
Changes in Endogenous Oxytocin Levels and the Effects of Exogenous Oxytocin Administration on Body Weight Changes and Food Intake in Polycystic Ovary Syndrome Model Rats

Abstract
:1. Introduction
2. Results
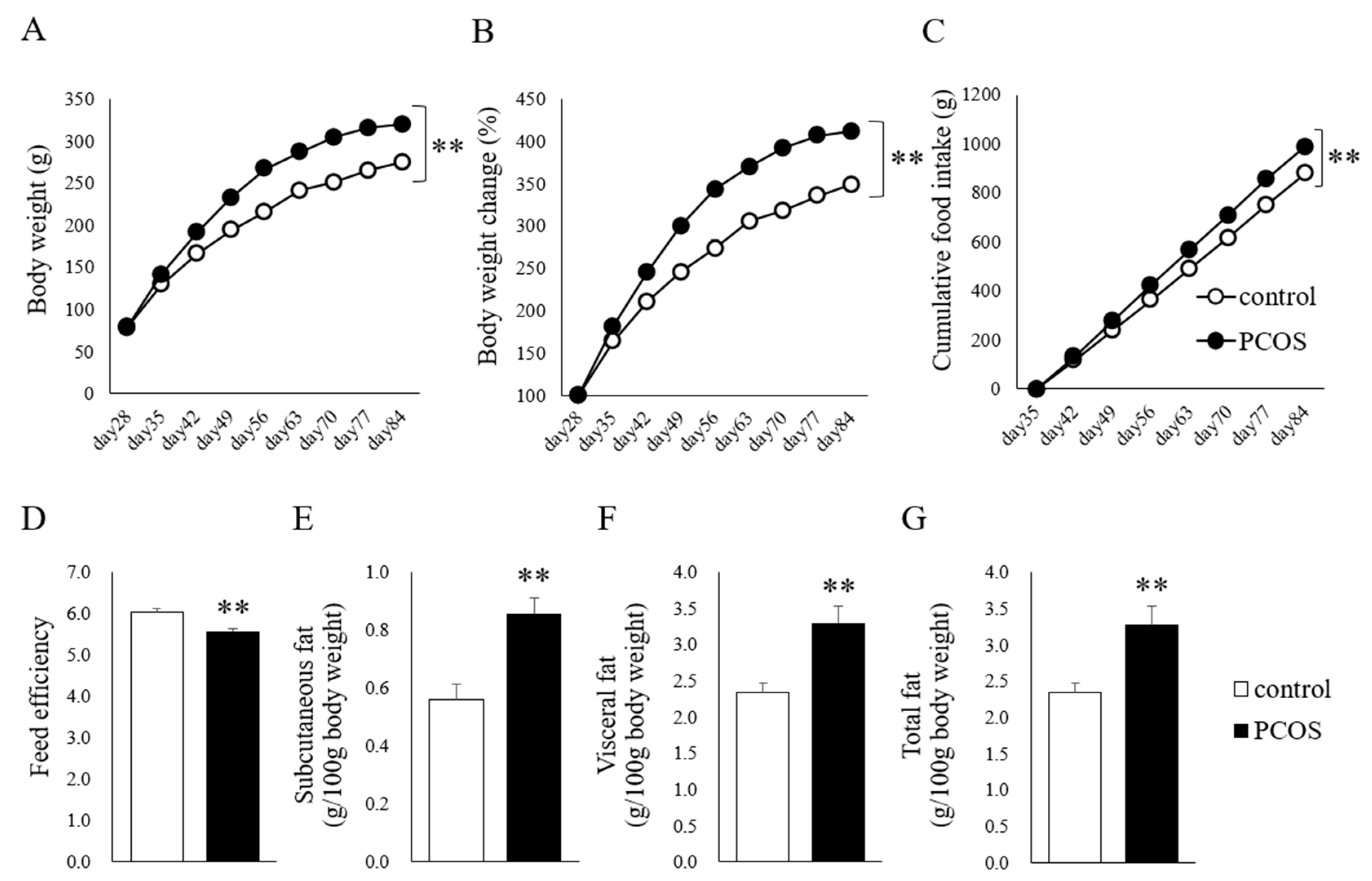
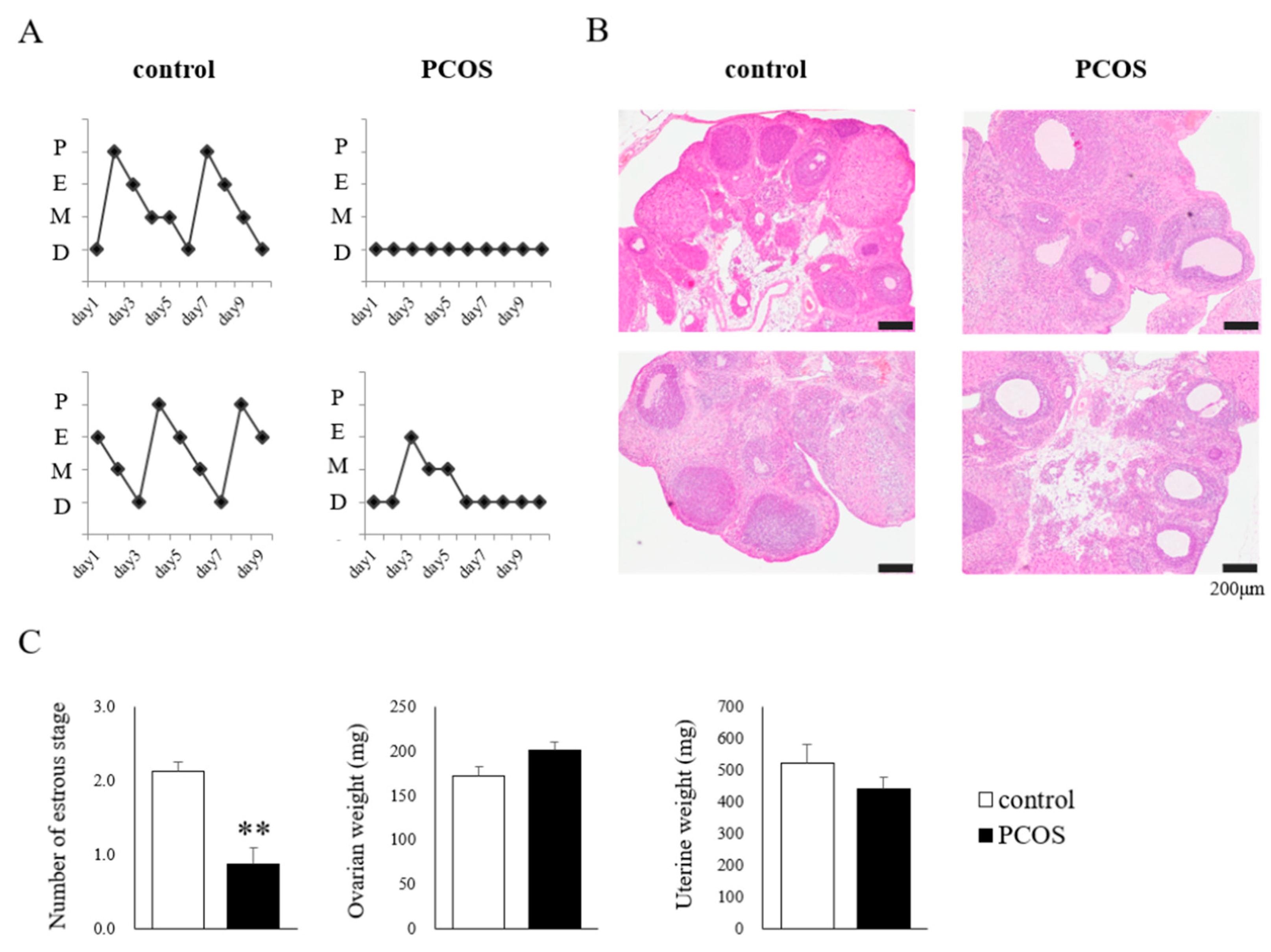
2.1. Reproductive and Metabolic Phenotypes
2.2. Central and Peripheral OT Levels
2.3. Effects of Acute OT Administration on Food Intake and Body Weight
2.4. Effects of Chronic OT Administration on Food Intake and Body Weight
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Analysis of the Reproductive and Metabolic Phenotypes and Tissue Sampling
4.3. Evaluation of the Effects of Acute OT Administration on Food Intake and Body Weight
4.4. Evaluation of the Effect of Chronic OT Administration on Food Intake and Body Weight
4.5. Hormone Assay
4.6. Histology
4.7. Real-Time Polymerase Chain Reaction (PCR)
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lauritsen, M.P.; Bentzen, J.G.; Pinborg, A.; Loft, A.; Thuesen, L.L.; Cohen, A.; Hougaard, D.M.; Andersen, A.N. The prevalence of polycystic ovary syndrome in a normal population according to the Rotterdam criteria versus revised criteria including anti-Mullerian hormone. Hum. Reprod. 2014, 29, 791–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfield, R.L.; Ehrmann, D.A. The pathogenesis of polycystic ovary syndrome (PCOS): The hypothesis of PCOS as functional ovarian hyperandrogenism revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Hardiman, P.J.; Petersen, I.; Wang, F.F.; Qu, F.; Baio, G. The prevalence of polycystic ovary syndrome in reproductive-aged women of different ethnicity: A systematic review and meta-analysis. Oncotarget 2017, 8, 96351–96358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotterdam ESHRE/ASRM-sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar] [CrossRef]
- Ehrmann, D.A. Polycystic ovary syndrome. N. Engl. J. Med. 2005, 352, 1223–1236. [Google Scholar] [CrossRef]
- Moran, L.J.; Misso, M.L.; Wild, R.A.; Norman, R.J. Impaired glucose tolerance, type 2 diabetes and metabolic syndrome in polycystic ovary syndrome: A systematic review and meta-analysis. Hum. Reprod. Update 2010, 16, 347–363. [Google Scholar] [CrossRef]
