Abstract
Obstructive sleep apnea (OSA), a disease associated with excessive sleepiness and increased cardiovascular risk, affects an estimated 1 billion people worldwide. The present study examined proteomic biomarkers indicative of presence, severity, and treatment response in OSA. Participants (n = 1391) of the Stanford Technology Analytics and Genomics in Sleep study had blood collected and completed an overnight polysomnography for scoring the apnea–hypopnea index (AHI). A highly multiplexed aptamer-based array (SomaScan) was used to quantify 5000 proteins in all plasma samples. Two separate intervention-based cohorts with sleep apnea (n = 41) provided samples pre- and post-continuous/positive airway pressure (CPAP/PAP). Multivariate analyses identified 84 proteins (47 positively, 37 negatively) associated with AHI after correction for multiple testing. Of the top 15 features from a machine learning classifier for AHI ≥ 15 vs. AHI < 15 (Area Under the Curve (AUC) = 0.74), 8 were significant markers of both AHI and OSA from multivariate analyses. Exploration of pre- and post-intervention analysis identified 5 of the 84 proteins to be significantly decreased following CPAP/PAP treatment, with pathways involving endothelial function, blood coagulation, and inflammatory response. The present study identified PAI-1, tPA, and sE-Selectin as key biomarkers and suggests that endothelial dysfunction and increased coagulopathy are important consequences of OSA, which may explain the association with cardiovascular disease and stroke.
1. Introduction
Obstructive sleep apnea (OSA) is characterized by recurrent partial or complete obstructive events of the upper airway, resulting in arousals and oxygen desaturations [1]. Nocturnal polysomnography (PSG) is the current gold standard for diagnosing and evaluating sleep apnea severity [2]. The presence and severity of sleep apnea is approximated by the apnea–hypopnea index (AHI), the number of abnormal respiratory events recorded per hour of sleep during PSG, whereby an AHI ≥ 5 but <15 is indicative of mild sleep apnea, AHI ≥ 15 but <30 is indicative of moderate sleep apnea, and AHI ≥ 30 is indicative of severe sleep apnea in adults [3].
Obstructive sleep apnea is an extremely common condition. A recent study estimated almost 1 billion people worldwide are affected by OSA, with prevalence exceeding 50% in some countries [4]. The prevalence of OSA is higher in men, increases with age, and is associated with higher body mass index (BMI) [5]. A recent study in adults older than 40 reported the prevalence of moderate-to-severe sleep-disordered breathing was 49.7% in men and 23.4% in women [6]. High levels of sleep apnea, typically of the obstructive type, are associated with increased cardiovascular risk, notably increased risk of high blood pressure [7], increased vascular aging [8], heart failure [9], arrythmias [10], and stroke [11], effects that seem more correlated with hypoxia than with sleep fragmentation [10,12]. Sleep fragmentation, in contrast, correlates more with arousal disturbances and daytime sleepiness, although some authors have found higher cardiovascular risk in sleepy versus non sleepy OSA patients [10,13]. Timely diagnosis and treatment of OSA is imperative to the health and prognosis of millions.
Recent work highlighted a need for blood-based biomarkers in sleep medicine to streamline more accurate and objective ascertainment of disorders [14]. Previously, we studied 1300 serum proteins in 351 patients with OSA compared to 362 controls [15], identifying 65 proteins associated with AHI and 9 differentially expressed proteins (DEPs) in moderate-to-severe OSA (AHI ≥ 15) compared to mild OSA/controls (AHI < 15). That study demonstrated that multivariate protein measurement could be used to predict the presence of moderate-to-severe OSA (AHI ≥ 15) compared with mild OSA/controls (AHI < 15) using a machine learning classifier and identified 3 proteins that were responsive to positive airway pressure (PAP) treatment in a cohort of 16 patients with OSA. Such results supported the initial efficacy of proteomic biomarkers for diagnosing OSA as well as their potential for measuring OSA severity and treatment response.
In the present study, we extend and validate the results of our previous findings [15] by identifying proteins cross-sectionally associated with the presence and severity of OSA (i.e., total AHI and OSA severity classifications) using an independent sample with a larger panel (~5000) of proteins in a larger (~1400) sample of clinic patients. We also examined the effect of OSA treatment (i.e., CPAP/PAP and supplemental oxygen versus off treatment) on identified proteins to explore potential pathophysiological mechanisms associated with managing symptoms and consequences of OSA.
2. Results
2.1. Stanford Technology Analytics and Genomics in Sleep (STAGES) Study Cohort
Demographic and clinical information for participants (n = 1391) included in the final analyses is presented in Table 1. The final sample with all relevant outcomes had almost equal representation of males (52%) and females (48%), with an average age of 46.1 ± 15.2 years, average BMI of 30.9 ± 8.7 kg/m2, and was primarily Caucasian/white (80%) and non-Hispanic (94%). Regarding PSG, average sleep duration was 5.6 ± 1.7 h, and patients had an average AHI of 15.6 ± 19.7 (range = 0.0–154.6). According to PSG technician notes, 7% of participants were provided CPAP during the PSG. Approximately 7% of participants self-reported using CPAP or PAP, less than 1% reported supplemental oxygen use, and less than 1% reported having previous sleep apnea surgery. Regarding blood sampling, most samples (67%) were collected between 12:00 PM and 23:59 PM, with an average time from draw to blood processing of 31.0 ± 14.0 h (range = 6.8–126.3 h), during which blood was transported to Stanford University for central processing.

Table 1.
Demographic and clinical characteristics of participants included in final analyses (n = 1391).
2.2. Effect of Time to Blood Processing
To analyze the effect of travel time at room temperature, we added 224 EDTA samples collected during the daytime that were immediately processed and were also measured using the same proteomic panel. Summary results of multivariate analyses of the combined cohort (n = 1654) including all STAGES plasma samples (n = 1430; time to processing range: 1–126 h) and the immediately processed samples (n = 224; time to processing = 0) are presented in Supplementary Table S1. Not surprisingly, most proteins (n = 4151) were affected by time from blood draw to processing, including 2505 positively associated proteins and 1646 negatively associated proteins, whereby relative protein expression increased or decreased, respectively, with longer time spent at room temperature. As this may be an important consideration in the interpretation of our results, time to blood processing was included as a covariate in all multivariate analyses. Further, we include false discovery rate (FDR) corrected p-values (i.e., FDR p combined time to processing) and beta coefficients (i.e., beta combined time to processing) from the analysis, including a comparison with immediately processed samples in supplementary tables alongside results of the following multivariate analyses for additional consideration in the interpretation of our results.
2.3. Proteins Associated with the Apnea–Hypopnea Index
Proteins significantly associated with total AHI as a continuous variable are presented in Table 2 and Supplementary Table S2. Multivariate analyses identified 84 proteins (47 positively and 37 negatively) associated with AHI after FDR correction. Proteins most positively associated with AHI were E-selectin (sE-Selectin), tissue-type plasminogen activator (tPA), plasminogen activator inhibitor 1 (PAI-1), scavenger receptor cysteine rich type 1 protein (sCD163), and tissue factor pathway inhibitor (TFPI). Pathways related to fatty acid oxidation and metabolism, RA and retinol biosynthesis and metabolism, complement and coagulation cascades, blood coagulation and clotting cascades, platelet amyloid precursor protein, and plasminogen activating cascades were involved (Supplementary Table S3B). Proteins most negatively associated with AHI include sex hormone-binding globulin (SHBG), amyloid-like protein 1, Desmoglein-2, serum albumin, and secretogranin-3 (SCG3), with pathways involved in insulin-like growth factor signaling and regulation, ghrelin regulation of food intake and energy homeostasis, genes encoding structural ECM glycoproteins, and post-translational modification of synthesis of GPI-anchored proteins (Supplementary Table S3C).
