
miRNA in Ischemic Heart Disease and Its Potential as Biomarkers: A Comprehensive Review

 ,
,
Abstract
:1. Introduction
2. Current Outlooks for miRNAs in Ischemic Heart Disease
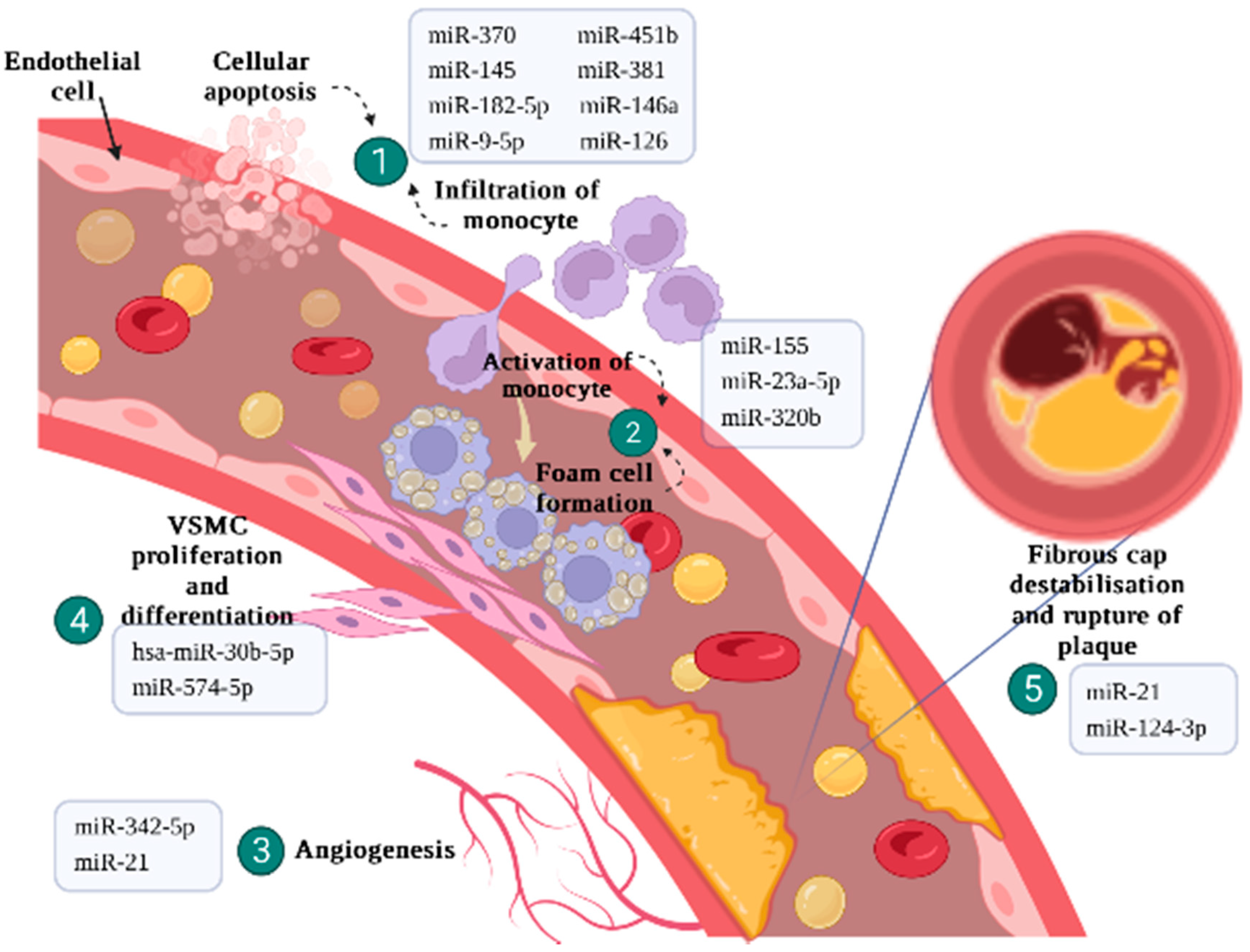
2.1. Endothelial Cells Regulation and Inflammatory Response
2.2. Activation of Monocytes and Differentiation of Macrophages
2.3. Plaque Angiogenesis
2.4. VSMC Proliferation and Differentiation
2.5. Fibrous Cap Destabilisation and Plaque Rupture
3. Therapeutic Potential of miRNAs in IHD
4. Caveats of Using miRNA as Diagnostic Biomarker in IHD
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P. Global burden of cardiovascular diseases and risk factors, 1990–2019: Update from the GBD 2019 study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Much, A.A.; Maor, E.; Asher, E.; Younis, A.; Xu, Y.; Lu, Y.; Liu, X.; Shu, J.; Bragazzi, N.L. Global, regional, and national burden of ischaemic heart disease and its attributable risk factors, 1990–2017: Results from the Global Burden of Disease Study 2017. Eur. Heart J. Qual. Care Clin. Outcomes 2022, 8, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Caceres, B.A.; Ancheta, A.J.; Dorsen, C.; Newlin-Lew, K.; Edmondson, D.; Hughes, T.L. A population-based study of the intersection of sexual identity and race/ethnicity on physiological risk factors for CVD among US adults (ages 18–59). Ethn. Health 2022, 27, 617–638. [Google Scholar] [CrossRef] [PubMed]
- Andersson, C.; Vasan, R.S. Epidemiology of cardiovascular disease in young individuals. Nat. Rev. Cardiol. 2018, 15, 230–240. [Google Scholar] [CrossRef]
- Bekelis, K.; Roberts, D.W.; Zhou, W.; Skinner, J.S. Fragmentation of care and the use of head computed tomography in patients with ischemic stroke. Circ. Cardiovasc. Qual. Outcomes 2014, 7, 430–436. [Google Scholar] [CrossRef]
- Dineva, S.; Uzunova, K.; Pavlova, V.; Filipova, E.; Kalinov, K.; Vekov, T. Comparative efficacy and safety of chlorthalidone and hydrochlorothiazide—meta-analysis. J. Hum. Hypertens. 2019, 33, 766–774. [Google Scholar] [CrossRef]
- Khan, M.A.; Hashim, M.J.; Mustafa, H.; Baniyas, M.Y.; Al Suwaidi, S.K.B.M.; AlKatheeri, R.; Alblooshi, F.M.K.; Almatrooshi, M.E.A.H.; Alzaabi, M.E.H.; Al Darmaki, R.S. Global epidemiology of ischemic heart disease: Results from the global burden of disease study. Cureus 2020, 12, e9349. [Google Scholar] [CrossRef]
- da Silva Júnior, W.F.; de Menezes, D.L.B.; de Oliveira, L.C.; Koester, L.S.; de Almeida, P.D.O.; Lima, E.S.; de Azevedo, E.P.; da Veiga Júnior, V.F.; Neves de Lima, Á.A. Inclusion complexes of β and HPβ-cyclodextrin with α, β amyrin and in vitro anti-inflammatory activity. Biomolecules 2019, 9, 241. [Google Scholar] [CrossRef]