- Ladson, G.; Dodson, W.C.; Sweet, S.D.; Archibong, A.E.; Kunselman, A.R.; Demers, L.M.; Willianms, N.I.; Coney, P.; Legro, R.S. The effects of metformin with lifestyle therapy in polycystic ovary syndrome: A randomized double-blind study. Fertil. Steril. 2011, 95, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Bates, G.W.; Legro, R.S. Longterm management of polycystic ovarian syndrome (PCOS). Mol. Cell. Endocrinol. 2013, 373, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Foster, G.D.; Wyatt, H.R.; Hill, J.O.; Angela, P.M.; Makris, P.; Rosenbaum, D.L.; Brill, C.; Stein, R.I.; Mohammed, B.S.; Miller, B.; et al. Weight and metabolic outcomes after 2 years on a low-carbohydrate versus low-fat diet: A randomized trial. Ann. Intern. Med. 2010, 153, 147–157. [Google Scholar] [CrossRef]
- Du Vigneaud, V.; Ressler, C.; Trippett, S. The sequence of amino acids in oxytocin, with a proposal for the structure of oxytocin. J. Biol. Chem. 1953, 205, 949–957. [Google Scholar] [CrossRef]
- Altirriba, J.; Poher, A.L.; Rohner-Jeanrenaud, F. Chronic oxytocin administration as a treatment against impaired leptin signaling or leptin resistance in obesity. Front. Endocrinol. 2015, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Maejima, Y.; Iwasaki, Y.; Yamahara, Y.; Kodaira, M.; Sedbaxar, U.; Yada, T. Peripheral oxytocin treatment ameliorates obesity by reducing food intake and visceral fat mass. Aging 2011, 3, 1169–1177. [Google Scholar] [CrossRef]
- Maejima, Y.; Rita, R.S.; Santoso, P.; Aoyama, M.; Hiraoka, Y.; Nishimori, K.; Gantulga, D.; Shimomura, K.; Yada, T. Nasal oxytocin administration reduces food intake without affecting locomotor activity and glycemia with c-Fos induction in limited brain areas. Neuroendocrinology 2015, 101, 35–44. [Google Scholar] [CrossRef]
- Maejima, Y.; Aoyama, M.; Sakamoto, K.; Jojima, T.; Aso, Y.; Takasu, K.; Takenoshita, S.; Shimomura, K. Impact of sex, fat distribution and initial body weight on oxytocin’s body weight regulation. Sci. Rep. 2017, 7, 8599. [Google Scholar] [CrossRef] [Green Version]
- Maejima, Y.; Yokota, S.; Nishimori, K.; Shimomura, K. The anorexigenic neural pathways of oxytocin and their clinical implication. Neuroendocrinology 2018, 107, 91–104. [Google Scholar] [CrossRef]
