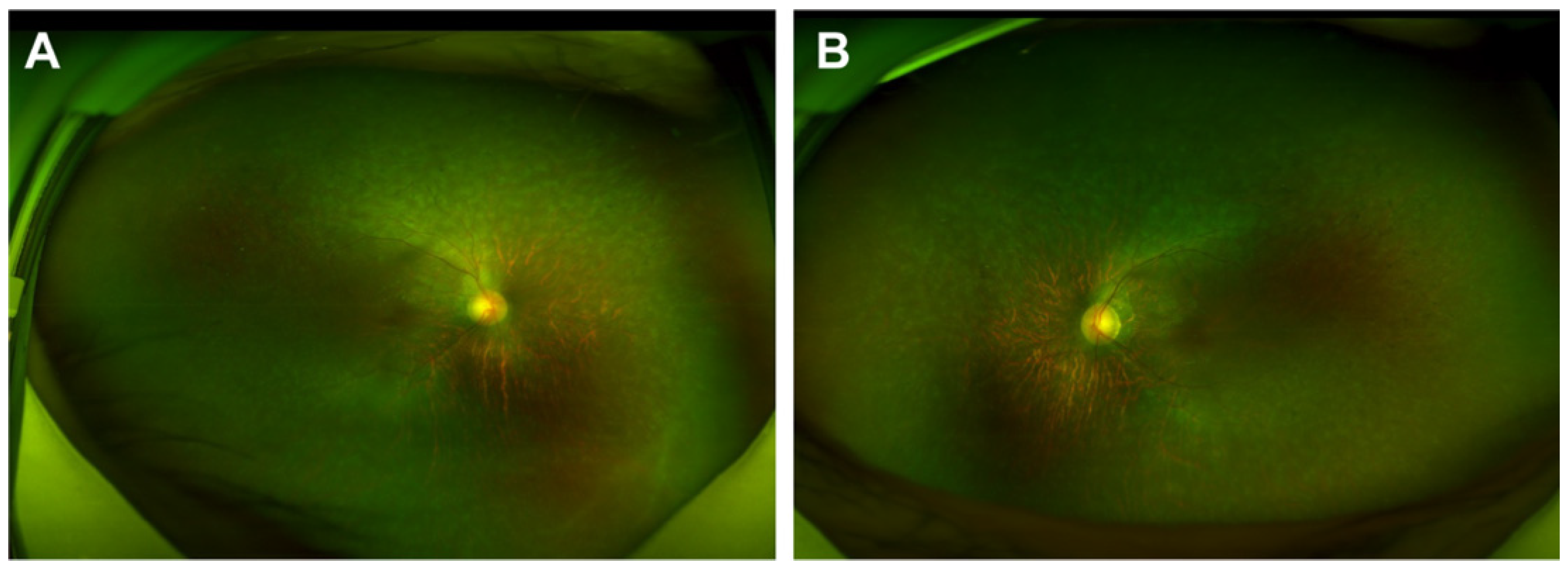
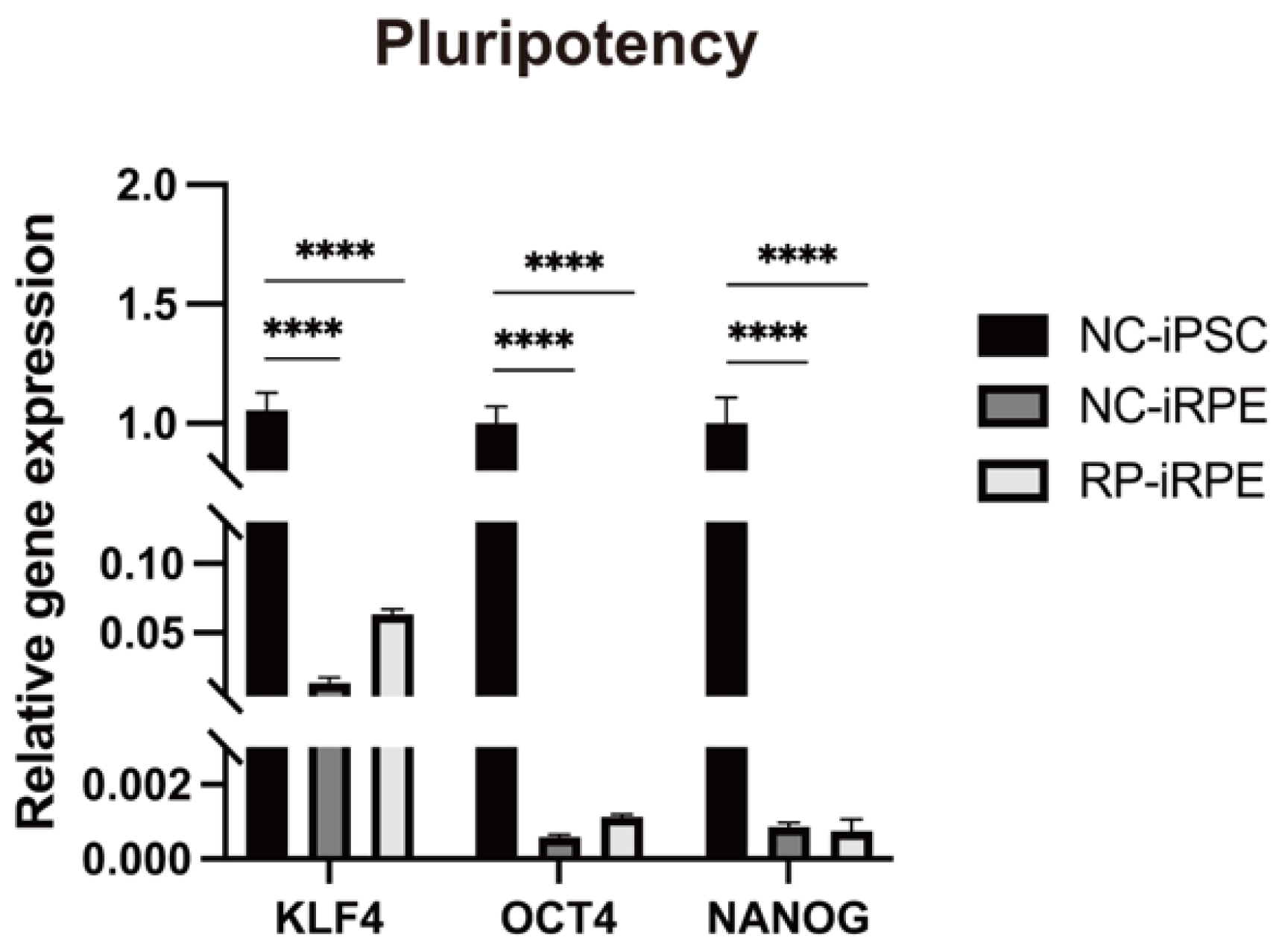
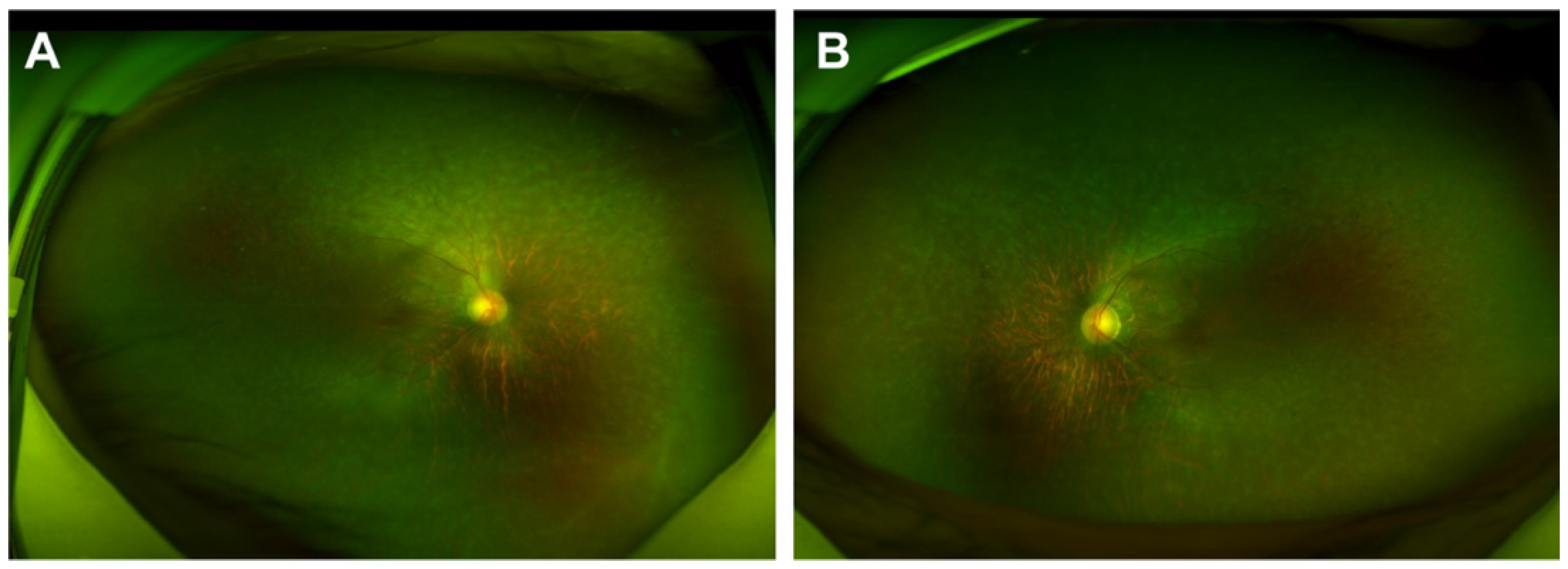
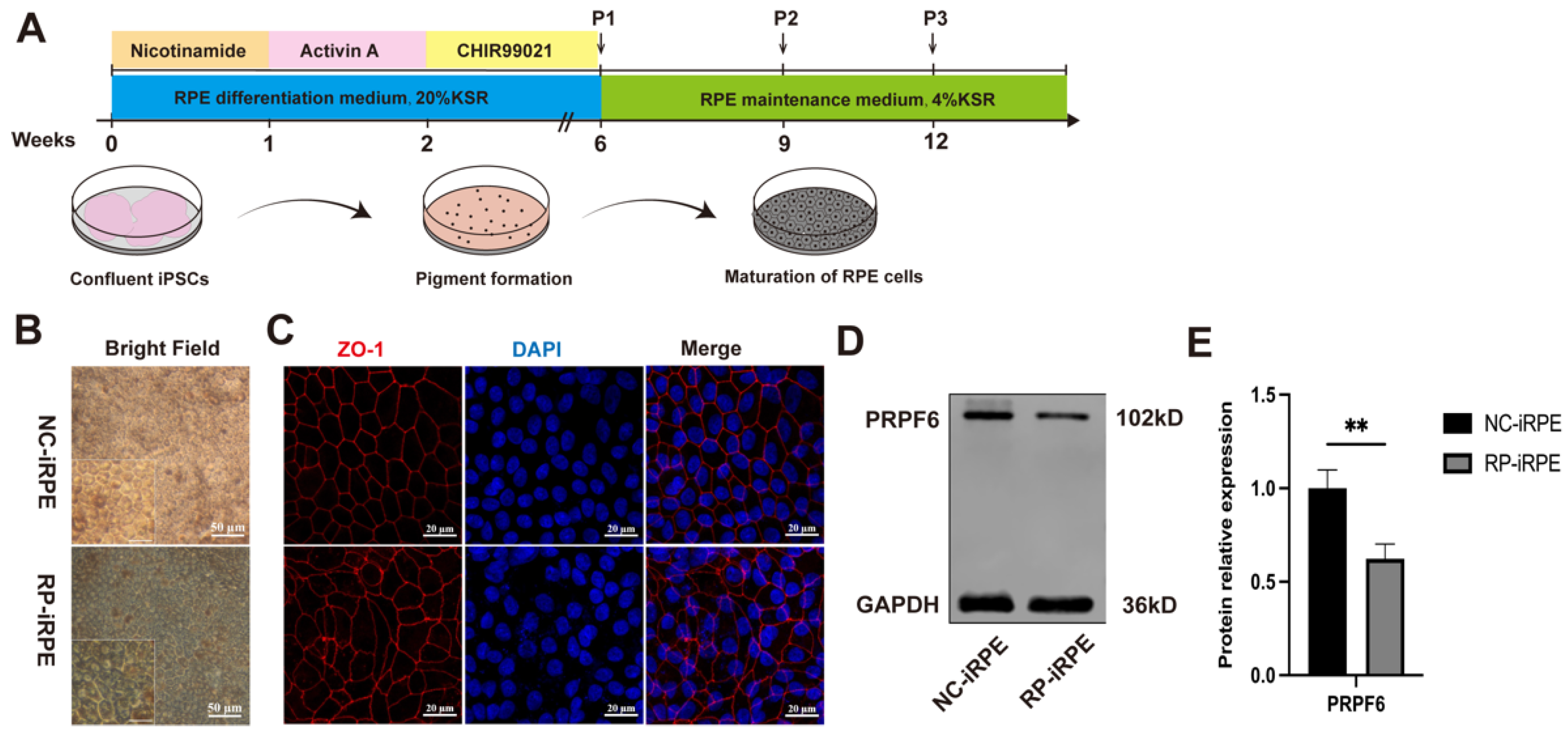
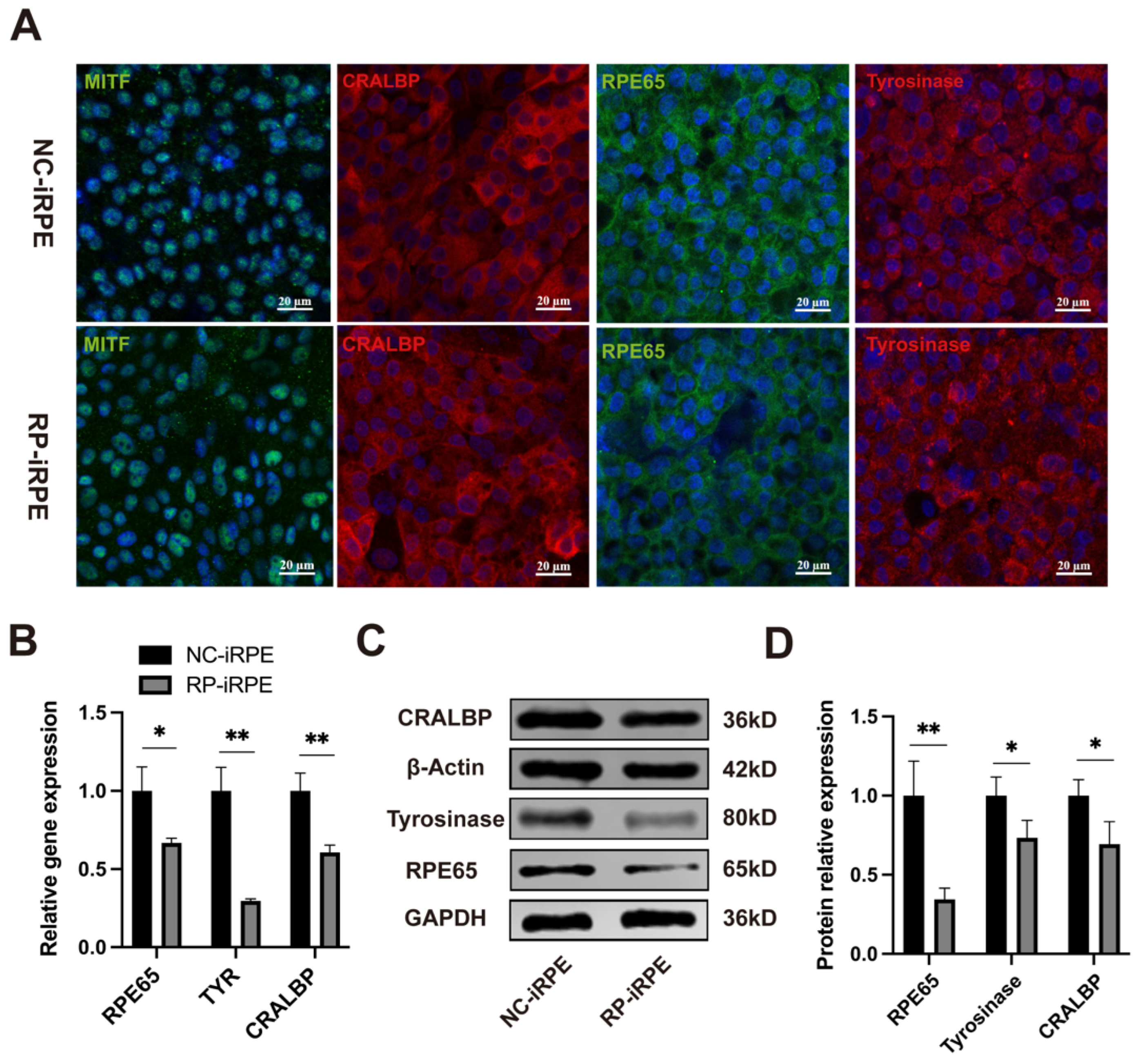
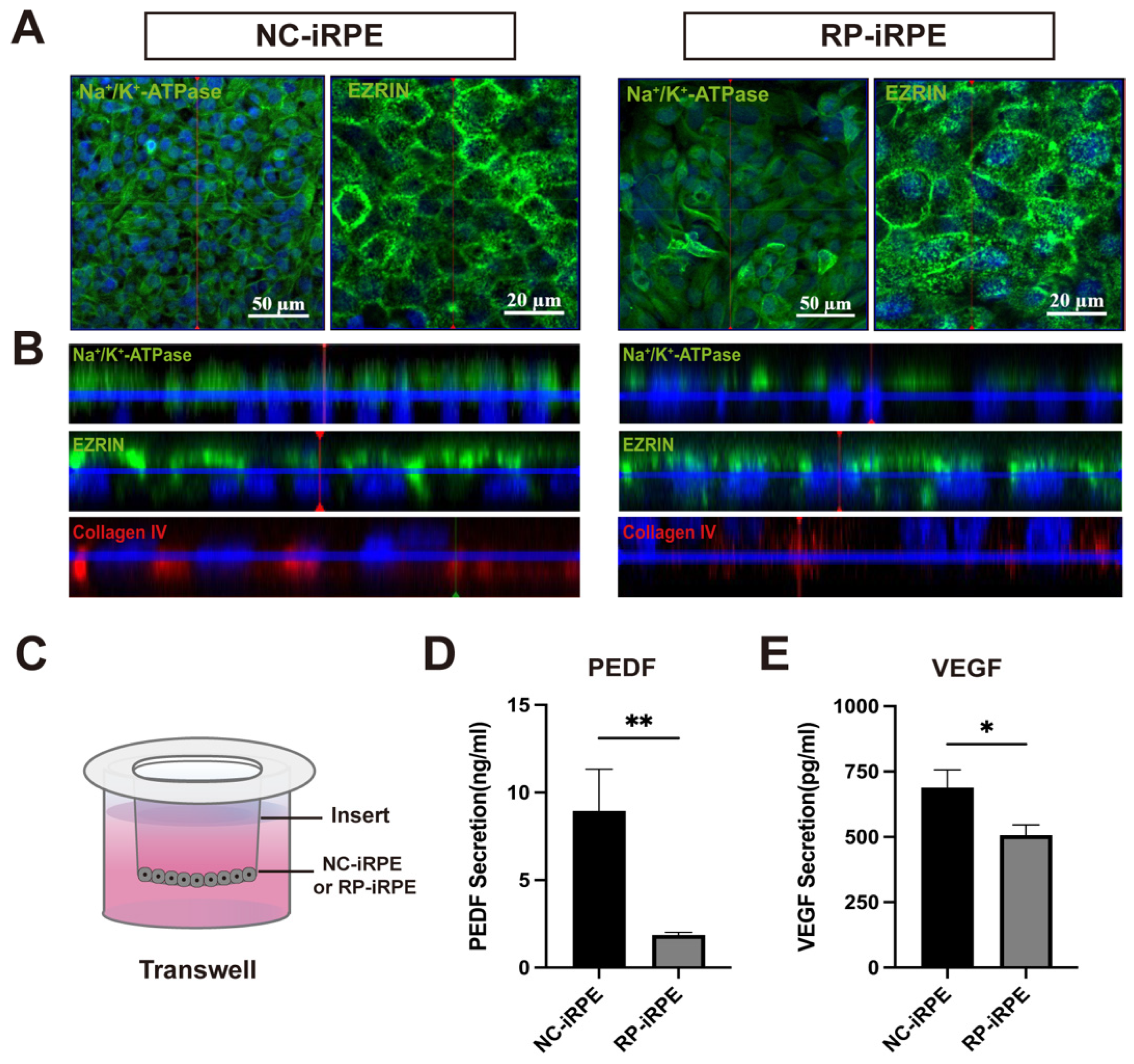
2.1. Generation and Characterization of NC-iRPE and RP-iRPE
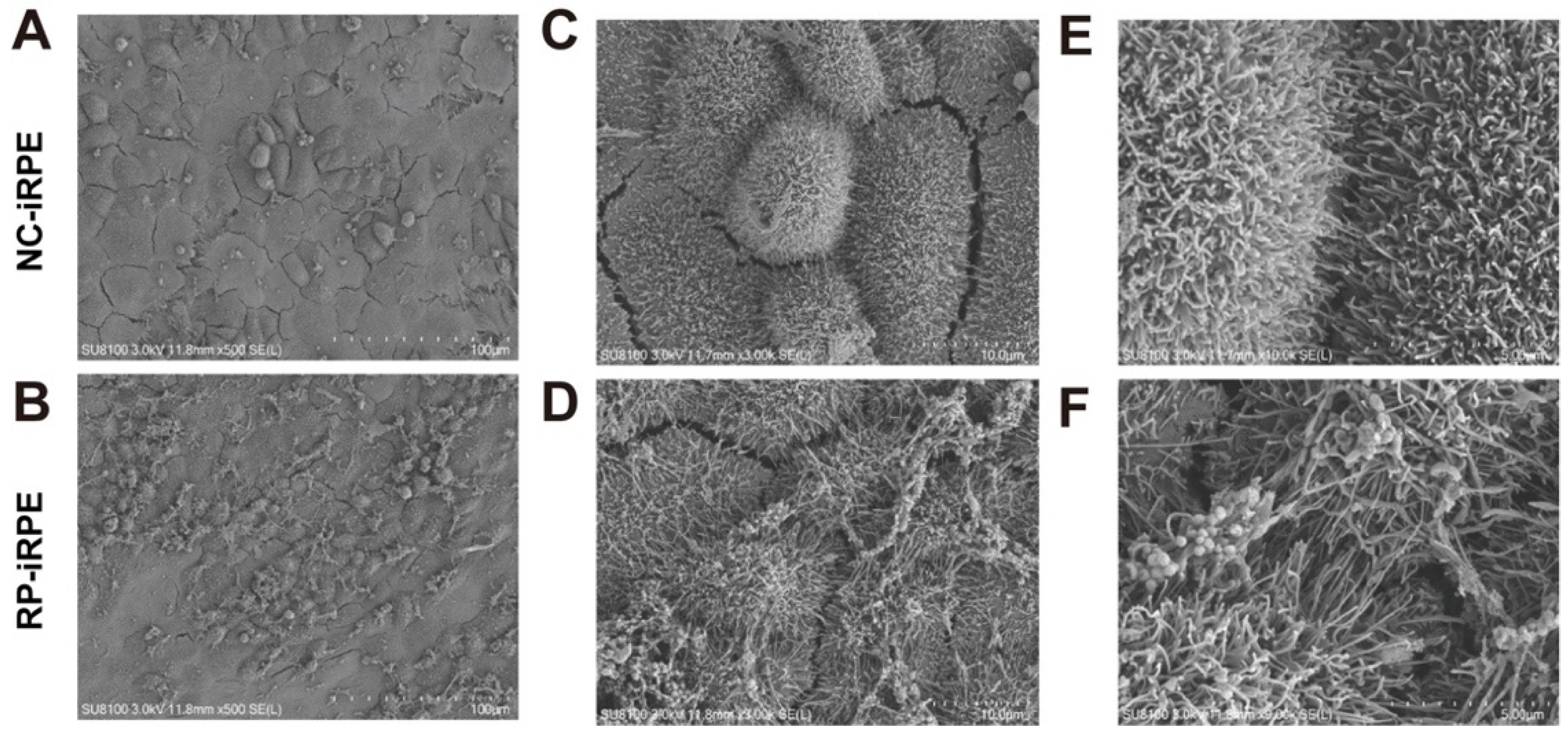
2.2. Association of PRPF6 Mutation with Changes in RPE Cell Polarity
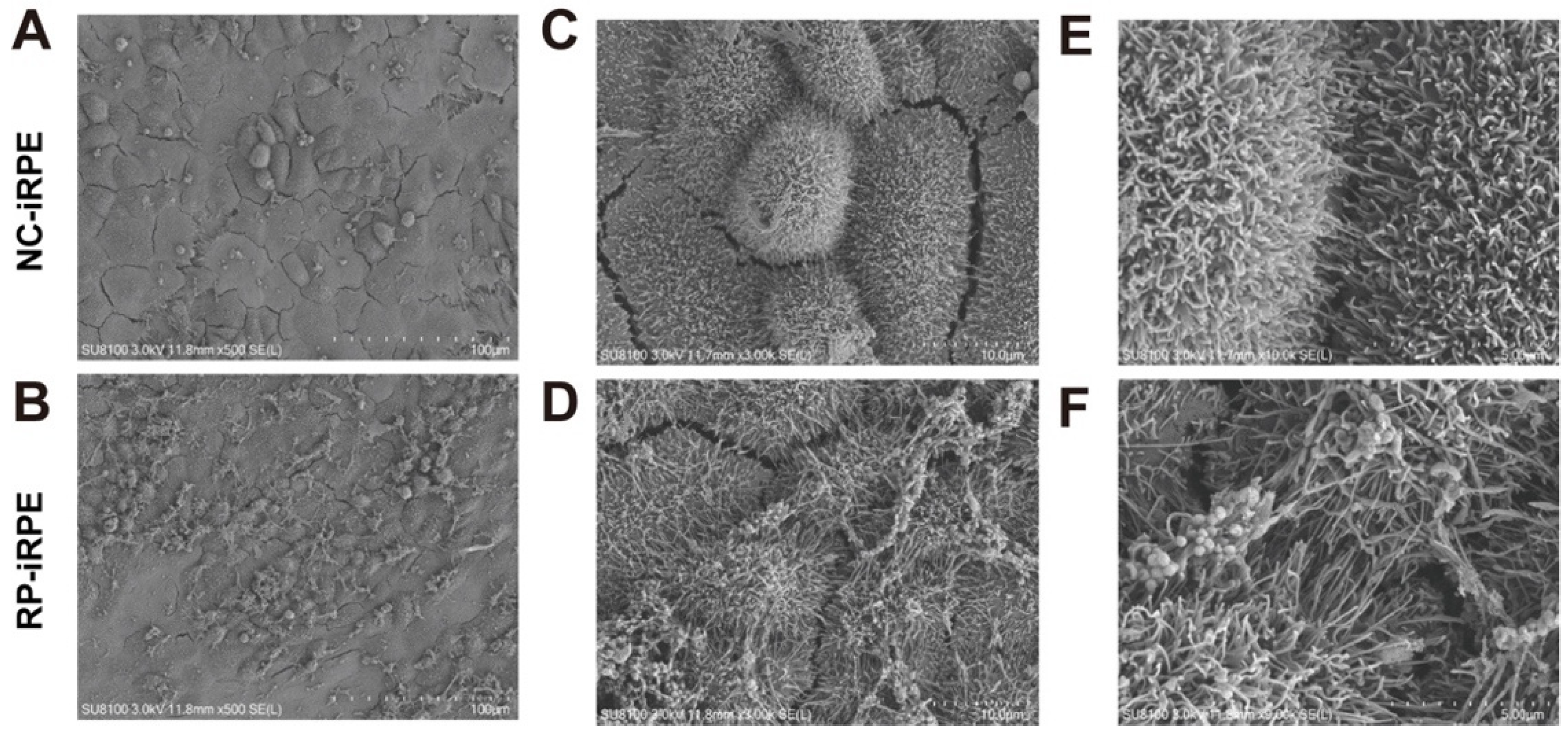
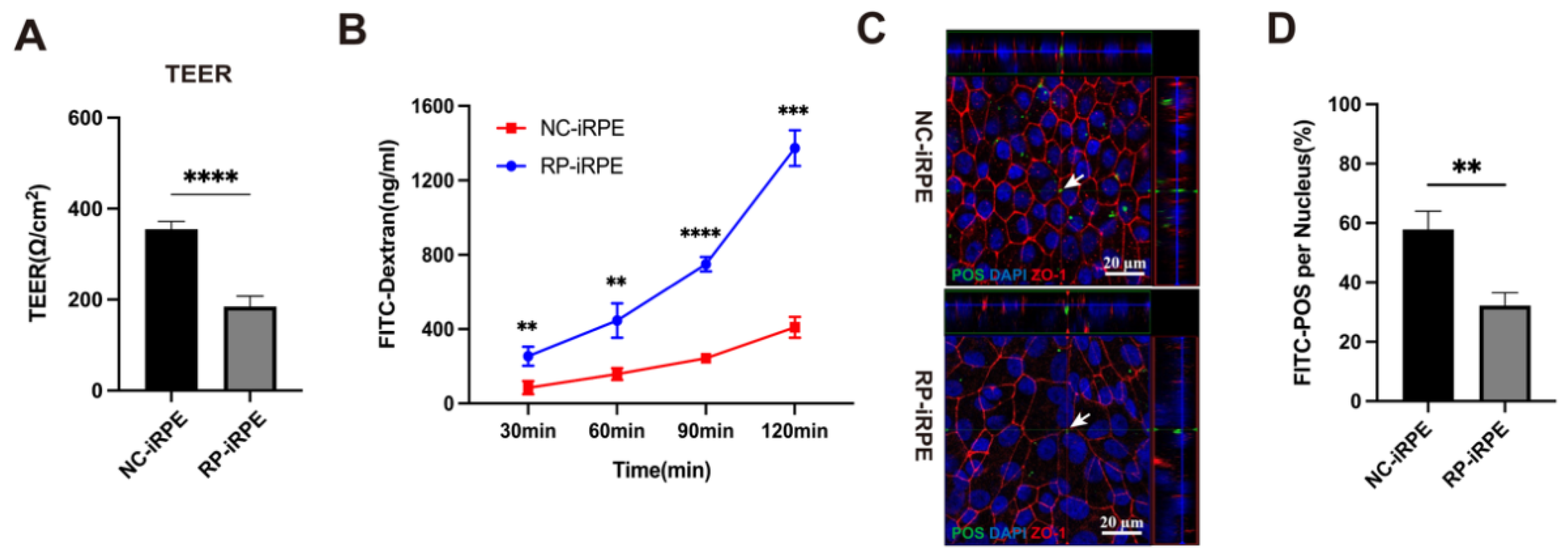
2.3. Aberrant Barrier Function in RPE Cells with PRPF6 Mutation
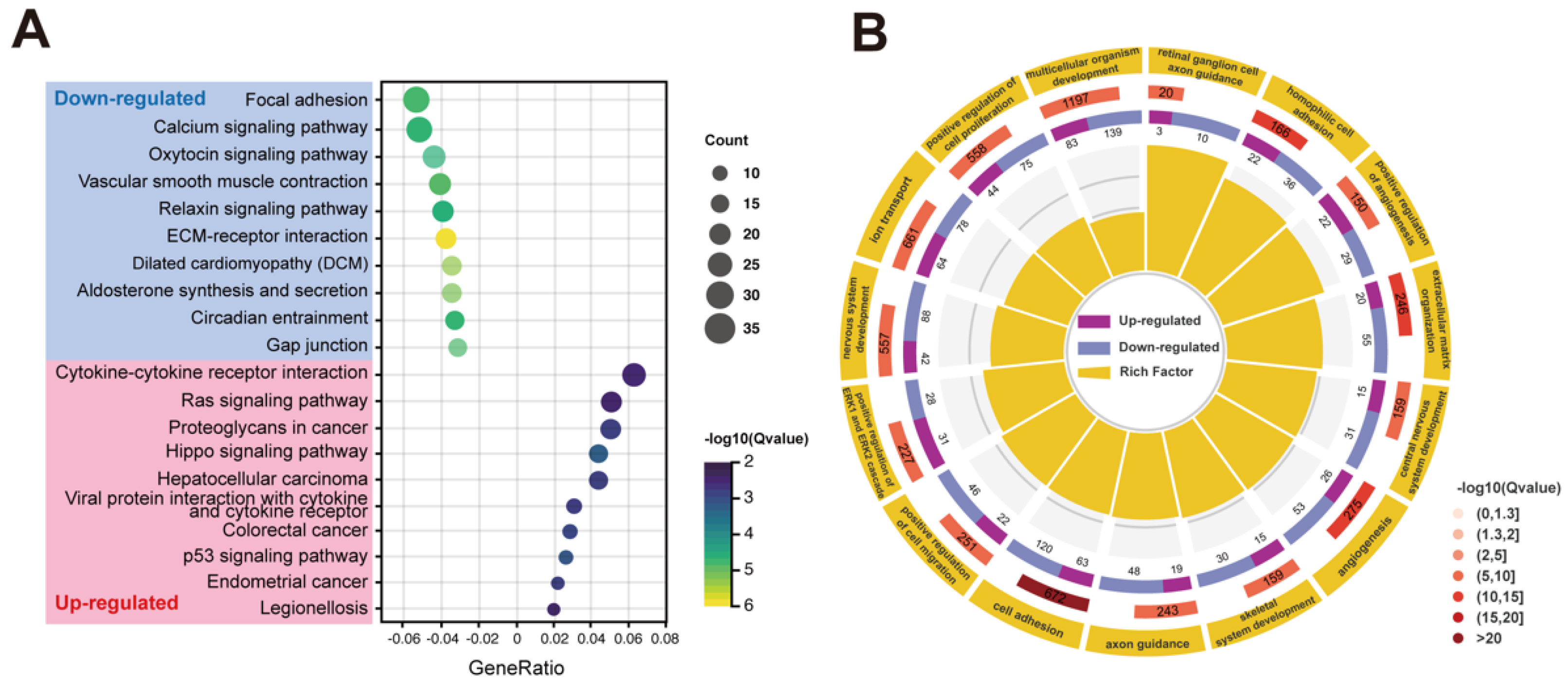
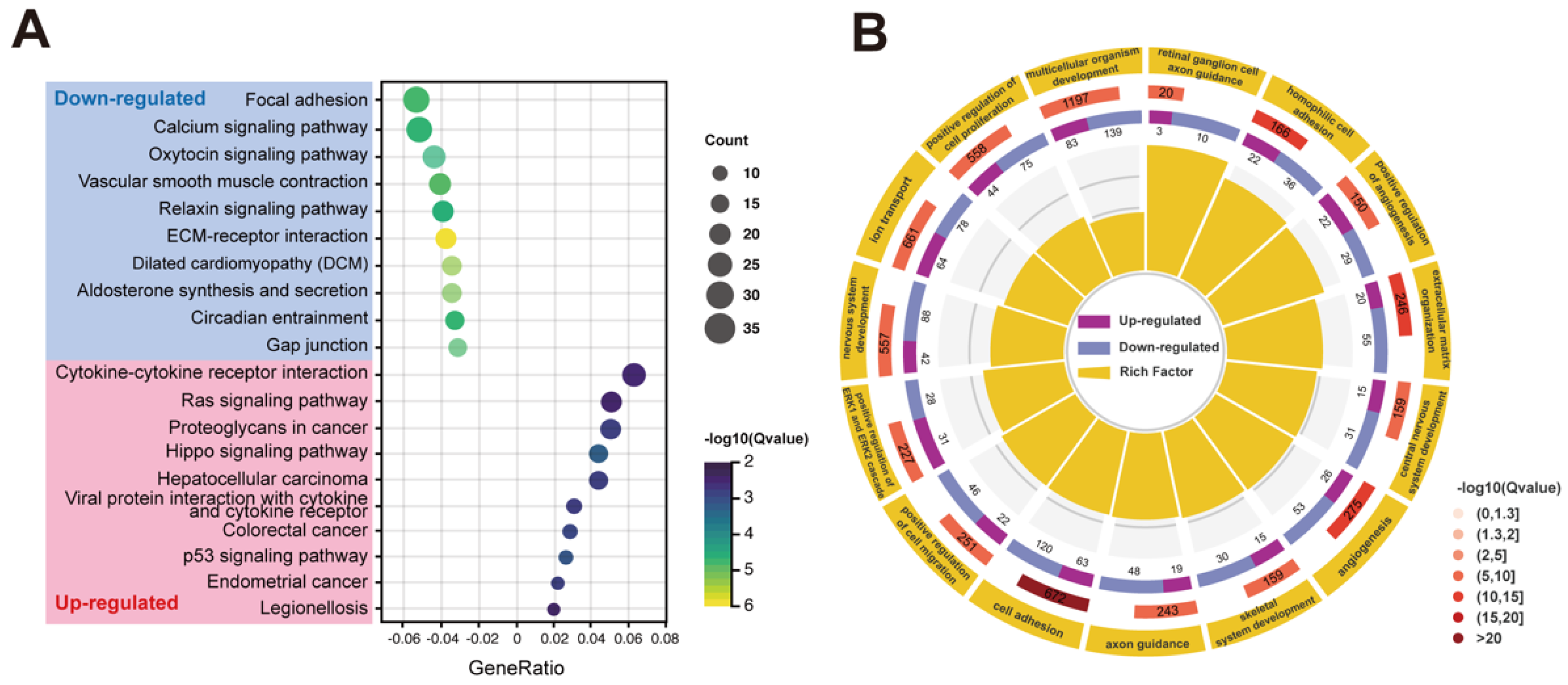
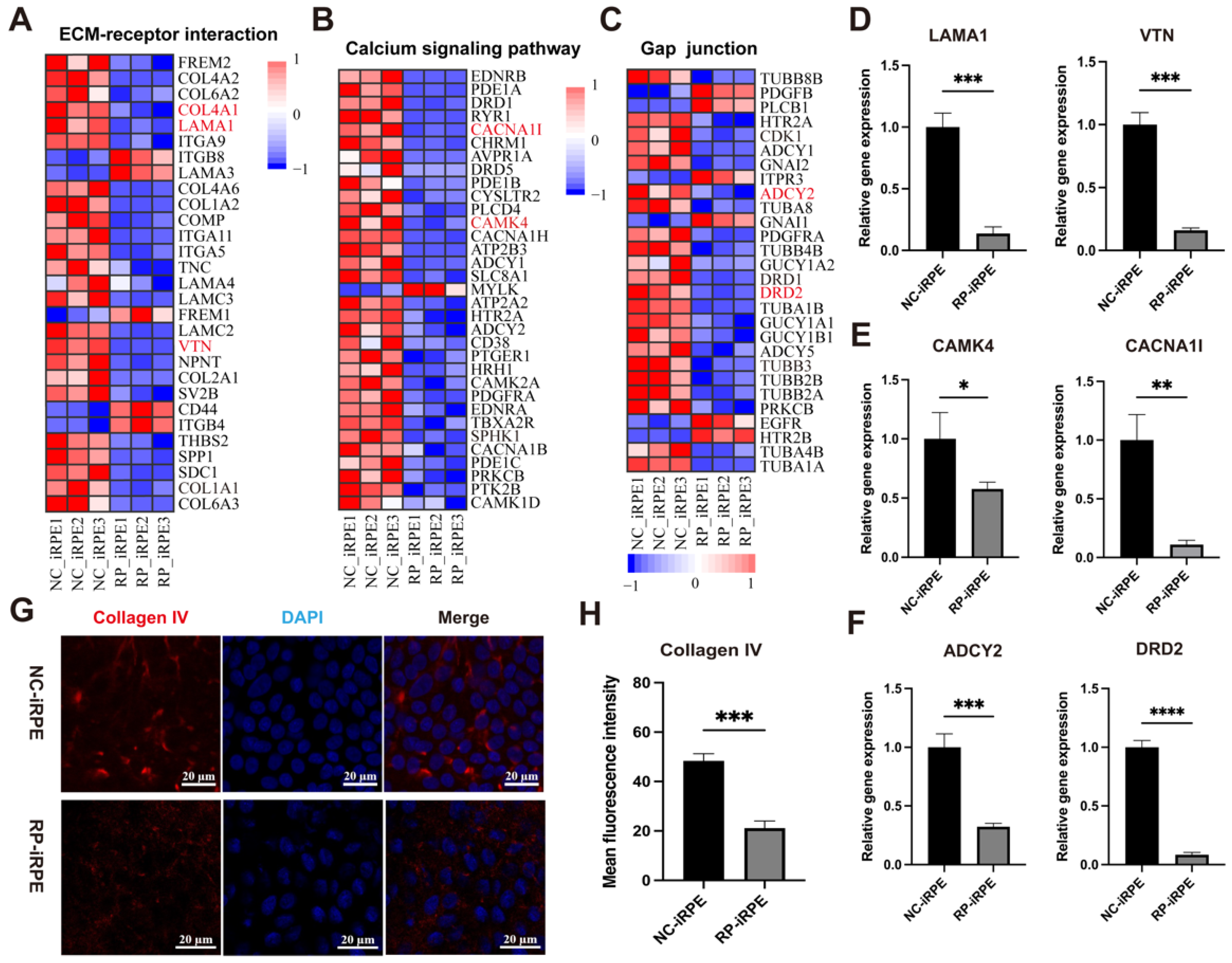
2.4. Transcriptome Analysis of RPE Cells with PRPF6 Mutation
3. Discussion
4. Materials and Methods
4.1. iPSCs Culture and Passaging
4.2. iPSCs Differentiation to RPE Cells
4.3. Immunofluorescence Staining
4.4. Reverse Transcription PCR and Quantitative Real-Time PCR
4.5. Western Blotting
4.6. TEER Assay
4.7. SEM Observation
4.8. ELISA
4.9. Permeability Assay
4.10. Phagocytosis Assay
4.11. RNA-Sequencing Analyses
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Species | Supplier | Catalog Number |
|---|---|---|---|
| ZO-1 | mouse | Invitrogen | 33-9100 |
| PRPF6 | rabbit | Invitrogen | PA5-61428 |
| MITF | rabbit | Invitrogen | MA5-32554 |
| CRALBP | rabbit | Proteintech | 15356-1-AP |
| RPE65 | mouse | Abcam | ab13826 |
| Tyrosinase | rabbit | Abcam | ab170905 |
| Ezrin | rabbit | GeneTex | GTX111709 |
| Na+/K+-ATPase | mouse | Santa Cruz | sc-48345 |
| Collagen IV | mouse | Abcam | ab86042 |
| ID | Pathway | Gene Num | Ratio | Q Value |
|---|---|---|---|---|
| Downregulated | ||||
| ko04512 | ECM-receptor interaction | 24 | 0.038 | 4.90 × 10−8 |
| ko05414 | Dilated cardiomyopathy (DCM) | 22 | 0.035 | 4.40 × 10−6 |
| ko04925 | Aldosterone synthesis and secretion | 22 | 0.035 | 6.31 × 10−6 |
| ko04270 | Vascular smooth muscle contraction | 26 | 0.041 | 1.19 × 10−5 |
| ko04540 | Gap junction | 20 | 0.032 | 1.36 × 10−5 |
| ko04510 | Focal adhesion | 34 | 0.054 | 1.50 × 10−5 |
| ko04713 | Circadian entrainment | 21 | 0.033 | 1.85 × 10−5 |
| ko04020 | Calcium signaling pathway | 33 | 0.052 | 1.96 × 10−5 |
| ko04926 | Relaxin signaling pathway | 25 | 0.040 | 2.34 × 10−5 |
| ko04921 | Oxytocin signaling pathway | 28 | 0.044 | 2.34 × 10−5 |
| Upregulated | ||||
| ko04390 | Hippo signaling pathway | 20 | 0.043 | 5.85 × 10−4 |
| ko04115 | p53 signaling pathway | 12 | 0.026 | 8.29 × 10−4 |
| ko05210 | Colorectal cancer | 13 | 0.028 | 1.31 × 10−3 |
| ko05213 | Endometrial cancer | 10 | 0.022 | 1.70 × 10−3 |
| ko05205 | Proteoglycans in cancer | 23 | 0.050 | 1.70 × 10−3 |
