1. Introduction
Parkinson’s disease (PD) is defined by α-synuclein aggregation and toxicity in ventral midbrain dopaminergic neurons, which is clinically diagnosed with characteristic motor symptoms, including bradykinesia, resting tremor, muscle rigidity, and postural instability [
1,
2]. Phosphorylated and misfolded α-synuclein is a major constituent of Lewy body inclusions and Lewy neurites in PD and related brain diseases such as α-synucleinopathy [
3]. Approximately 80% of PD patients progressively manifest cognitive deficits over 15 years since the diagnosis of motor dysfunction, which is associated with the pathological propagation of Lewy pathologies from the midbrain towards the limbic and cortical brain areas [
4]. Despite the extensive pathological involvement of diverse brain regions in PD patients, currently available therapies focus on mitigating overt symptoms mainly by supplementing the responsible neurotransmitters or augmenting specific receptor-regulated signaling pathways. However, these anti-symptomatic therapies fail to fundamentally target disease progression, which is thought to be mediated by α-synuclein pathology formation and propagation. For better and effective treatment of motor and diverse non-motor clinical symptoms in patients with PD, it is crucial to suppress α-synuclein aggregation or modulate signaling pathways that contribute to α-synuclein aggregation or propagation in PD environments.
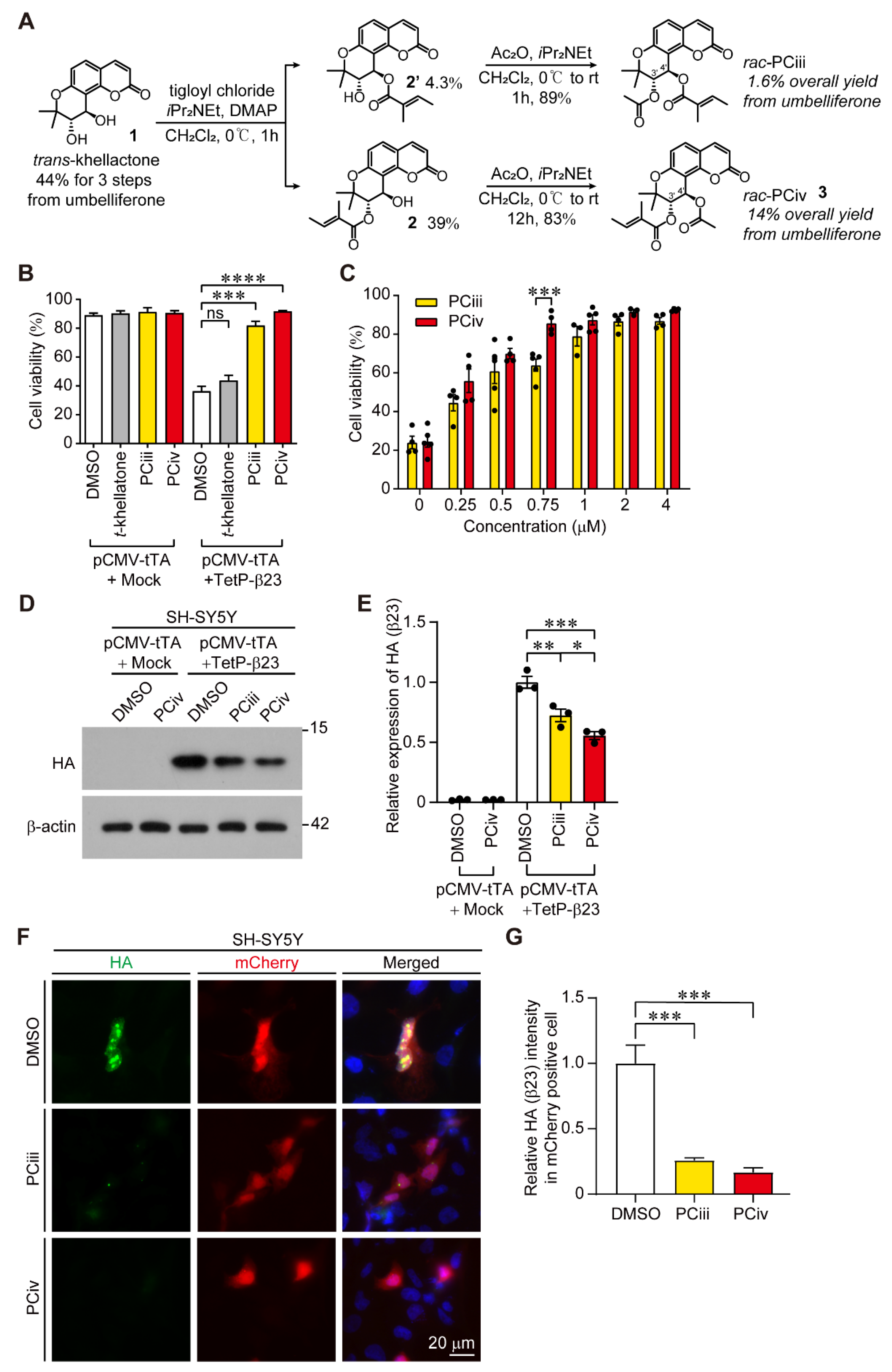
Current approaches to targeting pathological α-synuclein include anti-α-synuclein antibody therapies and small compounds with the ability to inhibit α-synuclein aggregation. Current clinical trials of anti-α-synuclein therapies may hold promise in specifically blocking the propagation of α-synuclein aggregation or receptor-mediated activation of microglia directly by extracellular α-synuclein aggregates. However, the low brain permeability of antibodies and limited therapeutic effect on intracellular α-synuclein aggregation reflects the need to develop small compounds with favorable brain permeability profiles that inhibit intracellular α-synuclein aggregation. In this regard, our group reported that peucedanocoumarin III (PCiii) exhibits strong inhibitory activity against repeated β sheet aggregate mimic β23 induced toxicity [
5,
6]. β23 was previously invented to represent the general β sheet aggregate structure of many brain disease proteins, including amyloid beta, α-synuclein, and tau [
7]. Both natural and synthetic PCiii compounds disaggregated and facilitated the proteasomal clearance of β23. Similarly, α-synuclein aggregation and neurotoxicity have been effectively prevented in a 6-OHDA-induced PD mouse model [
6]. Although the PCiii organic synthesis protocol has been well established, and this compound possesses promising anti-aggregate therapeutic activity in neurodegenerative diseases, further studies on PCiii derivatives with equivalent or advanced pharmacological properties have not yet been conducted.