- Zhang, G.; Bai, H.; Zhang, H.; Dean, C.; Wu, Q.; Li, J.; Guariglia, S.; Meng, Q.; Cai, D. Neuropeptide exocytosis involving synaptotagmin-4 and oxytocin in hypothalamic programming of body weight and energy balance. Neuron 2011, 69, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Cai, D. Circadian intervention of obesity development via resting-stage feeding manipulation or oxytocin treatment. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1004–E1012. [Google Scholar] [CrossRef]
- Roberts, Z.S.; Wolden-Hanson, T.; Matsen, M.E.; Ryu, V.; Vaughan, C.H.; Graham, J.L.; Havel, P.L.; Chukri, D.W.; Schwartz, M.W.; Morton, G.J.; et al. Chronic hindbrain administration of oxytocin is sufficient to elicit weight loss in diet-induced obese rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R357–R371. [Google Scholar] [CrossRef] [Green Version]
- Arletti, R.; Benelli, A.; Bertolini, A. Oxytocin inhibits food and fluid intake in rats. Physiol. Behav. 1990, 48, 825–830. [Google Scholar] [CrossRef]
- Blevins, J.E.; Thompson, B.W.; Anekonda, V.T.; Ho, J.M.; Graham, J.L.; Roberts, Z.S.; Hwang, B.H.; Ogimoto, K.; Wolden-Hanson, T.; Nelson, J. Chronic CNS oxytocin signaling preferentially induces fat loss in high-fat diet-fed rats by enhancing satiety responses and increasing lipid utilization. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R640–R658. [Google Scholar] [CrossRef] [Green Version]
- Deblon, N.; Veyrat-Durebex, C.; Bourgoin, L.; Caillon, A.; Bussier, A.L.; Petrosino, S.; Piscitelli, F.; Legros, J.J.; Geenen, V.; Foti, M.; et al. Mechanisms of the anti-obesity effects of oxytocin in diet-induced obese rats. PLoS ONE 2011, 6, e25565. [Google Scholar] [CrossRef] [Green Version]
- Morton, G.J.; Thatcher, B.S.; Reidelberger, R.D.; Ogimoto, K.; Wolden-Hanson, T.; Baskin, D.G.; Schwartz, M.W.; Belvins, J.E. Peripheral oxytocin suppresses food intake and causes weight loss in diet-induced obese rats. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E134–E144. [Google Scholar] [CrossRef] [PubMed]
- Olson, B.R.; Drutarosky, M.D.; Chow, M.S.; Hruby, V.J.; Stricker, E.M.; Verbalis, J.G. Oxytocin and an oxytocin agonist administered centrally decrease food intake in rats. Peptides 1991, 12, 113–118. [Google Scholar] [CrossRef]