| ko05225 | Hepatocellular carcinoma | 20 | 0.043 | 1.75 × 10−3 |
| ko04061 | Viral protein interaction with cytokine and cytokine receptor | 14 | 0.030 | 1.85 × 10−3 |
| ko04060 | Cytokine-cytokine receptor interaction | 29 | 0.063 | 3.58 × 10−3 |
| ko04014 | Ras signaling pathway | 24 | 0.052 | 4.27 × 10−3 |
| ko05134 | Legionellosis | 9 | 0.020 | 4.67 × 10−3 |
| ID | Term | Gene Num | Ratio | Q Value |
|---|---|---|---|---|
| GO:0007155 | cell adhesion | 183 | 0.27 | 5.70 × 10−21 |
| GO:0007156 | homophilic cell adhesion via plasma membrane adhesion molecules | 58 | 0.35 | 3.99 × 10−12 |
| GO:0030198 | extracellular matrix organization | 75 | 0.30 | 7.64 × 10−12 |
| GO:0001525 | angiogenesis | 79 | 0.29 | 5.73 × 10−11 |
| GO:0045766 | positive regulation of angiogenesis | 51 | 0.34 | 2.41 × 10−10 |
| GO:0007399 | nervous system development | 130 | 0.23 | 4.43 × 10−10 |
| GO:0007411 | axon guidance | 67 | 0.28 | 1.02 × 10−8 |
| GO:0030335 | positive regulation of cell migration | 68 | 0.27 | 1.71 × 10−8 |
| GO:0006811 | ion transport | 142 | 0.21 | 2.33 × 10−8 |
| GO:0031290 | retinal ganglion cell axon guidance | 13 | 0.65 | 1.88 × 10−7 |
| GO:0007417 | central nervous system development | 46 | 0.29 | 4.51 × 10−7 |
| GO:0008284 | positive regulation of cell proliferation | 119 | 0.21 | 4.83 × 10−7 |
| GO:0070374 | positive regulation of ERK1 and ERK2 cascade | 59 | 0.26 | 6.81 × 10−7 |
| GO:0007275 | multicellular organism development | 222 | 0.19 | 1.10 × 10−6 |
| GO:0001501 | skeletal system development | 45 | 0.28 | 1.17 × 10−6 |
References
- Liu, W.; Liu, S.; Li, P.; Yao, K. Retinitis Pigmentosa: Progress in Molecular Pathology and Biotherapeutical Strategies. Int. J. Mol. Sci. 2022, 23, 4883. [Google Scholar] [CrossRef]
- Sun, X.; Sun, P.; Liu, L.; Jiang, P.; Li, Y. Ferulic acid attenuates microglia-mediated neuroinflammation in retinal degeneration. BMC Ophthalmol. 2021, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Girardot, P.E.; Sellers, J.T.; Li, Y.; Wang, J.; Chrenek, M.A.; Wu, W.; Skelton, H.; Nickerson, J.M.; Pardue, M.T.; et al. Wheel running exercise protects against retinal degeneration in the I307N rhodopsin mouse model of inducible autosomal dominant retinitis pigmentosa. Mol. Vis. 2019, 25, 462–476. [Google Scholar]
- Ballios, B.G.; Place, E.M.; Martinez-Velazquez, L.; Pierce, E.A.; Comander, J.I.; Huckfeldt, R.M. Beyond Sector Retinitis Pigmentosa: Expanding the Phenotype and Natural History of the Rhodopsin Gene Codon 106 Mutation (Gly-to-Arg) in Autosomal Dominant Retinitis Pigmentosa. Genes 2021, 12, 1853. [Google Scholar] [CrossRef]