- Melak, T.; Baynes, H.W. Circulating microRNAs as possible biomarkers for coronary artery disease: A narrative review. Ejifcc 2019, 30, 179. [Google Scholar]
- Hodas, R.; Polexa, Ș.A.; Rareș, M.; Benedek, T. Coronary Computed Tomography Angiography for Assesment of Stable Coronary Artery Disease–a Cost-effectiveness Perspective. J. Interdiscip. Med. 2021, 6, 37–42. [Google Scholar] [CrossRef]
- Fazmin, I.T.; Achercouk, Z.; Edling, C.E.; Said, A.; Jeevaratnam, K. Circulating microRNA as a Biomarker for Coronary Artery Disease. Biomolecules 2020, 10, 1354. [Google Scholar] [CrossRef] [PubMed]
- Meder, B.; Keller, A.; Vogel, B.; Haas, J.; Sedaghat-Hamedani, F.; Kayvanpour, E.; Just, S.; Borries, A.; Rudloff, J.; Leidinger, P. MicroRNA signatures in total peripheral blood as novel biomarkers for acute myocardial infarction. Basic Res. Cardiol. 2011, 106, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Siasos, G.; Bletsa, E.; Stampouloglou, P.K.; Oikonomou, E.; Tsigkou, V.; Paschou, S.A.; Vlasis, K.; Marinos, G.; Vavuranakis, M.; Stefanadis, C.; et al. MicroRNAs in cardiovascular disease. Hell. J. Cardiol. 2020, 61, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Doench, J.G.; Petersen, C.P.; Sharp, P.A. siRNAs can function as miRNAs. Genes Dev. 2003, 17, 438–442. [Google Scholar] [CrossRef]
- Paul, S.; Ruiz-Manriquez, L.M.; Ledesma-Pacheco, S.J.; Benavides-Aguilar, J.A.; Torres-Copado, A.; Morales-Rodríguez, J.I.; De Donato, M.; Srivastava, A. Roles of microRNAs in chronic pediatric diseases and their use as potential biomarkers: A review. Arch. Biochem. Biophys. 2021, 699, 108763. [Google Scholar] [CrossRef]
- Song, Y.; Higgins, H.; Guo, J.; Harrison, K.; Schultz, E.N.; Hales, B.J.; Moses, E.K.; Goldblatt, J.; Pachter, N.; Zhang, G. Clinical significance of circulating microRNAs as markers in detecting and predicting congenital heart defects in children. J. Transl. Med. 2018, 16, 1–11. [Google Scholar] [CrossRef]
- Vegter, E.L.; van der Meer, P.; de Windt, L.J.; Pinto, Y.M.; Voors, A.A. MicroRNAs in heart failure: From biomarker to target for therapy. Eur. J. Heart Fail. 2016, 18, 457–468. [Google Scholar] [CrossRef]
- Romaine, S.P.; Tomaszewski, M.; Condorelli, G.; Samani, N.J. MicroRNAs in cardiovascular disease: An introduction for clinicians. Heart 2015, 101, 921–928. [Google Scholar] [CrossRef]
- Ren, J.; Zhang, J.; Xu, N.; Han, G.; Geng, Q.; Song, J.; Li, S.; Zhao, J.; Chen, H. Signature of circulating microRNAs as potential biomarkers in vulnerable coronary artery disease. PLoS ONE 2013, 8, e80738. [Google Scholar] [CrossRef]
