
Lack of Neuroprotection with a Single Intravenous Infusion of Human Amnion Epithelial Cells after Severe Hypoxia–Ischemia in Near-Term Fetal Sheep


 and
and
Abstract
:1. Introduction
2. Results
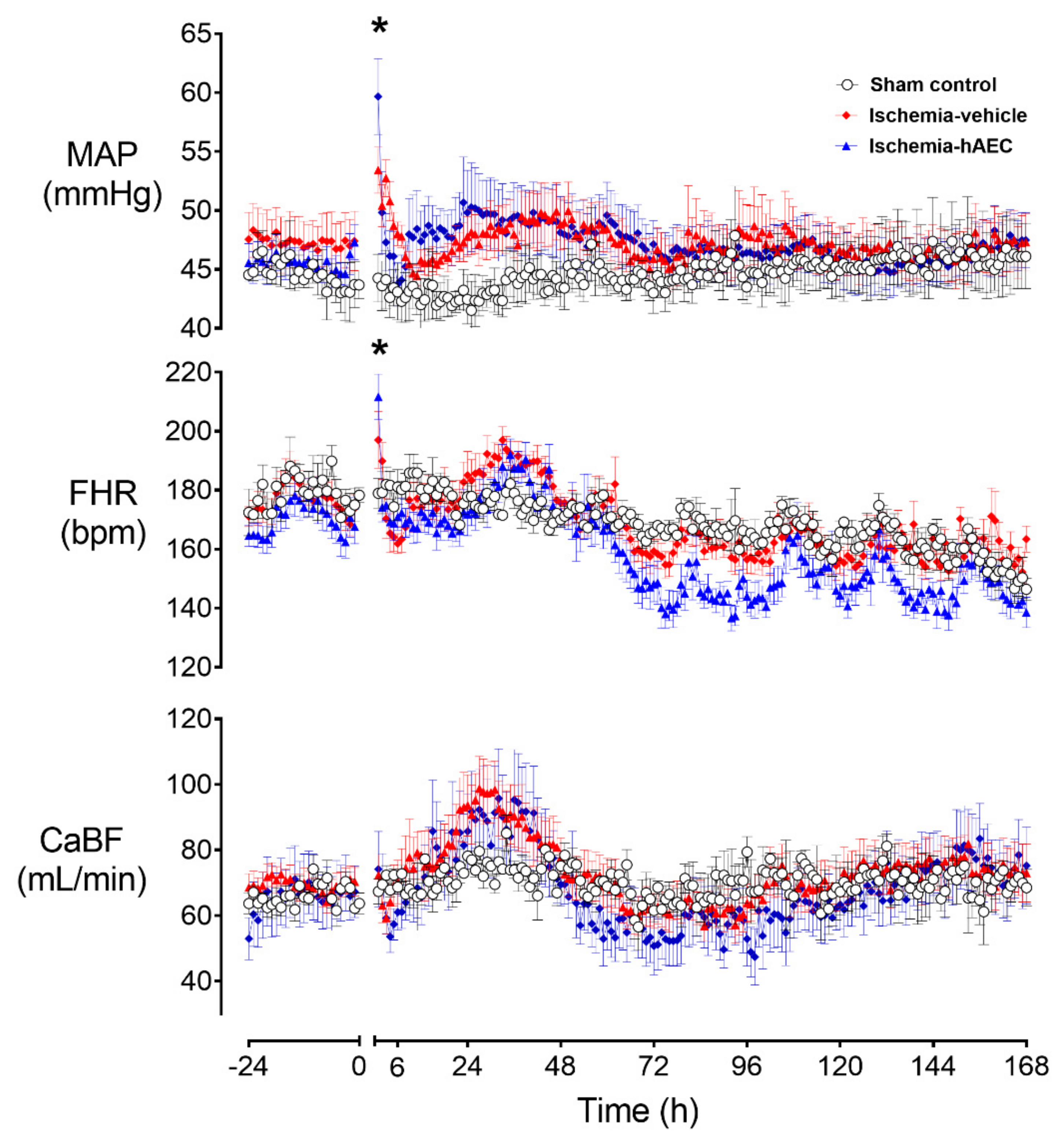
2.1. Physiology
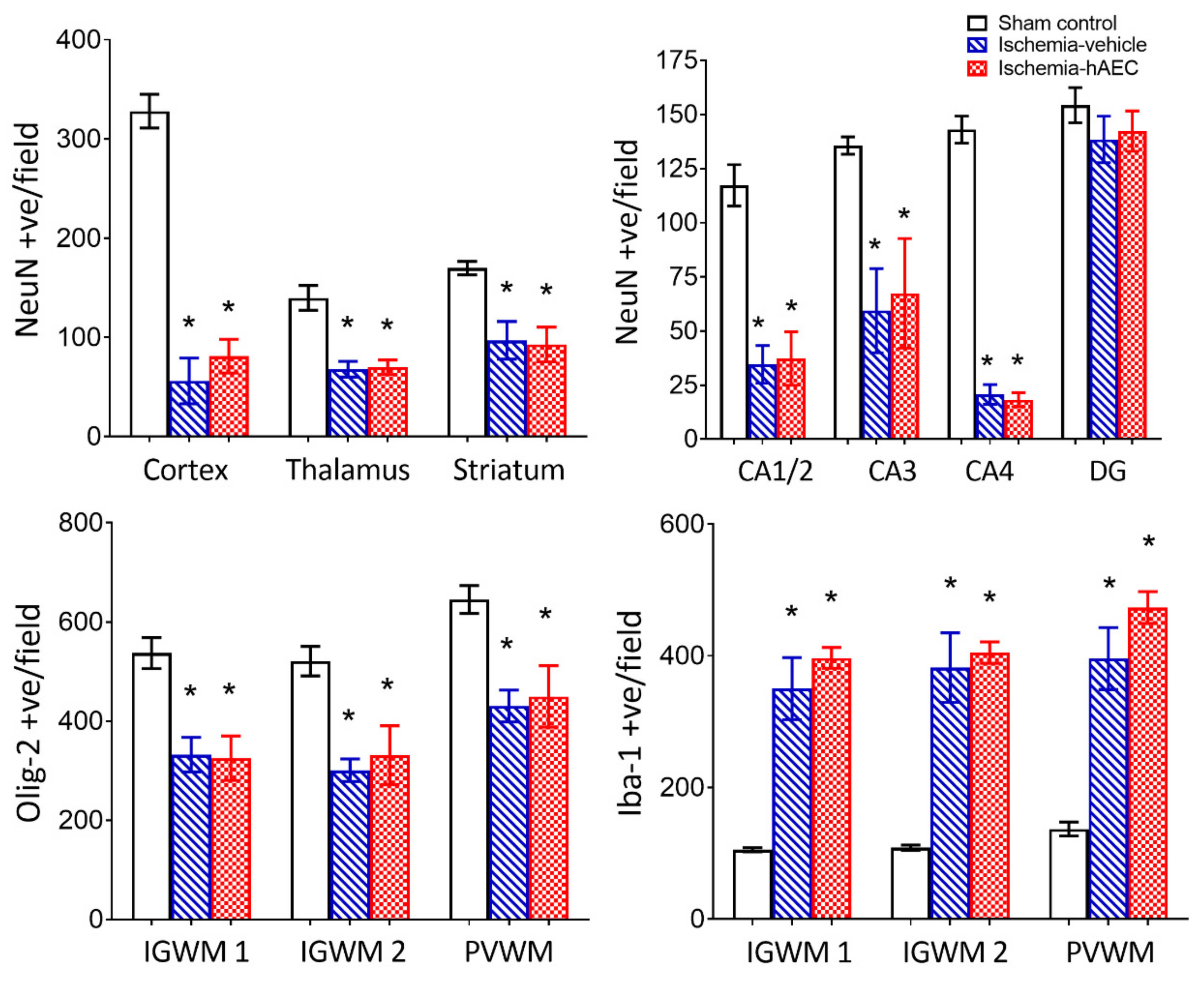
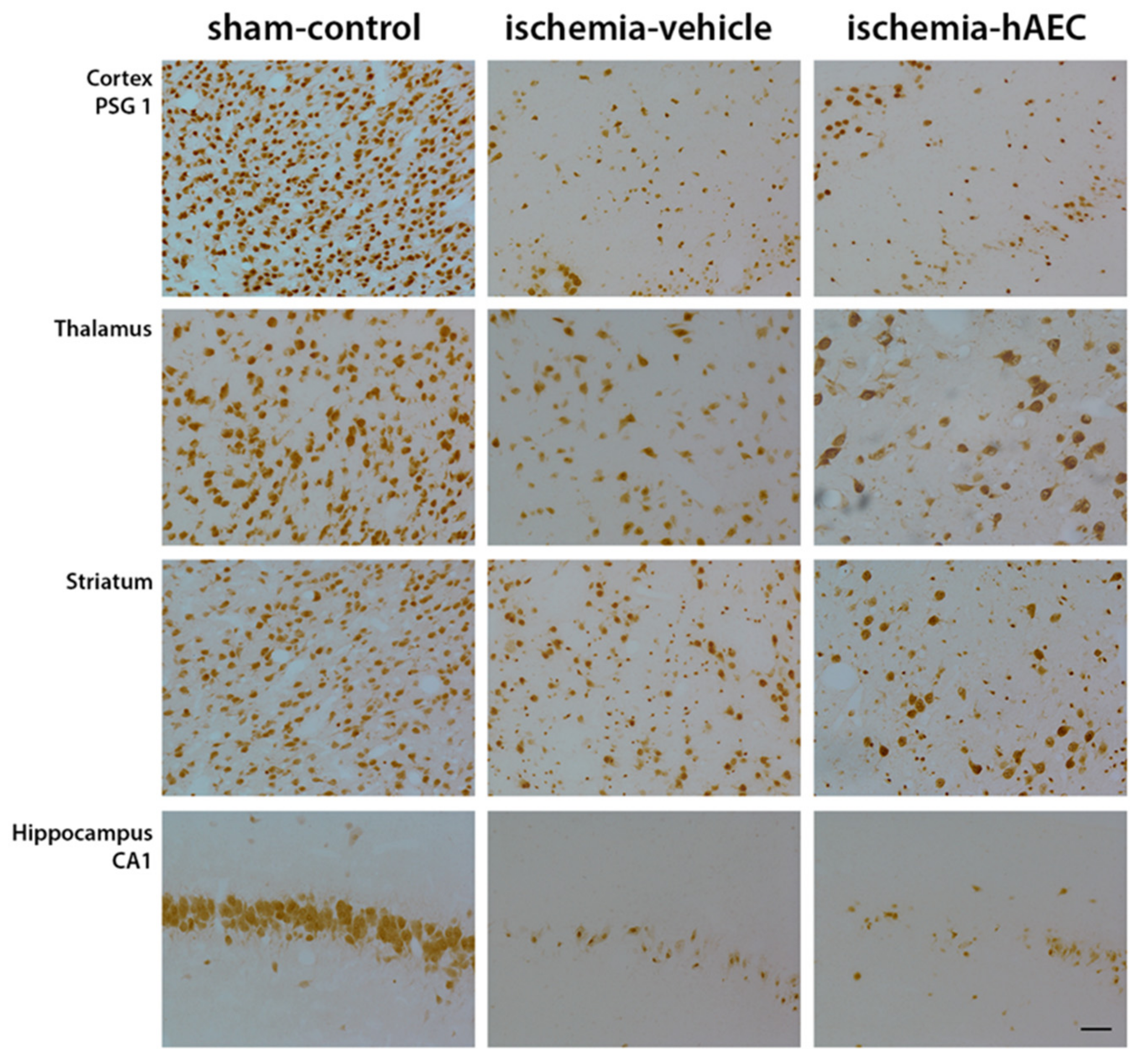
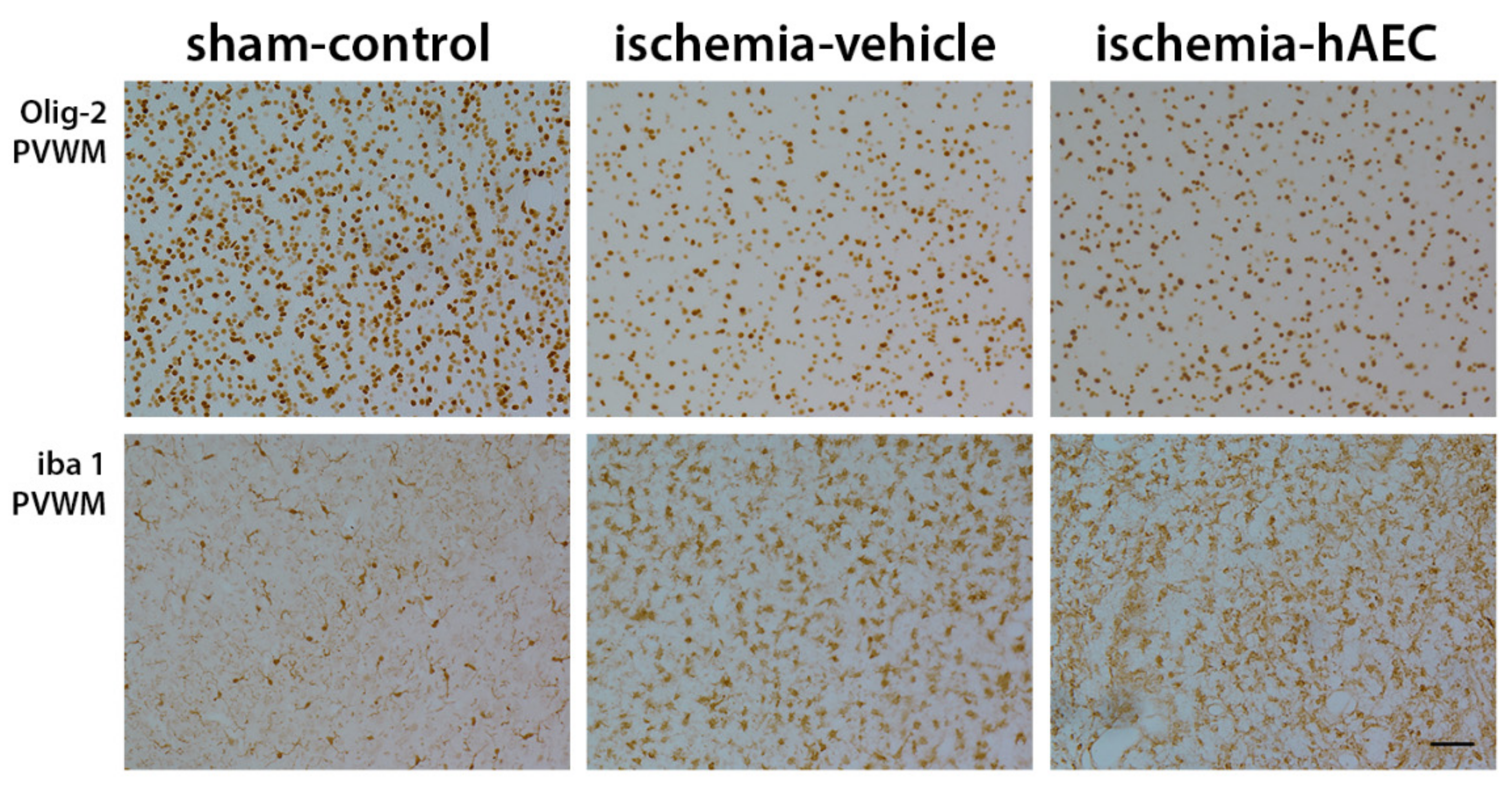
2.2. Histology
3. Discussion
4. Methods and Materials
4.1. Post-Operative Care
4.2. Experimental Protocol
4.3. Cell Preparation
4.4. Immunohistochemistry
4.5. Data Analysis
4.6. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ivain, P.; Montaldo, P.; Khan, A.; Elagovan, R.; Burgod, C.; Morales, M.M.; Pant, S.; Thayyil, S. Erythropoietin monotherapy for neuroprotection after neonatal encephalopathy in low-to-middle income countries: A systematic review and meta-analysis. J. Perinatol. 2021, 41, 2134–2140. [Google Scholar] [CrossRef] [PubMed]
- Graham, E.M.; Ruis, K.A.; Hartman, A.L.; Northington, F.J.; Fox, H.E. A systematic review of the role of intrapartum hypoxia-ischemia in the causation of neonatal encephalopathy. Am. J. Obstet. Gynecol. 2008, 199, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.; Kozuki, N.; Blencowe, H.; Vos, T.; Bahalim, A.; Darmstadt, G.L.; Niermeyer, S.; Ellis, M.; Robertson, N.J.; Cousens, S.; et al. Intrapartum-related neonatal encephalopathy incidence and impairment at regional and global levels for 2010 with trends from 1990. Pediatr. Res. 2013, 74 (Suppl. 1), 50–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, A.J.; Groenendaal, F. Delayed neuroprotection in the era of hypothermia: What can we add? J. Clin. Neonatol. 2016, 5, 3–7. [Google Scholar] [CrossRef]
- Azzopardi, D.; Strohm, B.; Marlow, N.; Brocklehurst, P.; Deierl, A.; Eddama, O.; Goodwin, J.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; et al. Effects of hypothermia for perinatal asphyxia on childhood outcomes. N. Engl. J. Med. 2014, 371, 140–149. [Google Scholar] [CrossRef] [Green Version]