Table 2.
The 84 proteins significantly associated with the apnea–hypopnea index (AHI) after 5% FDR correction.
2.4. Differential Expression of Proteins Stratified by Severity Index of AHI
Differences in characteristics of participants (n = 1391) by OSA classification status based on AHI are presented in Supplementary Table S4. Participants with moderate-to-severe OSA (AHI ≥ 15) were significantly older (50.5 ± 13.1 vs. 43.9 ± 15.7; p = 4.22 × 10−15), had higher BMI (33.6 ± 9.1 vs. 29.5 ± 8.1; p = 8.23 × 10−23), and consisted of a significantly higher proportion of males (63% vs. 41%; p = 1.63 × 10−15) compared with the mild-to-no OSA (AHI < 15) group (Supplementary Table S4A). A marginally longer time to processing was found in subjects with moderate-to-severe OSA (32.1 ± 14.4 vs. 30.5 ± 13.8, respectively; p = 0.004). Multivariate analyses identified 69 differentially expressed proteins (DEPs) between these two OSA groups, with 40 upregulated and 29 downregulated in those with moderate-to-severe OSA (Supplementary Table S5). Top upregulated proteins included Quinone oxidoreductase-like protein 1 (QORL1), Tumor protein p53-inducible protein 11 (P5I11), cystine-rich protein 1 (CRIP1), DCN1-like protein 5 (DCNL5), and Tartrate-resistant acid phosphatase type 5 (TrATPase), with pathways similar to those listed for proteins positively associated with AHI (Supplementary Table S6A). Top downregulated proteins included Amyloid-like protein 1, Desmoglein-2, SHBG, secretogranin-3 (SCG3), and vesicular, overexpressed in cancer, prosurvival protein 1 (ECOP), with pathways like those listed for proteins negatively associated with AHI, plus fibrinolysis pathways, complement system pathways, and others (Supplementary Table S6C). In comparison to AHI-associated proteins found using linear regression, 50 of 69 DEPs for this OSA classification were significant in AHI multivariate analyses.
Summary characteristics comparing severe OSA, moderate OSA, mild OSA, and controls are presented in Supplementary Table S4B–D. Thirty-two significant DEPs (20 upregulated, 12 downregulated) were found in severe OSA compared to controls, and 4 significant DEPs (1 upregulated, 3 downregulated) were found in moderate OSA compared to controls (presented in Supplementary Tables S7 and S8, respectively). No DEPs were found between mild OSA and controls.
2.5. Protein Markers Validated with a Machine Learning Classifier
Many proteins (n = 84) were significantly associated with AHI when modeled as a continuous variable, and the largest number of proteins (n = 69) was identified when AHI was stratified by severity using the cutoff score of 15 as a binary variable. The AHI ≥ 15 cutoff score was therefore next used to train a machine learning classifier on SomaScan protein measures using three different models. Model 1 incorporated protein measures and demographic variables and achieved 66.4% accuracy (F1 = 0.598) in classifying moderate-to-severe OSA. Model 2 incorporated only proteins and achieved 66.9% accuracy (F1 = 0.603). Model 3, with only demographics, achieved the lowest accuracy of 63.5% (F1 = 0.566). Receiver operating curves (ROC) and areas under the curves (AUCs) for each model are presented in Figure 1, classifier metrics are presented in Table 3, and the confusion matrix is in Supplementary Table S9. Of the top 15 classifier features from Model 1 (Supplementary Table S10), 10 proteins were identified as significant protein markers of AHI, and 8 were significant DEPs for moderate-to-severe OSA (Amyloid-like protein 1, tPA, PAI-1, CRIP1, SCG3, ECOP, Desmoglein-2, and SHBG), suggesting these proteins are robust predictors of the presence and severity of sleep apnea. When comparing the models, we see a trend in the differences for Model 1 compared with Model 3, and Model 2 compared with Model 3 (Supplementary Table S11A). It seems here that the improvement in prediction coming from the protein expression variables over the demographics is the main factor playing into the performance of the model.
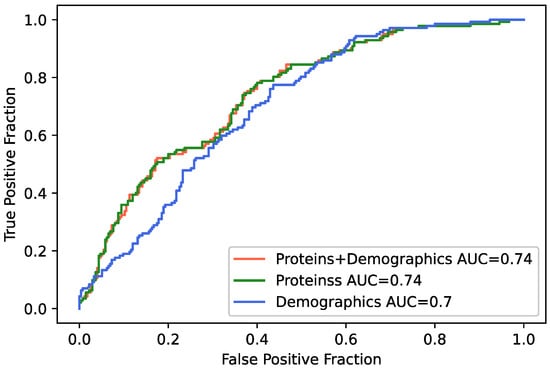
Figure 1.
Receiver operating characteristics (ROC) curves for machine learning classifier for moderate-to-severe obstructive sleep apnea. Model 1 (red line) included all 4985 proteins as well as demographic variables (i.e., age, sex, BMI); Model 2 (green line) included only the 4985 proteins; and Model 3 (blue line) included only demographic variables.
Table 3.
Performance metrics of a machine learning classifier for moderate-to-severe OSA (AHI ≥ 15) trained on 974 samples (331 cases, 643 controls) and validated on 417 samples (142 cases, 275 controls).
2.6. Effect of CPAP and Supplemental Oxygen on Protein Markers
Proteins associated with CPAP/PAP intervention are presented in Supplementary Table S12A. For this comparison, we only used the 84 proteins previously shown to be associated with total AHI. Nonparametric paired samples tests comparing pre- and post-intervention protein expression identified 5 proteins significantly reduced following use of CPAP/PAP: CRP (change = −12.3%; p = 0.004), PAI-1 (change = −10.6%; p = 0.007), TrATPase (change = −6.2%; p = 0.009), sE-Selectin (change = −8.2%; p = 0.030), and tPA (change = −14.1%; p = 0.047). These proteins are associated with pathways involved in blood coagulation, inflammatory response, fibrinolysis, and plasminogen activation. Regarding changes in protein expression with supplemental oxygen compared with air (i.e., sham), when patients were off CPAP, nonparametric paired comparisons identified significant differences in protein expression change for PAI-1 and TGF-b R III (Supplementary Table S12B), whereby PAI-1 decreased with supplemental oxygen and increased with air, whereas TGF-b R III increased with supplemental oxygen and decreased with air. Regarding the proteins with significant changes following CPAP/PAP, CRP and tPA decreased, and TrATPase and sE-Selectin increased with supplemental oxygen, whereas these proteins all increased with air. However, these differences did not reach statistical significance.
2.7. Replication with Our Prior Study in Serum
In comparison with our previous findings in serum [15], 44 (52%) of the 84 proteins associated with total AHI in the present study were not included in the SomaScan 1.3 panel used in our previous study, and therefore could not be included in this comparison (see Supplementary Table S13 for a complete list). Seven (18%) of the 40 remaining protein markers of AHI were replicated in the present study, including tPA, TFPI, Aminoacylase-1, LG3BP, Factor I, and Coagulation Factor IXab (all positively associated with AHI), and UNC5H4 (negatively associated with AHI). We further conducted multivariate linear regression analyses for the 1282 mutual proteins between the 1.3K and 5K panels in the combined STAGES/Stanford Sleep Study cohort (n = 1921). This analysis resulted in 77 proteins (49 positive, 28 negative) significantly associated with AHI (Supplementary Table S14), including 22 proteins identified in the STAGES cohort alone. Significant proteins included sE-Selectin, PAI-1, Coagulation Factor IXab, TFPI, IGFBP-2, UNC5H4, SHBG, CRP, TrATPase, and tPA, with pathway analysis identifying functions consistent with those previously described for AHI as a continuous variable (Supplementary Table S15).