- Matera, A.G.; Wang, Z. A day in the life of the spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Will, C.L.; Luhrmann, R. Spliceosome structure and function. Cold Spring Harb. Perspect. Biol. 2011, 3, a003707. [Google Scholar] [CrossRef]
- Griffin, C.; Saint-Jeannet, J.P. Spliceosomopathies: Diseases and mechanisms. Dev. Dyn. 2020, 249, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Arzalluz-Luque, A.; Cabrera, J.L.; Skottman, H.; Benguria, A.; Bolinches-Amoros, A.; Cuenca, N.; Lupo, V.; Dopazo, A.; Tarazona, S.; Delas, B.; et al. Mutant PRPF8 Causes Widespread Splicing Changes in Spliceosome Components in Retinitis Pigmentosa Patient iPSC-Derived RPE Cells. Front. Neurosci. 2021, 15, 636969. [Google Scholar] [CrossRef]
- Ruzickova, S.; Stanek, D. Mutations in spliceosomal proteins and retina degeneration. RNA Biol. 2017, 14, 544–552. [Google Scholar] [CrossRef]
- Klimesova, K.; Vojackova, J.; Radivojevic, N.; Vandermoere, F.; Bertrand, E.; Verheggen, C.; Stanek, D. TSSC4 is a component of U5 snRNP that promotes tri-snRNP formation. Nat. Commun. 2021, 12, 3646. [Google Scholar] [CrossRef] [PubMed]
- Wood, K.A.; Eadsforth, M.A.; Newman, W.G.; O’Keefe, R.T. The Role of the U5 snRNP in Genetic Disorders and Cancer. Front. Genet. 2021, 12, 636620. [Google Scholar] [CrossRef]
- Adler, A.S.; McCleland, M.L.; Yee, S.; Yaylaoglu, M.; Hussain, S.; Cosino, E.; Quinones, G.; Modrusan, Z.; Seshagiri, S.; Torres, E.; et al. An integrative analysis of colon cancer identifies an essential function for PRPF6 in tumor growth. Genes. Dev. 2014, 28, 1068–1084. [Google Scholar] [CrossRef]
- Song, H.; Sun, N.; Lin, L.; Wei, S.; Zeng, K.; Liu, W.; Wang, C.; Zhong, X.; Wang, M.; Wang, S.; et al. Splicing factor PRPF6 upregulates oncogenic androgen receptor signaling pathway in hepatocellular carcinoma. Cancer Sci. 2020, 111, 3665–3678. [Google Scholar] [CrossRef]
- Liu, W.; Wang, C.; Wang, S.; Zeng, K.; Wei, S.; Sun, N.; Sun, G.; Wang, M.; Zou, R.; Liu, W.; et al. PRPF6 promotes androgen receptor/androgen receptor-variant 7 actions in castration-resistant prostate cancer cells. Int. J. Biol. Sci. 2021, 17, 188–203. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, X.; Li, S.; Jiang, H.; Sun, W.; Wang, P.; Zhang, Q. Landscape of pathogenic variants in six pre-mRNA processing factor genes for retinitis pigmentosa based on large in-house data sets and database comparisons. Acta Ophthalmol. 2022. [Google Scholar] [CrossRef]
- Huang, X.F.; Huang, F.; Wu, K.C.; Wu, J.; Chen, J.; Pang, C.P.; Lu, F.; Qu, J.; Jin, Z.B. Genotype-phenotype correlation and mutation spectrum in a large cohort of patients with inherited retinal dystrophy revealed by next-generation sequencing. Genet. Med. 2015, 17, 271–278. [Google Scholar] [CrossRef]