In this study, we found that the structural isomer of PCiii (named “PCiv”) retained anti-aggregate functions against general β sheet aggregate β23 and α-synuclein aggregates. PCiv had a higher synthetic yield and better brain penetration compared with PCiii. Importantly, PCiv was shown to be effective in treating motor deficits and dopaminergic α-synucleinopathy pathologies in a sporadic PD mouse model of combinatorial recombinant adeno-associated virus expressing α-synuclein (rAAV-αSyn) and α-synuclein preformed fibril (PFF) injection, indicating its translational value.
3. Discussion
Here, we are the first to report the therapeutic biological function of PCiv in preventing α-synuclein aggregation and neurotoxicity in an animal model of PD. In our previous report characterizing PCiii, which was initially screened as a natural anti-amyloid compound [
5], PCiv was observed as merely a byproduct of higher synthetic yield without further evaluation of its potential biological activity. PCiv and PCiii are structural isomers differing in the positions of acetyl or tigloyl conjugation at the 3′- or 4′-carbon residues of
trans-khellactone. Attachment of the tigloyl moiety at the 3′-carbon of
trans-khellactone seems to provide slightly better cytoprotection against amyloid-induced cytotoxicity and a greater protective effect on PFF-induced α-synuclein aggregation in neurons. PCiv has never been studied for its potential inhibitory effect against α-synuclein aggregation, although there are some reports on the biological activity of the peucedanocoumarin analog modulating platelet aggregation [
14]. The comparable inhibitory effect of both PCiii and PCiv on recombinant α-synuclein protein aggregation in vitro suggests that the arrangement of acetyl or tigloyl moieties on
trans-khellactone is not critical for α-synuclein interaction and inhibition of its aggregation. Although the actual binding mode of PCiii or PCiv to α-synuclein aggregates is unclear, this result may suggest the potential presence of multiple binding pockets on α-synuclein aggregate structures or that there is some flexibility on the 3′- and 4′-C side of peucedanocoumarins for α-synuclein interaction. However, it is evident that acetyl and tigloyl conjugation is critically important for the biological activity of PCiii and PCiv since
trans-khellatone had no effect on cytoprotection against β23-induced cellular toxicity. It would be instructive to synthesize a more extensive repertoire of structural derivatives of PCiii or PCiv with different moieties conjugated to 3′ and 4′-C and evaluate their anti-protein aggregate biological activity. Moreover, it requires further investigation to understand which stereoisomer of racemic PCiii or PCiv mixture exerted actual biological effect on α-synuclein aggregation process. It would also be informative to narrow down which domain of α-synuclein aggregates PCiii and PCiv physically interacts during the disaggregation process. Hydrogen deuterium exchange mass spectrometry may be an excellent approach for investigating this issue in the future.