We further trained a similar machine learning classifier as previously described with the 5 proteins significant for AHI in the STAGES cohort, combined cohort, and that were responsive to CPAP (i.e., CRP, PAI-I, TrATPase, tPA, and sE-Selectin) for classifying participants into OSA or control categories based on an AHI cutoff score of 15. ROCs and AUCs for each model are presented in Figure 2, and classifier metrics are presented in Table 4. Performance metrics for Model 1 (proteins + demographics) and Model 2 (proteins only) were similar to, albeit slightly lower than, those from the classifier using the entire array of 4985 proteins. Model 1 with the targeted proteins achieved the highest accuracy of 65.7% (F1 = 0.571) compared to an accuracy of 66.4% (F1 = 0.598) from the entire protein array. There were significant differences between Model 1 and Model 2 (DeLong: Z = 2.94; p = 0.003; Bootstrap: D = 2.95; p = 0.003) and Model 1 and Model 3 (DeLong: Z = 2.44; p = 0.014; Bootstrap: D = 2.43; p = 0.015; Supplementary Table S11B). In this case, the integration of proteins with the demographics showed a clear improvement over the two individual data sources. Collectively, this suggests that this subset of proteins seems to contribute a significant proportion of the predictive ability of the presence, severity, and treatment response for obstructive sleep apnea.
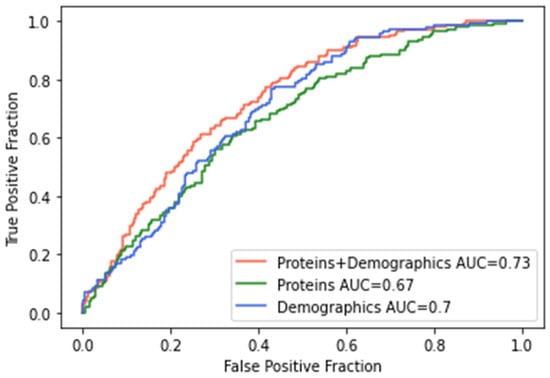
Figure 2.
Receiver operating characteristics (ROC) curves for machine learning classifier for moderate-to-severe obstructive sleep apnea in replicated and CPAP proteins. Model 1 (red line) included the five replicated proteins as well as demographic variables (i.e., age, sex, BMI) and achieved the highest accuracy of 66%; Model 2 (green line) included only the five proteins, and Model 3 (blue line) included only demographic variables.
Table 4.
Performance metrics of a machine learning classifier using only the 5 replicated proteins that were also CPAP responsive for moderate-to-severe OSA (AHI ≥ 15) trained on 974 samples (331 cases, 643 controls) and validated on 417 samples (142 cases, 275 controls).
3. Discussion
These results extend our prior study of 713 patients with 1500 proteins using an independent sample with a similar but larger platform and larger sample. In this study, we found 84 proteins (47 positive and 37 negatively) associated with total AHI, 50 of which were also significant DEPs between moderate-to-severe OSA (AHI ≥ 15) and mild/no OSA (AHI < 15). Of the 42 significant predictive features of the OSA machine learning classifier, 8 of the top 15 features were significant markers of total AHI and OSA from multivariate analysis, suggesting that these 8 proteins (Amyloid-like protein 1, tPA, PAI-1, CRIP1, SCG3, ECOP, Desmoglein-2, and SHBG) are likely predictors of the presence and severity of sleep apnea. Furthermore, five of the 84 significant protein markers for AHI (CRP, PAI-1, TrATPase, sE-Selectin, and tPA) were significantly affected by CPAP/PAP treatment and were replicated in a combined analysis with the samples from our previous study. These results suggest that this subset of proteins represents robust predictors of the presence and severity of sleep apnea and may be potential markers of treatment response in sleep apnea management.
The results obtained here must be considered more unbiased than candidate marker studies reported in the literature, as we studied over 5000 proteins, a study more akin to conducting a genome-wide association study versus candidate gene studies. Encouragingly, the present study validated markers identified in our previous findings with 1500 proteins in serum [15], notably increased tPA and sE-selectin, two factors that have been consistently reported in multiple studies to be associated with sleep apnea [16,17,18]. This is despite the fact the assays were done in EDTA plasma that had travelled for over 24 h at room temperature prior to processing in the STAGES samples. These two factors are primarily secreted by the endothelium in the setting of damage or injury, confirming that sleep apnea likely causes sleep fragmentation and damages the endothelium, perhaps as the result of hypoxia. These results support the notion that sleep apnea is associated with accelerated vascular aging [8]. Similarly, CRP, a marker of inflammation secreted by the liver and known to correlate with BMI, has been shown by multiple investigators to be higher in BMI-matched individuals with sleep apnea, and to also decrease with CPAP therapy [19,20].
A novel, potentially robust sleep apnea marker identified in this study is Tartrate-resistant acid phosphatase (TrATPase), also called acid phosphatase 5, tartrate resistant (ACP5). TrATPase is a glycosylated monomeric metalloprotein enzyme important to the function of osteoclasts, although it is also secreted by macrophages and immune cells. Deficiency in TrATPase is associated with spondylenchondrodysplasia, a disorder that combines abnormal bone development, various autoimmune diseases and central nervous abnormalities. Interestingly, OSA has been associated with osteoporosis in recent studies [21,22,23], a possible reflection of hypoxia.
The present study also identified PAI-1, another important coagulation factor, as significantly upregulated in sleep apnea, consistent with other studies [16,24]. Intriguingly, PAI-1 was not altered in our first published proteomic study, which used serum samples [15]. One possible explanation is that PAI-1 has a large platelet pool [25,26], thus measures in serum and plasma have different roles, explaining the discrepancy across the two studies, and suggesting that free PAI-1, the active form, is the culprit in the present findings. Another explanation could be that our first study was not powered to detect such an association in PAI-1, which would explain why PAI-1 was amongst the top proteins significantly associated with total AHI in the combined analysis.
In the context of pathobiological mechanisms underlying sleep apnea, PAI-1 is hypoxically regulated and an inhibitor of tPA, and dual elevation of PAI-1and tPA is associated with increases in coagulation and fibrinolysis turnover. Indeed, following injury, PAI-1 helps maintain fibrinolysis to local regions of tPA release [26,27]. Increased tPA and PAI-1 have also been suggested to increase local blood–brain barrier permeability [26]. More recently, increased PAI-1 and tPA have also demonstrated associations with post COVID-19 hypercoagulability [28,29], a state that induces thrombosis [30], a likely result of hypoxia. It is our hypothesis (Figure 3) that endothelial damage in sleep apnea, perhaps as the result of hypoxia, induces increased local fibrin aggregation and fibrinolysis. In this context, a dual increase of tPA and PAI-1 has been found to predict stroke [31], and they are further elevated post stroke [32,33], suggesting these to be possible mediators of the important association of stroke with sleep apnea. This may also explain the protective effect of CPAP in patients with OSA [34,35], whereby the present study identified significant reductions in PAI-1 and tPA, along with sE-Selectin, TrATPase, and CRP, after at least two weeks on CPAP/PAP. This suggests that treatment with CPAP/PAP reduces the hypoxic burden associated with sleep apnea, thereby reducing fibrinolysis, blood coagulation, plasminogen activation, and inflammatory responses.
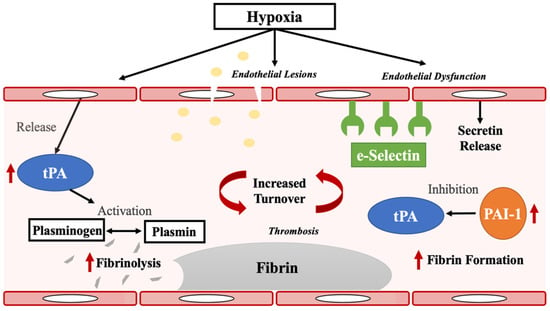
Figure 3.