- Tanackovic, G.; Ransijn, A.; Ayuso, C.; Harper, S.; Berson, E.L.; Rivolta, C. A missense mutation in PRPF6 causes impairment of pre-mRNA splicing and autosomal-dominant retinitis pigmentosa. Am. J. Hum. Genet. 2011, 88, 643–649. [Google Scholar] [CrossRef]
- O’Neill, A.C.; Uzbas, F.; Antognolli, G.; Merino, F.; Draganova, K.; Jack, A.; Zhang, S.; Pedini, G.; Schessner, J.P.; Cramer, K.; et al. Spatial centrosome proteome of human neural cells uncovers disease-relevant heterogeneity. Science 2022, 376, eabf9088. [Google Scholar] [CrossRef]
- Fields, M.A.; Del Priore, L.V.; Adelman, R.A.; Rizzolo, L.J. Interactions of the choroid, Bruch’s membrane, retinal pigment epithelium, and neurosensory retina collaborate to form the outer blood-retinal-barrier. Prog. Retin. Eye Res. 2020, 76, 100803. [Google Scholar] [CrossRef]
- Lakkaraju, A.; Umapathy, A.; Tan, L.X.; Daniele, L.; Philp, N.J.; Boesze-Battaglia, K.; Williams, D.S. The cell biology of the retinal pigment epithelium. Prog. Retin. Eye Res. 2020, 78, 100846. [Google Scholar] [CrossRef]
- Ao, J.; Wood, J.P.; Chidlow, G.; Gillies, M.C.; Casson, R.J. Retinal pigment epithelium in the pathogenesis of age-related macular degeneration and photobiomodulation as a potential therapy? Clin. Exp. Ophthalmol. 2018, 46, 670–686. [Google Scholar] [CrossRef]
- Chakrabarty, K.; Shetty, R.; Ghosh, A. Corneal cell therapy: With iPSCs, it is no more a far-sight. Stem Cell Res. Ther. 2018, 9, 287. [Google Scholar] [CrossRef]
- Liu, S.; Xie, B.; Song, X.; Zheng, D.; He, L.; Li, G.; Gao, G.; Peng, F.; Yu, M.; Ge, J.; et al. Self-Formation of RPE Spheroids Facilitates Enrichment and Expansion of hiPSC-Derived RPE Generated on Retinal Organoid Induction Platform. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5659–5669. [Google Scholar] [CrossRef]
- Reichman, S.; Slembrouck, A.; Gagliardi, G.; Chaffiol, A.; Terray, A.; Nanteau, C.; Potey, A.; Belle, M.; Rabesandratana, O.; Duebel, J.; et al. Generation of Storable Retinal Organoids and Retinal Pigmented Epithelium from Adherent Human iPS Cells in Xeno-Free and Feeder-Free Conditions. Stem Cells 2017, 35, 1176–1188. [Google Scholar] [CrossRef]
- Shrestha, R.; Wen, Y.T.; Tsai, R.K. Effective Differentiation and Biological Characterization of Retinal Pigment Epithelium Derived from Human Induced Pluripotent Stem Cells. Curr. Eye Res. 2020, 45, 1155–1167. [Google Scholar] [CrossRef]
- Regent, F.; Morizur, L.; Lesueur, L.; Habeler, W.; Plancheron, A.; Ben M’Barek, K.; Monville, C. Automation of human pluripotent stem cell differentiation toward retinal pigment epithelial cells for large-scale productions. Sci. Rep. 2019, 9, 10646. [Google Scholar] [CrossRef]