- Thayyil, S.; Pant, S.; Montaldo, P.; Shukla, D.; Oliveira, V.; Ivain, P.; Bassett, P.; Swamy, R.; Mendoza, J.; Moreno-Morales, M.; et al. Hypothermia for moderate or severe neonatal encephalopathy in low-income and middle-income countries (HELIX): A randomised controlled trial in India, Sri Lanka, and Bangladesh. Lancet Glob. Health 2021, 9, e1273–e1285. [Google Scholar] [CrossRef]
- Castillo-Melendez, M.; Yawno, T.; Jenkin, G.; Miller, S.L. Stem cell therapy to protect and repair the developing brain: A review of mechanisms of action of cord blood and amnion epithelial derived cells. Front. Neurosci. 2013, 7, 194. [Google Scholar] [CrossRef] [Green Version]
- Bennet, L.; Tan, S.; Van den Heuij, L.; Derrick, M.; Groenendaal, F.; Van Bel, F.; Juul, S.; Back, S.A.; Northington, F.; Robertson, N.J.; et al. Cell therapy for neonatal hypoxia-ischemia and cerebral palsy. Ann. Neurol. 2012, 71, 589–600. [Google Scholar] [CrossRef]
- Chicha, L.; Smith, T.; Guzman, R. Stem cells for brain repair in neonatal hypoxia-ischemia. Childs Nerv. Syst. 2014, 30, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Fleiss, B.; Guillot, P.V.; Titomanlio, L.; Baud, O.; Hagberg, H.; Gressens, P. Stem cell therapy for neonatal brain injury. Clin. Perinatol. 2014, 41, 133–148. [Google Scholar] [CrossRef]
- Donega, V.; van Velthoven, C.T.; Nijboer, C.H.; van Bel, F.; Kas, M.J.H.; Kavelaars, A.; Heijnen, C.J. Intranasal mesenchymal stem cell treatment for neonatal brain damage: Long-term cognitive and sensorimotor improvement. PLoS ONE 2013, 8, e51253. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; McDonald, C.A.; Fahey, M.C.; Jenkin, G.; Miller, S.L. Could cord blood cell therapy reduce preterm brain injury? Front. Neurol. 2014, 5, 200. [Google Scholar] [CrossRef]