Proposed mechanisms in the relationship between obstructive sleep apnea, PAI-1, tPA, and sE-Selectin. Hypoxia in sleep apnea causes endothelial damage or injury. This is associated with (1) increased release of sE-selectin; and (2) stimulation of local fibrin aggregation. Increased tPA and PAI-1 activity reflects enhanced fibrin formation/fibrinolysis turnover, perhaps contributing to the known increased risk of stroke in sleep apnea.
There are important limitations to consider when interpreting our results. Although the SomaScan assay has demonstrated good validity and reproducibility as well as stability in protein expression with varying time to processing [36,37], the blood samples from different sites were shipped to Stanford for processing at room temperature, and therefore may have been subject to blood composition changes associated with such a delay. All PSGs were conducted as part of standard-of-care according to the clinic protocol at each site and using the sampling rate/filters recommended by the datacenter; therefore, certain methodologies may have varied between study sites. Similarly, evaluation of the response to CPAP/PAP treatment was conducted in a combined sample of participants from two separate cohorts with different methodologies. Neither of the intervention cohorts included a control condition. Furthermore, the combined sample size of 25 in CPAP/PAP evaluation may not be sufficiently powered to examine such a relationship, and future research should include larger sample sizes and comparisons with controls. We relied on self-reported outcomes for demographic and secondary clinical conditions (e.g., BMI, cardiovascular disease). Finally, although we found biomarkers that were cross-sectionally associated with sleep apnea and that responded to CPAP, our machine learning models achieved relatively low accuracy, and addition of these biomarkers to demographics only marginally improved performance in our ROC curve analysis. One reason for this could be the dependance of these markers on demographics, and/or that these markers only contribute marginally to risk of OSA. This result outlines the need for finding additional biomarkers by increasing the CPAP sample size.
4. Materials and Methods
4.1. Stanford Technology Analytics and Genomics in Sleep Study Cohort
The Stanford Technology Analytics and Genomics in Sleep (STAGES) study, described previously [38,39], collected data from patients across 11 different sleep clinics between 2018 and 2020. Briefly, all participants were patients who attended an appointment with a physician at a sleep clinic and completed an overnight polysomnography (PSG) study, in addition to completing the Alliance Sleep Questionnaire (ASQ) and providing a blood sample. Although 1430 participants completed the study, a total of 1391 participants had relevant PSG, ASQ, and blood sampling data and were included in final analyses. All investigations were carried out following the rules of the Declaration of Helsinki of 1975, each institution’s Institutional Review Board approved all study procedures, and participants provided written informed consent. Data collected from STAGES, aside from the blood samples, are available through the National Sleep Research Resource (NSRR).
4.1.1. Polysomnography
All participants completed a Level 1 nocturnal PSG study in a sleep lab. All PSGs were conducted as part of standard-of-care according to the clinic protocol at each site and using the sampling rate/filters recommended by the datacenter. Outcomes of interest for the present study included total PSG duration (h), sleep duration (h), apnea–hypopnea index (AHI, with hypopnea defined as 3% desaturation or arousal). Individuals undergoing PSG exclusively for treatment purposes (e.g., oral appliance evaluation) were excluded at enrollment, but CPAP or other OSA treatment was used in some cases, and these participants were included in this analysis. AHI was scored based on the American Academy of Sleep Medicine’s recommended criteria for scoring a respiratory event as a hypopnea [3], and the presence and severity of OSA was characterized by the AHI score. A cutoff score of 15 was used to stratify patients to moderate-to-severe OSA (AHI ≥ 15) or mild-to-no OSA (AHI < 15). We also subclassified OSA cases into severe (AHI ≥ 30), moderate (30 > AHI ≥ 15), or mild (15 > AHI ≥ 5) in comparison to controls (AHI < 5) using standard cutoff scores [1,3].
4.1.2. Alliance Sleep Questionnaire
The ASQ is an electronic, comprehensive sleep disorder questionnaire that includes questions and validated measures designed to collect standardized subjective sleep data [39,40]. The questionnaire uses complex branching logic in a modular fashion to guide participants through a comprehensive set of questions to quantify demographic and clinical characteristics, including age, sex, body mass index (BMI), race, ethnicity, and associated medical history (e.g., medication use, presence of comorbidities, previous diagnosis of sleep conditions, sleep complaints). All patients completed the ASQ within approximately 4 weeks of the polysomnography.
4.1.3. Blood Sampling
Blood samples were typically collected during the scheduled PSG, either the evening before or the morning after the PSG, or within four weeks of the PSG. Times of blood draw were dichotomized (i.e., blood draw) into morning (i.e., 0:00–11:59) and evening (i.e., 12:00–23:59) samples to control for diurnal variation in blood composition. Blood samples were then shipped at room temperature, in most cases taking 1–2 days to arrive at Stanford, and processed for EDTA plasma and DNA extraction. As there was variability in the amount of time samples remained at room temperature (i.e., time to blood processing), time to processing was included in all analyses as a salient covariate. Time to processing was calculated as the total number of hours between time of blood sample collection to time plasma was frozen, which was then log-normalized, and extreme outliers (log-normalized processing time < 2.7) were removed (n = 7) to reduce variability in protein expression based on a long duration of processing time.
4.2. Intervention Cohorts
EDTA plasma samples from two different studies were used to examine the effect of CPAP/PAP and oxygen use on protein biomarkers of sleep apnea. One study recruited local community members and was conducted at Washington University in St. Louis (WashU) as described previously [41]. It included 16 patients with OSA (7 mild (5 < AHI < 15); 11 moderate-to-severe (AHI ≥ 15)) who underwent PAP treatment for 1–4 months and provided plasma samples pre- and post-treatment. Participants included in analyses were adherent to PAP, defined as usage for at least 4 h on at least 70% of the 30 preceding nights as recorded by the PAP machine. The other study recruited participants with a diagnosis of moderate-to-severe OSA who had been treated with CPAP for more than one year; this study was conducted at the University of Oxford, as described previously [42]. It included 25 participants in a two-arm randomized crossover design. Participants had moderate-to-severe OSA and had been treated with CPAP for more than 1 year, with average CPAP usage exceeding 4 h/night. After four nights of screening off CPAP, participants had at least 14 nights back on CPAP therapy before undergoing 14 nights of supplemental oxygen or 14 nights of air (both off CPAP). Participants then had a washout period of at least 14 nights back on CPAP before crossing over (e.g., air if oxygen was the first arm). Plasma samples were collected on days 0 (pre-arm 1), 14 (post-arm 1), 28 (pre-arm 2), and 42 (post-arm 2). The present study combined 16 pre-PAP samples (from the WashU study) with 25 post-air off-CPAP samples (from the Oxford study) as the “before CPAP/PAP” condition and 16 post-PAP samples (WashU) with 25 pre-air on-CPAP samples (Oxford) as the “after CPAP/PAP” condition, for a total sample size of 41 participants. We further examined the effect of supplemental oxygen compared with that of air from the samples collected (n = 25) in the Oxford study.
4.3. Stanford Sleep Cohort
We sought to extend and replicate our previous findings using the Stanford Sleep Study, as described previously [15]. Briefly, 1070 participants ages 18 to 91 were enrolled by the Stanford Sleep Clinic in 1999, and each participant provided a blood sample and completed overnight PSG. Blood samples were typically collected the morning following overnight PSG and allowed to clot for a minimum of 30 min. Serum was then aliquoted and stored at −80 °C before being shipped to SomaLogic for protein quantification. Of note, serum samples from this study were collected and processed with a mean delay of 11.6 years (i.e., from blood draw to SomaScan assay). PSG studies for cohort participants were scored using the alternate AASM hypopnea definition for AHI [43].