- Yang, C.; Georgiou, M.; Atkinson, R.; Collin, J.; Al-Aama, J.; Nagaraja-Grellscheid, S.; Johnson, C.; Ali, R.; Armstrong, L.; Mozaffari-Jovin, S.; et al. Pre-mRNA Processing Factors and Retinitis Pigmentosa: RNA Splicing and Beyond. Front. Cell Dev. Biol. 2021, 9, 700276. [Google Scholar] [CrossRef]
- Zhou, Y.; Jing, Y.; Mao, S.; Liu, J.; Cui, Z.; Wang, Y.; Chen, J.; Chan, H.f.; Tang, S.; Chen, J. Establishment of induced pluripotent stem cell line CSUASOi004-A by reprogramming peripheral blood mononuclear cells of a PRPF6-related dominant retinitis pigmentosa patient. Stem Cell Res. 2020, 45, 101793. [Google Scholar] [CrossRef]
- Reichman, S.; Terray, A.; Slembrouck, A.; Nanteau, C.; Orieux, G.; Habeler, W.; Nandrot, E.F.; Sahel, J.A.; Monville, C.; Goureau, O. From confluent human iPS cells to self-forming neural retina and retinal pigmented epithelium. Proc. Natl. Acad. Sci. USA 2014, 111, 8518–8523. [Google Scholar] [CrossRef]
- Marmorstein, A.D. The polarity of the retinal pigment epithelium. Traffic 2001, 2, 867–872. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, Y.N.; Yoon, Y.H.; Seo, E.J.; Seo, G.H.; Keum, C.; Lee, B.H.; Lee, J.Y. Diverse Genetic Landscape of Suspected Retinitis Pigmentosa in a Large Korean Cohort. Genes 2021, 12, 675. [Google Scholar] [CrossRef]
- Farkas, M.H.; Lew, D.S.; Sousa, M.E.; Bujakowska, K.; Chatagnon, J.; Bhattacharya, S.S.; Pierce, E.A.; Nandrot, E.F. Mutations in pre-mRNA processing factors 3, 8, and 31 cause dysfunction of the retinal pigment epithelium. Am. J. Pathol. 2014, 184, 2641–2652. [Google Scholar] [CrossRef]
- Valdes-Sanchez, L.; Calado, S.M.; de la Cerda, B.; Aramburu, A.; Garcia-Delgado, A.B.; Massalini, S.; Montero-Sanchez, A.; Bhatia, V.; Rodriguez-Bocanegra, E.; Diez-Lloret, A.; et al. Retinal pigment epithelium degeneration caused by aggregation of PRPF31 and the role of HSP70 family of proteins. Mol. Med. 2019, 26, 1. [Google Scholar] [CrossRef]
- Penney, J.; Ralvenius, W.T.; Tsai, L.H. Modeling Alzheimer’s disease with iPSC-derived brain cells. Mol. Psychiatry 2020, 25, 148–167. [Google Scholar] [CrossRef]
- O’Hara-Wright, M.; Mobini, S.; Gonzalez-Cordero, A. Bioelectric Potential in Next-Generation Organoids: Electrical Stimulation to Enhance 3D Structures of the Central Nervous System. Front. Cell Dev. Biol. 2022, 10, 901652. [Google Scholar] [CrossRef]
- Graziotto, J.J.; Farkas, M.H.; Bujakowska, K.; Deramaudt, B.M.; Zhang, Q.; Nandrot, E.F.; Inglehearn, C.F.; Bhattacharya, S.S.; Pierce, E.A. Three gene-targeted mouse models of RNA splicing factor RP show late-onset RPE and retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2011, 52, 190–198. [Google Scholar] [CrossRef]