- Antoniadou, E.; David, A.L. Placental stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 31, 13–29. [Google Scholar] [CrossRef]
- Diaz-Prado, S.; Muinos-Lopez, E.; Hermida-Gomez, T.; Cicione, C.; Rendal-Vázquez, M.E.; Fuentes-Boquete, I.; de Toro, F.J.; Blanco, F.J. Human amniotic membrane as an alternative source of stem cells for regenerative medicine. Differentiation 2011, 81, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Miki, T. Amnion-derived stem cells: In quest of clinical applications. Stem Cell Res. Ther. 2011, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Davidson, J.O.; van den Heuij, L.G.; Fraser, M.; Wassink, G.; Miller, S.L.; Lim, R.; Wallace, E.M.; Jenkin, G.; Gunn, A.J.; Bennet, L. Window of opportunity for human amnion epithelial stem cells to attenuate astrogliosis after umbilical cord occlusion in preterm fetal sheep. Stem Cells Transl. Med. 2020, 10, 427–440. [Google Scholar] [CrossRef]
- Van den Heuij, L.G.; Fraser, M.; Miller, S.L.; Jenkin, G.; Wallace, E.M.; Davidson, J.O.; Lear, C.A.; Lim, R.; Wassink, G.; Gunn, A.J.; et al. Delayed intranasal infusion of human amnion epithelial cells improves white matter maturation after asphyxia in preterm fetal sheep. J. Cereb. Blood Flow Metab. 2019, 39, 223–239. [Google Scholar] [CrossRef]
- Yawno, T.; Schuilwerve, J.; Moss, T.J.; Vosdoganes, P.; Westover, A.J.; Afandi, E.; Jenkin, G.; Wallace, E.M.; Miller, S.L. Human amnion epithelial cells reduce fetal brain injury in response to intrauterine inflammation. Dev. Neurosci. 2013, 35, 272–282. [Google Scholar] [CrossRef]
- Yawno, T.; Sabaretnam, T.; Li, J.; McDonald, C.; Lim, R.; Jenkin, G.; Wallace, E.M.; Miller, S.L. Human amnion epithelial cells protect against white matter brain injury after repeated endotoxin exposure in the preterm ovine fetus. Cell Transplant. 