- Ram, T.P.; Fomison-Nurse, I.; Gandhi, S.; Coffey, S.; Saxena, P.; Galvin, I.; Bunton, R.; Williams, M.J.A.; Lamberts, R.R.; Katare, R. The diagnostic sensitivity of circulating cardio-enriched microRNAs is increased after normalization of high-density lipoprotein levels. Int. J. Cardiol. 2017, 236, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, J.; Sun, D.; Wei, N. MiR-155 inhibits transformation of macrophages into foam cells via regulating CEH expression. Biomed. Pharm. 2018, 104, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Nader, J.; Metzinger, L.; Maitrias, P.; Caus, T.; Metzinger-Le Meuth, V. Aortic valve calcification in the era of non-coding RNAs: The revolution to come in aortic stenosis management? Noncoding RNA Res. 2020, 5, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.J. Macrophages in atherosclerosis regression. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 20–33. [Google Scholar] [CrossRef]
- Severino, P.; D’Amato, A.; Pucci, M.; Infusino, F.; Adamo, F.; Birtolo, L.I.; Netti, L.; Montefusco, G.; Chimenti, C.; Lavalle, C. Ischemic heart disease pathophysiology paradigms overview: From plaque activation to microvascular dysfunction. Int. J. Mol. Sci. 2020, 21, 8118. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Basatemur, G.L.; Jørgensen, H.F.; Clarke, M.C.; Bennett, M.R.; Mallat, Z. Vascular smooth muscle cells in atherosclerosis. Nat. Rev. Cardiol. 2019, 16, 727–744. [Google Scholar] [CrossRef]
- Li, Y.; Ren, M.; Wang, X.; Cui, X.; Zhao, H.; Zhao, C.; Zhou, J.; Guo, Y.; Hu, Y.; Yan, C. Glutaredoxin 1 mediates the protective effect of steady laminar flow on endothelial cells against oxidative stress-induced apoptosis via inhibiting Bim. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Kunadian, V.; Chieffo, A.; Camici, P.G.; Berry, C.; Escaned, J.; Maas, A.H.; Prescott, E.; Karam, N.; Appelman, Y.; Fraccaro, C. An EAPCI expert consensus document on ischaemia with non-obstructive coronary arteries in collaboration with European Society of Cardiology Working Group on Coronary Pathophysiology & Microcirculation Endorsed by Coronary Vasomotor Disorders International Study Group. Eur. Heart J. 2020, 41, 3504–3520. [Google Scholar]
- Ford, T.J.; Rocchiccioli, P.; Good, R.; McEntegart, M.; Eteiba, H.; Watkins, S.; Shaukat, A.; Lindsay, M.; Robertson, K.; Hood, S. Systemic microvascular dysfunction in microvascular and vasospastic angina. Eur. Heart J. 2018, 39, 4086–4097. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, X. Effect of microRNA-370 on coronary atherosclerosis and its underlying mechanism. Exp. Ther. Med. 2019, 17, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Sun, H.; Sun, B. MicroRNA-145 is involved in endothelial cell dysfunction and acts as a promising biomarker of acute coronary syndrome. Eur. J. Med. Res. 2020, 25, 1–10. [Google Scholar] [CrossRef]