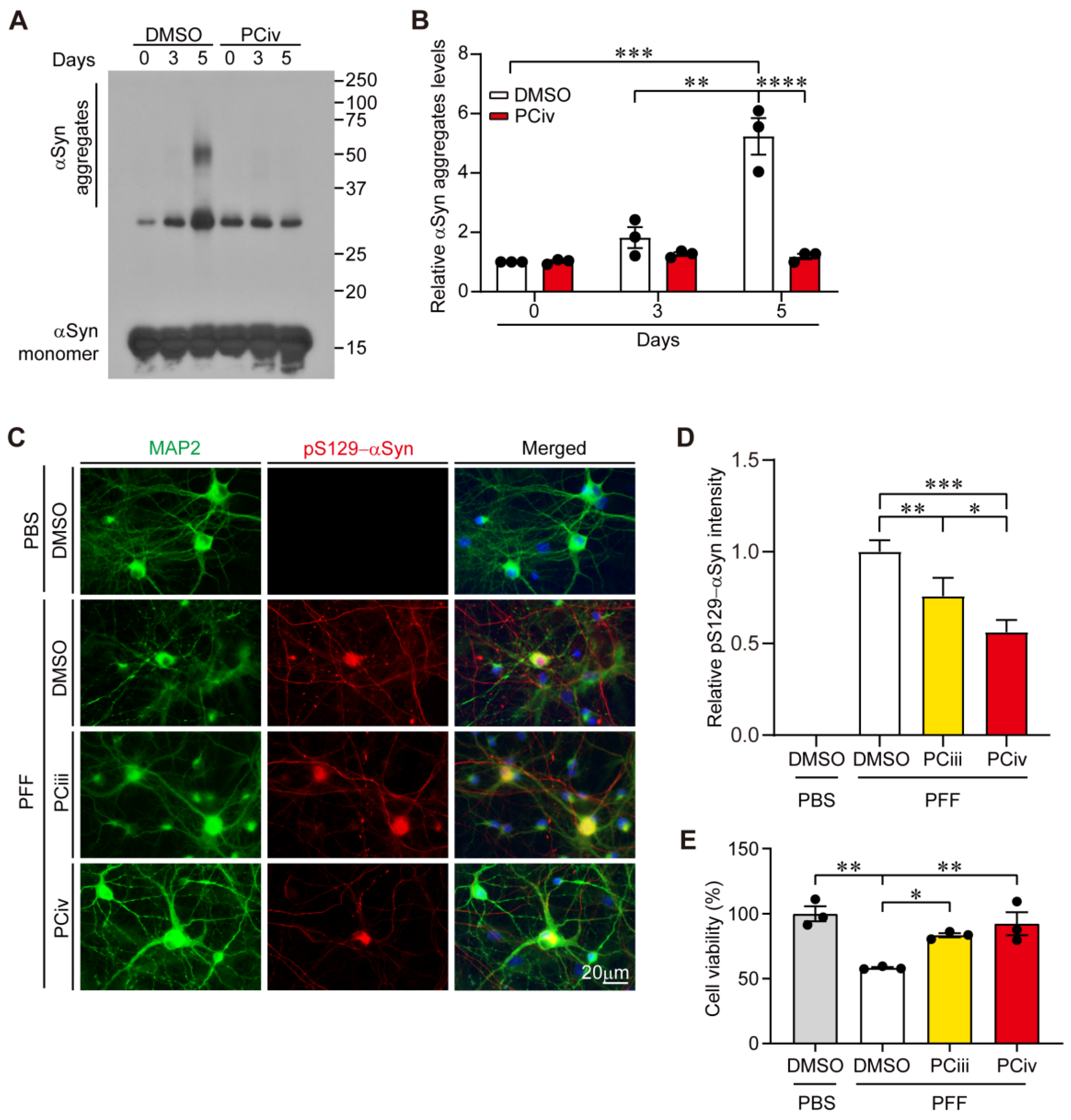
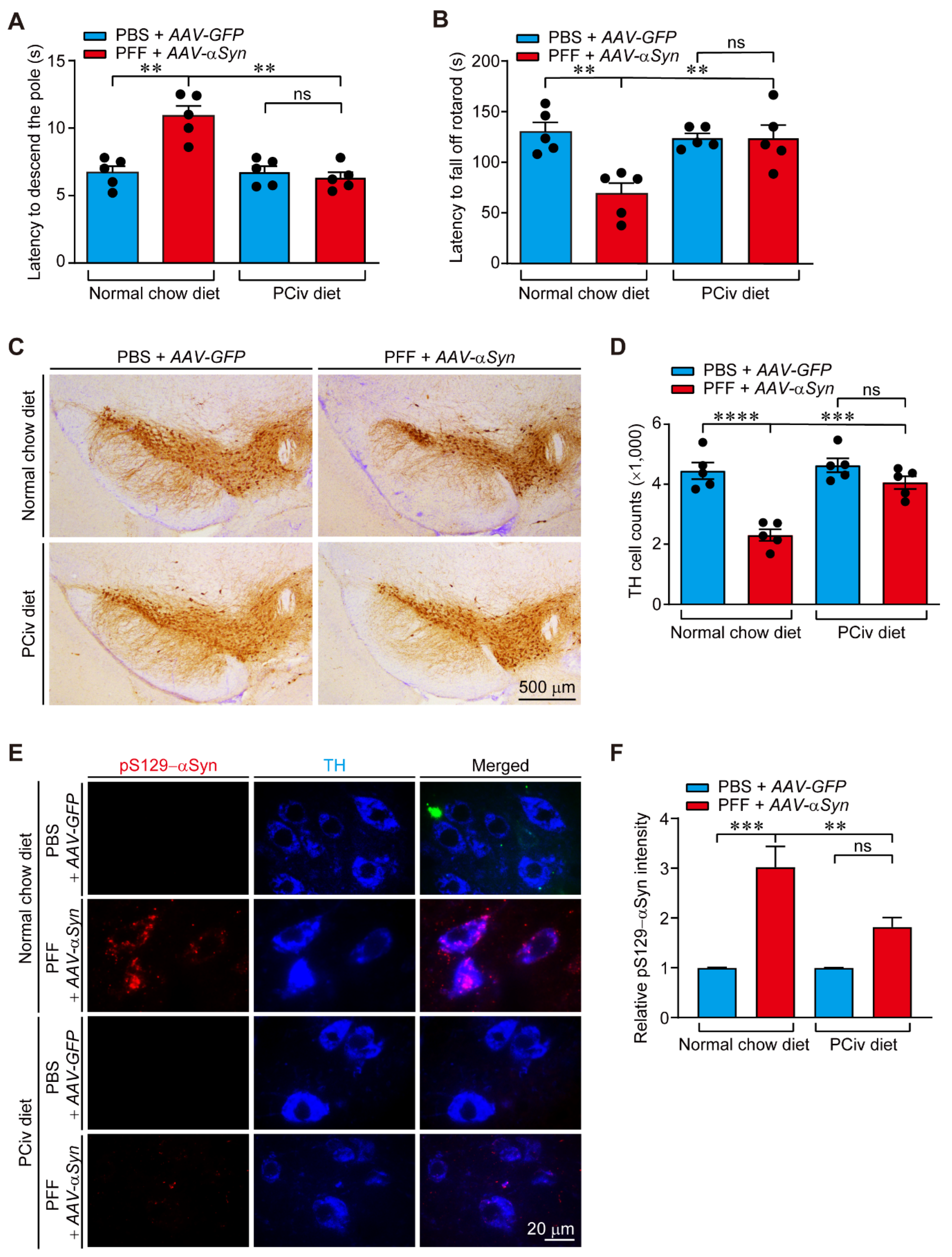
In this study, the therapeutic efficacy of PCiv was evaluated in sporadic PD mouse models. Although PCiii was reportedly neuroprotective in a 6-OHDA-induced PD mouse model [
6], PCiv showed more potent prevention of PFF-induced α-synuclein aggregation in mice cortical neurons (
Figure 2C), prompting us to further examine its therapeutic potential in a PD mouse model of α-synucleinopathy. With favorable brain penetration of PCiv in vivo, PCiv treatment prevented α-synuclein aggregation and dopaminergic neurodegeneration in a PD mouse model with a combinatorial PFF and
AAV-αSyn nigral injection, ultimately contributing to the behavioral rescue of motor functions. α-Synuclein aggregation pathologies are characterized by progressive impairment of brain function mediated by various brain subregions. According to Braak’s theory, which was supported by experimental data [
4,
9,
15], α-synuclein aggregation can be initiated in the intestinal peripheral nervous system or olfactory nerve; this Lewy pathology can then propagate to higher brain structures in the central nervous system through retrograde transmission. Indeed, targeting α-synuclein aggregation could be beneficial for terminating Lewy pathology propagation at any Braak stages. Furthermore, PCiv-mediated prevention of α-synuclein aggregation can preserve motor and non-motor functions mediated by specific brain subregions, even in environments with Lewy pathology infiltration. PCiv may be further evaluated in other α-synuclein transgenic mouse models, affecting multiple brain regions, to expand its therapeutic application to more diverse neurological disorders. Interestingly, the PCiv analog was reported as a potential probe in a cell model of huntingtin aggregation according to the PubChem Bioassay (CHEMBL1378488). PCiii was shown to be effective as an anti-aggregate compound for both α-synuclein and huntingtin proteins. Similarly, PCiv likely retains anti-aggregate functions that are applicable to a broader spectrum of disease-associated protein aggregation. Amyloid-mimic β23 clearance facilitated by PCiv supports this notion.