2017, 26, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Leaw, B.; Zhu, D.; Tan, J.; Muljadi, R.; Saad, M.I.; Mockler, J.C.; Wallace, E.M.; Lim, R.; Tolcos, M. Human amnion epithelial cells rescue cell death via immunomodulation of microglia in a mouse model of perinatal brain injury. Stem Cell Res. Ther. 2017, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Barlow, R.M. The foetal sheep: Morphogenesis of the nervous system and histochemical aspects of myelination. J. Comp. Neurol. 1969, 135, 249–262. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, G.H.; Baghurst, K.I.; Potter, B.J.; Hetzel, B.S. Foetal brain development in the sheep. Neuropathol. Appl. Neurobiol. 1979, 5, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Chan, K.Y.Y.; Papagianis, P.; Nitsos, I.; Zahra, V.; Allison, B.; Polglase, G.R.; McDonald, C.A. Umbilical Cord Blood Cells Do Not Reduce Ventilation-Induced Lung Injury in Preterm Lambs. Front. Physiol. 2020, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal stromal cells: Sensors and switchers of inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Robertson, N.J.; Meehan, C.; Martinello, K.A.; Avdic-Belltheus, A.; Boggini, T.; Mutshiya, T.; Lingam, I.; Yang, Q.; Sokolska, M.; Charalambous, X.; et al. Human umbilical cord mesenchymal stromal cells as an adjunct therapy with therapeutic hypothermia in a piglet model of perinatal asphyxia. Cytotherapy 2021, 23, 521–535. [Google Scholar] [CrossRef]
- Pendharkar, A.V.; Chua, J.Y.; Andres, R.H.; Wang, N.; Gaeta, X.; Wang, H.; De, A.; Choi, R.; Chen, S.; Rutt, B.K.; et al. Biodistribution of neural stem cells after intravascular therapy for hypoxic-ischemia. Stroke 2010, 41, 2064–2070. [Google Scholar] [CrossRef] [Green Version]
- Vu, Q.; Xie, K.; Eckert, M.; Zhao, W.; Cramer, S.C. Meta-analysis of preclinical studies of mesenchymal stromal cells for ischemic stroke. Neurology 2014, 82, 1277–1286. [Google Scholar] [CrossRef]