- Fichtlscherer, S.; De Rosa, S.; Fox, H.; Schwietz, T.; Fischer, A.; Liebetrau, C.; Weber, M.; Hamm, C.W.; Röxe, T.; Müller-Ardogan, M. Circulating microRNAs in patients with coronary artery disease. Circ. Res. 2010, 107, 677–684. [Google Scholar] [CrossRef]
- Gholipour, A.; Shakerian, F.; Zahedmehr, A.; Irani, S.; Mowla, S.J.; Malakootian, M. Downregulation of Talin-1 is associated with the increased expression of miR-182-5p and miR-9-5p in coronary artery disease. J. Clin. Lab. Anal. 2022, 36, e24252. [Google Scholar] [CrossRef]
- Lin, J.; Jiang, J.; Zhou, R.; Li, X.; Ye, J. MicroRNA-451b participates in coronary heart disease by targeting VEGFA. Open Med. 2020, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dabravolski, S.A.; Khotina, V.A.; Omelchenko, A.V.; Kalmykov, V.A.; Orekhov, A.N. The Role of the VEGF Family in Atherosclerosis Development and Its Potential as Treatment Targets. Int. J. Mol. Sci. 2022, 23, 931. [Google Scholar] [CrossRef]
- Chen, P.B.; Black, A.S.; Sobel, A.L.; Zhao, Y.; Mukherjee, P.; Molparia, B.; Moore, N.E.; Aleman Muench, G.R.; Wu, J.; Chen, W.; et al. Directed remodeling of the mouse gut microbiome inhibits the development of atherosclerosis. Nat. Biotechnol. 2020, 38, 1288–1297. [Google Scholar] [CrossRef]
- Severino, P.; D’Amato, A.; Prosperi, S.; Fanisio, F.; Birtolo, L.I.; Costi, B.; Netti, L.; Chimenti, C.; Lavalle, C.; Maestrini, V.; et al. Myocardial Tissue Characterization in Heart Failure with Preserved Ejection Fraction: From Histopathology and Cardiac Magnetic Resonance Findings to Therapeutic Targets. Int. J. Mol. Sci. 2021, 22, 7650. [Google Scholar] [CrossRef]
- Bruen, R.; Fitzsimons, S.; Belton, O. miR-155 in the Resolution of Atherosclerosis. Front. Pharm. 2019, 10, 463. [Google Scholar] [CrossRef]
- Yang, S.; Ye, Z.M.; Chen, S.; Luo, X.Y.; Chen, S.L.; Mao, L.; Li, Y.; Jin, H.; Yu, C.; Xiang, F.X.; et al. MicroRNA-23a-5p promotes atherosclerotic plaque progression and vulnerability by repressing ATP-binding cassette transporter A1/G1 in macrophages. J Mol. Cell. Cardiol. 2018, 123, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Yang, B.; Yang, H.; Wang, L.; Li, H.; Chen, S.; Lu, X.; Gu, D. MicroRNA-320b Modulates Cholesterol Efflux and Atherosclerosis. J. Atheroscler. Thromb. 2022, 29, 200–220. [Google Scholar] [CrossRef] [PubMed]
- Halimulati, M.; Duman, B.; Nijiati, J.; Aizezi, A. Long noncoding RNA TCONS_00024652 regulates vascular endothelial cell proliferation and angiogenesis via microRNA-21. Exp. Med. 2018, 16, 3309–3316. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Feng, J.; Ma, Y.; Cai, D.; Luo, Y.; Wang, Q.; Li, F.; Zhang, M.; Hu, Q. Down-regulation of microRNA-342-5p or up-Regulation of Wnt3a Inhibits Angiogenesis and Maintains Atherosclerotic Plaque Stability in Atherosclerosis Mice. Nanoscale Res. Lett. 2021, 16, 165. [Google Scholar] [CrossRef]