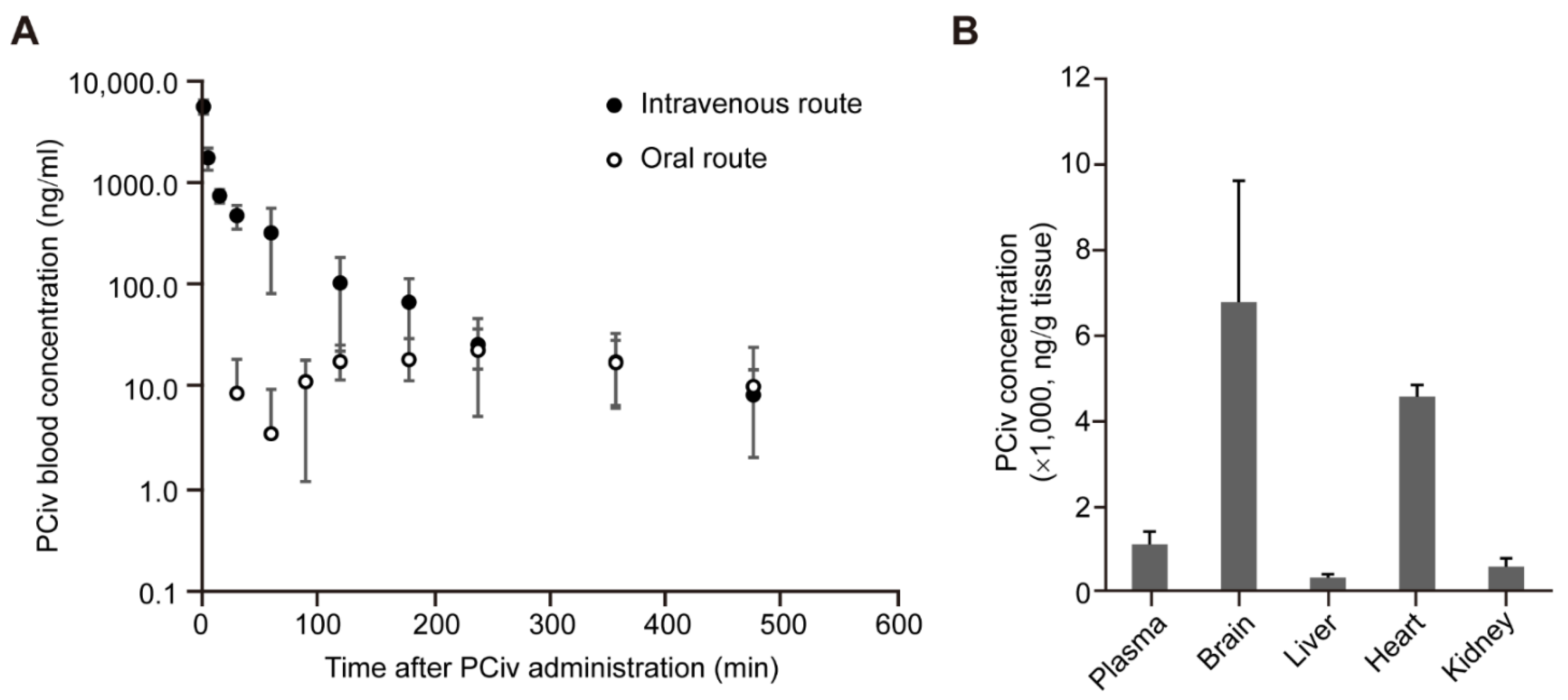
The advantages of PCiv compared to PCiii include a high yield in organic synthesis and a more favorable pharmacokinetic profile. Previously, we failed to analyze the detailed pharmacokinetic profiles of PCiii likely due to the poor pharmacokinetic properties, although it demonstrated brain distribution following intraperitoneal administration in mice [
6]. In contrast, PCiv was well-dissolved in the prepared vehicle solution (DMSO:PEG400:saline ratio of 5:70:25), and we were able to obtain pharmacokinetic profiles of PCiv following oral administration in rats. Although PCiv has approximately a 1.5-h half-life and relatively low bioavailability of 10%, its efficient distribution to the target organs (6.4-fold compared with the plasma concentration) poses strong advantages for PCiv in clinical applications for PD with oral administration. The pharmaceutical preparation of PCiv to enhance its oral bioavailability and absorption may be implemented for lead compound optimization. Moreover, toxicity evaluation of PCiv is critically important prior to the clinical evaluation of this drug. Although we did not observe any overt toxicity in cells and in vivo mouse models, prolonged and high-dose PCiv treatment in mice and characterization of behavior and organ pathology are required to ensure the safety of the drug, especially when PD treatment is likely to be prolonged because of the chronic nature of the disease.
4. Materials and Methods
4.1. Chemicals and Antibodies