- Bujakowska, K.; Maubaret, C.; Chakarova, C.F.; Tanimoto, N.; Beck, S.C.; Fahl, E.; Humphries, M.M.; Kenna, P.F.; Makarov, E.; Makarova, O.; et al. Study of gene-targeted mouse models of splicing factor gene Prpf31 implicated in human autosomal dominant retinitis pigmentosa (RP). Investig. Ophthalmol. Vis. Sci. 2009, 50, 5927–5933. [Google Scholar] [CrossRef]
- Buskin, A.; Zhu, L.; Chichagova, V.; Basu, B.; Mozaffari-Jovin, S.; Dolan, D.; Droop, A.; Collin, J.; Bronstein, R.; Mehrotra, S.; et al. Disrupted alternative splicing for genes implicated in splicing and ciliogenesis causes PRPF31 retinitis pigmentosa. Nat. Commun. 2018, 9, 4234. [Google Scholar] [CrossRef]
- Georgiou, M.; Yang, C.; Atkinson, R.; Pan, K.T.; Buskin, A.; Molina, M.M.; Collin, J.; Al-Aama, J.; Goertler, F.; Ludwig, S.E.J.; et al. Activation of autophagy reverses progressive and deleterious protein aggregation in PRPF31 patient-induced pluripotent stem cell-derived retinal pigment epithelium cells. Clin. Transl. Med. 2022, 12, e759. [Google Scholar] [CrossRef]
- Foltz, L.P.; Howden, S.E.; Thomson, J.A.; Clegg, D.O. Functional Assessment of Patient-Derived Retinal Pigment Epithelial Cells Edited by CRISPR/Cas9. Int. J. Mol. Sci. 2018, 19, 4127. [Google Scholar] [CrossRef]
- Booij, J.C.; Baas, D.C.; Beisekeeva, J.; Gorgels, T.G.; Bergen, A.A. The dynamic nature of Bruch’s membrane. Prog. Retin. Eye Res. 2010, 29, 1–18. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Chen, Y.H.; Zhang, T.; Liu, S.J.; Bai, X.Y.; Huang, X.Y.; Jiang, M.; Sun, X.D. Improvement of human embryonic stem cell-derived retinal pigment epithelium cell adhesion, maturation, and function through coating with truncated recombinant human vitronectin. Int. J. Ophthalmol. 2021, 14, 1160–1167. [Google Scholar] [CrossRef]
- da Cruz, L.; Fynes, K.; Georgiadis, O.; Kerby, J.; Luo, Y.H.; Ahmado, A.; Vernon, A.; Daniels, J.T.; Nommiste, B.; Hasan, S.M.; et al. Phase 1 clinical study of an embryonic stem cell-derived retinal pigment epithelium patch in age-related macular degeneration. Nat. Biotechnol. 2018, 36, 328–337. [Google Scholar] [CrossRef]
- Scimone, C.; Bramanti, P.; Ruggeri, A.; Katsarou, Z.; Donato, L.; Sidoti, A.; D’Angelo, R. Detection of Novel Mutation in Ccm3 Causes Familial Cerebral Cavernous Malformations. J. Mol. Neurosci. 2015, 57, 400–403. [Google Scholar] [CrossRef]