- Chen, L.Y.; Wang, X.; Qu, X.L.; Pan, L.N.; Wang, Z.Y.; Lu, Y.H.; Hu, H.Y. Activation of the STAT3/microRNA-21 pathway participates in angiotensin II-induced angiogenesis. J. Cell. Physiol. 2019, 234, 19640–19654. [Google Scholar] [CrossRef]
- Woo, C.C.; Liu, W.; Lin, X.Y.; Dorajoo, R.; Lee, K.W.; Richards, A.M.; Lee, C.N.; Wongsurawat, T.; Nookaew, I.; Sorokin, V. The interaction between 30b-5p miRNA and MBNL1 mRNA is involved in vascular smooth muscle cell differentiation in patients with coronary atherosclerosis. Int. J. Mol. Sci. 2019, 21, 11. [Google Scholar] [CrossRef]
- Lai, Z.; Lin, P.; Weng, X.; Su, J.; Chen, Y.; He, Y.; Wu, G.; Wang, J.; Yu, Y.; Zhang, L. MicroRNA-574-5p promotes cell growth of vascular smooth muscle cells in the progression of coronary artery disease. Biomed. Pharm. 2018, 97, 162–167. [Google Scholar] [CrossRef]
- Solly, E.L.; Dimasi, C.G.; Bursill, C.A.; Psaltis, P.J.; Tan, J.T.M. MicroRNAs as Therapeutic Targets and Clinical Biomarkers in Atherosclerosis. J. Clin. Med. 2019, 8, 2199. [Google Scholar] [CrossRef]
- He, W.; Zhu, L.; Huang, Y.; Zhang, Y.; Shen, W.; Fang, L.; Li, J.; Wang, Z.; Xie, Q. The relationship of MicroRNA-21 and plaque stability in acute coronary syndrome. Med. (Baltim.) 2019, 98, e18049. [Google Scholar] [CrossRef]
- Yu, F.; Di, M.; Li, M.; Chen, Y.; Zhang, Y.; Liu, X.; Huang, X.; Zhang, M. MicroRNA-124-3p inhibits collagen synthesis in atherosclerotic plaques by targeting prolyl 4-hydroxylase subunit alpha-1 (P4HA1) in vascular smooth muscle cells. Atherosclerosis 2018, 277, 98–107. [Google Scholar]
- Caroli, A.; Cardillo, M.T.; Galea, R.; Biasucci, L.M. Potential therapeutic role of microRNAs in ischemic heart disease. J. Cardiol. 2013, 61, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Taraldsen, M.D.; Wiseth, R.; Videm, V.; Bye, A.; Madssen, E. Associations between circulating microRNAs and coronary plaque characteristics: Potential impact from physical exercise. Physiol. Genom. 2022, 54, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.J.; Biessen, E.A.; Hohl, M.; Weber, C.; Van der Vorst, E.P.; Santovito, D. Small things matter: Relevance of MicroRNAs in cardiovascular disease. Front. Physiol. 2020, 11, 793. [Google Scholar] [CrossRef]
- Broderick, J.A.; Zamore, P.D. MicroRNA therapeutics. Gene Ther. 2011, 18, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Hu, T.; Du, Q.; Huang, W.; Yao, C. Nanoparticle-Delivered microRNA-153-3p Alleviates Myocardial Infarction-Induced Myocardial Injury in a Rat Model. ACS Biomater. Sci. Eng. 2022, 8, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xue, R.; Huang, P.; Wu, Y.; Fan, W.; He, X.; Dong, Y.; Liu, C. Modified Exosomes: A Good Transporter for miRNAs within Stem Cells to Treat Ischemic Heart Disease. J. Cardiovasc. Transl. Res. 2022, 15, 1–10. [Google Scholar] [CrossRef]