- Xu, J.; Feng, Z.; Wang, X.; Xiong, Y.; Wang, L.; Ye, L.; Zhang, H. hUC-MSCs Exert a Neuroprotective Effect via Anti-apoptotic Mechanisms in a Neonatal HIE Rat Model. Cell Transplant. 2019, 28, 1552–1559. [Google Scholar] [CrossRef]
- Baak, L.M.; Wagenaar, N.; van der Aa, N.E.; Groenendaal, F.; Dudink, J.; Tataranno, M.L.; Mahamuud, U.; Verhage, C.H.; Eijsermans, R.M.; Smit, L.S.; et al. Feasibility and safety of intranasally administered mesenchymal stromal cells after perinatal arterial ischaemic stroke in the Netherlands (PASSIoN): A first-in-human, open-label intervention study. Lancet Neurol. 2022, 21, 528–536. [Google Scholar] [CrossRef]
- Penny, T.R.; Pham, Y.; Sutherland, A.E.; Mihelakis, J.G.; Lee, J.; Jenkin, G.; Fahey, M.C.; Miller, S.L.; McDonald, C.A. Multiple doses of umbilical cord blood cells improve long-term brain injury in the neonatal rat. Brain Res. 2020, 1746, 147001. [Google Scholar] [CrossRef]
- Wassink, G.; Galinsky, R.; Drury, P.P.; Gunn, E.R.; Bennet, L.; Gunn, A.J. Does maturity affect cephalic perfusion and T/QRS ratio during prolonged umbilical cord occlusion in fetal sheep? Obstet. Gynecol. Int. 2014, 2014, 314159. [Google Scholar] [CrossRef] [Green Version]
- Drury, P.P.; Davidson, J.O.; van den Heuij, L.G.; Wassink, G.; Gunn, E.R.; Booth, L.C.; Bennet, L.; Gunn, A.J. Status epilepticus after prolonged umbilical cord occlusion is associated with greater neural injury in fetal sheep at term-equivalent. PLoS ONE 2014, 9, e96530. [Google Scholar] [CrossRef]
- Williams, C.E.; Gunn, A.J.; Mallard, C.; Gluckman, P.D. Outcome after ischemia in the developing sheep brain: An electroencephalographic and histological study. Ann. Neurol. 1992, 31, 14–21. [Google Scholar] [CrossRef]
- Van Bel, F.; Roman, C.; Klautz, R.J.; Teitel, D.F.; Rudolph, A.M. Relationship between brain blood flow and carotid arterial flow in the sheep fetus. Pediatr. Res. 1994, 35, 329–333. [Google Scholar]