- Scimone, C.; Bramanti, P.; Ruggeri, A.; Donato, L.; Alafaci, C.; Crisafulli, C.; Mucciardi, M.; Rinaldi, C.; Sidoti, A.; D’Angelo, R. CCM3/SERPINI1 bidirectional promoter variants in patients with cerebral cavernous malformations: A molecular and functional study. BMC Med. Genet. 2016, 17, 74. [Google Scholar] [CrossRef]
- Scimone, C.; Donato, L.; Alibrandi, S.; Esposito, T.; Alafaci, C.; D’Angelo, R.; Sidoti, A. Transcriptome analysis provides new molecular signatures in sporadic Cerebral Cavernous Malformation endothelial cells. Biochim. Biophys. Acta Mol. Bas. Dis. 2020, 1866, 165956. [Google Scholar] [CrossRef]
- Wimmers, S.; Karl, M.O.; Strauss, O. Ion channels in the RPE. Prog. Retin. Eye Res. 2007, 26, 263–301. [Google Scholar] [CrossRef]
- Viheriala, T.; Sorvari, J.; Ihalainen, T.O.; Moro, A.; Gronroos, P.; Schlie-Wolter, S.; Chichkov, B.; Skottman, H.; Nymark, S.; Ilmarinen, T. Culture surface protein coatings affect the barrier properties and calcium signalling of hESC-RPE. Sci. Rep. 2021, 11, 933. [Google Scholar] [CrossRef]
- El Ghaleb, Y.; Schneeberger, P.E.; Fernandez-Quintero, M.L.; Geisler, S.M.; Pelizzari, S.; Polstra, A.M.; van Hagen, J.M.; Denecke, J.; Campiglio, M.; Liedl, K.R.; et al. CACNA1I gain-of-function mutations differentially affect channel gating and cause neurodevelopmental disorders. Brain 2021, 144, 2092–2106. [Google Scholar] [CrossRef]
- Zech, M.; Bardakjian, T.M.; Stoklosa, M.; Ploski, R.; Jech, R.; Gonzalez-Alegre, P.; Winkelmann, J. A Neurodevelopmental Disorder With Dystonia and Chorea Resulting From Clustering CAMK4 Variants. Mov. Disord. 2021, 36, 520–521. [Google Scholar] [CrossRef]
- Rizzolo, L.J. Development and Role of Tight Junctions in the Retinal Pigment Epithelium. Int. Rev. Cytol. 2007, 258, 195–234. [Google Scholar]
- Ovando-Roche, P.; Georgiadis, A.; Smith, A.J.; Pearson, R.A.; Ali, R.R. Harnessing the Potential of Human Pluripotent Stem Cells and Gene Editing for the Treatment of Retinal Degeneration. Curr. Stem Cell Rep. 2017, 3, 112–123. [Google Scholar] [CrossRef]
- Achberger, K.; Probst, C.; Haderspeck, J.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging organoid and organ-on-a-chip technology to generate complex multi-layer tissue models in a human retina-on-a-chip platform. eLife 2019, 8, e46188. [Google Scholar] [CrossRef] [PubMed]
- Abdayem, R.; Callejon, S.; Portes, P.; Kirilov, P.; Demarne, F.; Pirot, F.; Jannin, V.; Haftek, M. Modulation of transepithelial electric resistance (TEER) in reconstructed human epidermis by excipients known to permeate intestinal tight junctions. Exp. Dermatol. 