4.4. Protein Quantification
Plasma samples from the STAGES cohort and intervention cohorts and serum samples from the Stanford Sleep Cohort followed the same protein quantification protocol. Relative expression levels (i.e., relative florescent units (RFU)) of proteins were assayed using the SomaScan aptamer-based multiplexed platform (SomaLogic Inc., Boulder, CO, USA), which utilizes aptamers and hybridization to quantify proteins from small amounts of human plasma [44,45,46]. The SomaScan platform was designed to have extended dynamic range and includes both extracellular and intracellular proteins (with soluble domains of membrane proteins). SomaScan assays have demonstrated good validity and reproducibility as well as stability in protein expression with varying time to processing [36,37]. SomaLogic further conducts standardized data quality control at the sample level and protein level to adjust for variability between and within samples, and provides population-based normalized outputs of relative protein expression levels. Detailed information on SomaLogic’s quality control technique can be found on the manufacturer’s website (https://somalogic.com/technology/, accessed on 17 July 2022). SomaLogic provided two output files, each with different levels of population-based normalization, whereby the present study utilized the most-normalized output based on our sample distributions. Plasma samples from the STAGES cohort and intervention cohorts used the SomaScan platform of 5287 proteins (i.e., version 5K). After removal of non-human proteins (e.g., mouse), a total of 4985 proteins were quantified and analyzed (see Supplementary Table S16 for a complete list of proteins included in analyses). Serum samples from the Stanford Sleep Cohort used the SomaScan platform of 1300 proteins (i.e., version 1.3K).
4.5. GWAS Principal Components
As genetic ancestry plays an integral role in disease phenotyping, the first five principal components from genome-wide association study (GWAS) analysis were included as important covariates in the present study. A total of 467 patients with OSA and 907 controls, as defined above, all genotyped using an Affymetrix PMRA array (Thermo Fisher Scientific, Santa Clara, CA, USA), were included. The cohort was imputed to the 1000 Genome Phase III [47] after haplotype phasing using QCTOOL version 2. Quality control was conducted to ensure no duplicated patients based on inheritance by descent using KING version 2.2.6 [48]. Variants and samples with missing call rates lower than 0.1, and variants with minimum allele frequency lower than 0.05 were excluded. Imputation calls with R2 ≥ 0.9 were selected to compute principal component analysis using PLINK version 1.9 [49,50]. Patients were matched with the closest controls in a 1:2 ratio based on the Euclidean distance of individuals’ principal components.
4.6. Statistical Analyses
4.6.1. Descriptive Statistics
Data were analyzed in Jupyter Notebook version 6.0.1 with Python 3.7.4 using the statsmodels library version 0.10.1 [51]. Descriptive statistics are reported as mean and standard deviation (SD) for continuous variables, and number and percentage for dichotomous variables. We examined differences in demographic and clinical characteristics between participants with OSA and controls using Mann–Whitney U tests, as most of our continuous variables were not normally distributed, and Chi squared tests for dichotomous variables, whereby an a priori alpha level of 0.05 indicated a significant difference between groups.
4.6.2. Multivariate Proteomic Analyses
Using the statsmodels 0.10.1 library in Python 3.7.4 [51], we executed a series of multiple linear regression models with log-normalized protein expression as the dependent variable and the associated sleep apnea feature as the independent variable. Important covariates such as age, sex, BMI, log-normalized blood time to processing, blood draw period (i.e., morning or evening), study site, and the first five principal components from GWAS analysis to control for ancestry were included. As BMI is highly correlated with sleep apnea, we sought to include additional features as covariates to control for any residual effect, including BMI2 and age × BMI × sex. We executed regression models with the following sleep apnea features as independent variables: (1) log-normalized AHI + 1; (2) mild-to-moderate OSA vs. mild/no OSA; (3) severe OSA vs. controls; (4) moderate OSA vs. controls; and (5) mild vs. controls. We applied the Benjamini–Hochberg procedure for controlling the FDR to all p-values with an a priori p-value of 0.05 for identifying statistically significant proteins in all models.
4.6.3. Effect of Time to Blood Processing on Protein Expression
As our samples were collected at 11 different sites across the country, time to blood processing varied considerably. To account for this, we examined the effect of time to processing in STAGES samples (n = 1430; time to processing range: 1–126 h) in comparison to 224 samples processed immediately after collection. Multiple linear regressions with log-normalized protein expression as the dependent variable and log-normalized time to blood processing as the independent variable with a 5% FDR correction were used for this comparison. An FDR-corrected a priori p-value of 0.05 was used for significance. To facilitate interpretation of our findings, p-values and beta coefficients for the effect of time to processing are presented in the Supplementary Materials alongside multivariate results for each sleep apnea feature performed in the STAGES dataset.
4.6.4. Pathway Analysis
The ToppFun module of the ToppGene suite (Division of Biomedical Informatics, Cincinnati Children’s Hospital Medical Center (BMI CCHMC), Cincinnati, OH, USA) was utilized for biological pathway enrichment analysis for all 5% FDR significant protein sets [52]. The ToppGene Suite is a free web portal (http://toppgene.cchmc.org, accessed on 25 April 2022) that executes candidate gene/protein prioritization using functional annotations or network analysis. The ToppFun module detects functional enrichment of the input gene list based on gene ontology, protein domains and interactions, regulome, ontologies, phenotype, pharmacome, and bibliome, and applies FDR and Bonferroni correction for determining statistical significance. The goal of this network-based prioritization is to identify proteins/genes that are relevant to pathways involved in biological processes or diseases. The NCBI Entrez gene IDs of all 5% FDR significant proteins were entered into the ToppFun module, and results are presented based on significant pathways/functions identified by ToppFun for all FDR significant proteins as well as upregulated and downregulated significant proteins separately.
4.6.5. Validation of Protein Markers with a Machine Learning Classifier
The Python Scikit-learn LogisticRegressionCV library (version 1.0.2) [53] was used to train an ElasticNet (L1 and L2) penalty regularized lasso model to classify participants as OSA cases (AHI ≥ 15) or mild-to-no OSA (AHI < 15) based on relative protein expression. Hyperparameters of the models were tuned via 5-fold cross-validation using RepeatedKFold model evaluation. We used a 70% train–30% test split, whereby data on 974 (331 cases, 643 controls) participants were used for model training, and the remaining 417 (142 cases, 275 controls) participants were used for model testing. Three models were trained to predict OSA, each with the following features: (1) protein measures in addition to demographic information (age, sex, and BMI); (2) protein measures alone; and (3) demographic information (age, sex, and BMI) alone. Preprocessing for the classifier included log-normalization of relative protein expression and time to blood processing, followed by the conversion of all continuous variables to a standardized scale using the sklearn.preprocessing.StandardScaler feature. All models and classifiers used the same preprocessing, test–train split, and elastic net-based approach for comparison. We used two strategies to compare the ROC curves with statistical tests: DeLong test and a bootstrap approach. The DeLong test is a generalization of the Mann–Whitney statistic approach for the comparison of ROC curves [54]. The null hypothesis is that both areas under the curve are equal. The nonparametric asymptotic behavior for U-statistics is valid and can generalize in this case. The bootstrap approach consists of resampling bootstrap replicates of the data and comparing the distribution of the differences of AUCs to the normal distribution.