- Ong, Z.Y.; Alhadeff, A.L.; Grill, H.J. Medial nucleus tractus solitarius oxytocin receptor signaling and food intake control: The role of gastrointestinal satiation signal processing. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R800–R806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maejima, Y.; Sakuma, K.; Santoso, P.; Gantulga, D.; Katsurada, K.; Ueta, Y.; Hiraoka, Y.; Nishimori, K.; Tanaka, S.; Shimomura, K.; et al. Oxytocinergic circuit from paraventricular and supraoptic nuclei to arcuate POMC neurons in hypothalamus. FEBS Lett. 2014, 58, 4404–4412. [Google Scholar] [CrossRef] [Green Version]
- Blevins, J.E.; Graham, J.L.; Morton, G.J.; Bales, K.L.; Schwartz, M.W.; Baskin, D.G.; Havel, P.J. Chronic oxytocin administration inhibits food intake, increases energy expenditure, and produces weight loss in fructose-fed obese rhesus monkeys. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R431–R438. [Google Scholar] [CrossRef] [Green Version]
- Lawson, E.A.; Marengi, D.A.; DeSanti, R.L.; Holmes, T.M.; Schoenfeld, D.A.; Tolley, C.J. Oxytocin reduces caloric intake in men. Obesity 2015, 23, 950–956. [Google Scholar] [CrossRef]
- Thienel, M.; Fritsche, A.; Heinrichs, M.; Peter, A.; Ewers, M.; Lehnert, H.; Born, J.; Hallschmid, M. Oxytocin’s inhibitory effect on food intake is stronger in obese than normal-weight men. Int. J. Obes. 2016, 40, 707–1714. [Google Scholar] [CrossRef] [Green Version]
- Altirriba, J.; Poher, A.L.; Caillon, A.; Arsenijevic, D.; Veyrat-Durebex, C.; Lyaute, J.; Dulloo, A.; Rohner-Jeanrenaud, F. Divergent effects of oxytocin treatment of obese diabetic mice on adiposity and diabetes. Endocrinology 2014, 155, 4189–4201. [Google Scholar] [CrossRef] [Green Version]
- Plante, E.; Menaouar, A.; Danalache, B.A.; Yip, D.; Broderick, T.L.; Chiasson, J.L.; Jankowski, M.; Gutkowska, J. Oxytocin treatment prevents the cardiomyopathy observed in obese diabetic male db/db mice. Endocrinology 2015, 156, 1416–1428. [Google Scholar] [CrossRef]
- Kamada, S.; Yamamoto, Y.; Aoki, H.; Tamura, K.; Takeda, A.; Minato, S.; Masaki, R.; Yanagihara, R.; Hayashi, N.; Yano, Y.; et al. A novel PCOS rat model and an evaluation of its reproductive, metabolic, and behavioral phenotype. Reprod. Med. Biol. 2021, 21, e12416. [Google Scholar] [CrossRef]