2015, 24, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Xie, M.; Gademann, F.; Cao, J.; Wang, P.; Guo, Y.; Zhang, L.; Su, T.; Zhang, J.; Chen, J. Protective effects of human iPS-derived retinal pigmented epithelial cells on retinal degenerative disease. Stem Cell Res. Ther. 2020, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Wang, Y.; Cui, Z.; Li, H.; Li, S.; Yang, X.; Yan, X.; Ding, C.; Tang, S.; Chen, J. The Construction of Retinal Pigment Epithelium Sheets with Enhanced Characteristics and Cilium Assembly Using iPS Conditioned Medium and Small Incision Lenticule Extraction Derived Lenticules. Acta Biomater. 2019, 92, 115–131. [Google Scholar] [CrossRef]
- Trudeau, K.; Roy, S.; Guo, W.; Hernandez, C.; Villarroel, M.; Simo, R.; Roy, S. Fenofibric acid reduces fibronectin and collagen type IV overexpression in human retinal pigment epithelial cells grown in conditions mimicking the diabetic milieu: Functional implications in retinal permeability. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6348–6354. [Google Scholar] [CrossRef] [PubMed]
- Parinot, C.; Rieu, Q.; Chatagnon, J.; Finnemann, S.C.; Nandrot, E.F. Large-scale purification of porcine or bovine photoreceptor outer segments for phagocytosis assays on retinal pigment epithelial cells. J. Vis. Exp. 2014, 94, e52100. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, F.; Mao, Y.; Finnemann, S.C. Advanced Analysis of Photoreceptor Outer Segment Phagocytosis by RPE Cells in Culture. Methods Mol. Biol. 2019, 1834, 95–108. [Google Scholar] [PubMed]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Tan, F.; Sun, X.; Cui, Z.; Gu, J.; Mao, S.; Chan, H.F.; Tang, S.; Chen, J. Aberrant Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem Cells of a Retinitis Pigmentosa Patient with the PRPF6 Mutation. Int. J. Mol. Sci. 2022, 23, 9049. https://doi.org/10.3390/ijms23169049
Liang Y, Tan F, Sun X, Cui Z, Gu J, Mao S, Chan HF, Tang S, Chen J. Aberrant Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem Cells of a Retinitis Pigmentosa Patient with the PRPF6 Mutation. International Journal of Molecular Sciences. 2022; 23(16):9049. https://doi.org/10.3390/ijms23169049
Chicago/Turabian StyleLiang, Yuqin, Feng Tan, Xihao Sun, Zekai Cui, Jianing Gu, Shengru Mao, Hon Fai Chan, Shibo Tang, and Jiansu Chen. 2022. "Aberrant Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem Cells of a Retinitis Pigmentosa Patient with the PRPF6 Mutation" International Journal of Molecular Sciences 23, no. 16: 9049. https://doi.org/10.3390/ijms23169049
APA StyleLiang, Y., Tan, F., Sun, X., Cui, Z., Gu, J., Mao, S., Chan, H. F., Tang, S., & Chen, J. (2022). Aberrant Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem Cells of a Retinitis Pigmentosa Patient with the PRPF6 Mutation. International Journal of Molecular Sciences, 23(16), 9049. https://doi.org/10.3390/ijms23169049