4.6.6. Effect of CPAP and Supplemental Oxygen Interventions on Sleep Apnea Proteins
Using the Python statsmodels library, Wilcoxon signed-rank sum tests, the nonparametric equivalent of a paired t-test, were conducted to examine if there were statistically significant differences in protein expression after CPAP/PAP compared to before CPAP/PAP in the combined WashU and Oxford cohort (n = 41). We further examined the effect of supplemental oxygen on proteins associated with AHI in comparison with air (i.e., sham) condition in the Oxford study (n = 25) by conducting Wilcoxon signed-rank sum tests between change scores in protein expression (i.e., post–pre values) after the oxygen condition and the air condition.
4.6.7. Replication with Our Prior Study in Serum
Lastly, we sought to replicate and validate our findings with multivariate and classifier analyses in a combined cohort of the samples from the present STAGES study (n = 1391) and samples from our previous study (i.e., Stanford Sleep Study [15]) including 530 participants. As our previous study utilized the SomaScan 1.3K platform, these analyses were conducted in a total of 1282 proteins who were present in both the 1.3K and 5K panels used in the STAGES study. We conducted multiple linear regression analysis in the combined cohort (n = 1921), including log-normalized protein expression as the dependent variable, log-normalized AHI + 1 as the independent variable, and covariates such as age, sex, BMI, BMI2, age × sex × BMI, blood draw period, GWAS principal components, and study (i.e., STAGES or Stanford Sleep Study).
5. Conclusions
The present study conducted the largest examination of proteins, to date, in adults with OSA. Confirming our results, PAI-I and tPA were robust predictors based on multivariate analyses and the machine learning classifier, and were replicated in the combined analysis. Both were also responsive to CPAP/PAP treatment. Reductions in PAI-1 after both supplemental oxygen and CPAP/PAP, but increases with the sham condition are consistent with previous studies [16,24], and reductions in sE-Selectin after CPAP/PAP is consistent with results of a recent meta-analysis [55]. Collectively, these findings support the use of PAI-I, tPA, and sE-Selectin as biomarkers of the presence and severity of sleep apnea, and they may provide a measure of treatment response in people with sleep apnea. These results may also inform considerations for additional preventive cardiovascular management for high-risk patients. Measurement of these key biomarkers may add to the patient profile and potentially inform additional therapeutic strategies even outside CPAP/PAP use, and creates an opportunity to inform cardiovascular disease management.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23147983/s1.
Author Contributions
Conceptualization, K.L.J.C. and E.M.; Data curation, A.M.M., A.B., P.K.S., S.K., R.B., C.A.K., Y.-E.S.J., N.P., C.D.T. and E.M.; Formal analysis, K.L.J.C., V.P.S., J.H. and E.M.; Funding acquisition, E.M.; Project administration, E.M.; Supervision, E.M.; Writing—original draft, K.L.J.C. and E.M.; Writing—review and editing, K.L.J.C., U.H., V.P.S., J.H., E.B.L., L.D.S., L.L., J.Z., A.M.M., A.B., P.K.S., S.K., R.B., C.A.K., Y.-E.S.J., N.P., C.D.T. and E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported, in part, by the National Heart, Lung, and Blood Institute (R24 HL114473, 75N92019R002, T32HL110952), the Bryte Foundation, and the Klarman Family Foundation. The SOX study was supported by the Oxford Radcliffe Hospital Charitable Funds, ResMed UK, and the National Institute for Health Research (NIHR) Oxford Biomedical Research Centre. The WashU study was supported by an investigator-initiated grant from Philips Respironics (no number), NIH awards (K23NS089922, UL1RR024992 subaward KL2TR000450), and a Washington University Institute of Clinical and Translational Sciences grant (UL1TR000448) from the NIH National Center for Advancing Translational Sciences. The content is solely the responsibility of the authors and does not necessarily represent the official view of the NIH. Dr. Nayia Petousi and Dr. Chris Turnbull were supported by NIHR Academic Clinical Lectureships. The views expressed are those of the authors and not necessarily of the NHS, the NIHR, or the Department of Health.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and was approved by the Institutional Review Board (or Ethics Committee) at all institutions, including Stanford University (protocol #39550), Bogan Sleep Consultants (protocol #420180158), Geisinger Medical Center (protocol #2018-0191), Mayo Clinic (protocol #17-007077), Toronto Sleep Institute (i.e., MedSleep, protocol #CR00110875), Washington University (protocol #201206112), and St. Luke’s Hospital (protocol #00002453).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data presented in this study from the STAGES cohort study, aside from the blood samples, are openly available as part of the National Sleep Research Resource (NSRR) at https://sleepdata.org/datasets/stages (accessed on 17 July 2022), and blood data may be made available upon request from the corresponding author.
Acknowledgments
We would like to thank John Stradling, Dushendree Sen, the staff at the Oxford Respiratory Trials Unit, and the study participants for their contributions to this manuscript.
Conflicts of Interest
Dr. Katie Cederberg, Mr. Umaer Hanif, Mr. Vicente Peris Sempere, Mr. Julien Hédou, Dr. Jing Zhang, Dr. Ling Lin, Dr. Suresh Kotagal, Dr. Adam Blackman, Dr. Clete Kushida, and Dr. Nayia Petousi declare no conflict of interest. Dr. Chris Turnbull declares consulting fees from Bayer and honoraria from Stowood, outside the scope of this work. Dr. Schneider reports personal fees from Jazz Pharmaceuticals, personal fees from Harmony Biosciences, personal fees from Eisai, outside the submitted work. Dr. Leary is an employee of Jazz Pharmaceuticals who, in the course of her employment, has received stock options exercisable for, and other stock awards of, ordinary shares of Jazz Pharmaceuticals plc. Dr. Anne Marie Morse has received consulting fees from Jazz Pharmaceuticals, Harmony Biosciences, and Avadel, outside the scope of this work. Dr. Paula Schweitzer has received consulting fees from Apnimed and Jazz Pharmaceuticals; her institution has received research funding from Apnimed, Avadel, Harmony Biosciences, Inspire Medical, and Suven Life Sciences, all outside the scope of this work. Dr. Richard Bogan is a shareholder in WaterMark Medical and Healthy Humming, LLC; serves on the Board of Directors for WaterMark Medical; is a consultant to Jazz, Harmony Biosciences, Takeda, Avadel, and Oventus; has industry funded research for Avadel, BresoTec, Idorsia, Suven, Jazz, Balance, Vanda, Merck, Eisai, Philips, Fresca, Takeda, Liva Nova, Roche, Sommetrics, NLS, Sanofi, Apnimed; and is on the Speakers Bureau for Jazz, Eisai, Harmony, Idorsia; all outside the scope of this work. Dr. Yo-El Ju reports consulting fees from Applied Cognition, outside the scope of this work. Dr. Emmanuel Mignot occasionally consults and has received contracts from Jazz Pharmaceuticals, Orexia/Centessa, Tekeda, Dreem, and ActiGraph; has received grant/clinical trial funding from Haromony, Tekeda, Apple, Humani, Sunovion, Indorsia, Eisai; is and has been a Principal Investigator on clinical trials using sodium oxybate and Solriamfetol, Jazz Pharmaceutical products, for the treatment of Type 1 narcolepsy; all outside the scope of this work.