Goat serum (#01-6201) and doxycycline (#D9891) were purchased from Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA and Sigma-Aldrich, St. Louis, MO, USA, respectively. The following primary antibodies were used: mouse antibody to HA (#PA1-985, 1:5000, Roche, Basel, Switzerland), rabbit antibody to filament α-synuclein (#ab209538, 1:5000, Abcam, Cambridge, UK), rabbit antibody to MAP2 (#ab32454, 1:1000, Abcam, Cambridge, UK), mouse antibody to α-synuclein (#610787, 1:3000, BD transduction laboratories, San Jose, CA, USA), mouse antibody to pS129-α-synuclein (#825702, 1:3000, Biolegend, San Diego, CA, USA), and rabbit antibody to TH (#NB300-109, 1:2000, Novus Biologicals, Centennial, CO, USA). The following secondary antibodies were used: horseradish peroxidase (HRP)-conjugated goat antibody to mouse immunoglobulin (Ig)G (#Gtx-213111-01, 1:5000, Genetex, Irvine, CA, USA), HRP-conjugated goat antibody to rabbit IgG (#Gtx-213110-01, 1:5000, Genetex, Irvine, CA, USA), biotin-conjugated goat antibody to rabbit IgG (#BA-1000, 1:1000, Vector Laboratories, Burlingame, CA, USA), Alexa Fluor 488-conjugated goat antibody to mouse IgG (A21202, 1:1000; Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA), Alexa Fluor 568-conjugated donkey antibody to rabbit IgG (#A10042, 1:1000; Invitrogen), and HRP-conjugated mouse antibody to β-actin (cat# A3854, 1:10,000, Sigma-Aldrich, St. Louis, MO, USA).
4.2. Organic Synthesis of PCiv
The general procedure was performed as follows. Unless otherwise noted, all starting materials and reagents were obtained from commercial suppliers and used without further purification. All solvents used for the routine isolation of products and chromatography were of reagent grade and glass-distilled. Reaction flasks were dried at 100 °C. Air- and moisture-sensitive reactions were performed in an argon atmosphere. Chemical shifts are expressed in parts per million (δ) downfield from tetramethylsilane and referenced to deuterated solvent (CHCl3). 1H-NMR data were reported in the order of chemical shift, multiplicity (s, singlet; d, doublet; t, triplet; q, quartet; m, multiplet or multiple resonances), number of protons, and coupling constant in hertz (Hz).