- Gupta, S.K.; Foinquinos, A.; Thum, S.; Remke, J.; Zimmer, K.; Bauters, C.; de Groote, P.; Boon, R.A.; de Windt, L.J.; Preissl, S. Preclinical development of a microRNA-based therapy for elderly patients with myocardial infarction. J. Am. Coll. Cardiol. 2016, 68, 1557–1571. [Google Scholar] [CrossRef]
- Li, Q.; Xie, J.; Li, R.; Shi, J.; Sun, J.; Gu, R.; Ding, L.; Wang, L.; Xu, B. Overexpression of micro RNA-99a attenuates heart remodelling and improves cardiac performance after myocardial infarction. J. Cell. Mol. Med. 2014, 18, 919–928. [Google Scholar] [CrossRef]
- Qin, Y.; Yu, Y.; Dong, H.; Bian, X.; Guo, X.; Dong, S. MicroRNA 21 inhibits left ventricular remodeling in the early phase of rat model with ischemia-reperfusion injury by suppressing cell apoptosis. Int. J. Med. Sci. 2012, 9, 413. [Google Scholar] [CrossRef]
- Yang, X.; Qin, Y.; Shao, S.; Yu, Y.; Zhang, C.; Dong, H.; Lv, G.; Dong, S. MicroRNA-214 inhibits left ventricular remodeling in an acute myocardial infarction rat model by suppressing cellular apoptosis via the phosphatase and tensin homolog (PTEN). Int. Heart J. 2016, 57, 247–250. [Google Scholar] [CrossRef]
- Song, C.-L.; Liu, B.; Diao, H.-Y.; Shi, Y.-F.; Li, Y.-X.; Zhang, J.-C.; Lu, Y.; Wang, G.; Liu, J.; Yu, Y.-P. The protective effect of microRNA-320 on left ventricular remodeling after myocardial ischemia-reperfusion injury in the rat model. Int. J. Mol. Sci. 2014, 15, 17442–17456. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, B.C.; Nguyen, S.S.; Winbanks, C.E.; Gao, X.M.; Boey, E.J.; Tham, Y.K.; Kiriazis, H.; Ooi, J.Y.; Porrello, E.R.; Igoor, S. Therapeutic silencing of miR-652 restores heart function and attenuates adverse remodeling in a setting of established pathological hypertrophy. FASEB J. 2014, 28, 5097–5110. [Google Scholar] [CrossRef] [PubMed]
- Bellera, N.; Barba, I.; Rodriguez-Sinovas, A.; Ferret, E.; Asín, M.A.; Gonzalez-Alujas, M.T.; Pérez-Rodon, J.; Esteves, M.; Fonseca, C.; Toran, N. Single intracoronary injection of encapsulated antagomir-92a promotes angiogenesis and prevents adverse infarct remodeling. J. Am. Heart Assoc. 2014, 3, e000946. [Google Scholar] [CrossRef] [PubMed]
- Hullinger, T.G.; Montgomery, R.L.; Seto, A.G.; Dickinson, B.A.; Semus, H.M.; Lynch, J.M.; Dalby, C.M.; Robinson, K.; Stack, C.; Latimer, P.A. Inhibition of miR-15 protects against cardiac ischemic injury. Circ. Res. 2012, 110, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-s.; Jin, J.-p.; Wang, J.-q.; Zhang, Z.-g.; Freedman, J.H.; Zheng, Y.; Cai, L. miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta Pharmacol. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Rawlings-Goss, R.A.; Campbell, M.C.; Tishkoff, S.A. Global population-specific variation in miRNA associated with cancer risk and clinical biomarkers. BMC Med. Genom. 2014, 7, 53. [Google Scholar] [CrossRef]