- Iwasa, T.; Matsuzaki, T.; Mayila, Y.; Kawakita, T.; Yanagihara, R.; Irahara, M. The effects of chronic oxytocin administration on body weight and food intake in DHT-induced PCOS model rats. Gynecol. Endocrinol. 2020, 36, 55–60. [Google Scholar] [CrossRef]
- Gajdosechova, L.; Krskova, K.; Segarra, A.B.; Spolcova, A.; Suski, M.; Olszanecki, R.; Zorad, S. Hypooxytocinaemia in obese Zucker rats relates to oxytocin degradation in liver and adipose tissue. J. Endocrinol. 2014, 220, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Zhu, T.; Tang, B.; Yu, S.; Hu, H.; Sun, W.; Pan, R.; Wang, J.; Wang, D.; Yang, L.; et al. Decreased circulating levels of oxytocin in obesity and newly diagnosed type 2 diabetic patients. J. Clin. Endocrinol. Metab. 2014, 99, 4683–4689. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.; Qian, W.; Pan, R.; Jia, J.; Jiang, D.; Yang, Q.; Wang, S.; Liu, Y.; Yu, S.; Hu, H.; et al. Reduced circulating oxytocin and high-molecular-weight adiponectin are risk factors for metabolic syndrome. Endocr. J. 2016, 63, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Balagura, S.; Harrell, L.E.; Roy, E. Effect of the light-dark cycle on neuroendocrine and behavioral responses to scheduled feeding. Physiol. Behav. 1975, 15, 245–247. [Google Scholar] [CrossRef]
- Iwasa, T.; Matsuzaki, T.; Mayila, Y.; Yanagihara, R.; Yamamoto, Y.; Kawakita, T.; Kuwahara, A.; Irahara, M. Oxytocin treatment reduced food intake and body fat and ameliorated obesity in ovariectomized female rats. Neuropeptide 2019, 75, 49–57. [Google Scholar] [CrossRef]
- Otgontsetseg, E.; Kato, T.; Kawakita, T.; Kasai, K.; Yoshida, K.; Iwasa, T.; Irahara, M. Effects of peripheral oxytocin administration on body weight, food intake, adipocytes, and biochemical parameters in peri- and postmenopausal female rats. Endocr. J. 2021, 68, 7–16. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Annealing T (°C) |
|---|---|---|
| OT forward | GAA CAC CAA CGC CAT GGC CTG CCC | 62 |
| OT reverse | TCG GTG CGG CAG CCA TCC GGG CTA | |
| OTR forward | CGA TTG CTG GGC GGT CTT | 67 |
| OTR reverse | CCG CCG CTG CCG TCT TGA | |
| NPY forward | GGG GCT GTG TGG ACT GAC CCT | 66 |
| NPY reverse | GAT GTA GTG TCG CAG AGC GGA G | |