References
- American Academy of Sleep Medicine. International Classification of Sleep Disorders, 3rd ed.; American Academy of Sleep Medicine: Darien, IL, USA, 2015. [Google Scholar]
- Rundo, J.V.; Downey, R., 3rd. Polysomnography. Handb. Clin. Neurol. 2019, 160, 381–392. [Google Scholar] [PubMed]
- Berry, R.B.; Budhiraja, R.; Gottlieb, D.J.; Gozal, D.; Iber, C.; Kapur, V.K.; Marcus, C.L.; Mehra, R.; Parthasarathy, S.; Quan, S.F.; et al. Rules for scoring respiratory events in sleep: Update of the 2007 AASM Manual for the Scoring of Sleep and Associated Events. Deliberations of the Sleep Apnea Definitions Task Force of the American Academy of Sleep Medicine. J. Clin. Sleep Med. 2012, 8, 597–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjafield, A.V.; Ayas, N.T.; Eastwood, P.R.; Heinzer, R.; Ip, M.S.M.; Morrell, M.J.; Nunez, C.M.; Patel, S.R.; Penzel, T.; Pépin, J.L.; et al. Estimation of the global prevalence and burden of obstructive sleep apnoea: A literature-based analysis. Lancet Respir. Med. 2019, 7, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Senaratna, C.V.; Perret, J.L.; Lodge, C.J.; Lowe, A.J.; Campbell, B.E.; Matheson, M.C.; Hamilton, G.S.; Dharmage, S.C. Prevalence of obstructive sleep apnea in the general population: A systematic review. Sleep Med. Rev. 2017, 34, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Heinzer, R.; Vat, S.; Marques-Vidal, P.; Marti-Soler, H.; Andries, D.; Tobback, N.; Mooser, V.; Preisig, M.; Malhotra, A.; Waeber, G.; et al. Prevalence of sleep-disordered breathing in the general population: The HypnoLaus study. Lancet Respir. Med. 2015, 3, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Peppard, P.E.; Young, T.; Palta, M.; Skatrud, J. Prospective study of the association between sleep-disordered breathing and hypertension. N. Engl. J. Med. 2000, 342, 1378–1384. [Google Scholar] [CrossRef]
- Lisan, Q.; van Sloten, T.; Boutouyrie, P.; Laurent, S.; Danchin, N.; Thomas, F.; Guibout, C.; Perier, M.C.; Dagnelie, P.; Henry, R.M.; et al. Sleep Apnea is Associated With Accelerated Vascular Aging: Results From 2 European Community-Based Cohort Studies. J. Am. Heart Assoc. 2021, 10, e021318. [Google Scholar] [CrossRef]
- Azarbarzin, A.; Sands, S.A.; Taranto-Montemurro, L.; Vena, D.; Sofer, T.; Kim, S.W.; Stone, K.L.; White, D.P.; Wellman, A.; Redline, S. The Sleep Apnea-Specific Hypoxic Burden Predicts Incident Heart Failure. Chest 2020, 158, 739–750. [Google Scholar] [CrossRef]
- Javaheri, S.; Barbe, F.; Campos-Rodriguez, F.; Dempsey, J.A.; Khayat, R.; Javaheri, S.; Malhotra, A.; Martinez-Garcia, M.A.; Mehra, R.; Pack, A.I.; et al. Sleep Apnea: Types, Mechanisms, and Clinical Cardiovascular Consequences. J. Am. Coll. Cardiol. 2017, 69, 841–858. [Google Scholar] [CrossRef]
- Baillieul, S.; Dekkers, M.; Brill, A.K.; Schmidt, M.H.; Detante, O.; Pépin, J.L.; Tamisier, R.; Bassetti, C.L.A. Sleep apnoea and ischaemic stroke: Current knowledge and future directions. Lancet Neurol. 2022, 21, 78–88. [Google Scholar] [CrossRef]
- Baumert, M.; Immanuel, S.A.; Stone, K.L.; Litwack Harrison, S.; Redline, S.; Mariani, S.; Sanders, P.; McEvoy, R.D.; Linz, D. Composition of nocturnal hypoxaemic burden and its prognostic value for cardiovascular mortality in older community-dwelling men. Eur. Heart J. 2020, 41, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Bock, J.; Covassin, N.; Somers, V. Excessive daytime sleepiness: An emerging marker of cardiovascular risk. Heart 2022. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J. Biomarkers, old and new: For sleep, sleepiness, circadian phase and much more. J. Sleep Res. 2016, 25, 255–256. [Google Scholar] [CrossRef] [Green Version]
- Ambati, A.; Ju, Y.E.; Lin, L.; Olesen, A.N.; Koch, H.; Hedou, J.J.; Leary, E.B.; Sempere, V.P.; Mignot, E.; Taheri, S. Proteomic biomarkers of sleep apnea. Sleep 2020, 43, zsaa086. [Google Scholar] [CrossRef]
- Steffanina, A.; Proietti, L.; Antonaglia, C.; Palange, P.; Angelici, E.; Canipari, R. The Plasminogen System and Transforming Growth Factor-β in Subjects with Obstructive Sleep Apnea Syndrome: Effects of CPAP Treatment. Respir. Care 2015, 60, 1643–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagai, K.; Muldowney, J.A., 3rd; Song, Y.; Wang, L.; Bagai, J.; Artibee, K.J.; Vaughan, D.E.; Malow, B.A. Circadian variability of fibrinolytic markers and endothelial function in patients with obstructive sleep apnea. Sleep 2014, 37, 359–367. [Google Scholar] [CrossRef]
- Fernández-Bello, I.; Monzón Manzano, E.; García Río, F.; Justo Sanz, R.; Cubillos-Zapata, C.; Casitas, R.; Sánchez, B.; Jaureguizar, A.; Acuña, P.; Alonso-Fernández, A.; et al. Procoagulant State of Sleep Apnea Depends on Systemic Inflammation and Endothelial Damage. Arch. Bronconeumol. 2022, 58, 117–124. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Y.N.; Zhang, L.Y.; Li, C.X.; Li, S.Q.; Li, H.P.; Zhang, L.; Li, N.; Yan, Y.R.; Li, Q.Y. Changes of circulating biomarkers of inflammation and glycolipid metabolism by CPAP in OSA patients: A meta-analysis of time-dependent profiles. Ther. Adv. Chronic Dis. 2022, 13, 20406223211070919. [Google Scholar] [CrossRef]
- Yokoe, T.; Minoguchi, K.; Matsuo, H.; Oda, N.; Minoguchi, H.; Yoshino, G.; Hirano, T.; Adachi, M. Elevated levels of C-reactive protein and interleukin-6 in patients with obstructive sleep apnea syndrome are decreased by nasal continuous positive airway pressure. Circulation 2003, 107, 1129–1134. [Google Scholar] [CrossRef] [Green Version]
- Daniel, S.; Cohen-Freud, Y.; Shelef, I.; Tarasiuk, A. Bone mineral density alteration in obstructive sleep apnea by derived computed tomography screening. Sci. Rep. 2022, 12, 6462. [Google Scholar] [CrossRef]
- Yen, C.M.; Kuo, C.L.; Lin, M.C.; Lee, C.F.; Lin, K.Y.; Lin, C.L.; Chang, S.N.; Sung, F.C.; Kao, C.H. Sleep disorders increase the risk of osteoporosis: A nationwide population-based cohort study. Sleep Med. 2014, 15, 1339–1344. [Google Scholar] [CrossRef]
- Sies, N.S.; Zaini, A.A.; de Bruyne, J.A.; Jalaludin, M.Y.; Nathan, A.M.; Han, N.Y.; Thavagnanam, S. Obstructive sleep apnoea syndrome (OSAS) as a risk factor for secondary osteoporosis in children. Sci. Rep. 2021, 11, 3193. [Google Scholar] [CrossRef]