- Moreno, J.A.; Hamza, E.; Guerrero-Hue, M.; Rayego-Mateos, S.; García-Caballero, C.; Vallejo-Mudarra, M.; Metzinger, L.; Metzinger-Le Meuth, V. Non-Coding RNAs in Kidney Diseases: The Long and Short of Them. Int. J. Mol. Sci. 2021, 22, 6077. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Steps of Atherosclerosis | miRNA | Regulation | Model | Target Genes | Specific Roles of miRNA | Reference |
|---|---|---|---|---|---|---|
| Endothelial cells regulation and inflammatory response | miR-370 | up | Human peripheral blood mononuclear cells Cell line: HUVEC | FOXO1 | - Promotes endothelial cell migration and invasion - Inhibits apoptosis | [32] |
| miR-145 | down | Human serum Sprague –Dawley mice | FOXO1 | Increases the concentration of endothelial injury biomarkers and the inflammatory cytokines | [33] | |
| miR-182-5pmiR-9-5p | up | Human serum Cell line: HEK293T | TLN1 | - Interferes the ECM constituents | [35] | |
| miR-451b | up | Human peripheral blood Cell line: HUVEC | VEGF-A | - Inhibits cell proliferation - Promotes cell apoptosis | [36] | |
| miR-381 | down | Human plasma Cell line: HUVEC | CXCR4 | - Promotes release of inflammatory cytokines - Reduces cell proliferation - Promotes early apoptosis | [38] | |
| Activation of monocyte and differentiation of macrophage | miR-155 | up | Human THP-1 cells | CEH ABCA1 SR-A | - Increases cholesterol efflux in macrophages - Decreases intracellular lipid accumulation - Inhibits foam cells formation | [22] |
| miR-23a-5p | up | Human peripheral blood plasma Apoe−/− mice Raw264.7 cells and HEK293T cells | ABCA1 ABCG1 | - Decreases cholesterol efflux in macrophages - Promotes foam cells formation | [41] | |
| miR-320b | up | Human peripheral blood mononuclear cells Apoe−/− mice Human THP-1 cells, Raw264.7 cells, HEK293T, HUVEC | ABCG1 EEPD1 | - Reduces cholesterol efflux rate in macrophages - Promotes inflammatory response - Impairs lipid profile - Promotes atherosclerotic plaque formation | [13] | |
| Plaque angiogenesis | miR-342-5p | up | Apoe−/− mice and C57BL/6J mice | WNT3A | - Interferes the micro vessel density level | [44] |
| miR-21 | N/A | Cell line: HMECs | PTEN | - Increases endothelial cell proliferation and migration - Increases angiogenesis | [45] | |
| VSMC proliferation and differentiation | hsa-miR-30b-5p | up | Human aortic wall tissue VSMC cell lines | MBNL1 | Reduces the common markers level for VSMC differentiation | [46] |
| miR-574-5p | up | Human serum Cell culture | ZDHHC14 | - Promotes VSMC proliferation - Inhibits cellular apoptosis | [47] | |