- Covert, R.F.; Schreiber, M.D.; Torgerson, L.J.; Torgerson, R.W.; Miletich, D.J. Prediction of cerebral blood flow in fetal lambs by carotid artery ultrasonic flow transducer. Reprod. Fertil. Dev. 1996, 8, 157–162. [Google Scholar] [CrossRef]
- Murphy, S.; Rosli, S.; Acharya, R.; Mathias, L.; Lim, R.; Wallace, E.; Jenkin, G. Amnion epithelial cell isolation and characterization for clinical use. Curr. Protoc. Stem Cell Biol. 2010, 13, 1E.6.1–1E.6.25. [Google Scholar] [CrossRef]
- Vosdoganes, P.; Lim, R.; Koulaeva, E.; Chan, S.T.; Acharya, R.; Moss, T.J.; Wallace, E.M. Human amnion epithelial cells modulate hyperoxia-induced neonatal lung injury in mice. Cytotherapy 2013, 15, 1021–1029. [Google Scholar] [CrossRef]
- Davidson, J.O.; Drury, P.P.; Green, C.R.; Nicholson, L.F.; Bennet, L.; Gunn, A.J. Connexin hemichannel blockade is neuroprotective after asphyxia in preterm fetal sheep. PLoS ONE 2014, 9, e96558. [Google Scholar] [CrossRef]
- Scher, M.S.; Aso, K.; Beggarly, M.E.; Hamid, M.Y.; Steppe, D.A.; Painter, M.J. Electrographic seizures in preterm and full-term neonates: Clinical correlates, associated brain lesions, and risk for neurologic sequelae. Pediatrics 1993, 91, 128–134. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Sex | Singleton/Twin | Body Weight (g) | Brain Weight (g) |
|---|---|---|---|---|
| Sham Control | 5F3M | 8S | 4588.3.3 ± 359 | 40.1± 1.4 |
| Ischemia-vehicle | 3M3F1U | 6S1T | 4697.1.3 ± 240 | 39 ± 1.2 |
| Ischemia-hAEC | 5F1M | 6S | 4112.3 ± 238 | 38 ± 1.4 |
| Group | Baseline | 2 h | 4 h | 6 h | 1 d | 2 d | 3 d | 4 d | 5 d | 6 d | 7 d | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sham-control | pH | 7.38 ± 0.01 | 7.38 ± 0.01 | 7.38 ± 0.01 | 7.39 ± 0.01 | 7.38 ± 0.01 | 7.37 ± 0.01 | 7.37 ± 0.01 | 7.36 ± 0.01 | 7.35 ± 0.01 | 7.34 ± 0.01 | 7.35 ± 0.01 |
| Ischemia-vehicle | 7.4 ± 0.01 | 7.39 ± 0.01 | 7.41 ± 0.01 | 7.40 ± 0.01 | 7.38 ± 0.02 | 7.39 ± 0.01 | 7.37 ± 0.01 | 7.37 ± 0.01 | 7.37 ± 0.01 | 7.37 ± 0.01 | 7.37 ± 0.01 | |