- von Känel, R.; Loredo, J.S.; Ancoli-Israel, S.; Dimsdale, J.E. Association between sleep apnea severity and blood coagulability: Treatment effects of nasal continuous positive airway pressure. Sleep Breath 2006, 10, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Kruithof, E.K.; Nicolosa, G.; Bachmann, F. Plasminogen activator inhibitor 1: Development of a radioimmunoassay and observations on its plasma concentration during venous occlusion and after platelet aggregation. Blood 1987, 70, 1645–1653. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wan, Y.; Xu, H.; Wu, Y.; Hu, B.; Jin, H. The role of endogenous tissue-type plasminogen activator in neuronal survival after ischemic stroke: Friend or foe? Cell. Mol. Life Sci. 2019, 76, 1489–1506. [Google Scholar] [CrossRef] [PubMed]
- Moore, H.B.; Moore, E.E. Temporal Changes in Fibrinolysis following Injury. Semin. Thromb. Hemost. 2020, 46, 189–198. [Google Scholar] [CrossRef]
- Miltiades, A.; Houck, P.J.; Monteleone, M.; Harrison, N.L.; Cabrera-Garcia, D.; Roh, D.; Wagener, G. Insights into Fibrinogen-Mediated COVID-19 Hypercoagubility in Critically Ill Patients. J. Neurosurg. Anesthesiol. 2022, 34, 136–140. [Google Scholar] [CrossRef]
- Zuo, Y.; Warnock, M.; Harbaugh, A.; Yalavarthi, S.; Gockman, K.; Zuo, M.; Madison, J.A.; Knight, J.S.; Kanthi, Y.; Lawrence, D.A. Plasma tissue plasminogen activator and plasminogen activator inhibitor-1 in hospitalized COVID-19 patients. Sci. Rep. 2021, 11, 1580. [Google Scholar] [CrossRef] [PubMed]
- Wygrecka, M.; Birnhuber, A.; Seeliger, B.; Michalick, L.; Pak, O.; Schultz, A.S.; Schramm, F.; Zacharias, M.; Gorkiewicz, G.; David, S.; et al. Altered fibrin clot structure and dysregulated fibrinolysis contribute to thrombosis risk in severe COVID-19. Blood Adv. 2022, 6, 1074–1087. [Google Scholar] [CrossRef]
- Johansson, L.; Jansson, J.H.; Boman, K.; Nilsson, T.K.; Stegmayr, B.; Hallmans, G. Tissue plasminogen activator, plasminogen activator inhibitor-1, and tissue plasminogen activator/plasminogen activator inhibitor-1 complex as risk factors for the development of a first stroke. Stroke 2000, 31, 26–32. [Google Scholar] [CrossRef]
- Lindgren, A.; Lindoff, C.; Norrving, B.; Astedt, B.; Johansson, B.B. Tissue plasminogen activator and plasminogen activator inhibitor-1 in stroke patients. Stroke 1996, 27, 1066–1071. [Google Scholar] [CrossRef]
- Margaglione, M.; Di Minno, G.; Grandone, E.; Vecchione, G.; Celentano, E.; Cappucci, G.; Grilli, M.; Simone, P.; Panico, S.; Mancini, M. Abnormally high circulation levels of tissue plasminogen activator and plasminogen activator inhibitor-1 in patients with a history of ischemic stroke. Arter. Thromb. 1994, 14, 1741–1745. [Google Scholar] [CrossRef] [Green Version]
- Azarbarzin, A.; Zinchuk, A.; Wellman, A.; Labarca, G.; Vena, D.; Gell, L.; Messineo, L.; White, D.P.; Gottlieb, D.J.; Redline, S.; et al. Cardiovascular Benefit of CPAP in Adults with Coronary Artery Disease and OSA without Excessive Sleepiness. Am. J. Respir. Crit. Care Med. 2022. [Google Scholar] [CrossRef]
- Lao, M.; Cheng, Y.; Gao, X.; Ou, Q. The interaction among OSA, CPAP, and medications in patients with comorbid OSA and cardiovascular/cerebrovascular disease: A randomized controlled trial. BMC Pulm. Med. 2022, 22, 99. [Google Scholar] [CrossRef] [PubMed]
- Candia, J.; Cheung, F.; Kotliarov, Y.; Fantoni, G.; Sellers, B.; Griesman, T.; Huang, J.; Stuccio, S.; Zingone, A.; Ryan, B.M.; et al. Assessment of Variability in the SOMAscan Assay. Sci. Rep. 2017, 7, 14248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.H.; Tworoger, S.S.; Stampfer, M.J.; Dillon, S.T.; Gu, X.; Sawyer, S.J.; Chan, A.T.; Libermann, T.A.; Eliassen, A.H. Stability and reproducibility of proteomic profiles measured with an aptamer-based platform. Sci. Rep. 2018, 8, 8382. [Google Scholar] [CrossRef] [PubMed]
- Hanif, U.R.; Leary, E.B.; Schneider, L.D.; Paulsen, R.R.; Morse, A.M.; Blackman, A.; Schweitzer, P.K.; Kushida, C.A.; Liu, S.Y.; Jennum, P.; et al. Estimation of Apnea-Hypopnea Index using Deep Learning on 3D Craniofacial Scans. IEEE J. Biomed. Health Inform. 2021, 25, 4185–4194. [Google Scholar] [CrossRef]
- Leary, E.; Malunjkar, S.; Qadri, S.; Ruoff, C.; Sullivan, S.; Walsh, J.; Mignot, E. Deployment of the Alliance Sleep Questionnaire (ASQ) at the Stanford Sleep Disorders Clinic (SSDC) as Standard of Care. Sleep 2013, 36 (Suppl. S1), A413. [Google Scholar]
- Leary, E.; Malunjkar, S.; Ruoff, C.; Walsh, J.; Mignot, E. Validation of the Alliance Sleep Questionnaire (ASQ) Restless Legs Syndrome Module Module in Sleep Disordered Patients. Sleep 2015, 38, A432. [Google Scholar]
- Ju, Y.S.; Zangrilli, M.A.; Finn, M.B.; Fagan, A.M.; Holtzman, D.M. Obstructive sleep apnea treatment, slow wave activity, and amyloid-β. Ann. Neurol. 2019, 85, 291–295. [Google Scholar] [CrossRef]
- Turnbull, C.D.; Sen, D.; Kohler, M.; Petousi, N.; Stradling, J.R. Effect of Supplemental Oxygen on Blood Pressure in Obstructive Sleep Apnea (SOX). A Randomized Continuous Positive Airway Pressure Withdrawal Trial. Am. J. Respir. Crit. Care Med. 2019, 199, 211–219. [Google Scholar] [CrossRef]
- Berry, R.B.; Brooks, R.; Gamaldo, C.; Harding, S.M.; Lloyd, R.M.; Quan, S.F.; Troester, M.T.; Vaughn, B.V. AASM Scoring Manual Updates for 2017 (Version 2.4). J. Clin. Sleep Med. 2017, 13, 665–666. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, S.; Vaught, J.D.; Bock, C.; Gold, L.; Katilius, E.; Keeney, T.R.; Kim, N.; Saccomano, N.A.; Wilcox, S.K.; Zichi, D.; et al. From SOMAmer-based biomarker discovery to diagnostic and clinical applications: A SOMAmer-based, streamlined multiplex proteomic assay. PLoS ONE 2011, 6, e26332. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Ngo, D.; Finkel, N.; Pitts, R.; Gleim, S.; Benson, M.D.; Keyes, M.J.; Farrell, L.A.; Morgan, T.; Jennings, L.L.; et al. Application of Large-Scale Aptamer-Based Proteomic Profiling to Planned Myocardial Infarctions. Circulation 2018, 137, 1270–1277. [Google Scholar] [CrossRef]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]
- Manichaikul, A.; Mychaleckyj, J.C.; Rich, S.S.; Daly, K.; Sale, M.; Chen, W.M. Robust relationship inference in genome-wide association studies. Bioinformatics 2010, 26, 2867–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and statistical modeling with python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June 2010; p. 61. [Google Scholar]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Xiao, J.; Kang, J.; Sun, H.; Mu, Z.; Tong, D.; Li, M. Effects of Continuous Positive Airway Pressure on Cell Adhesion Molecules in Patients with Obstructive Sleep Apnea: A Meta-Analysis. Lung 2021, 199, 639–651. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).