To a solution of 2 (340 mg, 1.0 mmol) in CH2Cl2 (10 mL), iPr2NEt (0.35 mL, 2.0 mmol) and acetic anhydride (0.14 mL, 1.5 mmol) were added at 0 °C. The reaction mixture was then warmed to room temperature and stirred for 12 h. After aq.NH4Cl was added to the reaction mixture, it was transferred to a separatory funnel and extracted with CH2Cl2 twice. The organic layers were dried over MgSO4, filtered, and evaporated in vacuo. The residue was purified by column chromatography on silica gel (ethyl acetate: n-hexane = 1:2) to afford PCiv 3 (320 mg, 83%) as an amorphous solid.
PCiv 3: 1H-NMR (CDCl3, 500 MHz) δ 7.58 (1H, d, J = 15.6 HZ), 7.35 (1H, d, J = 14.5 Hz), 6.80 (1H, m,), 6.78 (1H, d, J = 14.5 Hz), 6.24 (1H, d, J = 7.5 Hz), 6.21 (1H, d, J = 15.6 Hz), 5.31 (1H, d, J = 8.0 Hz), 2.08 (3H, s), 1.78 (3H, t, J = 1.0 Hz), 1.74 (3H, dd, J = 12.0, 1.5 Hz), 1.41 (3H, s), 1.33 (3H, s); 13C-NMR (CDCl3, 125 MHz) δ 169.5, 166.3, 159.9, 156.5, 154.0, 143.3, 138.9, 129.1, 127.7, 114.4, 113.1, 112.5, 107.0, 71.9, 63.8, 24.2, 22.6, 20.7, 14.5, 12.1. LRMS (FAB) m/z 386 (M+); HRMS (FAB) Calcd for C21H22O7 (M+): 386.1366, Found 386.1381.
4.3. Cell Culture and Transfection
Human neuroblastoma SH-SY5Y cells (ATCC, Manassas, VA, USA) were grown in complete media containing DMEMs, 10% fetal bovine serum (vol/vol), and antibiotics (penicillin-streptomycin 100 U/mL, Thermo Fisher Scientific, Waltham, MA, USA). The atmospheric condition used to preserve the cells consisted of 5% CO
2/95% air at 37 °C and maintained a humidified environment. X-tremeGENE HP transfection reagents (Roche, Basel, Switzerland) were used for transient transfection of DNA constructs (pCMV-tTA, TetP-β23, and pTet-dual2 [
5]) following the manufacturer’s instructions.
4.4. Cell Viability Assay
SH-SY5Y cells were plated in 12-well plates at a density of 0.2 × 106 cells per well. Following transient transfection with the indicated constructs and subsequent treatment protocol, the cells were harvested by trypsinization, thereby yielding single-cell suspensions. The cells were washed twice with phosphate-buffered saline (PBS) and resuspended in serum-free DMEM. The resuspended cells were mixed with an equal volume of 0.4% trypan blue (w/v) and incubated for 2 min at room temperature. Live and dead cells were counted using a Countess II Automated Cell Counter (Life Technologies, Carlsbad, CA, USA) for assessing cell viability.
4.5. Fluorescence Imaging
The SH-SY5Y cells were fixed with 4% paraformaldehyde in PBS and blocked with a blocking solution (5% normal goat serum [Invitrogen], 0.1% Triton X-100 [Sigma-Aldrich, St. Louis, MO, USA]) in PBS, pH 7.4) for 1 h at room temperature and incubated with primary antibodies against HA-tagged β23 at 4 °C overnight. Then, the cells were washed with wash buffer (0.1% Triton x-100 in PBS, pH 7.4) and incubated with corresponding secondary antibodies conjugated with fluorescent dyes at room temperature for 1 h. After simple washing with wash buffer, the samples were incubated with DAPI (Sigma-Aldrich, St. Louis, MO, USA) solution. The cells were washed with DAPI solution in PBS and mounted on a microscope slide using Immu-Mount solution (Thermo Fisher Scientific, #9990402). Images were obtained using a fluorescent microscope (Axio Imager M2; Carl Zeiss, Oberkochen, Germany).
4.6. Western Blotting
Lysis buffer was added to the SH-SY5Y cells washed with ice-cold PBS (1% Nonidet P40 in PBS, pH 7.4) that was supplemented with protease/phosphatase inhibitors. To obtain total protein lysate, we performed freezing and thawing cycles three times using dry ice and ice water. The samples were then centrifuged at 14,000 g for 30 min. The supernatants were quantified through BCA protein assessment. The samples were mixed with 2X Laemmli buffer (Bio-Rad, Hercules, CA, USA) supplemented with β-mercaptoethanol (Sigma-Aldrich) and boiled for 5 min at 95 °C. The proteins were separated by SDS-PAGE and transferred to nitrocellulose membranes for immunoblotting. Western blotting was performed using specific antibodies and the immune response bands were visualized using chemiluminescence (Pierce, Thermo Fisher Scientific, Waltham, MA, USA). Next, band quantification was performed using ImageJ software. (National Institutes of Health, Bethesda, MD, USA).