| AgRP forward | TGA AGA AGA CAG CAG CAG ACC | 63 |
| AgRP reverse | AAG GTA CCT GTT GTC CCAAGC | |
| POMC forward | CCT CAC CAC GGA AAG CA | 66 |
| POMC reverse | TCA AGG GCT GTT CAT CTC C | |
| pporexin forward | GCC GTC TCT ACG AAC TGT TG | 60 |
| pporexin reverse | CGA GGA GAG GGG AAA GTT AG | |
| GAPDH forward | ATG GCA CAG TCAAGG CTG AGA | 70 |
| GAPDH reverse | CGC TCC TG GAA GAT GGT GAT | |
| 18S rRNA forward | GAC GGA CCA GAG CGA AAG C | 64 |
| 18S rRNA reverse | AAC CTC CGA CTT TCG TTC TTG A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, S.; Noguchi, H.; Takeda, A.; Arakaki, R.; Uchishiba, M.; Imaizumi, J.; Minato, S.; Kamada, S.; Kagawa, T.; Yoshida, A.; et al. Changes in Endogenous Oxytocin Levels and the Effects of Exogenous Oxytocin Administration on Body Weight Changes and Food Intake in Polycystic Ovary Syndrome Model Rats. Int. J. Mol. Sci. 2022, 23, 8207. https://doi.org/10.3390/ijms23158207
Yamamoto S, Noguchi H, Takeda A, Arakaki R, Uchishiba M, Imaizumi J, Minato S, Kamada S, Kagawa T, Yoshida A, et al. Changes in Endogenous Oxytocin Levels and the Effects of Exogenous Oxytocin Administration on Body Weight Changes and Food Intake in Polycystic Ovary Syndrome Model Rats. International Journal of Molecular Sciences. 2022; 23(15):8207. https://doi.org/10.3390/ijms23158207
Chicago/Turabian StyleYamamoto, Shota, Hiroki Noguchi, Asuka Takeda, Ryosuke Arakaki, Maimi Uchishiba, Junki Imaizumi, Saki Minato, Shuhei Kamada, Tomohiro Kagawa, Atsuko Yoshida, and et al. 2022. "Changes in Endogenous Oxytocin Levels and the Effects of Exogenous Oxytocin Administration on Body Weight Changes and Food Intake in Polycystic Ovary Syndrome Model Rats" International Journal of Molecular Sciences 23, no. 15: 8207. https://doi.org/10.3390/ijms23158207
APA StyleYamamoto, S., Noguchi, H., Takeda, A., Arakaki, R., Uchishiba, M., Imaizumi, J., Minato, S., Kamada, S., Kagawa, T., Yoshida, A., Kawakita, T., Yamamoto, Y., Yoshida, K., Kon, M., Shinohara, N., & Iwasa, T. (2022). Changes in Endogenous Oxytocin Levels and the Effects of Exogenous Oxytocin Administration on Body Weight Changes and Food Intake in Polycystic Ovary Syndrome Model Rats. International Journal of Molecular Sciences, 23(15), 8207. https://doi.org/10.3390/ijms23158207