| Ischemia-hAEC | 7.34 ± 0.03 | 7.32 ± 0.03 * | 7.33 ± 0.03 # | 7.35 ± 0.02 # | 7.35 ± 0.02 | 7.35 ± 0.01 | 7.34 ± 0.01 | 7.31 ± 0.01 *,# | 7.33 ± 0.01 | 7.33 ± 0.02 | 7.35 ± 0.01 | |
| Sham-control | pCO2 | 48.8 ± 4.3 | 47.4 ± 2.8 | 48.8 ± 1.5 | 47.3 ± 0.1 | 47.2 ± 1.1 | 48.3 ± 1.3 | 46.4 ± 1.2 | 48.7 ± 0.7 | 50.3 ± 0.5 | 49.5 ± 1.8 | 46.8 ± 0.1 |
| Ischemia-vehicle | 44.6 ± 1.4 | 42.6 ± 1.6 | 43.4 ± 1.4 | 44.9 ± 1.9 | 44.4 ± 2.7 | 44.1 ± 1.3 | 44.5 ± 1.9 | 45.5 ± 1.2 | 46.7 ± 1.9 | 46.7 ± 1.6 | 49.4 ± 1.9 * | |
| Ischemia-hAEC | 55.3 ± 1.3 *,# | 52.3 ± 1.3 *,# | 53.1 ± 1.2 *,# | 54.0 ± 1.0 *,# | 54.2 ± 0.8 *,# | 52.8 ± 1.3 # | 53 ± 1.3 # | 54.2 ± 1.2 # | 51.8 ± 1.4 | 55.5 ± 1.2 # | 53.9 ± 1.0 * | |
| Sham-control | pO2 | 21.4 ± 0.3 | 21.6 ± 1.2 | 21.2 ± 1.1 | 20.9 ± 1.2 | 20.1 ± 1.3 | 22.6 ± 3.1 | 22.2 ± 2.9 | 22.9 ± 2.2 | 24.3 ± 3.6 | 26 ± 2.7 | 23.7 ± 2.2 |
| Ischemia-vehicle | 26.0 ± 1.1 | 25.8 ± 1.2 | 25.9 ± 1.3 | 25.4 ±1.9 | 25.0 ± 2.0 | 24.6 ±1.5 | 26.7 ± 1.6 | 25.5 ± 1.9 | 26.0 ± 1.9 | 24.7 ± 1.4 | 22.9 ± 1.1 | |
| Ischemia-hAEC | 22.9 ± 1.3 | 20.6 ± 1.2 | 20.1 ± 0.6 | 20.6 ± 1.0 | 19.1 ± 1.2 | 19.5 ± 1.4 | 26.9 ± 1.0 | 26.6 ± 0.9 | 24.8 ± 1.2 | 24.3 ± 1.2 | 24.2 ± 0.8 | |
| Sham-control | Lactate | 0.9 ± 0.2 | 1.0 ± 0.2 | 1.2 ± 0.0 | 1.2 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.2 | 0.9 ±0.4 | 1.0 ± 0.2 | 0.8 ± 0.2 | 0.8 ± 0.3 | 0.8 ± 0.4 |
| Ischemia-vehicle | 1.1 ± 0.1 | 2.4 ± 0.5 * | 2.3 ± 0.5 * | 2.1 ± 0.4 * | 4.7 ± 1.1 * | 0.9 ± 0.1 | 1.0 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.1 | |
| Ischemia-hAEC | 1.2 ± 0.1 | 3.8 ± 0.7 * | 3.9 ± 0.8 * | 3.5 ± 0.7 * | 2.5 ± 0.7 * | 2.6 ± 0.7 * | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.1 | |
| Sham-control | Glucose | 0.6 ± 0.3 | 0.6 ± 0.2 | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.7 ± 0.1 | 0.6 ± 0.3 | 0.6 ± 0.1 | 0.6 ± 0.2 | 0.6 ± 0.1 | 0.5 ± 0.2 |