| Fibrous cap destabilisation and plaque rupture | miR-21 | N/A | Human blood samples | PTEN MMP-2 | - Reduces fibrous cap thickness - Increases necrotic core area - Increases the macrophages surrounding fibrous cap - Promotes plaque instability | [49] |
| miR-124-3p | up | Apoe−/− mice Human aortic smooth muscle cell lines | P4HA1 | - Inhibits collagen synthesis of VSMCs - Reduces plaque stability | [45] |
| Model | Intervention | Outcomes | Reference |
|---|---|---|---|
| HL-1 cell line transfected with microRNA | administration of miR-22 anti-miRs | activates cardiac autophagy to prevent post-infarction remodeling and improve cardiac function | [57] |
| Murine model | Lentivirus-mediated miR-99a delivery | improved survival rate and cardiac function | [58] |
| Rat model | adenovirus -delivered miR-214 or miR-21 improved | improved LV remodeling and decreased myocardial apoptosis | [59,60] |
| Rat model | administration of miR-320 anti-miRs | reduced the degree of myocardial fibrosis and apoptosis in LV remodeling | [61] |
| Rat model | inhibition of miR-132 | rescues cardiac hypertrophy and heart failure | [62] |
| Mouse model | administration of a locked nucleic acid anti-miR-652 | attenuation of cardiac hypertrophy. Improved heart function was associated with reduced cardiac fibrosis | [62] |
| Porcine model | single intracoronary administration of encapsulated anti-miR-92a | prevented left-ventricular remodeling | [63] |
| Porcine model | administration of locked nucleic acid modified anti-miR-15 | cardiomyocytes showed resistant to hypoxia-induced cardiomyocyte cell death | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, A.S.-Y.; Lai, K.-S.; Lim, S.-H.E.; Sivalingam, S.; Loh, J.-Y.; Maran, S. miRNA in Ischemic Heart Disease and Its Potential as Biomarkers: A Comprehensive Review. Int. J. Mol. Sci. 2022, 23, 9001. https://doi.org/10.3390/ijms23169001
Kong AS-Y, Lai K-S, Lim S-HE, Sivalingam S, Loh J-Y, Maran S. miRNA in Ischemic Heart Disease and Its Potential as Biomarkers: A Comprehensive Review. International Journal of Molecular Sciences. 2022; 23(16):9001. https://doi.org/10.3390/ijms23169001
Chicago/Turabian StyleKong, Amanda Shen-Yee, Kok-Song Lai, Swee-Hua Erin Lim, Sivakumar Sivalingam, Jiun-Yan Loh, and Sathiya Maran. 2022. "miRNA in Ischemic Heart Disease and Its Potential as Biomarkers: A Comprehensive Review" International Journal of Molecular Sciences 23, no. 16: 9001. https://doi.org/10.3390/ijms23169001
APA StyleKong, A. S.-Y., Lai, K.-S., Lim, S.-H. E., Sivalingam, S., Loh, J.-Y., & Maran, S. (2022). miRNA in Ischemic Heart Disease and Its Potential as Biomarkers: A Comprehensive Review. International Journal of Molecular Sciences, 23(16), 9001. https://doi.org/10.3390/ijms23169001