4.7. α-Synuclein Aggregation Assessment and PFF Preparation
Plasmid pRK172-human α-synuclein was transformed into BL21-competent Escherichia coli using heat-shock transformation. Recombinant α-synuclein was induced and purified, as described above. Purified α-synuclein was incubated with or without PCiv in sodium acetate buffer (100 mM, pH 7.5) in a shaking incubator (300 rpm, Vision Scientific, Daejeon, Korea) at 37 °C. After the indicated days of incubation, fibril formation in the protein solution was assessed by Western blotting. α-Synuclein fibrils (5 mg/mL) generated from monomeric α-synuclein incubation for 7 days were processed by sonication (Vibra Cell, Sonics & Materials, Inc., Newtown, CT, USA) to prepare α-synuclein PFFs. To prevent the solution from overheating, 60 pulses with a 0.5 s duration of 10% of the power level were applied with a short pause after every 10 pulses.
4.8. Mouse Cortical Neuron Culture and PFF Treatment and Neurotoxicity Assessment
CD1 mice were purchased from Orient Co. for use in the mouse primary neuron culture. E14 embryos from CD1 mice were dissected for primary cortical neurons in Neurobasal A medium (Gibco, Thermo Fisher Scientific Inc., Waltham, MA, USA) supplemented with B-27 (Gibco, Thermo Fisher Scientific Inc., Waltham, MA, USA) and 1 mM L-glutamine. Next, the primary cortical neurons were maintained in a poly L-lysine-coated tissue culture plate, and the medium was changed every 2–3 days. After 7 days, PFF was added to the cortical neurons and incubated for 14 days to observe the p-S129-α-synuclein signal which was used for immunofluorescence analysis. For neurotoxicity assessment, mouse cortical neurons following 14 days of PFF treatment were incubated with Neurobasal A medium containing CCk-8 (cat# B1007-500, Cellrix, MediFab, Seoul, Korea) solutions for 1.5 h in 37 ℃. The optical value was read at 450 nm using a multi-detection microplate reader (Synergy, BioTek Instruments Inc., Winooski, VT, USA).
4.9. Animal Experiments
All animal experiments were approved by the Ethical Committee of Sungkyunkwan University Animal Care and Use Committee of Gachon University (Approval number: SKKUIACUC2019-06-15-1; GIACUC-R2021006) and were conducted in accordance with all applicable international guidelines. Male C57BL/6N mice (2 months old) were obtained from Orient Bio (Suwon, Korea). The animals were housed in air-controlled rooms with a 12-h dark/light cycle. Mice and rats were provided ad libitum access to diet and water. All efforts were made to minimize animal suffering and the number of animals used in the experiments. A synthetic PCiv diet was supplied to the mice. The PCiv diet administration (50 mg/kg/day for PFF PD mouse models) began on day 0 and was continued for 30 days. Intranigral injection of the rAAV-αSyn and PFF mixture was performed on day 0. Mice brains were prepared for analysis, as described below. To test the therapeutic effects of PCiv, we prepared a diet containing PCiv. To facilitate each mouse consuming 2 mg of PCiv per day, we generated a mixture of 400 mg of PCiv per kg of rodent diet and then supplied it to the mice.
4.10. Rat Pharmacokinetic and Tissue Distribution of PCiv
In vivo rat pharmacokinetic studies were performed for intravenous and oral administration of PCiv as previously reported [
16,
17,
18]. Sprague-Dawley rats (8 weeks old, 260–280 g) were purchased from Orient Bio Inc. (Seongnam, Korea). Rats were maintained with free access to food and water under a 12:12 h light/dark cycle for 1 week. For pharmacokinetic studies, under anesthetic conditions (20 mg/kg tiletamine/zolazepam [Zoletil; Vibrac Laboratories, Carros, France] administered intramuscularly), the femoral vein and artery were surgically cannulated using polyethylene tubing (PE50; Clay Adams, Parsippany, NJ, USA) in fasted rats. Then, PCiv was administered intravenously and orally at a dose of 10 mg/kg (n = 4–5). The drug was dissolved in a vehicle (DMSO:PEG400:saline at a ratio of 5:70:25). Blood samples (approximately 100 μL) were collected at predetermined time points (0, 1, 5 (intravenous administration only), 15, 30, 60, 120, 180, 240, 360, and 480 min) after drug administration. To prevent blood clots and fluid loss, the cannula to the femoral artery was flushed with 20 IU/mL heparinized saline, and the same volume of saline was pushed to the cannula for the femoral vein. The plasma fraction was immediately obtained after centrifugation at 14,000 rpm for 15 min at 4 °C and then stored at −20 °C prior to analysis using the ultra-high-performance liquid chromatography (UHPLC)-diode array detector (DAD) method. Similarly, a tissue distribution study was performed 15 min after the intravenous bolus injection (10 mg/kg), as previously described [
17,
18]. Four major tissues, including tissues from the brain, heart, kidneys, and liver, were collected. Based on the wet weights of the tissue samples, a two-fold volume of PBS was added and homogenized. The tissue homogenates were stored at −20 °C before UHPLC-DAD analysis. The UHPLC-DAD system consisted of an Agilent Technologies 1290 Infinity II equipped with a DAD (G7117A), an autosampler (G71678B), a binary pump (G7104A), and a multicolumn thermostat (G7116B). The PCiv level was determined using a CapcellpakTM C18 5 μm column (250 × 1.5 mm, Shiseido Fine Chemical Co., Tokyo, Japan), as previously described [
6]. The mobile phase was composed of 0.1% phosphoric acid and acetonitrile and eluted in the gradient mode at a flow rate of 0.2 mL/min. A DAD detector was applied at 321 nm, with an injection volume of 10 μL. The calibration range was 20–10,000 ng/mL with good linearity (R = 0.99783 with 1/× weighting). The tissue concentration levels were calculated as the PCiv amount (ng) per gram of organ weight.