| Ischemia-vehicle | 1.0 ± 0.1 | 1.3 ± 0.1 * | 1.3 ± 0.1 * | 1.3 ± 0.1 * | 1.5 ± 0.1 * | 0.9 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | |
| Ischemia-hAEC | 1.0 ± 0.1 | 1.6 ± 0.1 * | 1.4 ± 0.1 * | 1.4 ± 0.1 * | 1.1 ± 0.2 | 1.3 ± 0.3 * | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.9 ± 0.1 | 0.8 ± 0.1 | 0.9 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davidson, J.O.; van den Heuij, L.G.; Dhillon, S.K.; Miller, S.L.; Lim, R.; Jenkin, G.; Gunn, A.J.; Bennet, L. Lack of Neuroprotection with a Single Intravenous Infusion of Human Amnion Epithelial Cells after Severe Hypoxia–Ischemia in Near-Term Fetal Sheep. Int. J. Mol. Sci. 2022, 23, 8393. https://doi.org/10.3390/ijms23158393
Davidson JO, van den Heuij LG, Dhillon SK, Miller SL, Lim R, Jenkin G, Gunn AJ, Bennet L. Lack of Neuroprotection with a Single Intravenous Infusion of Human Amnion Epithelial Cells after Severe Hypoxia–Ischemia in Near-Term Fetal Sheep. International Journal of Molecular Sciences. 2022; 23(15):8393. https://doi.org/10.3390/ijms23158393
Chicago/Turabian StyleDavidson, Joanne O., Lotte G. van den Heuij, Simerdeep K. Dhillon, Suzanne L. Miller, Rebecca Lim, Graham Jenkin, Alistair J. Gunn, and Laura Bennet. 2022. "Lack of Neuroprotection with a Single Intravenous Infusion of Human Amnion Epithelial Cells after Severe Hypoxia–Ischemia in Near-Term Fetal Sheep" International Journal of Molecular Sciences 23, no. 15: 8393. https://doi.org/10.3390/ijms23158393
APA StyleDavidson, J. O., van den Heuij, L. G., Dhillon, S. K., Miller, S. L., Lim, R., Jenkin, G., Gunn, A. J., & Bennet, L. (2022). Lack of Neuroprotection with a Single Intravenous Infusion of Human Amnion Epithelial Cells after Severe Hypoxia–Ischemia in Near-Term Fetal Sheep. International Journal of Molecular Sciences, 23(15), 8393. https://doi.org/10.3390/ijms23158393