4.11. Nigral Co-Injection of PFF and AAV-αSyn
The α-synucleinopathy PD model was created by co-injecting PFF (10 µg; ventral tegmental area: AP, −3.4 mm; ML, −0.5 mm; DV, −4.3 mm) and rAAV-αSyn (AAV serotype 1, 1 µL of titer 5 × 1011 GC/mL; substantia nigra pars compacta: AP, −3.4 mm; ML, −1.3 mm; DV, −4.3 mm) into a 2-month-old C57BL/6N mouse. rAAV-GFP was used as the control. PFF was prepared from pure recombinant human α-synuclein (Proteos, Inc., Kalamazoo, MI, USA) according to a previous report. Before use, sonicated PFFs were evaluated using Western blotting and a functional assay in cortical neuron cultures (pS129-αSyn induction and neurotoxicity). After co-injecting PFF and rAAV-αSyn, PD models were fed a PCiv or normal diet for a month. For stereological analysis, the animals were perfused 1 month after intranigral injection and fixed intracardially with ice-cold PBS. The tissues were post-fixed with 4% paraformaldehyde overnight. The fixed mice brains were cryoprotected in 30% sucrose (wt/vol) in PBS and subsequently processed for immunohistochemistry.
4.12. TH Stereological Cell Counting and Immunohistochemistry
Coronal sections (thickness, 35 µm) of the fixed and cryoprotected brains were cut, including the substantia nigra. The sections were incubated with rabbit polyclonal anti-TH (1:1000, Novus Biologicals, Centennial, CO, USA) antibody followed by sequential incubations with biotinylated goat anti-rabbit IgG and streptavidin-conjugated HRP using a Vectastain ABC kit (Vector Laboratories, Burlingame, CA, USA) according to the manufacturer’s instructions. To visualize TH-positive cells, 3,3-diaminobenzidine (DAB, cat# D4293, Sigma-Aldrich, St. Louis, MO, USA) was used as the HRP substrate. The total number of TH-positive neurons in the substantia nigra pars compacta was determined using the Optical Fractionator probe in Stereo Investigator software (MicroBrightfield, Williston, VT, USA). All stereological counting was performed in a manner blinded to the mouse treatments. For immunofluorescence labeling, coronal ventral midbrain sections were blocked in a blocking buffer (1× PBS, 5% normal goat serum, and 0.2% Triton-X) for 1 h at room temperature. The brain sections were incubated with the primary antibody mixture overnight at 4 °C, followed by brief washing and subsequent incubation in PBS containing fluorescent dye-labeled secondary antibodies. Fluorescent images were obtained using a fluorescence microscope (Axiovert, 200 M; Carl Zeiss, Oberkochen, Germany) for image acquisition.
4.13. Behavior Tests
4.13.1. Open Field Test
The open field uses a square plastic box (40 cm × 40 cm × 40 cm) divided into 64 (8 × 8) equal areas (5 cm × 5 cm). The field was divided into border, peripheral, and central areas. The central area contained four central squares (2 × 2). The peripheral area contained 12 squares surrounding the central region, and the border area comprised the remaining squares. The mice were placed in the middle of the box and left for 15 min. After each test, 70% ethanol was used to clean the open fields. Every trial used Smart v3.0 software (Panlab, Barcelona, Spain) to record the distance traveled as a measure of locomotor activity and the time spent in and entries to the center as an anxiolytic indicator.
4.13.2. Rotarod Test
Before the test trial, the mice were trained to walk in the rotarod for 2 days. On the test day (day 3), the mice were placed on the accelerating cylinder, and the latency and speed of falling off the rotarod were recorded. The speed of the rotarod was increased from 4 to 40 rpm within 5 min. The test was terminated when the mouse fell off the rotarod.
4.13.3. Pole Test
The pole is a bandaged metal rod that is 58 cm long and 10 mm in diameter. For the pole test, mice were allowed to adapt to the cage for at least 5 min. When the mice were adapted to the cage with the pole, they were placed at the end of the stick facing down. The total time until the animal came down the pole and put its forefoot on the floor was recorded. Latency was calculated as the average time of the three trials.
4.14. Statistics
Quantitative data are presented as the mean ± standard error of the mean. Statistical significance was assessed using an analysis of variance test with Tukey’s honest significant difference post hoc analysis for comparisons of more than three groups. Differences were considered statistically significant at p < 0.05. GraphPad Prism software (GraphPad Software, San Diego, CA, USA) was used to prepare all plots and perform all statistical analyses.




 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}